
Abstract
Aim
This study investigated the efficacy of a new strength training method on strength gain, hypertrophy, and neuromuscular fatigability.
Methods
The training exercise consisted of elbow flexion against a load of ~ 70% of one repetition maximal (1RM). A new method (3/7 method) consisting of five sets of an increasing number of repetitions (3 to 7) during successive sets and brief inter-set intervals (15 s) was repeated two times after 150 s of recovery and compared to a method consisting of eight sets of six repetitions with an inter-set interval of 150 s (8 × 6 method). Subjects trained two times per week during 12 weeks. Strength gain [1RM load and maximal isometric voluntary contraction (MVC)], EMG activity of biceps brachii and brachioradialis, as well as biceps’ brachii thickness were measured. Change in neuromuscular fatigability was assessed as the maximal number of repetitions performed at 70% of 1RM before and after training.
Results
Both 3/7 and 8 × 6 methods increased 1RM load (22.2 ± 7.4 and 12.1 ± 6.6%, respectively; p < 0.05) and MVC force (15.7 ± 8.2 and 9.5 ± 9.5%; p < 0.05) with a greater 1RM gain (p < 0.05) for the 3/7 method. Normalized (%Mmax) EMG activity of elbow flexors increased (p < 0.05) similarly (14.5 ± 23.2 vs. 8.1 ± 20.5%; p > 0.05) after both methods but biceps’ brachii thickness increased to a greater extent (9.6 ± 3.6 vs. 5.5 ± 3.7%; p < 0.05) for the 3/7 method. Despite subjects performing more repetitions with the same absolute load after training, neuromuscular fatigability increased (p < 0.05) after the two training methods.
Conclusion
The 3/7 method provides a better stimulus for strength gain and muscle hypertrophy than the 8 × 6 method.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Even though it is well documented that the gain in muscle strength in response to a long-term strength training program mainly reflects an increase in muscle mass (Häkkinen et al. 1998; Aagaard et al. 2001; Duchateau and Baudry 2011), the stimulus triggering muscle hypertrophy remains a matter of debate (Spiering et al. 2008; Schoenfeld 2013; Dankel et al. 2017). High mechanical loading of the muscle [> 60–70% of the one repetition maximal (1RM)] has long been considered as the main stimulus for hypertrophy through mechanotransduction process (Vierck et al. 2000; Kraemer and Ratamess 2004; Ogborn and Schoenfeld 2014). However, some studies have also suggested that the use of load greater than 60% of 1RM and brief rest interval between sets might provide a superior stimulus for muscle hypertrophy than longer rest periods (Kraemer et al. 1990; Bottaro et al. 2009; de Salles et al. 2009). In addition to the production of metabolites (lactate, H+, Pi, etc.) induced by high-load training, the brief recovery periods between sets further favour their intracellular accumulation. Such metabolic stimulus has been proposed to play a role in the increase in muscle mass and strength, either directly by its action as an anabolic signal (Goto et al. 2005; Ozaki et al. 2016; Nalbandian and Takeda 2016), or indirectly through the intensification of muscle activation (Dankel et al. 2017).
In this context, a recent work underscored the efficacy of a new training design (3/7 method) to promote strength gains (Laurent et al. 2016). The 3/7 method is characterized by an incremental number of repetitions (from 3 to 7 repetitions) during 5 successive sets, a load of ~ 70% of 1RM, and very brief rest interval between sets (15 s). When compared to a more “classical” method using constant repetitions per set and, similar load and training volume (4 × 6 method: 4 sets of 6 repetitions with 150-s rest interval between sets), the 3/7 method showed a greater strength gain in bench press exercise after 12 weeks of training (Laurent et al. 2016). In addition to the very brief inter-set interval, the incremental number of repetitions in the successive sets should favour metabolites’ accumulation within the muscle at the completion of the exercise that may have induced a greater stimulus for muscle hypertrophy than the 4 × 6 method. In agreement with this suggestion, a greater deficit in tissue oxygenation, recorded by near-infrared spectroscopy (NIRS), was observed in the biceps brachii (BB) and brachioradialis (BR) when lifting and lowering a load (70% of 1RM) with the elbow-flexor muscles for the 3/7 method compared with a more classical strength training method. This observation suggests that the brief rest duration between successive sets and perhaps the incremental number of repetitions during the successive series have substantial influence that likely explains the greater efficacy of the 3/7 method (Laurent et al. 2016).
No attempt was made to analyse the neuromuscular adaptations associated with strength gain in the study by Laurent and colleagues (2016). Therefore, to improve our understanding of the mechanisms underlying the greater efficacy of the 3/7 method compared with the 4 × 6 method, the present study compares the training-related gains in maximal strength, muscle mass and electromyographic (EMG) activity in a single muscle group, the elbow flexors, after a 12-weeks training program with a load of ~ 70% of 1RM. To reach a substantial amount of repetitions (~ 50) per session, the 3/7 method was repeated two times with a 150 s rest in between and compared to a method consisting of 8 sets of 6 repetitions (8 × 6 method) with 150 s of rest between sets. As the 3/7 method promotes greater metabolic changes (deoxygenation) than the 8 × 6 method (Penzer et al. 2016), we also assessed changes in neuromuscular fatigability after the two training programs. We hypothesized that the 3/7 method would induce greater strength gain and muscle hypertrophy, and improve muscle endurance to a greater extent than the 8 × 6 method.
Methods
Subjects
A total of 43 healthy subjects aged between 18 and 32 years participated in this study. Subjects were not engaged in any strength training program for the last 6 months prior to their participation to the study and were instructed to maintain their habitual activity for the entire duration of the study. Individuals with orthopaedic problems of the upper limb were excluded from this study. During the first experimental sessions, subjects’ assignment to the training groups (3/7 method, n = 16 or 8 × 6 method, n = 15) was performed so that age, initial strength, and number of women and men in each group was almost similar (Table 1). All these subjects completed the 12-week training program. In addition, a group of 12 subjects (5 women and 7 men), who did not train, was used to test data reliability of the main parameters at several weeks’ interval. The evaluation of the intraclass correlation coefficient (ICC) and the coefficient of variation (CV) values revealed an excellent day-to-day reliability agreement (mean values > 0.94) for the main dependent variables assessed in these subjects (Table 2) whereas ICC values for the fatigue-related parameters (i.e., the number of repetitions) indicated a moderate reliability (mean value 0.80).
Approval for the project was obtained from the local Ethics Committee, and each subject signed an informed consent document prior to their enrolment in the study. All procedures used in this study conformed to the Declaration of Helsinki.
Experimental apparatus
Subjects were seated in an adjustable chair with the right arm resting on a padded support in a horizontal position with the forearm supinated. A custom-made orthosis was placed around the wrist that was either connected to a force transducer or to an inertial load depending of the task to be performed (Baudry et al. 2013). The force transducer (linear range 0–1 KN, Maywood Instruments Ltd, Basingstoke, UK) was used to assess the isometric force exerted by the elbow flexors during maximal voluntary contraction (MVC) performed at an elbow angle of 90°. The inertial load used for the exercise was connected to the orthosis via a movable cable and pulley system at the same location as the force transducer. An inclinometer (precision 0.1°; ASM GmbH, München, Germany) fixed to the lateral side of the forearm was used to continuously monitor the angular position of the forearm relative to the arm (placed horizontally) in the elbow flexion–extension direction. The signals from the force transducer (MVCs) and inclinometer (dynamic contractions) were A/D sampled at 200 Hz (Power 1401, 16-bit resolution, Cambridge Electronic Design, Cambridge, UK).
Electromyogram
The EMG was recorded by means of self-adhesive Ag-AgCl electrodes of 8-mm in diameter and placed in a bipolar configuration with an inter-electrode distance of 2 cm over the belly of the BB (long head) and BR. To reduce the impedance at the skin–electrode interface, the skin was shaved when necessary and cleaned with a solution of alcohol, ether, and acetone. The reference electrodes were fastened over the ulna bone at the elbow level. The EMG signals were amplified (1000×) and bandpass filtered (10–1000 Hz) prior to being A/D sampled at 2 kHz (Power 1401, 16-bit resolution, Cambridge Electronic Design, UK). To position EMG electrodes at the same location in the different sessions, anatomical landmarks were carefully determined during the first experimental session. The location of each electrode was determined with the elbow flexed at 90° and the forearm in supination. For the BR, the longitudinal distance of the electrodes location from the radial styloid process in direction of the lateral epicondyle and the lateral distance of its perpendicular projection to this line were measured. For the BB, the longitudinal distance of the electrodes location from the acromion and the lateral distance of its perpendicular projection to a line going from the lateral epicondyle to the acromion were determined.
NIRS
A tissue oximeter (NIRO-200, Hamamatsu Photonics, Japan) that used continuous wave light and a multi-distance approach was used to measure the average tissue O2 saturation based on spatially resolved spectroscopy (Ferrari et al. 2004). NIRS measurements (sampling rate 5 Hz) were performed from the BB with the emission and detection probes positioned 3 cm apart along the longitudinal axis of the arm at about mid-distance between the acromion and the lateral epicondyle, close to the EMG electrodes. The probes were maintained by a rigid rubber shell, and firmly attached with double-sided adhesive tape. The same anatomical landmarks than for BB EMG electrodes were used to place NIRS probes at the same location in the different sessions.
Ultrasonography
Muscle thickness of the BB long head was chosen as being representative of this muscle group and assessed from images obtained with a realtime B-mode echograph (DP-6900Vet, Shenzhen Mindray Bio-Medical Electronics CO, Ltd., China) with a 6-cm width linear-array probe (7.5 MHz; 75L60EA). During these recordings, subjects were standing upright with the right arm slightly abducted (± 25°) and the forearm placed in supination. This position was kept in place by a second experimenter allowing the subject’s arm to be completely relaxed. The location and length of the belly of the BB long head were first determined by means of palpation and ultrasound (transversal and longitudinal) images, along a line between the acromion and its distal insertion. Thereafter, the probe was placed longitudinally on the muscle belly along the longitudinal axis of the muscle and ultrasound images were recorded at three adjacent locations (mid-belly and, proximal and distal to this position). These anatomical landmarks were noted to replace each probe at the same location before and after the training period. The probe was coated with a water-soluble transmission gel to provide acoustic contact. Particular care was taken to minimize the pressure of the probe to the skin during image acquisition.
Electrical nerve stimulation
Electrical stimuli (rectangular pulses, 0.2-ms duration) were delivered to the brachial plexus at Erb’s point via a constant current stimulator (DS7AH, Digitimer, Hertfordshire, UK) to evoke a M wave in BB and BR. The cathode and the anode (silver disks, 8 mm in diameter) were located at the supraclavicular fossa and over the superior portion of the trapezius muscle, respectively. To ensure maximal muscle activation throughout the experimental session, stimulus intensity was set at 120% of the smallest intensity eliciting a maximal M-wave (Mmax) in both BB and BR. The Mmax was used to normalize the EMG signals recorded during voluntary contractions. Although this procedure has some limitations (see Keenan et al. 2005), it takes into account differences in recording conditions between subjects and from session to session for a given subject.
Fatiguing test
The test consisted of performing elbow flexions as many times as possible against an inertial load equivalent to 70% of 1RM at a fixed cadence of 25 repetitions per min. One repetition consisted of lifting and lowering the load over the entire range of motion (30°; from 105° to 75° of elbow flexion). This range of motion was chosen because we observed in previous study (Baudry et al. 2013) that for greater angular displacement, the NIRS signal was altered due to movement artifacts. The elbow angle signal was displayed online on a monitor in front of the subject with two horizontal cursors indicating the lower and upper limits of the range of motion. To assess training-related changes in neuromuscular endurance, the fatiguing test was performed at the same relative load (70% of 1RM of the session). Furthermore, to assess the local endurance performance (number of repetitions at the same absolute load), a second fatiguing test was performed during the post-training session using the same absolute load that was used in the pre-training session. A rest interval of 20 min was given between the two fatiguing tests, the relative-load fatiguing test being always performed before the absolute-load fatiguing test (Baudry et al. 2013).
Experimental procedures
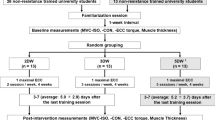
Each subject participated in one familiarization session and two experimental sessions (one session before and one session after the training program). The familiarization session was scheduled 3–4 days before the pre-training session and the post-training session was performed 3–4 days after the last training session to avoid residual fatigue. Each experimental session began with the ultrasonography recordings. After the positioning of the EMG electrodes and the NIRS probes, subjects performed brief (~ 3 s) isometric MVCs with the elbow flexors. During the MVCs, subjects were verbally encouraged to produce their maximal force. At least two trials were performed, with subjects resting for 120 s between trials. When peak force of these two MVCs was within 5% of each other, the greatest value was taken as the maximum. Otherwise, additional trials were performed until the 5% criterion was achieved. All subjects met this criterion within a maximal of five trials. Then, the location and stimulation intensity for Mmax were determined. The Mmax was recorded during a low-intensity contraction of 20% MVC.
Thereafter, the 1RM load was assessed (0.25-kg accuracy) by increasing the inertial load until the subject was unable to lift it over the entire range of motion (30°; from 105° to 75° of elbow flexion). Between 4 and 8 trials were needed to determine the 1RM load. A minimum of 90 s was provided between trials. Then, the fatiguing tests with the relative (before and after training) and absolute load (only after training) were performed.
Training
Subjects trained the right elbow flexors (two subjects in each group were left-handed) two times per week during 12-weeks, for a total of 24 sessions. Two successive training sessions were separated by a minimum interval of 48 h, each session being supervised by one investigator or specialized person. Every training session began by a warm-up consisting of two sets of 10 repetitions with a load corresponding to 50% of the training load. The training exercise was performed on a dual adjustable pulley equipment (Life fitness®, Brunswick Corporation, Lake Forest, IL). The weight was suspended to a cable passing over the two pulleys and connected to a handle at the other extremity. Subjects were comfortably seated with the right arm resting on a padded support in a horizontal position and the forearm supinated. The training exercise required lifting and lowering the load (70% of 1RM) over the same range of motion and cadence as for the testing sessions. The 3/7 method sessions were composed of two bouts (150 s of rest between bouts) of five sets performed with an increment of one repetition per set, starting with three repetitions for the first set. A rest interval of 15 s was given between successive sets. During the 8 × 6 method sessions, subjects performed 8 sets of 6 repetitions with a resting interval of 150 s between sets and a similar movement cadence as the 3/7 method. For both methods, when subjects were unable to perform the prescribed numbers of movements, the supervisor provided the least amount of assistance necessary to enable the subject to complete the required number of repetitions. The initial load used at the beginning of the training period corresponded to the maximal load with which a subject was able to complete between 10 and 12 repetitions. This load was determined during the first session of training by gradually increasing the load. The training load was readjusted after the first five training sessions and thereafter during the training period when subjects were able to complete the training volume with the prescribed load during two consecutive sessions without any assistance. The load was increased by a minimum increment of 0.5 kg, corresponding to 2.5–5% of the previous training load. The subjects completed the 12-weeks training program with a compliance to training sessions of 100%.
Data analysis
Muscle thickness of BB, defined as the distance from the superficial aponeurosis to the deep aponeurosis (Fig. 1), was measured at the proximal, mid, and distal portion of the muscle. For the three probe locations, the distance between deep and superficial aponeuroses was measured at the middle and the two extremities of each image. Muscle thickness value used for analysis corresponded to the average of these nine locations. Data from two subjects (1 subject in each training group) were not used in the analysis due to a technical problem.

Sagittal reconstruction of the BB long head from ultrasound images in one subject before and after 12 weeks of strength training with the 3/7 method. The thickness of the BB was defined as the distance between the superficial and deep aponeuroses (vertical lines) for nine locations along the muscle
The peak force produced during each MVC was recorded and the greatest value was taken for subsequent analysis. The averaged value of the rectified EMG (aEMG) of the BB and BR was measured over a 1-s epoch around the peak force during each MVC. The aEMG of BB and BR was analyzed separately and then averaged to represent the aEMG of the elbow-flexor muscles. The peak-to-peak amplitude of the Mmax was measured from the unrectified EMG signal. For the BB and BR, the aEMG measured during the MVC was normalized to the corresponding Mmax.
During the fatiguing test, the aEMG of these muscles was measured over the entire range of movement, with the onset and end of the elbow flexion–extension movements determined from the inclinometer signal. The aEMG (expressed relative to aEMG recorded during MVC) of the first and last repetitions was used for statistical analysis.
The NIRS measurement provides a tissue oxygenation index (TOI, expressed in %) that is assumed to represent a dynamic balance between oxygen supply and utilization (Ferrari et al. 2004). While previous studies suggested that TOI reflected the mean arteriolar, capillary, and venous hemoglobin O2 saturation levels with a minor contribution from myoglobin (< 20%; Ferrari et al. 2004), more recent studies reported a predominant myoglobin contribution (> 50–70%) to the NIRS signal (Davis and Barstow 2013; Bendahan et al. 2017). TOI was measured at the end of each concentric phase (75° of elbow flexion) of the repetitions during the fatiguing test. The values of the first and last repetitions were used for statistical comparison.
Statistics
Prior to the comparison of each dependent variable, the Gaussian distribution of the data was verified by the Kolmogorov–Smirnov test. Unpaired Student t tests were used to compare 1RM load, MVC force and muscle thickness between the two methods before training. Two-way ANOVAs (methods × training) were used to compare the effects of the two training methods on the 1RM load, training load, MVC force, muscle thickness, aEMG during MVC and Mmax, and the variables extracted from the fatiguing tests [number of repetitions, MVC (% pre-fatigue), aEMG during MVC (% pre-fatigue)]. In addition, the changes (%) for each variable were compared between methods by unpaired Student’s t tests. Three-way ANOVAs (method × training × repetition) with repeated measures design for training and repetition were used to analyse the influence of the training methods on aEMG, TOI measured during the relative-load fatiguing test. Tukey post-hoc test was used when significant interactions were found to determine specific differences between mean values. Coefficients of determination extracted from Pearson product–moment correlations were calculated for the association between training-related changes in 1RM load or MVC force with muscle thickness, 1RM load with MVC force, 1RM load with number of repetitions during the absolute-load fatiguing test. The level of statistical significance was set at p ≤ 0.05 for all comparisons. Values are expressed as mean ± SD in the text.
Results
Training load
The training load did not differ significantly between the two groups neither at the beginning (3/7 method 11.4 ± 3.1 kg; 8 × 6 method 10.8 ± 2.3 kg; p = 0.98), nor at the end of the training program (3/7 method 14.0 ± 3.4 kg; 8 × 6 method 14.3 ± 3.1 kg; p = 0.99). However, a greater increase (p < 0.01) in the training load was observed for the 8 × 6 method (32.4 ± 7.3%) compared with the 3/7 method (23.6 ± 7.3%). A difference in the amount of increase (p < 0.001) in the training load between the two methods was observed after the first 5 sessions (6.4 ± 2.8%; p < 0.001) for the 8 × 6 method with no change (0.3 ± 1.1%; p = 0.34) for the 3/7 method. Thereafter, the training load increased linearly at a similar rate (p = 0.32) for the 8 × 6 and 3/7 method (1.4 vs. 1.3%/session, respectively).
Strength-related parameters
Before training, no statistical difference (0.34 > p > 0.98) was observed between the two training groups for all dependent variables (Table 3). The 1RM load was significantly (p < 0.001) increased after the two training methods (Table 3). However, the increase was greater (p < 0.01) for the 3/7 method compared with 8 × 6 methods (22.2 ± 7.4 vs. 12.1 ± 6.6%). The MVC force of the elbow flexors was also significantly (p < 0.001) increased after the two training methods (Table 3) with a trend (p = 0.07) toward a greater gain (15.7 ± 8.2 vs. 9.5 ± 9.5%) for the 3/7 method compared with the 8 × 6 method. The gain in 1RM load was positively associated (r2 = 0.47; p < 0.001) with the increase in MVC force when data from the two methods were pooled together.
For the two training methods, both raw and normalized (% Mmax) aEMG of the elbow-flexor muscles were increased after training (p < 0.05; Table 3). When each muscle was analyzed individually, the normalized aEMG increased after training but only significantly for BB, regardless of the method. It is worth noting that if the amplitude of the Mmax recorded in BB did not change after training, it increased (p < 0.05) in BR.
The thickness of the BB long head increased significantly (p < 0.01) after training for the two methods with a greater gain (p < 0.01) for the 3/7 compared with the 8 × 6 method (9.6 ± 3.6 vs. 5.5 ± 3.7%; Table 3). The gains in 1RM load and MVC force were associated with the increase in muscle thickness when data from the two methods were pooled together (r2 = 0.38; p < 0.001 and r2 = 0.32; p < 0.01, respectively; Fig. 2). In contrast, no significant association was found between the increase in raw or normalized aEMG, and the gain in 1RM load or MVC force.

Relation between changes in muscle thickness and 1RM load (top panel) or MVC force (bottom panel). Data from the two methods were pooled together
Fatigue-related parameters
When the fatiguing test was performed at similar absolute load before and after training, the number of repetitions was greater (p < 0.001) after than before training for the two training methods (Table 4), with a greater increase (p < 0.05) for the 3/7 method compared with the 8 × 6 method (69.6 ± 46.9 vs. 31.4 ± 29.9%). This increase was positively associated with the gain in 1RM load when data from the two methods were pooled together (r2 = 0.41; p < 0.001).
When the same relative load (70% of 1RM of the testing session) was used before and after training, the number of repetitions decreased after training, regardless of the method (p < 0.001; Table 4). This decrease was greater (p = 0.05) for the 3/7 (− 17.6 ± 9.9%) than the 8 × 6 method (− 7.3 ± 16.5%). Both before and after training, an increase (p < 0.001) in aEMG of the elbow flexors throughout the fatiguing test was observed when values for BB and BR were averaged. However, for both training methods the magnitude of this increase was less after than before training (p < 0.05; Table 4). When each muscle was analyzed individually, the normalized aEMG increased (p < 0.001) from the beginning to the end of the fatiguing test (Table 4). At task failure, the BR aEMG was less increased (p < 0.001) after than before training in the two methods (Table 4). No difference was observed for the BB aEMG after the training period (Table 4).
Regardless of the training methods, TOI decreased (p < 0.001) from the beginning to the end of the fatiguing test in BB. The magnitude of change during the fatiguing test did not differ (p = 0.61) before and after training, regardless of the training group (Table 4).
Discussion
The main finding of the present study is that the 3/7 method is more effective for improving maximal strength and muscle hypertrophy than the 8 × 6 method for a similar load intensity and training volume. Furthermore, neuromuscular fatigability tested at the same relative load was similarly increased after both training methods.
Maximal strength
In agreement with previous studies (Duchateau and Hainaut 1988; Kawakami et al. 1995; Häkkinen et al. 1998; Abe et al. 2000; Aagaard et al. 2001; Erskine et al. 2014; Laurent et al. 2016), the 12-week training program increased maximal strength. As the strength-related data of the subjects that participated to the test–retest study displayed a high degree of reliability (> 0.94), we are confident that these adaptations are truly due to the training program. As expected, a greater increase in 1RM load (22.2 vs. 12.1%; p < 0.01) and a tendency of a greater gain in MVC strength (15.7 vs. 9.5%; p = 0.07) was observed for the 3/7 method compared with the 8 × 6 method. According to the principle of training specificity (Sale and MacDougall 1981; Duchateau and Baudry 2011), the greater gain in 1RM load compared with MVC force is not surprising as the training exercise was performed under dynamic contraction. Nonetheless, the gain in 1RM load was positively associated with the increase in MVC force when data from the two training groups were pooled. As a whole, the results of the present study confirm those of a field study using bench press exercise (Laurent et al. 2016) indicating that the 3/7 method is more effective to increase maximal strength than a more classical method (4 sets of 6 repetitions with 150 s rest interval between sets).
Neural adaptations
It is known for many years that a strength training program may induce neural adaptations that contribute to the increase in maximal strength (Moritani and deVries 1979; Duchateau and Hainaut 1988; Aagaard et al. 2000; Duchateau et al. 2006; Stragier et al. 2016). To avoid the confounding effect of training-related changes on muscle fibre membrane and to assess more carefully the increase in muscle activity, voluntary EMG has been normalized to the Mmax. After using this procedure, our data indicate that the aEMG of the elbow flexors increased in the two training methods suggesting an enhanced muscle activation by greater motor unit recruitment and/or discharge rate. Even if this procedure is not perfect (Keenan et al. 2005), a careful interpretation of the data suggests that neural adaptations have contributed to the increase in maximal strength following the two training methods. However, the absence of significant difference (p = 0.46) in the increase of the normalized aEMG between the two groups suggests that neural adaptations should not be responsible of the superiority of the 3/7 method.
Muscular adaptation
In response to long-term strength training, muscle hypertrophy is classically reported (Aagaard et al. 2001; Kawakami et al. 1995; Blazevich et al. 2003; Bellamy et al. 2014; Erskine et al. 2014; Damas et al. 2016). An increase in protein synthesis in response to strength training has long been considered to require the use of load greater than 60% of 1RM (i.e., mechanical stress; Vierck et al. 2000; Kraemer et al. 2002; Kraemer and Ratamess 2004). In the present study, muscle thickness of the BB long head, used as an index of muscle hypertrophy, increased significantly in both groups after 12-weeks of strength training, but to a greater extent for the 3/7 method than the 8 × 6 method (9.6 vs. 5.5%; p < 0.01). These results are in agreement with the hypothesis that brief rest interval between sets might provide a superior stimulus for muscle hypertrophy than longer rest period (Kraemer et al. 1990; Goto et al. 2004; Bottaro et al. 2009; de Salles et al. 2009; Henselmans and Schoenfeld 2014).
More recently, several papers have shown that small loads (20–40% of 1RM), mobilized under blood flow restriction, may also induce an increase in muscle mass and maximal strength (Takarada et al. 2002; Wernbom et al. 2008; Manini and Clark 2009; Dankel et al. 2017). Similarly to training under blood flow restriction, the very brief rest period (15 s) between sets in the 3/7 method, likely induced a greater cumulative fatigue-related metabolites (lactate, H+, Pi, etc.) in the involved muscles by limiting their removal compared with longer rest period. Furthermore, the incremental organisation of the repetitions within each set may have also played a role by inducing a greater level of metabolite accumulation at the end of the 5th and 10th set of the 3/7 method (Penzer et al. 2016). Even if a more intense muscle activation during the 3/7 method than the 8 × 6 method can be expected, especially during the first 5 sessions, our results support metabolic stress as an additive factor of the greater hypertrophy after the 3/7 method. This argument is reinforced by the greater deficit in tissue oxygenation recorded by NIRS during the 3/7 method (Penzer et al. 2016) and data from other studies reporting that the metabolic stress induced during the training exercise or the entire session can initiate anabolic signalling for muscle hypertrophy (Rooney et al. 1994; Schott et al. 1995; Goto et al. 2005; Nalbandian and Takeda 2016; Ozaki et al. 2016; de Freitas et al. 2017). Nevertheless, future investigations need to demonstrate whether metabolites produced during strength training have either a direct effect on the mechanisms associated with muscle hypertrophy or an indirect action through the intensification of muscle activation (Dankel et al. 2017). Although our study was not designed to elucidate the molecular mechanisms responsible for the greater increase in muscle mass with the 3/7 method, the current results demonstrate its greater efficacy to increase muscle hypertrophy and maximal strength than the 8 × 6 method.
Fatigue-related changes
When the same absolute load was used during the fatiguing test before and after training, subjects performed more repetitions in the latter condition. As the maximal strength increased after training, the load used during the fatiguing test represented a lesser percentage of the 1RM load after than before training. According to the load-repetition relation, the lesser is the load, the greater is the number of repetitions performed by the subject (Shimano et al. 2006). As the maximal strength increased to a greater extent after the 3/7 method, it is therefore not surprising that the subjects trained with this method were able to perform more repetitions after the training program than those of the 8 × 6 method.
To assess changes in neuromuscular fatigability, it is nevertheless necessary to use the same relative load before and after training to take into account the training-related change in muscle strength. In our study, when comparing the number of repetitions for the same relative load (70% of the actual 1RM load), it decreased significantly after training regardless of the method. Furthermore, throughout the fatiguing test, the aEMG of two main elbow-flexor muscles (BB and BR) increased before and after training for both methods. The increase in aEMG likely represents an enhanced voluntary activation of the muscles associated with the recruitment of additional motor units and increase in discharge rate of the newly recruited motor units to overcome fatigue that progressively develops with repetitions (Carpentier et al. 2001).
Although consistent with the greater neuromuscular fatigability observed after the two training programs, the smaller augmentation in aEMG after training, which reflects a lesser muscle activation at the end of the fatiguing test, is surprising. A first explanation may be related to the fact that, in addition to possible alterations in neuromuscular propagation and adjustments in the central nervous system, surface EMG is also influenced by the loss of signal due to amplitude cancellation that occurs when overlapping positive and negative phases of muscle action potentials are summated (Keenan et al. 2005). As the amount of cancellation increases during fatiguing contractions, due to the slowing of the action potential of the individual motor units, this might explain the lowest increase in elbow flexors aEMG after than before training for the two methods. However, the similar overall reduction in the increase of both BB and BR aEMG at the end of the fatiguing test after both training methods, while the number of repetitions was reduced to a greater extent for the 3/7 method than the 8 × 6 method, cannot explain these results.
Another potential reason for the greater neuromuscular fatigability after training may be due to an enhanced intramuscular pressure due to the increase in muscle mass that should reduce blood flow in the elbow-flexor muscles (Vedsted et al. 2006; Hunter 2014). Indeed, the greater reduction in muscle oxygenation, as attested by the reduction in TOI (Table 4), and the progressive accumulation of metabolites may have interfered with the contractile function (Russ and Kent-Braun 2003) and limited muscle activation through the inhibitory action of metaboreceptors (Kaufman and Forster 1996; Taylor et al. 2016). These greater metabolic alterations likely induced an early cessation of the task, explaining the lower number of repetitions after than before training. However, if this hypothesis might explain the slightly greater increase in neuromuscular fatigability after training with the 3/7 method compared with the 8 × 6 method, as muscle thickness and strength augmented to a greater extent after the 3/7 method, in contrast no difference in aEMG and TOI changes was observed. A lack of sensitivity of these techniques to measure accurately motor unit activation and subtle intramuscular changes is not excluded and further studies are thus needed to document these observations. Regardless of the underlying mechanisms, this increase in fatigability after the training program indicates that despite the enhanced maximal strength, it is at the expense of neuromuscular endurance, as already reported for the quadriceps by Izquierdo and colleagues (2011).
Study limitations
Two limitations must be acknowledged. First, we chose the BB long head as the representative muscle of the overall hypertrophy of the elbow flexors in response to the strength training programs. This muscle is only one part of the four major components of the elbow flexors (short and long heads of BB, BR and brachialis), with the brachialis contributing for almost the same force as the entire BB (Edgerton et al. 1986; Kawakami et al. 1994). In that context, it has been reported that the magnitude of the activation of each elbow flexor muscles can differ in function of the characteristics of the movement (i.e., load, speed, contraction type, muscle length, etc.; see Brown et al. 1993; Kulig et al. 2001). However, despite potential specific training-related adaptations, a similar hypertrophy has been reported for BB, BR and brachialis following 12 weeks of strength training against a load of ~ 70% of 1RM (Erskine et al. 2014). Therefore and as the general movement pattern was similar in our two training methods, we assume that the observed difference in muscle hypertrophy in the BB long head was representative of the whole elbow flexors.
Second, it can be argued that the superiority of the 3/7 method over the 8 × 6 method may be simply due because, in the latter, the load was not optimal and sets were not performed until failure. However, this was also the case in the first sets of each bout of the 3/7 method and, it is worth noting, that some of our subjects were unable to complete the total workload of the 8 × 6 method during the first few training sessions without a small help of the supervisor in the last few sets. Furthermore, the greater increase in load after the first 5 training sessions for the 8 × 6 method likely minimized differences in workload between the two methods in the last 19 sessions. These arguments and the absence of clear evidence that muscular failure during strength training is absolutely necessary for maximizing hypertrophy and increase in muscle strength in untrained individuals (Nobrega and Libardi 2016; de Salles et al. 2009), we believe the superiority of the 3/7 method should not simply be due to a suboptimal loading during the 8 × 6 method.
Conclusion
The results of the present study extend the superiority of the 3/7 methods over the 8 × 6 method for a similar training volume (Laurent et al. 2016). As neural adaptation was similar after the two strength training methods, the greater efficacy of the 3/7 method was mainly due to changes located at muscular level. It is therefore suggested that the 3/7 method offers a more effective combination of mechanical and metabolic stimuli to induce muscular adaptations. However, the neuromuscular fatigability, assessed at the same relative load, was increased after both training methods and contrary to our expectation, without any superiority of the 3/7 method over the 8 × 6 method in the improvement of local muscle endurance.
Abbreviations
- 1RM:
-
One repetition maximal
- aEMG:
-
Averaged value of the rectified EMG
- ANOVA:
-
Analysis of variance
- BB:
-
Biceps brachii
- BR:
-
Brachioradialis
- CV:
-
Coefficient of variation
- EMG:
-
Electromyography
- ICC:
-
Intraclass correlation coefficient
- M max :
-
Maximal motor wave
- MVC:
-
Maximal voluntary contraction
- NIRS:
-
Near-infrared spectroscopy
- TOI:
-
Tissue oxygenation index
References
Aagaard P, Simonsen EB, Andersen JL, Magnusson SP, Halkjaer-Kristensen J, Dyhre-Poulsen P (2000) Neural inhibition during maximal eccentric and concentric quadriceps contraction: effects of resistance training. J Appl Physiol 89:2249–2257
Aagaard P, Andersen JL, Dyhre-Poulsen P, Leffers AM, Wagner A, Magnusson SP, Halkjaer-Kristensen J, Simonsen EB (2001) A mechanism for increased contractile strength of human pennate muscle in response to strength training: changes in muscle architecture. J Physiol 534:613–623
Abe T, DeHoyos DV, Pollock ML, Garzarella L (2000) Time course for strength and muscle thickness changes following upper and lower body resistance training in men and women. Eur J Appl Physiol 81:174–180
Baudry S, Sarrazin S, Duchateau J (2013) Effects of load magnitude on muscular activity and tissue oxygenation during repeated elbow flexions until failure. Eur J Appl Physiol 113:1895–1904
Bellamy LM, Joanisse S, Grubb A, Mitchell CJ, McKay BR, Phillips SM, Baker S, Parise G (2014) The acute satellite cell response and skeletal muscle hypertrophy following resistance training. PLoS One 9:e109739
Bendahan D, Chatel B, Jue T (2017) Comparative NMR and NIRS analysis of oxygen-dependent metabolism in exercising finger flexor muscles. Am J Physiol Regul Integr Comp Physiol 313:R740–R753
Blazevich AJ, Gill ND, Bronks R, Newton RU (2003) Training-specific muscle architecture adaptation after 5-wk training in athletes. Med Sci Sports Exerc 35:2013–2022
Bottaro M, Martins B, Gentil P, Wagner D (2009) Effects of rest duration between sets of resistance training on acute hormonal responses in trained women. J Sci Med Sport 12:73–78
Brown JM, Solomon C, Paton M (1993) Further evidence of functional differentiation within biceps brachii. Electromyogr Clin Neurophysiol 33:301–309
Carpentier A, Duchateau J, Hainaut K (2001) Motor unit behaviour and contractile changes during fatigue in the human first dorsal interosseus. J Physiol 534:903–912
Damas F, Phillips SM, Libardi CA, Vechin FC, Lixandrão ME, Jannig PR, Costa LA, Bacurau AV, Snijders T, Parise G, Tricoli V, Roschel H, Ugrinowitsch C (2016) Resistance training-induced changes in integrated myofibrillar protein synthesis are related to hypertrophy only after attenuation of muscle damage. J Physiol 594:5209–5222
Dankel SJ, Mattocks KT, Jessee MB, Buckner SL, Mouser JG, Loenneke JP (2017) Do metabolites that are produced during resistance exercise enhance muscle hypertrophy? Eur J Appl Physiol 117:2125–2135
Davis ML, Barstow TJ (2013) Estimated contribution of haemoglobin and myoglobin to near infrared spectroscopy. Respir Physiol Neurobiol 186:180–187
de Salles BF, Simão R, Miranda F, Novaes Jda S, Lemos A, Willardson JM (2009) Rest interval between sets in strength training. Sports Med 39:765–777
de Freitas MC, Gerosa-Neto J, Zanchi NE, Lira FS, Rossi FE (2017) Role of metabolic stress for enhancing muscle adaptations: practical applications. World J Methodol 7:46–54
Duchateau J, Baudry S (2011) Training adaptation of the neuromuscular system. In: Komi PV (ed) Neuromuscular aspects of sport performance. Wiley-Blackwell, Oxford, pp 216–253
Duchateau J, Hainaut K (1988) Training effects of sub-maximal electrostimulation in a human muscle. Med Sci Sports Exerc 20:99–104
Duchateau J, Semmler JG, Enoka RM (2006) Training adaptations in the behavior of human motor units. J Appl Physiol 101:1766–1775
Edgerton VR, Roy RR, Apor P (1986) Specific tension of human elbow flexor muscles. In: Saltin B (ed) Biochemistry of exercise VI. Human Kinetics, Champaign, pp 487–500
Erskine RM, Fletcher G, Folland JP (2014) The contribution of muscle hypertrophy to strength changes following resistance training. Eur J Appl Physiol 114:1239–1249
Ferrari M, Mottola L, Quaresima V (2004) Principles, techniques, and limitations of near infrared spectroscopy. Can J Appl Physiol 29:463–487
Goto K, Nagasawa M, Yanagisawa O, Kizuka T, Ishii N, Takamatsu K (2004) Muscular adaptations to combinations of high- and low-intensity resistance exercises. J Strength Cond Res 18:730–737
Goto K, Ishii N, Kizuka T, Takamatsu K (2005) The impact of metabolic stress on hormonal responses and muscular adaptations. Med Sci Sports Exerc 37:955–963
Häkkinen K, Newton RU, Gordon SE, McCormick M, Volek JS, Nindl BC, Gotshalk LA, Campbell WW, Evans WJ, Häkkinen A, Humphries BJ, Kraemer WJ (1998) Changes in muscle morphology, electromyographic activity, and force production characteristics during progressive strength training in young and older men. J Gerontol A Biol Sci Med Sci 53:415–423
Henselmans M, Schoenfeld BJ (2014) The effect of inter-set rest intervals on resistance exercise-induced muscle hypertrophy. Sports Med 44:1635–1643
Hunter SK (2014) Sex differences in human fatigability: mechanisms and insight to physiological responses. Acta Physiol (Oxf) 210:768–789
Izquierdo M, González-Izal M, Navarro-Amezqueta I, Calbet JA, Ibañez J, Malanda A, Mallor F, Häkkinen K, Kraemer WJ, Gorostiaga EM (2011) Effects of strength training on muscle fatigue mapping from surface EMG and blood metabolites. Med Sci Sports Exerc 43:303–311
Kaufman MP, Forster HV (1996) Reflexes controlling circulatory, ventilatory and airway responses to exercise. In: Rowell LB, Shepherd JT (eds) Handbook of physiology section 12: exercise: regulation and integration of multiple systems. Oxford University Press, New York, pp 381–447
Kawakami Y, Nakazawa K, Fujimoto T, Nozaki D, Miyashita M, Fukunaga T (1994) Specific tension of elbow flexor and extensor muscles based on magnetic resonance imaging. Eur J Appl Physiol Occup Physiol 68:139–147
Kawakami Y, Abe T, Kuno SY, Fukunaga T (1995) Training-induced changes in muscle architecture and specific tension. Eur J Appl Physiol Occup Physiol 72:37–43
Keenan KG, Farina D, Maluf KS, Merletti R, Enoka RM (2005) Influence of amplitude cancellation on the simulated surface electromyogram. J Appl Physiol 98:120–131
Kraemer W, Ratamess N (2004) Fundamentals of resistance training: Progression and exercise prescription. Med Sci Sports Exerc 36:674–688
Kraemer WJ, Marchitelli L, Gordon SE, Harman E, Dziados JE, Mello R, Frykman P, McCurry D, Fleck SJ (1990) Hormonal and growth factor responses to heavy resistance exercise protocols. J Appl Physiol 69:1442–1450
Kraemer WJ, Adams K, Cafarelli E, Dudley GA, Dooly C, Feigenbaum MS, Fleck SJ, Franklin B, Fry AC, Hoffman JR, Newton RU, Potteiger J, Stone MH, Ratamess NA, Triplett-McBride T (2002) American College of Sports Medicine. American College of Sports Medicine position stand. Progression models in resistance training for healthy adults. Med Sci Sports Exerc 34:364–380
Kulig K, Powers CM, Shellock FG, Terk M (2001) The effects of eccentric velocity on activation of elbow flexors: evaluation by magnetic resonance imaging. Med Sci Sports Exerc 33:196–200
Laurent C, Penzer F, Letroye B, Carpentier A, Baudry S, Duchateau J (2016) Effect of a strength training method characterized by an incremental number of repetitions across sets and a very short rest interval. Sci Sports 31:115–121
Manini TM, Clark BC (2009) Blood flow restricted exercise and skeletal muscle health. Exerc Sport Sci Rev 37:78–85
Moritani T, deVries HA (1979) Neural factors versus hypertrophy in the time course of muscle strength gain. Am J Phys Med 58:115–130
Nalbandian M, Takeda M (2016) Lactate as a signaling molecule that regulates exercise-induced adaptations. Biology (Basel) 5(4):38
Nobrega SR, Libardi CA (2016) Is resistance training to muscular failure necessary? Front Physiol 7:10
Ogborn D, Schoenfeld B (2014) The role of fiber types in muscle hypertrophy: implications for loading strategies. Strength Cond J 36:20–25
Ozaki H, Loenneke JP, Buckner SL, Abe T (2016) Muscle growth across a variety of exercise modalities and intensities: contributions of mechanical and metabolic stimuli. Med Hypotheses 88:22–26
Penzer F, Cabrol A, Baudry S, Duchateau J (2016) Comparison of muscle activity and tissue oxygenation during strength training protocols that differ by their organisation, rest interval between sets, and volume. Eur J Appl Physiol 116:1795–1806
Rooney KJ, Herbert RD, Balnave RJ (1994) Fatigue contributes to the strength training stimulus. Med Sci Sports Exerc 26:1160–1164
Russ DW, Kent-Braun JA (2003) Sex differences in human skeletal muscle fatigue are eliminated under ischemic conditions. J Appl Physiol 94:2414–2422
Sale D, MacDougall D (1981) Specificity in strength training: a review for the coach and athlete. Can J Appl Sport Sci 6:87–92
Schoenfeld BJ (2013) Potential mechanisms for a role of metabolic stress in hypertrophic adaptations to resistance training. Sports Med 43:179–194
Schott J, McCully K, Rutherford OM (1995) The role of metabolites in strength training. II. Short versus long isometric contractions. Eur J Appl Physiol Occup Physiol 71:337–341
Shimano T, Kraemer WJ, Spiering BA, Volek JS, Hatfield DL, Silvestre R, Vingren JL, Fragala MS, Maresh CM, Fleck SJ, Newton RU, Spreuwenberg LP, Häkkinen K (2006) Relationship between the number of repetitions and selected percentages of one repetition maximum in free weight exercises in trained and untrained men. J Strength Cond Res 20:819–823
Spiering BA, Kraemer WJ, Anderson JM, Armstrong LE, Nindl BC, Volek JS, Maresh CM (2008) Resistance exercise biology: manipulation of resistance exercise programme variables determines the responses of cellular and molecular signalling pathways. Sports Med 38:527–540
Stragier S, Baudry S, Poortmans J, Duchateau J, Carpentier A (2016) Leucine-enriched protein supplementation does not influence neuromuscular adaptations in response to a 6-month strength training programme in older adults. Exp Gerontol 82:58–66
Takarada Y, Sato Y, Ishii N (2002) Effects of resistance exercise combined with vascular occlusion on muscle function in athletes. Eur J Appl Physiol 86:308–314
Taylor JL, Amann M, Duchateau J, Meeusen R, Rice CL (2016) Neural contributions to muscle fatigue: from the brain to the muscle and back again. Med Sci Sports Exerc 48:2294–2306
Vedsted P, Blangsted AK, Søgaard K, Orizio C, Sjøgaard G (2006) Muscle tissue oxygenation, pressure, electrical, and mechanical responses during dynamic and static voluntary contractions. Eur J Appl Physiol 96:165–177
Vierck J, O’Reilly B, Hossner K, Antonio J, Byrne K, Bucci L, Dodson M (2000) Satellite cell regulation following myotrauma caused by resistance exercise. Cell Biol Int 24:263–272
Wernbom M, Augustsson J, Raastad T (2008) Ischemic strength training: a low-load alternative to heavy resistance exercise? Scand J Med Sci Sports 18:401–416
Acknowledgements
The authors thank Angélique Manier, Maxime Tomi and Joachim Ribanje for their assistance in supervising training sessions and in collecting data.
Funding
This study was supported by a grant of the Sports Ministry of the Wallonia-Brussels Federation of Belgium.
Author information
Authors and Affiliations
Contributions
JD, SB, AC conceived the study. SS collected and analysed the data and prepared the figures. All authors interpreted the results, contributed to the writing of the paper and edited the final draft of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Olivier Seynnes.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Stragier, S., Baudry, S., Carpentier, A. et al. Efficacy of a new strength training design: the 3/7 method. Eur J Appl Physiol 119, 1093–1104 (2019). https://doi.org/10.1007/s00421-019-04099-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-019-04099-5