
Abstract
Purpose
The purpose of this study was to analyze the impact of an 8-week strength training program on the neuromuscular characteristics and pacing adopted by runners during a self-paced endurance running.
Methods
Eighteen endurance runners were allocated into either strength training group (STG, n = 9) or control group (CG, n = 9) and performed the following tests before and after the training period: (a) incremental test, (b) running speed-constant test, (c) 10-km running time trial, (d) drop jump test, (e) 30-s Wingate anaerobic test, (f) maximum dynamic strength test (1RM). During 1RM, the electromyographic activity was measured.
Results
In the STG, the magnitude of improvement for 1RM (23.0 ± 4.2 %, P = 0.001), drop jump (12.7 ± 4.6 %, P = 0.039), and peak treadmill speed (2.9 ± 0.8 %, P = 0.013) was significantly higher compared to CG. This increase in the 1RM for STG was accompanied by a tendency to a higher electromyographic activity (P = 0.080). The magnitude of improvement for 10-km running performance was higher (2.5 %) for STG than for CG (−0.7 %, P = 0.039). Performance was improved mainly due to higher speeds during the last seven laps (last 2800 m) of the 10-km running trial. There were no significant differences between before and after training period for maximal oxygen uptake, respiratory compensation point, running economy, and anaerobic performance for both groups (P > 0.05).
Conclusions
These findings suggest that a strength training program offers a potent stimulus to counteract fatigue during the last parts of a 10-km running race, resulting in an improved overall running performance.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Pacing strategy has been defined as the distribution of speed, and consequently energy expenditure, during an athletic competition (Abbiss and Laursen 2008), being an important determinant of athletic performance (Bertuzzi et al. 2014). It is believed that athletes choose an appropriate running speed in order to avoid premature exhaustion and therefore optimize their overall performance (Thiel et al. 2012). Some researchers have suggested that pacing might be regulated by perception of effort (Faulkner et al. 2008; St Clair Gibson et al. 2006). Faulkner et al. (2008) reported that during long-distance running, the speed distribution was characterized by a U-shaped profile (i.e., start and finish speeds higher than in the middle), while the rating of perceived exertion (RPE) increased in a linear fashion regardless of the changes in the speed. It appears that this linear RPE profile may reflect a centrally regulated system that monitors exercise intensity in order to protect whole-body homeostasis (Ulmer 1996). Therefore, athletes would adjust their running speed by comparing moment-to-moment the actual with the desirable RPE for a given distance (St Clair Gibson et al. 2006). This would implicate that the exercise intensity is continually adjusted during the race through the interpretations of multiple internal signals integrated within the central nervous system (St Clair Gibson et al. 2006).
Previous findings suggested that momentary internal signals utilized in pacing monitoring may originate mainly from the physiological and neuromuscular systems. For example, Lima-Silva et al. (2010) observed that runners with a higher running economy (RE), peak treadmill speed (PTS), and a faster speed corresponding to the onset of blood lactate accumulation were able to adopt a more aggressive U-shaped speed curve, probably due to reduced afferent signals during the race. Using a stepwise multiple regression model, a recent study showed that maximum dynamic strength (1RM), PTS, and maximal oxygen uptake (\( {\dot{\text{V}}} \)O2max) explained 80 % of the speed variation during the middle (400–9600 m) of a 10-km running time trial (Bertuzzi et al. 2014). Interestingly, PTS, which integrates aerobic power, anaerobic capacity, and neuromuscular capability, was the only variable able to predict the end-spurt (last 400 m), accounting alone for 66 % of the pacing variance. An interesting, but untested, corollary of these findings is that the interventions (e.g., physical training and dietary manipulation) were able to alter these physiological and neuromuscular characteristics, and hence, the afferent signals originating from their muscles may allow the athletes to change the pacing (Lambert et al. 2005). However, it is still unknown whether the pacing can be altered in response to adaptations in these parameters produced by physical training.
Over the past three decades, many studies have reported beneficial effects of strength training on these parameters related to pacing strategy (Hakkinen and Komi 1983; Mikkola et al. 2011; Taipale et al. 2010). For example, Støren et al. (2008) reported significant increases in 1RM (~33 %), RE (5 %), and explosive strength (~26 %) after 8 weeks of strength training performed by long-distance runners. In a similar manner, Mikkola et al. (2011) observed a significant increase in the PTS (~3 %) in long-distance runners after an 8-week strength training period, while Taipale et al. (2010) found a higher electromyographic activity (iEMG) of the lower limbs during 1RM and an improved anaerobic performance in long-distance runners after 8 weeks of strength training. These adaptations would contribute to alter motor unit recruitment patterns and the capacity to produce and maintain force application during running. This seems especially important during the final stages of a race, when athletes attain higher speeds. Previous findings have demonstrated that running speed and iEMG of the lower limbs were increased during the final lap of a 5-km running time trial, suggesting that athletes increased their muscle recruitment despite the developing muscle fatigue (Nummela et al. 2006). Thus, it could be expected that an increased force application after a strength training program may be accompanied by a faster end-spurt.
It is interesting to observe that although these findings suggest beneficial effects of the strength training on endurance performance, it is still unknown which part of the self-paced endurance running (i.e., start, middle, and end phases) could be changed by strength training and possible mechanisms by which these changes could occur. Therefore, the aim of this randomized controlled trial was to analyze the impact of a strength training program on the neuromuscular characteristics and pacing during a 10-km running time trial. In light of previous findings suggesting that the neuromuscular system is critical to maintain high running speeds during the final laps of long-distance running (Nummela et al. 2006), it was hypothesized that the athletes would adopt a more aggressive end-spurt after a strength training program due to improvements in the neuromuscular system.
Methods
Subjects
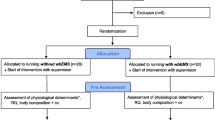
Twenty-six male recreational long-distance runners from local running clubs were invited to participate in this study. Figure 1 shows the flow of participants through the trial. Seven runners failed to complete all testing sessions and were excluded from the study. They were randomly allocated (by a computer-generated random sequence using a 1:1 ratio) into either strength training group (STG, n = 10, age 34.1 ± 7.7 years, body mass 67.5 ± 9.2 kg, height 173.7 ± 11.4 cm) or control group (CG, n = 9, age 32.9 ± 9.2 years, body mass 71.0 ± 11.3 kg, height 174.3 ± 9.9 cm). One participant belonging to the STG group discontinued the intervention because of logistical issues. Thus, the final sample size was eighteen subjects. They participated in local competitions, and their best performances in 10-km competitions ranged from 35 to 45 min. All participants performed only low-intensity continuous aerobic training (50–70 % \( {\dot{\text{V}}} \)O2max) before the beginning of the study, and they were instructed to maintain a similar aerobic training during the experimental period. The subjects’ running training volumes were expressed as the mean distances covered (Taipale et al. 2010; Sedano et al. 2013), which were assessed through a training log recorded for 3 weeks prior to the beginning of the study and for the last 3 weeks before the study completion. The exclusion criteria included previous strength or plyometric training experience, use of dietary supplement, smokers, vegetarian diet, neuromuscular disorders, and cardiovascular dysfunctions. The participants received a verbal explanation about the possible benefits, risks, and discomforts associated with the study and signed a written informed consent before participating in the study. The study was approved by the local Ethics Committee for Human Studies and was performed in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki.

Flow diagram of the progress through the phases of a parallel randomized trial of two groups
Study design
The current study was conducted using a parallel-group design as previously suggested (Schulz et al. 2010). The athletes performed the strength training (ST) program from January to July 2013 during a non-competitive season. Before and after the 8-week ST program, the runners were required to visit the laboratory on four different occasions separated by at least 72 h over a 2-week period. During the first visit, after anthropometric assessments, the participants completed a constant load and a maximal incremental treadmill test to determine cardiorespiratory parameters. A 10-km time trial on an outdoor track was performed during the second visit to determine endurance running performance and pacing strategy. Drop jump and 30-s Wingate anaerobic test were performed in the third visit to determine the ability to produce explosive strength and anaerobic performance, respectively. During the fourth visit, the maximum dynamic strength was assessed using the one-repetition maximum half-squat test (1RM). Familiarizations with the drop jump and 1RM procedures were conducted at the end of the first and second visits, with 20 min of passive recovery between them. With exception of the 10-km time trial, all tests were performed at the same time of day at a constant room temperature (20–24 °C), and at least 2 h after the last meal. The participants were instructed to refrain from any exhaustive or unaccustomed exercise 48 h before the tests and to refrain from taking nutritional supplements during the experimental period.
Anthropometric measurements
Anthropometric measurements were performed as described by Norton and Olds (1996). Participants were weighed to the nearest 0.1 kg using an electronic scale (Filizola, model ID 1500, São Paulo, Brazil). Height was measured to the nearest 0.1 cm using a stadiometer. Skinfold thickness were measured to the nearest 0.2 mm at eight body sites (i.e., triceps brachii, suprailiac, abdominal, chest, subscapular, midaxillar, anterior thigh, and calf) using a Harpenden caliper (West Sussex, UK). The median of three values was used for data analysis. Measurements were performed by an experienced investigator. Body density and body fat were estimated using the equations of Jackson and Pollock (1985), and Brozek et al. (1963), respectively.
Maximal incremental treadmill test
The subjects performed a maximal incremental test on a motor-driven treadmill (model TK35, CEFISE, Nova Odessa, Brazil). After a 3-min warm-up at 8 km h−1, the speed was increased by 1 km h−1 every 3 min until exhaustion. The participants received strong verbal encouragement to ensure attainment of maximal values. Gas exchange was measured breath-by-breath using a gas analyzer (Cortex Metalyzer 3B, Cortex Biophysik, Leipzig, Germany) and subsequently averaged over 20-s intervals throughout the test. Before each test, the gas analyzer was calibrated according to the manufacturer’s recommendations. Maximal heart rate (HRMAX) was defined as the highest value obtained at the end of the test. \( {\dot{\text{V}}} \)O2max was determined when two or more of the following criteria were met: an increase in oxygen uptake of less than 2.1 ml kg−1 min−1 between two consecutive stages, a respiratory exchange ratio greater than 1.1, and a ± 10 bpm of the predicted maximal heart rate (i.e., 220-age) (Howley et al. 1995). The respiratory compensation point (RCP) was determined by three independent investigators as the point of a nonlinear increase in the VE/VCO2, a constant increase in the VE/VO2, and the first decrease in the expiratory fraction of CO2 (Meyer et al. 2005). The highest velocity achieved during the test was recorded as the peak treadmill speed (PTS). When the participants could not complete the entire last stage (<3 min), the PTS was calculated using fractional time supported in the last stage multiplied by the increment rate.
Running economy
A standardized warm-up consisting of a 5-min run at 8 km h−1, followed by a 3-min passive recovery, was performed before the test. Treadmill speed was adjusted to 12 km h−1 after the warm-up, and the subjects ran for 6 min at this speed. The test began with the participant’s feet astride the moving belt and hands holding the handrail. The measurement of \( {\dot{\text{V}}} \)O2 was similar to the procedures adopted during the maximal incremental treadmill test. The RE was determined from the mean \( {\dot{\text{V}}} \)O2 during the last 30 s of the test.
Anaerobic performance
The 30-s Wingate anaerobic test, as described by Bar-Or (1987), was used to determine the lower limbs anaerobic performance. The test was preceded by a 5-min warm-up period against the inertial resistance of the equipment and included two 4-s sprints performed in last seconds of the second and fourth minutes. After a 10-min rest, the participants were instructed to pedal “all-out” for 30 s against a resistance of 0.075 kg kg−1 body mass. Verbal encouragement was provided throughout the test. The external power output was calculated every 1 s (Ergometric 6.0, Cefise, Nova Odessa, Brazil). The highest external power output in the first 5-s interval of the test was used to calculate the peak power (PP), whereas the average power generated over the entire 30 s corresponded to the mean power (MP).
Self-paced running endurance performance
Participants performed a 10-km running time trial on an outdoor 400-m track. They were instructed to finish the race as quickly as possible, as in a competitive event. Before the test, the participants warmed up for 10 min at 8 km h−1. They were instructed to maintain regular water consumption within 6 h of testing, and water was provided ad libitum during the entire event. Verbal encouragement was provided during the entire race. However, runners were not advised of their lap splits. Speed was registered every 100 m via a global positioning system (GPS Forerunner® 405, Garmin, Kansas City, Oregon, USA). RPE was reported by participants every 1000 m using the Borg 15-point scale (Borg 1982). Copies of this scale were laminated and reduced to 10 cm by 5-cm cards and affixed to the wrist of the dominant arm of the individuals. The 10-km running tests were performed at the same time of day and in similar conditions for the same subjects. Ambient temperature and humidity were provided by the Institute of Astronomy, Geophysics and Atmospheric Sciences of the University of São Paulo, Brazil. The mean ± SD values for temperature and humidity were 22.8 ± 3.8 °C and 61.0 ± 8.2 %, respectively.
Explosive strength
The drop jump test was used to determine the ability of the runners to produce explosive strength. The drop jump height (DJH) and contact time (CT) were measured using a contact mat (Jump System Pro, Cefise, São Paulo, Brazil). Runners stepped off a 40-cm box and attempted to achieve the greatest vertical height with a short ground contact time (close to 200 ms) (Young and Behm 2003). Subjects were instructed to minimize knee flexion and extension during the drop jump. Demonstrations were provided in all sessions by the same investigator. All jumps were performed with hands on the hips, and five repetitions were performed with a 30-s rest interval between them. The largest and smallest values were discarded, and the average of the remaining three jumps was calculated and used for statistical analysis. The jump height was determined by flight time, and the reactive strength index (RSI) was determined as jump height divided by contact time.
Maximum dynamic strength and electromyographic activity
The participants were familiarized with all the procedures, equipment, and proper exercise techniques prior to data collection. Body position and foot placement were controlled by measuring tapes fixed on both the bar and the ground, respectively. An adjustable-height wooden seat was placed behind the subjects to keep the bar displacement and knee flexion angle constant on each half-squat repetition. The subjects’ settings on the Smith machine were recorded to guarantee the same positioning across familiarization and testing sessions. The maximum dynamic strength (1RM half-squat) test was performed according to standard procedures (Brown and Weir 2001). The participants performed a brief general warm-up exercise composed of a 5-min treadmill run at 8 km h−1 followed by light lower limbs stretching. Following the initial warm-up run, the participants performed a specific warm-up protocol composed of two sets of half-squat exercise, interspaced by a 3-min interval. In the first set, five repetitions at 50 % of the estimated 1RM were performed. In the second set, three repetitions at 70 % of the estimated 1RM were performed. After the second set, the subjects rested for 3 min before performing five trials to achieve the 1RM load (i.e., the maximum weight that could be lifted once using the proper technique) with a 3-min interval between the attempts.
During the 1RM test, the EMG signal was recorded from the left leg. Disposable dual Ag/AgCl snap electrodes with a 1-cm diameter and a 2-cm center-to-center spacing (Noraxon, Scottsdale, AZ, USA) were placed on the belly of the vastus medialis (VM) muscle before starting the test. The SENIAM guidelines were followed for skin preparation, electrode placement, and orientation. Electrode positions were marked with small ink tattoos on the skin during the first testing session to ensure that electrode placement over the entire experimental period would be consistent (Hakkinen and Komi 1983). The electromyographic signals were registered with a telemetric EMG system, which had a gain of 1000, a bandwidth (−3 dB) of 10 to 500 Hz, and a common mode rejection ratio >85 dB. The signal was relayed to the computer via a 16-bit A/D converter (Myotrace 400, Noraxon, Scottsdale, AZ, USA). The EMG data were band-pass filtered at 20–400 Hz, and an envelope representing the muscle activation was determined with a moving RMS filter with a window of 50 ms. Both concentric and eccentric phases were included, and only the iEMG signal of the maximal repetition was chosen for subsequent statistical analyses. The period of activation was determined as the period where the signal was above a threshold of 15 % of the maximum activity of that muscle during the trial for at least 100 ms. For each bout of EMG activation, the integrated EMG (iEMG) was calculated (i.e., the area under the EMG versus time curve divided by the period of activation).
Strength training program
The strength training program was focused on strength development of the leg extensors, which have been considered the major muscle group recruited during running (Taipale et al. 2010). In addition to their normal endurance training, the STG performed strength training sessions twice weekly (separated by 72 h) during 8 weeks. The training sessions were composed by four exercises (Table 1). The main investigator monitored all training sessions, and careful attention was given to each athlete to ensure appropriated technique. During this 8-week training period, the runners were instructed to maintain their previous endurance training routine and to perform the strength training on different days than the endurance training. The training was prescribed with the intensity between 3 and 10RM according to Table 1. A 3-min resting interval between sets and exercises was used throughout the training period. A standardized warm-up protocol consisting of a 5-min treadmill run at 8 km h−1 followed by light lower limb stretching exercises was performed before each training session. In addition, all participants were instructed to perform the exercises with a natural pacing and were required to use the same athletic shoes during the training sessions.
Statistical analysis
Data normality was assessed using the Shapiro–Wilk test. All variables were normally distributed. The results of descriptive statistics were reported as mean ± SD. The effects of training were tested using a two-way (group × time) analysis of variance (ANOVA). When a significant F value was detected, Tukey’s post hoc test was used to identify significant differences. Comparisons between groups for percentage change (%) found after the completion of the study were also performed with unpaired t test. Significance level was set at P < 0.05. All statistical analyses were performed using a statistical software package (Statistica 8, StataSoft Inc., Tulsa, OK, USA).
Results
Participant characteristics
All subjects in the STG completed over 95 % of the scheduled strength training sessions. Participants did not report any discomfort or any adverse event produced by the ST program. There were no significant differences in percentage body fat before (STG 11.2 ± 1.2 %; CG 11.8 ± 1.7 %) and after the training period (STG 9.5 ± 1.1 %; CG 11.3 ± 1.2 %, P = 0.372–0.558). There were also no significant differences in body mass comparing the groups before (STG 67.5 ± 3.1 kg; CG 71.0 ± 3.8 kg) and after training (STG 67.2 ± 2.7 kg; CG 74.7 ± 3.8 kg, P = 0.479–0.435). In addition, no statistical differences were observed in the running training volume expressed as the mean weekly distance covered before (STG 38.6 ± 4.4 km week−1, CG 36.3 ± 7.7 km week−1) and after (STG 41.3 ± 9.1 km week−1, CG 37.2 ± 4.2 km week−1) the completion of the study (P = 0.288–0.982).
Parameters related to endurance performance
There were no significant differences between the groups for baseline values of the parameters related to endurance performance (Table 2). After the training period, there were no significant changes in the \( {\dot{\text{V}}} \)O2max, RCP, or RE for either group (P = 0.198–0.895). However, there was a significant difference between groups for percentage change in PTS, with the STG having a greater percentage change compared to CG (P = 0.013).
Neuromuscular characteristics and anaerobic performance
Table 3 shows the neuromuscular characteristics and anaerobic performance before and after the 8-week strength training period, whereas Fig. 2 shows the percentage change for neuromuscular characteristics within the groups after strength training. No significant group differences were observed in the baseline values (P = 0.258–0.980). The ANOVA revealed a significant group x training effect interaction for 1RM (P = 0.003). The 1RM increased in STG, but did not change in CG. The increased 1RM was accompanied by a tendency to increase the percentage change in iEMG (P = 0.080). In addition, there were a significant higher percentage change for 1RM (P = 0.001), DJH (P = 0.039), CT (P = 0.048), and RSI (P = 0.004) in the STG than in CG. No statistical differences were observed between the other neuromuscular and anaerobic parameters obtained before and after training for both groups (P = 0.153–0.950).

Percentage changes from pre- to post-intervention in the strength training group and the control group. *P < 0.05, between-group differences. 1RM maximum dynamic strength, iEMG surface electromyography of the vastus medialis, DJH drop jump height, CT contact time, RSI reactive strength index
10-km running time trial
Before the training period, there were no significant differences between groups in the overall 10-km running performance (STG = 2626 ± 307 s; CG = 2514 ± 226 s; P = 0.393). After the training period, the STG improved their performance time by 2.5 %, which was higher than that found for CG (−0.7 %) (P = 0.039). Figure 3 shows the running speed and RPE before and after training for STG (panels A and B) and CG (panels C and D), while the panels E and F show, respectively, the percentage change in running speed and in RPE in both groups. It was observed that in the last seven sections of the 10-km running performance, the percentage change was significantly higher for STG than CG (P = 0.006–0.044). In turn, no statistical differences were observed between the RPE response before and after training for both groups (P = 0.203–0.780).

Impact of 8-week strength training on the pacing strategy and rating of perceived exertion during a 10-km running time trial. Data are presented as mean ± SEM. STG strength training group (n = 9), CG control group (n = 9), RPE rating of perceived exertion. *Significantly different from control group (P < 0.05)
Discussion
The main objective of this study was to ascertain whether a strength training program performed during 8 weeks would alter the pacing selected by long-distance runners during a 10-km time trial. Our principal findings corroborated our initial hypothesis, revealing that the ST program produced meaningful improvements in neuromuscular characteristics, which were accompanied by a faster end-spurt for the same RPE. These changes also resulted in an improved overall 10-km performance for STG compared with CG. While the influence of a strength training on endurance performance has already been reported (Bertuzzi et al. 2013, Mikkola et al. 2011), to the best of our knowledge, this is the first study to demonstrate that a ST program was able to influence self-paced endurance running, contributing to an improved overall performance through a more elevated speed during the middle-to-last phases of a 10-km running.
The ability of the skeletal muscles to produce force has been related to both neural and metabolic factors (Aagaard and Mayer 2007). In this respect, previous studies have used vertical jump (Mikkola et al. 2011), all-out exercises (Rønnestad et al. 2014), iEMG (Mikkola et al. 2007), and 1RM tests (Bertuzzi et al. 2014) to assess the neuromuscular and anaerobic adaptations provided by strength training in long-distance athletes. An improved ability to produce maximal and explosive strength would be expected when participants who have no prior strength training experience complete a 4- to 8-week ST program. In particular, strength training may enhance the endurance performance by at least two distinct mechanisms. First, the higher motor unit synchronization after a training period might result in strength potentiation, which improves efficiency, coordination, and delays the onset of signals related to fatigue (Creer et al. 2004). As a result, the runners would be able to produce higher running speeds due to an improved ability to rapidly absorb and create force against the ground (Beattie et al. 2014). In our study, we found an improvement of 26.7 % for 1RM, 47 % for iEMG, 10.3 % for drop jump height, 12.5 % for contact time measured during the drop jump test, and 24 % for reactive strength index in the STG. These findings are in agreement with previous studies, which found a significant increase in 1RM (~18.0 %), iEMG (16.2 %), and jump height (6.1 %) in a similar group of athletes after 6–8 weeks of strength training (Bertuzzi et al. 2014; Mikkola et al. 2011; Taipale et al. 2010). Second, an increase in maximal strength provokes a subsequent lower relative force generated per stride, which would result in a higher recruitment of type I fibers and delayed recruitment of type II fibers, thereby delaying fatigue (Tanaka and Swensen 1998). This adaptation could allow runners to maintain a relatively higher speed over the last third of the race due to a higher ATP availability via aerobic metabolism (Tanaka and Swensen 1998). Based on these findings, it is possible that runners of the STG were able to increase the speed during the last part of a 10-km running time trial after the training period due to changes in the neuromuscular system related to motor unit synchronization and recruitment, increasing the ability to produce force (Mikkola et al. 2011; Taipale et al. 2010).
On the other hand, no statistical differences were detected for anaerobic performance in both STG and CG after the training period. This finding differs from other studies that have reported improvement in the anaerobic performance in long-distance runners after 8–9 weeks of ST program (Mikkola et al. 2011, Paavolainen et al. 1999). These discrepancies among the studies might be partly explained by differences in training regimens. The training protocol used in the current study had higher intensity and lower volume than most of the previous studies conducted with long-distance runners (Paavolainen et al. 1999; Taipale et al. 2010). As a result, our protocol could have produced a lower stress in the muscle energy systems and muscle lactate production, which did not improve the ability of the anaerobic lactic metabolism to resynthesize ATP. This is in accordance with previous findings indicating that strength training increases the anaerobic capacity in athletes who traditionally use training regimens with higher volume than that used in the present study (Pizza et al. 1996). Furthermore, it has been reported that increases in muscle strength after training at higher loads (4–6RM), such as those used in the last weeks of our training program (last 4 weeks), could be attributed to neural adaptations including enhanced motor unit activation, faster firing frequency of motor units, improved synchronization, and co-contraction of antagonists (Docherty and Sporer 2000). Thus, these findings could suggest that the characteristics of our training protocol may have led to greater adaptations in neuromuscular characteristics than anaerobic metabolism.
Our results revealed that the average 2.5 % improvement in the time to cover the 10-km running time trial observed in the STG appears to be the result of an enhanced running speed in the last third of the race. Interestingly, this increase in running speed was not accompanied by an alteration in the RPE, indicating that athletes were able to maintain higher speeds with similar perception of effort after a strength training period. Previous studies proposed that the RPE is generated as a result of the numerous afferent signals during exercise and acts as a mediator of subsequent alterations in skeletal muscle activation (De Morree et al. 2012). In this scenario, the RPE represents the integration of the alterations in physiological systems during dynamic exercise and is considered a primary regulator of pacing strategy (Tucker and Noakes 2009). It therefore appears that the improvements in the neuromuscular characteristics induced by strength training may have allowed the athletes to exercise at a greater intensity for the same level of perceived exertion. These findings support the notion that neuromuscular mechanisms related to peripheral fatigue are some of the possible variables utilized by central nervous system to regulate exercise intensity, particularly during the final phase of a running time trial.
With respect to parameters related to endurance performance, our results are consistent with previous studies that observed no alterations for \( {\dot{\text{V}}} \)O2max and RCP in long-distance athletes participating in a ST program (Bertuzzi et al. 2014; Mikkola et al. 2011; Storen et al. 2008). It appears that changes in these physiological variables are dependent on initial aerobic fitness. Previous studies that observed changes in \( {\dot{\text{V}}} \)O2max and RCP after strength training utilized sedentary or aerobically unfit subjects (\( {\dot{\text{V}}} \)O2max < 40 ml kg−1 min−1) (Gettman et al. 1978). It is believed that an acute bout of strength training requires oxygen consumption values of less than 50 % of maximal capacity (Hurley et al. 1984). This is unlikely to provide a strong stimulus for improving aerobic capacity, since endurance training, which results in improvements of aerobic fitness in trained runners, is typically performed at intensities requiring at least 70–85 % of \( {\dot{\text{V}}} \)O2max (Billat et al. 2004). However, it is interesting that we observed a significant increase in the percentage change in PTS for the STG after training program. It has been proposed that the PTS is influenced not only by maximal aerobic power, but also by the RE, anaerobic performance, and neuromuscular characteristics of the subjects (Mikkola et al. 2011; Noakes 1988). In the current study, the ST regimen did not change the RE and anaerobic performance, but neuromuscular parameters were improved (i.e., 1RM, DJ, CT, and RSI). Therefore, the results of the present study support the concept that a slight increase in PTS induced by the ST program was caused by changes in the neuromuscular system, increasing the ability to produce force.
It is necessary to acknowledge some of the limitations of the present study. First, it is important to note that our sample was composed of moderately trained athletes. Furthermore, participants had no previous strength or plyometric training experience. These athletes may be more responsive to strength training than highly trained runners. Thus, caution should be exercised in extrapolating these findings to highly trained athletes who frequently perform training sessions using exercises that are strength- or power-oriented. Second, the training protocol was limited to four exercises, which may have resulted in a lower strength training volume than those often used in the “real world.” Thus, future trials should compare the efficacy of various exercise modalities, intensities, and duration to determine which training regimens are most effective to alter the pacing strategy selected by long-distance athletes during running time trials.
In conclusion, the results of the present study showed that 8 weeks of strength training improved the neuromuscular characteristics of endurance runners, resulting in a faster and more-sustained end-spurt during a 10-km running time trial. These findings suggest that the neuromuscular characteristics could be an important determinant of exercise intensity during the middle-to-last phases of a 10-km running. In addition, it is possible to suggest that the strength gain after a ST program may contribute to reduced peripheral fatigue in long-distance athletes during the final phases of an endurance race, resulting in an improved overall running performance.
Abbreviations
- 1RM:
-
Maximum dynamic strength
- CG:
-
Control group
- CT:
-
Contact time
- iEMG:
-
Electromyographic activity
- DJH:
-
Drop jump height
- MP:
-
Mean power
- PP:
-
Peak power
- PTS:
-
Peak treadmill speed
- RCP:
-
Respiratory compensation point
- RE:
-
Running economy
- RPE:
-
Rate of perceived exertion
- RSI:
-
Reactive strength index
- STG:
-
Strength training group
- ST:
-
Strength training
- \( {\dot{\text{V}}} \)O2max:
-
Maximal oxygen uptake
- VM:
-
Vastus medialis
References
Aagaard P, Mayer F (2007) Neuronal adaptations to strength training. Deutsche Zeit Sport med 58(2):50–53
Abbiss CR, Laursen PB (2008) Describing and understanding pacing strategies during athletic competition. Sports Med 38(3):239–252
Bar-Or O (1987) The Wingate anaerobic test: an update on methodology, reliability and validity. Sports Med 50:273–282
Beattie K, Kenny IC, Lyons M, Carson BP (2014) The effect of strength training on performance in endurance athletes. Sports Med 44(6):845–865
Bertuzzi R, Pasqua LA, Bueno S, Damasceno MV, Lima-Silva AE, Bishop D, Tricoli V (2013) Strength-training with whole-body vibration in long-distance runners: a randomized trial. Int J Sports Med 34:917–923
Bertuzzi R, Lima-Silva AE, Pires FO, Damasceno MV, Bueno S, Pasqua LA, Bishop DJ (2014) Pacing strategy determinants during a 10-km running time trial: contributions of perceived effort, physiological, and muscular parameters. J Strength Cond Res 28(6):1688–1696
Billat VL, Sirvent P, Lepretre PM, Koralsztein JP (2004) Training effect on performance, substrate balance and blood lactate concentration at maximal lactate steady state in master endurance runners. Pflugers Arch 447(6):875–883
Borg GA (1982) Psychophysical bases of perceived exertion. Med Sci Sports Exerc 14(5):377–381
Brown LE, Weir JP (2001) Procedures recommendation I: accurate assessment of muscular strength and power. J Exerc Physiol 4:1–21
Brozek J, Grande F, Anderson JT, Keys A (1963) Densitometric analysis o body composition: revision of some quantitative assumptions. Ann NY Acad Sci 26(110):113–140
Creer AR, Ricard MD, Conlee RK, Hoyt GL, Parcell AC (2004) Neural, metabolic, and performance adaptations to four weeks of high-intensity sprint-interval training in trained cyclists. Int J Sports Med 25:92–98
De Morree HM, Klein C, Marcora SM (2012) Perception of effort reflects central motor command during movement execution. Psychophysiology 49:1242–1253
Docherty D, Sporer A (2000) Proposed model for examining the interference phenomenon between concurrent aerobic and strength training. Sports Med 30(6):385–394
Faulkner J, Parfitt G, Eston R (2008) The rating of perceived exertion during competitive running scales with time. Psychophysiology 45:977–985
Gettman LR, Ayres JJ, Pollock ML, Jackson A (1978) The effect of circuit weight training on strength, cardiorespiratory function, and body composition of adult men. Med Sci Sports Exerc 10(3):171–176
Hakkinen K, Komi PV (1983) Electromyographic changes during strength training and detraining. Med Sci Sports Exerc 15(6):455–460
Howley ET, Bassett DR Jr, Welch HG (1995) Criteria for maximal oxygen uptake: review and commentary. Med Sci Sports Exerc 27:1292–1301
Hurley BF, Seals DR, Seals DR, Ehsani AA, Cartier LJ, Dalsky GP, Hagberg JM, Holloszy JO (1984) Effects of high-intensity strength training on cardiovascular function. Med Sci Sports Exerc 16(5):483–488
Jackson AS, Pollock ML (1985) Practical assessment of body composition. Phys Sports Med 19:76–90
Lambert EV, St Clair Gibson A, Noakes TD (2005) Complex systems model of fatigue: integrative homoeostatic control of peripheral physiological systems during exercise in humans. Br J Sports Med 39:52–62
Lima-Silva AE, Bertuzzi RCM, Pires FO, Barros RV, Gagliardi JF, Hammond J, Kiss MA, Bishop DJ (2010) Effect of performance level on pacing strategy during a 10-km running race. Eur J Appl Physiol 108(5):1045–1053
Meyer T, Lucía A, Earnest CP, Kindermann W (2005) A conceptual framework for performance diagnosis and training prescription from submaximal gas exchange parameters—theory and application. Int J Sports Med 26:S38–S48
Mikkola J, Rusko H, Nummela A, Pollari T, Häkkinen K (2007) Concurrent endurance and explosive type strength training improves neuromuscular and anaerobic characteristics in young distance runners. Int J Sports Med 28:602–611
Mikkola J, Vesterinen V, Taipale R, Capostagno B, Häkkinen K, Nummela A (2011) Effect of resistance training regimens on treadmill running and neuromuscular performance in recreational endurance runners. J Sports Sci 29:1359–1371
Noakes TD (1988) Implications of exercise testing for prediction of athletic performance: a contemporary perspective. Med Sci Sports Exerc 20:319–330
Norton K, Olds T (1996) Measurement technique in anthropometry. In: Norton K, Olds T (eds) Anthropometrica. University of New South Wale Press, Sydney, pp 25–75
Nummela A, Paavolainen LM, Sharwood KA, Lambert MI, Noakes TD, Rusko HK (2006) Neuromuscular factors determining 5 km running performance and running economy in well-trained athletes. Eur J Appl Physiol 97:1–8
Paavolainen L, Hakkinen K, Hamalainen I, Nummela A, Rusko H (1999) Explosive-strength training improves 5-km running time by improving running economy and muscle power. J Appl Physiol 86(5):1527–1533
Pizza FX, Naglieri TA, Holtz RW, Mitchell JB, Starling RD, Phillips MD, Cavender DL, Braun WA (1996) Maximal accumulated oxygen deficit of resistance-trained men. Can J Appl Physiol 21(5):391–402
Rønnestad BR, Hansen J, Hollan I, Ellefsen S (2014) Strength training improves performance and pedaling characteristics in elite cyclists. Scand J Med Sci Sports 25(1):e89–e98
Schulz KF, Altman DG, Moher D (2010) CONSORT 2010 Statement: updated guidelines for reporting parallel group randomised trials. PLoS Med 7(3):e1000251
St Clair Gibson A, Lambert EV, Rauch LH, Tucker R, Baden DA, Foster C, Noakes TD (2006) The role of information processing between the brain and peripheral physiological systems in pacing and perception of effort. Sports Med 36(8):705–722
Storen O, Helgerud J, Stoa EM, Hoff J (2008) Maximal strength training improves running economy in distance runners. Med Sci Sports Exerc 40:1087–1092
Taipale RS, Mikkola J, Nummela A, Vesterinen V, Capostagno B, Walker S, Gitonga D, Kraemer WJ, Häkkinen K (2010) Strength training in endurance runners. Int J Sports Med 31:468–476
Tanaka H, Swensen T (1998) Impact of resistance training on endurance performance. A new form of cross-training? Sports Med 25(3):191–200
Thiel C, Foster C, Banzer W, De Koning J (2012) Pacing in Olympic track races: competitive tactics versus best performance strategy. J Sports Sci 30(11):1107–1115
Tucker R, Noakes TD (2009) The anticipatory regulation of performance: the physiological basis for pacing strategies and the development of a perception-based model for exercise performance. Br J Sports Med 43(6):392–400
Ulmer HV (1996) Concept of an extracellular regulation of muscular metabolic rate during heavy exercise in humans by psychophysiological feedback. Experientia 52(5):416–420
Young WB, Behm DG (2003) Effects of running, static stretching and practice jumps on explosive force production and jumping performance. J Sports Med Phys Fitness 43:21–27
Acknowledgments
The authors thank each of the individuals for their participation. The study was supported by grant from São Paulo Research Foundation (FAPESP 2011/10742-9). Mayara Vieira Damasceno is supported by a master scholarship from São Paulo Research Foundation (FAPESP 2013/00371-9).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Guido Ferretti.
Rights and permissions
About this article

Cite this article
Damasceno, M.V., Lima-Silva, A.E., Pasqua, L.A. et al. Effects of resistance training on neuromuscular characteristics and pacing during 10-km running time trial. Eur J Appl Physiol 115, 1513–1522 (2015). https://doi.org/10.1007/s00421-015-3130-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-015-3130-z