
Abstract
Introduction
The purpose of this study was to investigate the relationship between running performance (best official record in a 5000 m race) and elastic properties of tendon structures in knee extensors and plantar flexors among a large population of long distance runners.
Methods
Sixty-four highly trained male long distance runners participated in this study. Elongation of tendon structures in the knee extensors and plantar flexors was measured using ultrasonography while subjects performed ramp isometric contractions up to the voluntary maximum. The relationship between the estimated muscle force and tendon elongation was fit to a linear regression, the slope of which was defined as the stiffness of the tendon structures. Muscle and tendon thicknesses were also measured in the knee extensors and plantar flexors using ultrasonography.
Results
The best official record in a 5000 m race was negatively correlated to the stiffness of the tendon structures in the knee extensors (r = −0.341, p < 0.01), whereas it was positively correlated to the stiffness of the tendon structures in the plantar flexors (r = 0.414, p < 0.001). In both the knee extensors and plantar flexors, the other measured variables of muscle and tendon structures did not correlate with the best official record in a 5000 m race.
Conclusion
These results suggested that better long distance runners had stiffer tendon structures in the knee extensors and more compliant ones in the plantar flexors. These results implied that tendon structures in knee extensors and plantar flexors adapted to endurance running training and contributed to running performance of long distance runners separately.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tendon elasticity is important for both muscle power production and efficiency during various exercises, such as jumping and running (e.g., Cavagna 1977). Recent advances in technology have made it possible to investigate the dynamics of muscle and tendon in vivo (e.g., Fukunaga et al. 2001). Previous studies have demonstrated that muscle fibers contract nearly isometrically just after the end of the transition from dorsi to plantar flexion during an ankle bending exercise (Kubo et al. 2000b) and counter-movement jumps (Kawakami et al. 2002). This behavior of the muscle fibers was related to greater muscle power and the elastic recoil of tendon structures during the concentric phase of the movement. Furthermore, in the stance phase, the muscle fibers contracted nearly isometrically during walking and shortened during running, whereas the tendon structures were stretched (Farris and Sawicki 2012; Fukunaga et al. 2001; Ishikawa and Komi 2007). Indeed, we reported that the stiffness of tendon structures was inversely correlated to the relative difference in jump height between the jumps performed with and without counter-movements, i.e., pre-stretch augmentation (Kubo et al. 1999, 2000a, 2005, 2007b). Running induces stretch–shortening cycles in the muscle–tendon complex in the lower limbs, in which the lengthening and shortening actions of the muscle–tendon complex are repeated. Based on these findings, the elastic properties of tendon structures in lower limbs would contribute to store and reuse the elastic energy during endurance running. Therefore, we expected that lower tendon stiffness in lower limbs was related to the higher efficiency and performance for long distance runners.
To date, however, previous findings concerning the relationship between tendon elasticity and running performance are conflicting (Arampatzis et al. 2006; Fletcher et al. 2010; Kubo et al. 2010b). We previously reported that lower tendon stiffness in both the knee extensors and plantar flexors was significantly correlated to the best official record in a 5000 m race (Kubo et al. 2010b). Arampatzis et al. (2006) showed that the knee extensors of the most economical runners had a more compliant tendon and aponeurosis, whereas they showed higher contractile strength and higher tendon stiffness for plantar flexors. Fletcher et al. (2010) demonstrated that the higher stiffness of the Achilles tendon was associated with lower oxygen consumption (i.e., higher running economy). In addition, according to the longitudinal study (Albracht and Arampatzis 2013), isometric training for plantar flexor muscles increased tendon-aponeurosis stiffness, and thus improved running economy. Although the reasons for the discrepancy among previous studies are unknown, the influence of tendon elasticity on running performance may have differed between the knee extensors and plantar flexors. Furthermore, since the number of subjects participating in these studies was small; therefore, whether the elastic properties of tendon structures are correlated to the running performance of long distance runners currently remains unclear. Accordingly, it is necessary to determine the relationship between the tendon properties and running performance with a large sample of long distance runners.
In the present study, we investigated the relationship between running performance (best official record in a 5000 m race) and the elastic properties of tendon structures in both the knee extensors and plantar flexors among a large population of long distance runners.
Methods
Subjects
Sixty-four highly trained male long distance runners participated in this study. The duration of training experience ranged from 2.5 to 14.5 years (7.3 ± 2.5 years). They had participated in competitive meets at the regional or intercollegiate level within the preceding year. The best official record in a 5000 m race within 1 year prior to these tests ranged from 14:11 to 16:16 (14:54 ± 0:28) (min:s). They were with the track and field club of University, and their major goal is to win at the “Hakone Ekiden” (this relay race, about 20 km every one section, has been held on January 2nd and 3rd every year). They were not the specialists of a 5000 m race, since they have take part in various events, e.g., 5000 m race, half-marathon, etc. They performed training (mainly endurance training) at least 6 days/week for up to 4 h/day. The subjects were fully informed of the procedures to be utilized as well as the purpose of this study. Written informed consent was obtained from all subjects. This study was approved by the office of the Department of Sports Sciences, The University of Tokyo, and complied with their requirements for human experimentation.
Muscle and tendon thicknesses
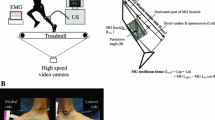
The muscle thicknesses of the knee extensors (three anatomical sites) and plantar flexors (three anatomical sites) were measured with an ultrasonic apparatus (SSD-900, Aloka, Japan). Subjects remained in a supine position for the measurements of the knee extensors and a prone position for those of the plantar flexors at 90° (anatomical position) of ankle joint with legs straight and the muscles relaxed. The anthropometric locations of the measurement sites were first precisely determined and marked before the ultrasonic measurement. A transducer with a 7.5 MHz scanning head was coated with water-soluble transmission gel, which provided acoustic contact without depressing the dermal surface. The thickness of each site was measured to the nearest 0.1 mm. The anatomical sites for the measurements are noted below (Fig. 1). In the knee extensors, muscle thickness values at the central (rectus femoris and vastus intermedius muscles), lateral (vastus lateralis and vastus intermedius muscles), and medial (vastus medialis and vastus intermedius) sides at the level of 50 % of the distance between the lateral condyle of the femur and the greater trochanter were adopted as representative of each part. In the plantar flexors, muscle thickness values at the levels of 30 % (medial gastrocnemius muscle, lateral gastrocnemius muscle) and 50 % (soleus muscle) of the distance between the popliteal crease and the center of the lateral malleolus were adopted as representative of each muscle. For medial gastrocnemius and lateral gastrocnemius muscles, the mediolateral width of each muscle was measured over the skin at that level, and then the position of one-half of the width was chosen as the measured site for each muscle (Kawakami et al. 1998). For soleus muscle, the position of the greatest thickness in the lateral half of the muscle was chosen as the measured site at the level as mentioned earlier (Kawakami et al. 1998). Furthermore, the mean value of three measured sides (for knee extensors) and muscles (for plantar flexors) was adopted as the muscle size of the knee extensors and plantar flexors, respectively.

Thick bars represent the locations of sonographic scanning sites for knee extensors (three sites) and plantar flexors (three sites)
In addition, the thicknesses of the patellar and Achilles tendons were measured at 50 % of the patellar tendon length and at the height of the lateral malleolus of the Achilles tendon, respectively. The knee and ankle joint angles were 90° during the measurement of tendon thickness.
Elastic properties of tendon structures
A maximal voluntary isometric contraction (MVC) was measured by means of specially designed dynamometers (Applied Office, Tokyo, Japan) for knee extension and plantar flexion. All measurements were performed on the right lower limb. During each task, subjects exerted isometric torque from zero (relax) to MVC within 5 s. Torque signals were amplified and sampled at 1 kHz using a 16-bit A/D converter (PowerLab/16SP, AD Instruments, Australia). The hips and back were held tightly in the seat using adjustable lap belts during the knee extension task. The right ankle was firmly attached to the lever arm of the dynamometer with a strap and fixed with the knee joint flexed at an angle of 90° (full extension = 0°). During the plantar flexion task, subjects lay prone on the test bench with the waist and shoulders being secured by adjustable lap belts and held in position. The ankle joint was set at 90° with the knee joint at full extension, and the right foot was securely strapped to a footplate connected to the lever arm of the dynamometer.
The elongation of tendon structures (outer tendon and aponeurosis) in the knee extensors and plantar flexors was assessed during isometric contractions. An ultrasonic apparatus (SSD-6500, Aloka, Tokyo, Japan) with an electronic linear array probe (7.5 MHz wave frequency with 80 mm scanning length; UST 5047-5, Aloka) was used to obtain longitudinal ultrasonic images of the vastus lateralis and medial gastrocnemius muscles by procedures described previously (Kubo et al. 2010b). Two measured sites were selected for measurements: at 50 % of the distance between the greater trochanter and the lateral epicondyle of the femur for the vastus lateralis muscle and at 30 % of the distance between the popliteal crease and the centre of the lateral malleolus for the medial gastrocnemius muscle. The probe was longitudinally attached to the dermal surface of the subject with adhesive tape, which prevented the probe from sliding. A marker was placed between the skin and the ultrasonic probe as the landmark to confirm that the probe did not move during measurements (Kubo et al. 2000a). Ultrasonic images were recorded on videotape at 30 Hz and synchronized with recordings of a clock timer for subsequent analyses. The point at which one fascicle was attached to the aponeurosis was visualized on ultrasonic images. The displacement of this point was considered to indicate lengthening of the deep aponeurosis and distal tendon. To correct measurements taken for tendon and aponeurosis elongation, additional measurements were performed under passive conditions (Kubo et al. 2010b). In each subject, the displacement of each site obtained from ultrasonic images could be corrected for that attributed to joint rotation alone. In this study, only values corrected for angular rotation were reported. The elongation of tendon structures (L) was converted to strain by the following equation (Kubo et al. 2007a):
where TL is the length of the tendon structure at rest. We measured the distance between the measurement site for L and the insertion of the patellar and Achilles tendons (confirmed using ultrasonography).
Torque (TQ) measured during isometric contractions was converted to muscle force (Fm) by the following equation (Kubo et al. 2010b):
where k is the relative contribution of physiological cross-sectional area in vastus lateralis muscle within knee extensors (Akima et al. 2001) and medial gastrocnemius muscle within plantar flexors (Fukunaga et al. 1996), and MA is the moment arm length in quadriceps femoris muscles at 90° and triceps surae muscle at 90°, which was estimated from the limb length of each subject (Greive et al. 1978; Visser et al. 1990). In this study, the slope of Fm–L curve above 50 % of MVC (about 20 points) was defined as stiffness (Kubo et al. 2010b), which in all case exceeded r 2 = 0.95. The reliability of tendon stiffness measurements using ultrasonography has been confirmed in our previous studies (Kubo et al. 2001).
Statistics
Descriptive data represent mean ± SD. Significant differences in the physical characteristics, duration of training experience, and measured variables of muscles and tendon structures between the Faster and Slower groups (described in “Results”) were examined by an unpaired t test. Pearson product–moment correlation coefficients were computed to assess the relationships among the measured variables of muscles and tendon structures and the best official record in a 5000 m race. The level of significance was set at p < 0.05.
Results
The best official records in a 5000 m race were 14:34 (range 14:11–14:47) in the Faster group and 15:14 (range 14:48–16:16) in the Slower group (p < 0.001). No significant differences were observed in age or physical characteristics between the Faster and Slower groups, while the duration of training experience was significantly longer for the Faster group than for the Slower group (Table 1).
No significant differences were noted in MVC values in either the knee extensors or plantar flexors between the two groups (Table 2). Muscle thicknesses for the soleus muscle (p = 0.020) and entire of the plantar flexors (p = 0.024) were significantly greater in the Faster group than in the Slower group; however, no significant differences were noted in the muscle thickness of any site or entire of the knee extensors between the two groups.
In the knee extensors, the elongation of tendon structures was slightly lower in the Faster group than in the Slower group, although no significant differences were observed in the elongation of tendon structures between the two groups at any force production level in the plantar flexors (Fig. 2). The stiffness of the tendon structures in the knee extensors was significantly higher in the Faster group than in the Slower group (p = 0.022), whereas that in the plantar flexors was significantly lower in the Faster group than in the Slower group (p = 0.010) (Table 3; Fig. 1). No significant differences in initial length, maximal elongation and strain of tendon structures, and tendon thickness were found in either the knee extensors or plantar flexors between the two groups (Table 3).

Relationship between estimated muscle force and the elongation of tendon structures in knee extensors (a) and plantar flexors (b) for the Faster (open) and Slower (closed) groups. Values are mean ± SD. Asterisk indicates significantly different from the Slower group at p < 0.05
The best official record in a 5000 m race was negatively correlated to the stiffness of tendon structures in the knee extensors, but was positively correlated to the stiffness of tendon structures in the plantar flexors (Table 4; Fig. 3). In both the knee extensors and plantar flexors, the other measured variables of muscle and tendon structures did not correlate with the best official record in a 5000 m race (Table 4).

Relationship between the best official record in a 5000 m race and stiffness in tendon structures in knee extensors (a) and plantar flexors (b)
Discussion
The main result of this study was that better long distance runners had stiffer tendon structures in their knee extensors and more compliant ones in their plantar flexors. These results implied that the relationship between tendon properties and running performance would differ between the knee extensors and plantar flexors.
In the knee extensors, the stiffness of tendon structures was significantly higher in the Faster group than in the Slower group (Table 3; Fig. 2a). Furthermore, we found that the best official record in a 5000 m race was negatively correlated to the stiffness of tendon structures (Table 4; Fig. 3a). These results conflicted with the findings of Arampatzis et al. (2006) and Kubo et al. (2010b). Possible reasons for this discrepancy are the small sample size and differences in the best record of track-and-field events. In our previous study (Kubo et al. 2010b), the number of subjects was only 15, and the range of the best official record of a 5000 m race was very narrow (from 14:11 to 14:53 min:s). In Arampatzis et al. (2006), the number of subjects in each group (10 runners in the high running economy group, 12 runners in the moderate running economy group, and 6 runners in the low running economy group) was also smaller, and running performance would be lower according to the training volume and body mass of the subjects. Our previous studies showed that maximal elongation of tendon structures in the knee extensors was significantly lower in long distance runners than in untrained subjects (Kubo et al. 2000a, 2010b). These previous findings implied that greater mechanical stresses imposed on the knee extensors during long-term running training were responsible for the differences in the tendon properties in the knee extensors between long distance runners and untrained subjects. Therefore, since the faster runners had performed large amounts of running training, their tendon structures in the knee extensors may have been stiffer due to the greater mechanical stresses imposed by long-term running training. The duration of training experience was significantly longer in the Faster group than in the Slower group (Table 1), although no significant correlation coefficient was found between tendon stiffness in the knee extensors and the duration of training experience (data not shown; r = −0.085, p = 0.504). However, we currently do not have concrete evidence for this speculation because the training volume of each runner during a long-term period was unknown. Furthermore, to clarify this point, we need to investigate the genetic effects on the tendon properties for long distance runners in the future study.
Previous studies suggested that an increase in the stiffness of the tendon structures resulted in a higher rate of force development and shorter time to peak torque and electromechanical delay (Bojsen-Moller et al. 2005; Kubo et al. 2001; Ohta 2012). However, it is currently unknown whether stiffer tendon structures in the knee extensors make the transmission of exerted muscle force more efficient during running. Fujimoto et al. (2000) reported using positron emission tomography that glucose uptake by the knee extensors was lower than that by the plantar flexors. Kyrolainen et al. (2005) also indicated that the activation level of the knee extensor muscles was lower than that of the plantar flexor muscles at the stance phase during running. In addition, the positive work done during stance phase of running was considerably lower at the knee joint than at the ankle joint, although no difference in the negative work between the knee and ankle joints (Schache et al. 2011). Considering these findings, it is possible that the elastic properties of tendon structures in the knee extensors do not so much contributed to store and reuse the elastic energy during endurance running, although this point has not yet been proven experimentally.
Another important result of the present study was that the stiffness of tendon structures in the plantar flexors was significantly lower in the Faster group than in the Slower group (Table 3; Fig. 2b). Furthermore, we found that the best official record in a 5000 m race was positively correlated to the stiffness of tendon structures in the plantar flexors (Table 4; Fig. 3b). These results agreed with our previous finding (Kubo et al. 2010b). The findings of musculoskeletal modeling studies (Hof et al. 2002; Sasaki and Neptune 2006) and cadaveric studies (Alexander and Bennet-Clark 1977; Ker et al. 1987) showed that the lower metabolic energy expenditure by muscle fibers during walking and running was associated with the elastic energy stored in the Achilles tendon. Furthermore, the previous studies using ultrasonography demonstrated that the muscle fibers shortened during the first half of stance phase (braking phase) of running and jumping, whereas the tendon structures were stretched (Ishikawa and Komi 2007; Farris and Sawicki 2012). This would mean that the elastic properties of tendon structures contributed to store the elastic energy during exercises. During the concentric phase (propulsion phase) of running and jumping, the rapid shortening of tendon structures played a role in lowering the velocity of muscle fibers (Kawakami et al. 2002; Kubo et al. 2000a). This will enable the working muscles to develop more tension and power in relation to their force–velocity properties. These findings together with the present results suggested that the lower tendon stiffness in the plantar flexors is suitable for storing elastic energy at the stance phase during running, and, thus, be contributed to achieving higher official record in a 5000 m race. On the other hand, the present result on the tendon structures in the plantar flexors disagreed with the other previous findings (Albracht and Arampatzis 2013; Fletcher et al. 2010). Albracht and Arampatzis (2013) reported that isometric training for plantar flexor muscles increased tendon–aponeurosis stiffness, and thus improved running economy. Possible reasons for the discrepancy are the differences in the competition ability in the long distance race event and the index of running performance (the best official record in a 5000 m race in the present study, running economy in the previous studies). Although this may be affected to hysteresis in tendon structures, this was not calculated in the present study. Finni et al. (2013) reported that the large variability in hysteresis in human tendons was caused by individual differences or methodological uncertainties. In future studies, a reliable technique needs to be employed to measure hysteresis in human tendon structures in vivo.
The present results indicated that the relationship between tendon properties and running performance differed between the knee extensors and plantar flexors. Although the underlying mechanisms responsible for these results are unknown, they may be due to differences in the “plasticity” of tendon structures due to long-term running training between the measured sites. We previously reported that no significant difference was observed in relative increases in the stiffness of tendon structures between the knee extensors and plantar flexors following resistance training for 3 months (Kubo et al. 2010a). However, site differences in the changes in tendon stiffness may be observed after the long-term training. Our previous studies demonstrated that the tendon structures of long distance runners were stiffer than those in untrained subjects in the knee extensors, but not in the plantar flexors (Kubo et al. 2000a, 2010b). Other studies also found no significant difference in the stress–strain relationship for the triceps surae aponeurosis and tendons between runners and non-runners (Arampatzis et al. 2007; Rosager et al. 2002). Based on these findings, we suggested that long-term running training stiffened tendon structures in the knee extensors, but not in the plantar flexors.
In addition to the tendon properties discussed above, the thickness of the plantar flexor muscles (especially the soleus muscle) was significantly greater in the Fast group than in the Slower group (Table 2). However, the muscle thickness of the plantar flexors did not correlate with the best official record in a 5000 m race (Table 4). To the best of our knowledge, few studies have investigated the relationship between muscle sizes in the lower limbs and the performance of long distance runners (Berg et al. 1998). Berg et al. (1998) reported that the cross-sectional area of lower limb muscles did not correlat with the endurance running performance (10 km run time). We previously reported that the relative muscle thickness (to body mass1/3) of long distance runners was significantly greater than that of untrained subjects in the plantar flexors, but not in the knee extensors (Kubo et al. 2010b). According to the previous studies using electromyogram and positron emission tomography (Fujimoto et al. 2000; Kyrolainen et al. 2005), the activation level of the plantar flexor muscles was greater than that of the knee extensor muscles at the stance phase during running. These previous findings and the present results suggested that long distance running training modified the plantar flexor muscles, especially the soleus muscle, which predominantly contains type I fibers. However, these training-induced changes in muscle size of plantar flexors may not be associated with the running performance of long distance runners.
In the present study, we must draw the attention to some limitations of the in vivo approach followed. First, we measured the thickness, but not cross-sectional area, of tendon as a size of tendon. Finni et al. (2009) reported using ultrasonography that the tendon thickness was significantly correlated with the tendon cross-sectional area. Therefore, we considered that this point did not affect the obtained result on the size of tendon. Second, the force–elongation relationship of the tendon structures should be converted to a stress–strain relationship to determine accurately the relationship between tendon properties and running performance. However, we could not calculate the Young’s modulus from stress–strain relationship of tendon, since we did not measure the elastic properties of outer tendon (i.e., patellar and Achilles tendons), but those of tendon structures (including outer tendon and aponeurosis). Third, the moment arm length was estimated from the lower leg length of each subject to estimate the tendon force. According to the previous studies (e.g., Maganaris et al. 1998), this assumption was not always true. The moment arm length changes of each subject during the measurement of tendon properties would be necessary for an accurate absolute tendon force determination. In the present study, however, we aimed to study whether the tendon properties were related to the endurance running performances. Fourth, there was a difference in strain rate of the tendon structures between the running and tendon measurement. Previous studies showed that loading rate during the measurement of tendon elongation did not affect the stiffness of Achilles tendon (Kubo et al. 2002; Peltonen et al. 2013). In future studies, the difference in the tendon properties between the dynamic and static conditions should be tested.
In conclusion, better long distance runners had stiffer tendon structures in the knee extensors and more compliant ones in the plantar flexors. These results implied that the greater loading imposed during long-term running training stiffened tendon structures in the knee extensors, but not in the plantar flexors. In addition, the lower stiffness of tendon structures would contribute to the running performance of long distance runners in the plantar flexors, but not in the knee extensors, since the mechanical work of ankle joint was higher than that of knee joint during endurance running (e.g., Schache et al. 2011).
Abbreviations
- Fm:
-
Muscle force
- L:
-
Elongation of tendon structures
- MA:
-
Moment arm length
- MVC:
-
Maximal voluntary contraction
- TL:
-
Length of tendon structures
- TQ:
-
Torque
References
Akima H, Kubo K, Imai M, Kanehisa H, Suzuki Y, Gunji A, Fukunaga T (2001) Inactivity and muscle: effect of resistance training during bed rest on muscle size in the lower limb. Acta Physiol Scand 172:269–278
Albracht K, Arampatzis A (2013) Exercise-induced changes in triceps surae tendon stiffness and muscle strength affect running economy in humans. Eur J Appl Physiol 113:1605–1615
Alexander RM, Bennet-Clark HC (1977) Storage of elastic strain energy in muscle and other tissues. Nature 265:114–117
Arampatzis A, De Monte G, Karamanidis K, Morey-Klapsing G, Stafilidis S, Bruggemann GP (2006) Influence of the muscle–tendon unit’s mechanical and morphological properties on running economy. J Exp Biol 209:3345–3357
Arampatzis A, Karamanidis K, Morey-Klapsing G, De Monte G, Stafilidis S (2007) Mechanical properties of the triceps surae tendon and aponeurosis in relation to intensity of sport activity. J Biomech 40:1946–1952
Berg K, Latin RW, Coffey C (1998) Relationship of somatotype and physical characteristics to distance running performance in middle age runners. J Sports Med Phys Fitness 38:253–257
Bojsen-Moller J, Magnusson SP, Rasmussen LR, Kjaer M, Aagaard P (2005) Muscle performance during maximal isometric and dynamic contractions is influenced by the stiffness of the tendinous structures. J Appl Physiol 99:986–994
Cavagna GA (1977) Storage and utilization of elastic energy in skeletal muscle. Exerc Sports Sci Rev 5:89–129
Farris DJ, Sawicki GS (2012) Human medial gastrocnemius force–velocity behavior shifts with locomotion speed and gait. Proc Natl Acad Sci 109:977–982
Finni T, Kovanen V, Ronkainen P, Pollanen E, Bashford GR, Kaprio J, Alen M, Kujala UM, Sipila S (2009) Combination of hormone replacement therapy and high physical activity is associated with differences in Achilles tendon size in monozygotic female twin pairs. J Appl Physiol 106:1332–1337
Finni T, Peltonen J, Stenroth L, Cronin NJ (2013) Viewpoint: on the hysteresis in the human Achilles tendon. J Appl Physiol 114:515–517
Fletcher JR, Esau SP, MacIntosh BR (2010) Changes in tendon stiffness and running economy in highly trained distance runners. Eur J Appl Physiol 110:1037–1046
Fujimoto T, Itoh M, Tashiro M, Yamaguchi K, Kubota K, Ohmori H (2000) Glucose uptake by individual skeletal muscles during running using whole-body positron emission tomography. Eur J Appl Physiol 83:297–302
Fukunaga T, Roy RR, Shellock FG, Hodgson JA, Edgerton VR (1996) Specific tension of human plantar flexors and dorsiflexors. J Appl Physiol 80:158–165
Fukunaga T, Kubo K, Kawakami Y, Fukashiro S, Kanehisa H, Maganaris CN (2001) In vivo behavior of human muscle tendon during walking. Proc R Soc Lond B 268:229–233
Greive DW, Pheasant S, Cavagna PR (1978) Prediction of gastrocnemius length from knee and ankle joint posture. In: Asmussen E, Jorgensen K (eds) Biomechanics, vol VI. University Park, Baltimore, pp 405–412
Hof AL, Van Zandwijk JP, Bobbert MF (2002) Mechanics of human triceps surae muscle in walking, running and jumping. Acta Physiol Scand 174:17–30
Ishikawa M, Komi PV (2007) The role of the stretch reflex in the gastrocnemius muscle during human locomotion at various speeds. J Appl Phyiol 103:1030–1036
Kawakami Y, Ichinose Y, Fukunaga T (1998) Architectural and functional features of human triceps surae muscles during contraction. J Appl Physiol 85:398–404
Kawakami Y, Muraoka T, Ito S, Kanehisa H, Fukunaga T (2002) In vivo muscle fibre behaviour during conter-movement exercise in humans reveals a significant role for tendon elasticity. J Physiol 540:635–646
Ker RF, Bennett MB, Bibby SR, Kester RC, Alexander RM (1987) The spring in the arch of the human foot. Nature 325:147–149
Kubo K (2005) In vivo elastic properties of human tendon structures in lower limb. Int J Sports Health Sci 3:143–151
Kubo K, Kawakami Y, Fukunaga T (1999) Influence of elastic properties of tendon structures on jump performance in humans. J Appl Physiol 87:2090–2096
Kubo K, Kanehisa H, Kawakami Y, Fukunaga T (2000a) Elastic properties of muscle–tendon complex in long-distance runners. Eur J Appl Physiol 81:181–187
Kubo K, Kanehisa H, Takeshita D, Kawakami Y, Fukashiro S, Fukunaga T (2000b) In vivo dynamics of human medial gastrocnemius muscle–tendon complex during stretch-shortening cycle exercise. Acta Physiol Scand 170:127–135
Kubo K, Kanehisa H, Ito M, Fukunaga T (2001) Effects of isometric training on the elasticity of human tendon structures in vivo. J Appl Physiol 91:26–32
Kubo K, Kanehisa H, Kawakami Y, Fukunaga T (2002) Measurement of viscoelastic properties of tendon structures in vivo. Scand J Med Sci Sports 12:3–8
Kubo K, Ishida Y, Komuro T, Tsunoda N, Kanehisa H, Fukunaga T (2007a) Age-related differences in the force generation capabilities and tendon extensibilities of knee extensors and plantar flexors in men. J Gerontol Med Sci 62:1252–1258
Kubo K, Morimoto M, Komuro T, Tsunoda N, Kanehisa H, Fukunaga T (2007b) Influences of tendon stiffness, joint stiffness, and electromyographic activity on jump performances using single joint. Eur J Appl Physiol 99:235–243
Kubo K, Ikebukuro T, Yata H, Tsunoda N, Kanehisa H (2010a) Effects of training on muscle and tendon in knee extensors and plantar flexors in vivo. J Appl Biomech 26:316–323
Kubo K, Tabata T, Ikebukuro T, Igarashi K, Yata H, Tsunoda N (2010b) Effects of mechanical properties of muscle and tendon on performance in long distance runners. Eur J Appl Physiol 110:507–514
Kyrolainen RJ, Watson JS, Weir J (2005) Changes in muscle activity with increasing running speed. J Sports Sci 23:1101–1109
Maganaris CN, Baltzopoulos V, Sargeant AJ (1998) Changes in Achilles tendon moment arm from rest to maximal isometric plantarflexion: in vivo observations in man. J Physiol 510:977–985
Ohta Y (2012) Effects of tendinous tissue properties on force output evoked by 2-pulse trains at different inter-pulse intervals in the human tibialis anterior muscle. Acta Bioeng Biomech 14:45–52
Peltonen J, Cronin NJ, Stenroth L, Finni T, Avela J (2013) Viscoelastic properties of the Achilles tendon in vivo. Springerplus 2:212
Rosager S, Aagaard P, Dyhre-Poulsen P, Neergaard K, Kjaer M, Magnusson SP (2002) Load-displacement properties of the human triceps surae aponeurosis and tendon in runners and non-runners. Scand J Med Sci Sports 12:90–98
Sasaki K, Neptune RR (2006) Muscle mechanical work and elastic energy utilization during walking and running near the preferred gait transition speed. Gait Posture 23:383–390
Schache AG, Blanch PD, Dorn TW, Brown NAT, Rosemond D, Pandy MG (2011) Effect of running speed on lower limb joint kinetics. Med Sci Sports Exerc 43:1260–1271
Visser JJ, Hoogkamer JE, Bobbert MF, Huijing PA (1990) Length and moment arm of human leg muscles as a function of knee and hip-joint angles. Eur J Appl Physiol 61:453–460
Acknowledgments
This study was supported by a Grant-in-Aid for Young Scientists (A) (21680047 to K. Kubo) from the Japan Society for the Promotion of Science.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Olivier Seynnes.
Rights and permissions
About this article

Cite this article
Kubo, K., Miyazaki, D., Shimoju, S. et al. Relationship between elastic properties of tendon structures and performance in long distance runners. Eur J Appl Physiol 115, 1725–1733 (2015). https://doi.org/10.1007/s00421-015-3156-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-015-3156-2