
Abstract
Many temperate ectotherms undergo thermal acclimation to remain functional over a wide range of body temperatures, but few studies have investigated whether populations of a single species have evolved differences in the thermal plasticity of locomotor performance. Therefore, we asked whether the thermal plasticity of locomotor performance has diverged between northern and southern populations of eastern newts (Notophthalmus viridescens). We acclimated eastern newts from Florida and Maine to cold (6 °C) or warm (28 °C) conditions for 12 weeks. Following acclimation, we measured the burst speed of newts at 6, 11.5, 17, 22.5, 28, and 33.5 °C. We also measured the activities of creatine kinase (CK) and lactate dehydrogenase (LDH) in skeletal muscle of newts. The newts from Maine were better able to acclimate to low temperature compared to newts from Florida. Regardless of acclimation, the thermal sensitivity of burst speed was higher in the Florida compared to the Maine population. In general, newts from Maine performed better at low temperatures, whereas newts from Florida performed better at high temperatures. The activities of CK and LDH were lower in cold compared to warm-acclimated newts in the Florida population, but acclimation did not affect the activities of these enzymes in the Maine population. The activities of CK and LDH do not explain differences in the thermal plasticity of locomotor performance between populations. Our results demonstrate that the thermal sensitivity and plasticity of locomotor performance differ between northern and southern populations of eastern newts, suggesting that these traits readily adapt to the thermal environment.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The rates of enzyme-mediated reactions depend on temperature. As body temperature (T b) increases, the rates of physiological processes (e.g., metabolism, growth, and locomotion) increase before peaking at a thermal optimum. However, these rate processes decrease rapidly if T b rises above the thermal optimum (Huey and Stevenson 1979). Consequently, the physiological performance of ectotherms fluctuates with environmental temperature (T a), unless behavior prevents T b from changing, or physiological mechanisms buffer the effects of temperature on organismal function (Hochachka and Somero 2002). Given that temperature affects traits directly related to fitness, it is not surprising that the thermal environment exerts strong selection on the thermal sensitivity of physiological performance in ectotherms (Huey and Stevenson 1979).
Despite the effects of temperature on fitness, ectotherms are found in diverse thermal environments across the globe. Different species, or different populations of the same species, adapt to local conditions over time so that performance is matched to the range of T bs experienced by the organism. At the cellular level, modifications in the catalytic efficiencies of metabolic enzymes and/or changes in the mitochondrial content of the tissues can counteract the effects of temperature on metabolism (Hochachka and Somero 2002). For example, the skeletal muscles of fish living at low temperatures have more mitochondria than those of species living at high temperatures (Guderley 2004). Additionally, the thermal sensitivity of sprint speed in several species of Anolis lizards matches the body temperatures that these lizards experience in the field (Van Berkum 1986), and species of tree frogs from high latitudes jump further at low temperatures compared to species from low latitudes (Alder-John et al. 1988). Whereas interspecific comparisons have demonstrated that species adapt to the thermal environment, earlier studies comparing closely related species, or populations of the same species, suggested that thermal adaptation is constrained within taxa. For instance, the thermal sensitivity of locomotion does not differ between populations of tree frogs from high and low latitudes (Alder-John et al. 1989) or between agamid lizards living in different thermal environments (Hertz et al. 1983). However, recent reports show that jumping performance at low temperatures is higher in populations of striped marsh frogs from cold climates compared to those from warm climates, whereas populations from warm climates perform better at high temperatures than those from cold climates (Wilson 2001). Furthermore, Anolis lizards introduced to Florida tolerate lower temperatures than native populations from Puerto Rico (Gunderson and Leal 2012). Thus, these data suggest that the thermal physiology of ectotherms evolves more readily than previously thought.
Many eurythermal ectotherms in temperate climates rely on phenotypic plasticity to contend with seasonal fluctuations in T b during their lifetime. Thermal acclimatization occurs in response to predictable cues (e.g., photoperiod or temperature) and is characterized by biochemical modifications that compensate for thermodynamic effects on physiological performance. Thermal acclimation often results in changes in the oxidative capacity of muscle tissue (Berner and Bessay 2006; Berner and Puckett 2010; Guderley 2004). As a result, metabolic capacity is maintained despite seasonal fluctuations in T b. Thermal acclimation also affects locomotor performance, as animals acclimated to a particular temperature perform better at that temperature compared to animals acclimated to other temperatures (Glanville and Seebacher 2006; Gvozdik et al. 2007; Wilson and Franklin 1999; Wilson et al. 2000).
In species with broad geographic distributions, different thermal environments may cause thermal plasticity to diverge among populations (Seebacher 2005). If thermal acclimatization is beneficial in environments where temperature fluctuates on a seasonal basis, species or populations from environments with greater seasonal temperature variation may have a greater capacity for thermal acclimatization compared to those from environments with less seasonal variation. Furthermore, populations that experience differences in minimum and maximum temperatures may differ in their capacity to acclimatize to low or high temperatures, respectively. In general, tropical ectotherms lack the capacity for thermal acclimation compared to temperate species (Feder and Lynch 1982; Rogowitz 1996). As interspecific comparisons may be confounded by phylogenetic differences, intraspecific comparisons are better suited for determining whether the thermal environment affects the capacity for thermal acclimatization. Few studies have investigated within-species differences in thermal plasticity in ectotherms. However, evidence for intraspecific variation in thermal plasticity was shown in mosquito fish, although variation between populations was not explained by environmental conditions (Seebacher et al. 2012). Also, cold acclimation increases the aerobic capacity of populations of killifish from high latitudes to a greater extent than those from low latitudes (Dhillon and Schulte 2011; Fangue et al. 2009), suggesting that thermal plasticity is labile. Because locomotion is a predictor of fitness (Husak et al. 2006), selection is expected to favor thermal plasticity of locomotor performance in variable thermal environments. However, studies comparing within-species differences in the thermal plasticity of locomotor performance are needed to elucidate how the thermal environment influences this trait.
The geographic distribution of the eastern newt (Notophthalmus viridescens) ranges from Canada (49°N) south to Florida (25°N). Adults of this species are aquatic, and thermal acclimation of metabolic rate and metabolic enzymes in muscle (Berner and Bessay 2006; Berner and Puckett 2010) appear to be partly responsible for allowing these animals to remain active throughout the year. Surface temperatures of ponds in Florida are ~15 °C in winter and can exceed 30 °C in summer (Escobar et al. 2009; Kushlan 1979; waterdata.usgs.gov). In Maine, the ponds freeze over in the winter and rarely rise above ~25 °C in the summer (waterdata.usgs.gov). Since northern and southern populations experience differences in the variability of seasonal temperatures, as well as differences in minimum and maximum temperatures, these animals allow us to investigate whether the plasticity of performance has diverged among populations from different thermal environments. Therefore, we compared the thermal plasticity of locomotor performance and the biochemical properties of skeletal muscle between populations of eastern newts from northern (Maine) and southern (Florida) latitudes.
Materials and methods
In spring 2012, eastern newts were collected in Florida (Marion and Putnam counties) and Maine (Brunswick County) and transported to the animal facility at Miami University. Newts were held at 17 °C (12:12 LD) for a minimum of 6 weeks in order to habituate to captivity. The newts were housed in groups of 8–10 individuals per 76-L aquarium located inside a temperature-controlled environmental chamber (Thermolinear, Cincinnati, OH, USA). The aquaria contained dechlorinated water at a depth of 15 cm. Each aquarium was equipped with a water filter (ReptoFilter, Tetrafauna, Blacksburg, VA, USA) and contained rocks and plastic plants for refuge. The newts were fed bloodworms ad libitum (Sally’s, San Francisco Bay Brand, Newark, CA, USA), and the water in the aquaria was changed every other day.
Each population was separated into one of two acclimation regimes: cold (6 °C, 10:14 LD) or warm (28 °C, 14:10 LD). The temperature was changed from 17 °C to the appropriate acclimation temperature over a period of 2 weeks. The newts were held at their acclimation temperature for 12 weeks. After 12 weeks, a subset of these animals was sacrificed by cervical severing, and trunk and tail muscle was collected and immediately frozen in liquid nitrogen. The muscle tissue was stored at −80 °C for enzyme analysis. The rest of the animals were used for locomotion experiments (as described below). Following the completion of the locomotor performance experiments, we measured the body mass, snout-vent length, and vent-tail length of each newt before sacrifice. Permission to collect eastern newts was granted by the Florida Fish and Wildlife Conservation Commission (permit no LSSC-12-00018) and the Maine Department of Inland Fisheries and Wildlife (permit no 2012-365). All animal experimentation was approved by the Institutional Animal Care and Use Committee of Miami University (protocol no 830) and complied with the “Principles of Animal Care,” publication no. 86–23, revised 1985, of the National Institutes of Health as well as the laws of the USA.
Locomotor performance assays
We measured the burst swimming performance of cold- and warm-acclimated newts from Maine (cold acclimated, n = 20; warm acclimated, n = 19) and Florida (cold acclimated, n = 18; warm acclimated, n = 21) at 6, 11.5, 17, 22.5, 28, and 33.5 °C in a 70 cm × 10 cm × 5 cm plexiglass track. The track was filled with 3 cm of water to limit the newts from swimming vertically. The bottom of the track was marked at 1 cm increments. The track was equipped with a water jacket that connected to a temperature-controlled circulating water bath (model F12, Julabo, Allentown, PA, USA). Prior to the burst swimming trials, the body temperature of the newts was changed to the experimental temperature at a rate of 5 °C/h. To change body temperature, newts were placed in small plastic containers filled with 1 cm of water, and the water in the containers was changed to the experimental temperature in a biological incubator (model I-30VL, Percival, Perry, IA, USA). All animals remained at the test temperature for 30 min prior to the experiments. Locomotor performance experiments were performed in an environmental chamber set to within ±1 °C of the desired test temperature. Burst swimming trials were performed at one test temperature per day, with 1 day in between test temperatures.
Individuals were prompted to swim by touching the tail with a small paintbrush, and the ensuing escape response was filmed (30 frames/s) with a video camera (Sony DCR-SX65) positioned 90° above the track. Because newts do not achieve maximal escape responses every trial (previous experiments indicate that the standard error of the three responses ranges from 1 to 20 % of the mean of the responses across the experiment), three escape responses were filmed for each individual. The time required to swim 10 cm during the escape response was calculated by Tracker Video Motion Analysis and Modeling Tool software (Open Source Physics, www.opensourcephysics.org) and used to calculate the burst swimming speed of each individual. We used the highest burst swimming speeds from the three escape responses filmed. Individuals that failed to swim were omitted from the analysis (2 individuals from the warm-acclimated Florida group at 6 °C, 1 individual from the cold-acclimated Maine group at 11.5 °C, 2 individuals from the warm-acclimated Maine group at 22.5 °C, 3 individuals from the warm-acclimated Maine group at 28 °C, 1 individual from the warm-acclimated Florida group at 33.5 °C, and 3 individuals from the cold-acclimated Florida group at 33.5 °C). Burst swimming speed was measured in the order of 17, 22.5, 6, 33.5, 11.5, and 28 °C. To demonstrate that our results are repeatable and that these experiments did not influence locomotor performance, we re-tested the burst swimming speed of each individual at 17 °C (Wilson et al. 2000). There was no difference between the initial and final measurements of burst speed at 17 °C for either population or acclimation group. To determine the thermal sensitivity of burst speed, we calculated temperature coefficients (Q 10 values) for the burst speed of each individual as: Q 10 = (R 2/R 1)10/(T2–T1), where R 2 is the burst speed at the higher temperature (T 2), and R 1 is the burst speed at the lower temperature (T 1).
Enzyme assays
The activities of lactate dehydrogenase (LDH) and creatine kinase (CK) in skeletal muscle of cold and warm-acclimated Florida and Maine newts were measured at 6, 17, and 28 °C. Tissue homogenates were prepared following the methods of Berner and Puckett (2010). Trunk and tail muscle were weighed and homogenized at 4 °C in nine volumes of homogenization buffer (50 mM imidazole, 2 mM MgCl2, 5 mM EDTA, 1 mM glutathione, and 0.1 % Triton X 100, pH = 7.5). Homogenized tissue was centrifuged at 300g, and the resultant supernatant was collected for enzyme analysis.
Enzyme activities were calculated from the change in absorbance using a temperature-controlled spectrophotometer (Lambda 35, PerkinElmer, Waltham, MA, USA). LDH activity was measured at 340 nm in assay medium containing 100 mM potassium phosphate (pH = 7.0), 0.16 mM NADH, and 0.4 mM pyruvate. The conditions for the CK activity assay were adapted from Dalzeil et al. (2012) and optimized for newt muscle. CK activity was measured at 340 nm in reaction medium containing 100 mM Tris–HCl (pH = 7.4), 25 mM PCr, 3 mM ADP, 5 mM AMP, 1 mM NADP, 5 mM glucose, 2 U/ml hexokinase, 1 U/ml G6PDH, 5 mM MgCl2, and 3.25 mM DTT. All assays were performed in duplicate. Enzyme activity is expressed as units g−1 of tissue, where one unit is equal to 1 µmol of substrate modified per minute.
Statistical analysis
The effects of population and acclimation on burst speed were tested with a mixed model ANOVA (Verbeke and Molenberghs 2009), with sex, temperature, acclimation condition, and population as fixed effects, and individual as a random effect. Body mass and snout-vent length did not differ between populations or treatment groups, but the Florida population had longer tails than the Maine population (Table 1; F 1,1 = 9.8, P < 0.01), and within each population, the warm-acclimated newts had longer tails compared to the cold-acclimated newts (F 1,1 = 5.0, P < 0.05). Therefore, vent-tail length was also included in the model. Bonferroni-adjusted tests were used to compare mean burst speeds between populations and acclimation conditions. For the enzyme experiments, the effects of population and acclimation on the activities of CK and LDH were tested with a mixed model ANOVA, with temperature, acclimation, and population as fixed effects, and individual as a random effect. In the case of a population × acclimation interaction, the effects of acclimation on the activities of CK and LDH were analyzed separately for each population using a mixed effect model, with temperature and acclimation as fixed effects, and individual as a random effect. The effects of population and acclimation conditions on body mass, snout-vent length, vent-tail length, and Q 10 values were tested using a factorial ANOVA. Data that did not meet parametric assumptions of normality and equal variance were log-transformed prior to the analysis. All statistical tests were performed in JMP Pro (version 10.0). Data are presented as mean ± SEM. Significance was determined whether P < 0.05.
Results
Locomotor performance
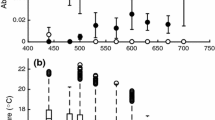
There was no effect of vent-tail length or sex on the burst speed of eastern newts. Thermal acclimation affected the thermal sensitivity of burst speed (Fig. 1; temperature × acclimation interaction, F 5,351 = 48.7, P < 0.0001) of both populations (temperature × acclimation × population interaction, F 5,350 = 2.1, P = 0.07). In both populations, Q 10 values for burst speed were higher in warm-acclimated newts compared to cold-acclimated newts (Table 2; F 1,1 = 122.9, P < 0.001). The effect of thermal acclimation on burst speed differed between populations (Fig. 1; population × acclimation interaction, F 1,70 = 4.4, P < 0.05). In the Maine population, the burst speed of the cold-acclimated newts was 66 % higher than that of the warm-acclimated newts at 6 °C (t = 7.6, P < 0.05) and 37 % higher than that of the warm-acclimated newts at 11.5 °C (t = 5.4, P < 0.05). In the Florida population, the burst speed of the cold-acclimated newts was 30 % higher than the warm-acclimated newts at 6 °C (t = 4.7, P < 0.0001). Within the cold-acclimated newts, the Maine population performed better at low temperatures compared to the Florida population, as the burst swimming speed of the cold-acclimated Maine newts was higher than the cold-acclimated Florida newts at 6 °C (t = 6.4, P < 0.05) and 11.5 °C (t = 4.5, P < 0.05).

Effect of temperature on the burst speed of eastern news acclimated to 6 °C (Maine, n = 20; Florida, n = 18) or 28 °C (Maine, n = 19; Florida, n = 21) for 12 weeks. *Significant differences between acclimation groups in the Maine population. $Significant difference between acclimation groups in the Florida population. †Significant difference between cold-acclimated Maine and Florida newts. #Significant difference between warm-acclimated Maine and Florida newts. Data are presented as mean ± SEM. Significance was determined whether P < 0.05
Thermal acclimation also affected the locomotor performance of the newts at high temperatures. In the Maine population, the burst speed of the warm-acclimated newts was 33 % higher than that of the cold-acclimated newts at 28 °C (t = 4.3, P < 0.05) and 47 % higher than that of the cold-acclimated newts at 33.5 °C (t = 4.0, P < 0.05). In the Florida population, burst speed did not differ between warm- and cold-acclimated newts at 25 °C, but the burst speed of the warm-acclimated newts was 60 % higher than that of the cold-acclimated newts at 33.5 °C (t = 5.9, P < 0.05). At high temperature, the Florida population performed better than the Maine population, as the warm-acclimated Florida newts had a higher burst speed than the warm-acclimated Maine newts at 33.5 °C (t = 3.5, P < 0.05).
The thermal sensitivity of burst speed also differed between populations (Fig. 1; population × temperature interaction, F 5,350 = 14.2, P < 0.0001). To determine how the thermal sensitivity of locomotor performance differed between populations, we calculated Q 10 values of burst speed (Table 1). When calculated over the whole temperature range (6–33.5 °C), Q 10 values were higher in the Florida population than in the Maine population when compared at similar acclimation temperatures (F 1,1 = 36.7, P < 0.0001). At 6–17 °C, Q 10 values were higher in the Florida population compared to the Maine population (F 1,1 = 29.5, P < 0.0001), but Q 10 values were not affected by population when calculated over other temperature ranges.
Enzyme activities
The effect of thermal acclimation on LDH activity differed between populations (Fig. 2; population × acclimation interaction, F 1,19 = 7.8, P < 0.05). Therefore, the effect of acclimation on LDH activity was analyzed separately for each population. Whereas acclimation had no affect on LDH activity in the Maine population, LDH activity was higher in warm-acclimated Florida newts compared to the cold-acclimated Florida newts (F 1,10 = 14.9, P < 0.01). Because the effect of acclimation on CK activity also differed between populations (Fig. 3; population × acclimation interaction, F 1,19 = 10.4, P < 0.05), the effect of acclimation on CK activity was analyzed separately for each population. Acclimation did not affect CK activity in the Maine population, but CK activity was higher in the warm compared to cold-acclimated Florida newts (F 1,10 = 5.3, P < 0.05).

Effect of temperature on the LDH activity of eastern newts acclimated to either 6 °C (Maine, n = 6; Florida, n = 6) or 28 °C (Maine, n = 5; Florida, n = 6) for 12 weeks. Data are presented as mean ± SEM. Significance was determined whether P < 0.05

Effect of temperature on the CK activity of eastern newts acclimated to either 6 °C (Maine, n = 6; Florida, n = 6) or 28 °C (Maine, n = 5; Florida, n = 6) for 12 weeks. Data are presented as mean ± SEM. Significance was determined whether P < 0.05
Discussion
Our results demonstrate that the thermal plasticity of locomotor performance differs between northern and southern populations of eastern newts (N. viridescens). Cold acclimation benefits locomotion at low temperatures in both populations, but the Maine population is better able to acclimate to low temperatures. Regardless of thermal plasticity, the thermal sensitivity of locomotor performance is higher in the Florida population. As a result of these differences in thermal plasticity and thermal sensitivity, the Maine population performs better at low temperatures, whereas the Florida newts perform better at high temperatures. These results indicate that the thermal sensitivity and plasticity of locomotor performance can diverge between populations from different thermal environments.
Traditionally, the thermal physiology of amphibians and reptiles has been viewed as conservative, as early studies found geographic variation in the thermal sensitivity of performance among closely related species, and different populations of the same species, to be limited (Hertz et al. 1983; Alder-John et al. 1989). However, the thermal tolerance of Anolis lizards introduced to Florida has diverged in the past 35 years (Gunderson and Leal 2012), suggesting that the thermal physiology of ectotherms can undergo rapid differentiation. Furthermore, the thermal sensitivity of jumping performance has diverged among different populations of striped marsh frogs (Wilson 2001), and our results suggest that the thermal sensitivity of performance has also diverged between northern and southern populations of eastern newts, regardless of thermal plasticity. Geographic variation in the thermal plasticity of aerobic capacity in killifish (Fangue et al. 2009; Dhillon and Schulte 2011) suggests that thermal plasticity may also differentiate among populations, and our study extends these observations by demonstrating that the thermal plasticity of locomotor performance is also labile in an amphibian.
In ectotherms that use behavioral thermoregulation to avoid temperatures that negatively affect performance, the capacity for thermoregulation should impact selection on the thermal sensitivity and plasticity of performance. Previous studies in Anolis lizards indicate that they adapt to low temperature more readily than high temperature, perhaps because these animals can avoid high temperature more effectively than low temperature (Gunderson and Leal 2012; Munoz et al. 2014). However, aquatic environments offer less opportunity for behavioral thermoregulation than terrestrial environments, and selection may be stronger on the thermal sensitivity and thermal plasticity of performance at both low and high temperature in aquatic ectotherms. While we do not have data on how the body temperatures of eastern newts vary seasonally, the results of our study are consistent with the predicted environmental temperatures of these populations. The surface temperatures of ponds in Florida are ~15 °C during winter (waterdata.usgs.gov; Kushlan 1979; Escobar et al. 2009), whereas the ponds in Maine are covered with ice. Therefore, the Maine population experiences near freezing temperatures during winter, necessitating physiological adjustments that compensate for the effects of temperature. In summer, the surface temperatures of ponds in Florida exceed 30 °C (waterdata.usgs.gov; Kushlan 1979; Escobar et al. 2009), but rarely rise above ~25 °C in Maine (waterdata.usgs.gov). In deep lakes, thermal stratification may allow animals to avoid high temperature in summer, but eastern newts are common in shallow bodies of water. Escobar et al. (2009) recorded water temperatures of 27 °C at ~3 m deep in a large Florida pond (17.5 m deep) during the warmest part of the year, and newts were located in areas where water temperature was ~28 °C in April 2012 (Mineo and Schaeffer, unpublished observations). Therefore, the Florida population likely cannot avoid high temperature as effectively as the Maine population, perhaps explaining why performance at high temperature has diverged between populations.
Regardless of acclimation conditions, the difference in the thermal sensitivity of burst speed between the Maine and Florida populations indicates differences between these populations that are not explained by phenotypic plasticity. Due to the interaction between population and thermal acclimation, we cannot completely separate the effects of plasticity and population on locomotor performance. However, Q 10 values indicate that the thermal sensitivity of locomotor performance was higher in the Florida population than that of the Maine population, and this difference between populations was similar across acclimation conditions. Therefore, regardless of plasticity, the locomotor performance of the Florida population is more sensitive to temperature compared to the Maine population. Whereas the high thermal sensitivity negatively affects the performance of the Florida population at low temperatures, the high thermal sensitivity results in better performance at high temperatures relative to the Maine population.
The differences in the thermal sensitivity and thermal plasticity of locomotor performance between Florida and Maine newts could be the result of either genetic adaptation or developmental plasticity. To our knowledge, no studies have investigated how thermal conditions during development influence the thermal plasticity of locomotor performance of adult amphibians, but recent work has shown that temperature experienced during development may affect the capacity for thermal acclimation in fish (Schnurr et al. 2014). Furthermore, the thermal sensitivity of burst speed is affected by temperature during development in lizards (Elphick and Shine 1998; Downes and Shine 1999). The incubation period of eastern newts is ~35 days, and because southern populations of eastern newts oviposit in early winter, and northern populations oviposit from March to July (Petranka 2010), these populations may experience similar temperatures during development. Nonetheless, future studies are needed to determine whether thermal conditions during development affect the thermal plasticity of adult amphibians. Also, because we varied photoperiod and acclimation temperature in order to simulate summer and winter conditions, we cannot eliminate the effect that photoperiod may have had on the performance of newts.
The mechanisms responsible for the differences in the thermal sensitivity and plasticity of locomotor performance between the Maine and Florida populations are not known. The activities of the metabolic enzymes (CK and LDH) were not upregulated in cold-acclimated newts, and the activities of these enzymes do not explain differences in locomotor performance between populations or acclimation groups. Because locomotion is the result of neural, metabolic, and mechanical properties, compensation at any of these levels of organization may be responsible for acclimation of whole-animal performance in ectotherms. As metabolic properties of skeletal muscle do not explain differences in performance, future studies are required to determine whether neural or whole-muscle properties underlie differences in these properties.
The lower activity of CK and LDH in the cold compared to warm-acclimated newts in the Florida population could be due to a shift in the composition of the muscle from an anaerobic to aerobic phenotype. Cold acclimation is associated with upregulation of aerobic enzymes in eastern newts from Tennessee, but the activity of LDH in muscle was not affected by thermal acclimation in those studies (Berner and Bessay 2006; Berner and Puckett 2010). The higher activities of CS and CCO in cold-acclimated newts from Tennessee were not associated with a higher amount of mitochondria in skeletal muscle, suggesting that the other mechanisms, such as changes in membrane composition, are responsible for the higher levels of enzyme activity (Berner et al. 2012). It is possible that cold acclimation results in a shift from a glycolytic to oxidative phenotype in the Florida population, explaining the lower activities of CK and LDH in the cold-acclimated newts from Florida. In contrast, upregulation of aerobic enzymes in the Tennessee population (Berner and Bessay 2006; Berner and Puckett 2010) may be due to other mechanisms, such as changes in membrane composition, allowing LDH activity to be maintained despite upregulation of aerobic capacity in cold-acclimated newts. Nevertheless, the activity of CK and LDH in skeletal muscle responds to thermal acclimation differently between populations, suggesting that the thermal plasticity of the anaerobic capacity of skeletal muscle has diverged between populations.
In conclusion, the thermal sensitivity and plasticity of locomotor performance differs between northern and southern populations of eastern newts. In conjunction with previous work (Wilson 2001), these results suggest that the thermal sensitivity of locomotor performance can diverge in amphibian populations from different thermal environments. Furthermore, our results demonstrate the thermal plasticity of locomotor performance is labile and can undergo differentiation among populations of the same species. The ability for thermal sensitivity and plasticity to differentiate among populations suggests that ectotherms may be better able to adapt to climate change than previously thought.
References
Alder-John HB, Morin PJ, Lawler S (1988) Thermal physiology, phenology, and distribution of tree frogs. Am Nat 132:506–520
Alder-John HB, Barnhart MC, Bennett AF (1989) Thermal sensitivity of swimming performance and muscle contraction in northern and southern populations of tree frogs (Hyla crucifer). J Exp Biol 142:357–372
Berner NJ, Bessay EP (2006) Correlation of seasonal acclimatization in metabolic enzyme activity with preferred body temperature in the eastern red spotted newt (Notophthalmus viridescens viridescens). Comp Biochem Physiol A 144:429–436
Berner NJ, Puckett RE (2010) Phenotypic flexibility and thermoregulatory behavior in the eastern red-spotted newt (Notophthalmus viridescens viridescens). J Exp Zool A 313:231–239
Berner NJ, Bullock JR, Jerome WG, Marquand TH, Waldrup CL (2012) Tissue-specific acclimation of enzyme activity in the eastern red spotted newt (Notophthalmus viridescens viridescens). Trends Comp Biochem Physiol 16:61–70
Dalziel AC, Ou M, Schulte PM (2012) Mechanisms underlying parallel reductions in aerobic capacity in non-migratory threespine stickleback (Gasterosteus aculeatus) populations. J Exp Biol 215:746–759
Dhillon RS, Schulte PM (2011) Intraspecific variation in the thermal plasticity of mitochondria in killifish. J Exp Biol 214:3639–3648
Downes SJ, Shine R (1999) Do incubation-induced changes in a lizard’s phenotype influence its vulnerability to predators? Oecologia 120:9–18
Elphick MJ, Shine R (1998) Longterm effects of incubation temperatures on the morphology and locomotor performance of hatchling lizards (Bassiana duperreyi, Scincidae). Biol J Linn Soc 63:429–447
Escobar J, Buck DG, Brenner M, Curtis JH, Hoyos N (2009) Thermal stratification, mixing, and heat budgets of Florida lakes. Fund Appl Limnol 174:283–293
Fangue NA, Richards JG, Schulte PM (2009) Do mitochondrial properties explain intraspecific variation in thermal tolerance? J Exp Biol 212:514–522
Feder ME, Lynch JF (1982) Effects of latitude, season, elevation, and microhabitat on field body temperatures of neotropical and temperate zone salamanders. Ecology 63:1657–1664
Glanville EJ, Seebacher F (2006) Compensation for environmental change by complementary shifts of thermal sensitivity and thermoregulatory behavior in an ectotherm. J Exp Biol 209:4869–4877
Guderley H (2004) Metabolic responses to low temperature in fish muscle. Biol Rev 79:409–427
Gunderson AR, Leal M (2012) Geographic variation in vulnerability to climate warming in a tropical Caribbean lizard. Funct Ecol 26:783–793
Gvozdik L, Puky M, Sugerkova M (2007) Acclimation is beneficial at extreme test temperatures in the Danube crested newt, Triturus dobrogicus (Caudata, Salamandridae). Biol J Linnean Soc 90:627–636
Hertz PE, Huey RB, Nevo E (1983) Homage to Santa Anita: thermal sensitivity of sprint speed in Agamid lizards. Evolution 37:1075–1084
Hochachka PW, Somero GN (2002) Biochemical adaptation. Mechanism and process in physiological evolution. Oxford University Press, New York
Huey RB, Stevenson RD (1979) Integrating thermal physiology and ecology of ectotherms: a discussion of approaches. Am Zool 19:357–366
Husak JF, Fox SF, Lovern MB, Van Den Bussche RA (2006) Faster lizards sire more offspring: sexual selection on whole-animal performance. Evolution 60:2122–2130
Kushlan JA (1979) Temperature and oxygen in an everglades alligator pond. Hydrobiologia 67:267–271
Munoz MM, Stimola MA, Algar AC, Conover A, Rodriguez AJ, Landestoy MA, Bakken GS, Losos JB (2014) Evolutionary stasis and lability in thermal physiology in a group of tropical lizards. Proc R Soc B 281:20132433
Petranka JW (2010) Salamanders of the United States and Canada. Smithsonian Books, Washington
Rogowitz GL (1996) Evaluation of thermal acclimation and altitudinal variation of metabolism in a neotropical lizard (Anolis gundlachii). Copeia 1996:535–542
Schnurr ME, Yin Y, Scott GR (2014) Temperature during embryonic development has persistent effects on metabolic enzymes in the muscle of zebrafish. J Exp Biol 217:1370–1380
Seebacher F (2005) A review of thermoregulation and physiological performance in reptiles: what is the role of phenotypic flexibility? J Comp Physiol B 175:453–461
Seebacher F, Holmes S, Roosen NJ, Nouvian M, Wilson RS, Ward AJW (2012) Capacity for thermal acclimation differs between populations and phylogenetic lineages within a species. Funct Ecol 26:1418–1428
Van Berkum FH (1986) Evolutionary patterns of the thermal sensitivity of sprint speed in Anolis lizards. Evolution 40:594–604
Verbeke G, Molenberghs G (2009) Linear mixed models for longitudinal data. Springer, New York
Wilson RS (2001) Geographic variation in thermal sensitivity of jumping performance in the frog Limnodynastes peronii. J Exp Biol 204:4227–4326
Wilson RS, Franklin CE (1999) Thermal acclimation of locomotor performance in tadpoles of the frog Limnodynastes peronii. J Comp Physiol B 169:445–451
Wilson RS, James RS, Johnston IA (2000) Thermal acclimation of locomotor performance in tadpoles and adults of the aquatic frog (Xenopus laevis). J Comp Physiol B 170:117–124
Acknowledgments
We thank Nancy J. Berner for sharing the protocol for the LDH assay and for giving us helpful advice on caring for newts, Anne C. Dalziel and Patricia M. Schulte for sharing their protocol for the CK assay, Michael Oxendine for his assistance collecting newts, and Keely Corder for her assistance with enzyme assays. Rick Wilson and Andrew McCullough from the Cathance River Preserve and Brunswick High School also deserve thanks for their assistance in locating and collecting newts in Maine. A. John Bailer, Jon P. Costanzo, Kathleen A. Killian, and Richard E. Lee, Jr. contributed constructive critiques of this manuscript. We also thank Rachael Morgan-Kiss for allowing use of her spectrophotometer. This work was funded by the Department of Biology at Miami University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by G. Heldmaier.
Rights and permissions
About this article

Cite this article
Mineo, P.M., Schaeffer, P.J. The thermal plasticity of locomotor performance has diverged between northern and southern populations of the eastern newt (Notophthalmus viridescens). J Comp Physiol B 185, 103–110 (2015). https://doi.org/10.1007/s00360-014-0869-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00360-014-0869-1