
Abstract
The growth, development, and quality of rice are inhibited by the presence of cadmium (Cd) in the soil, the uptake of which depends strongly on the pH of the growth medium. Here, a hydroponics-based experiment was conducted to characterize the relationship between Cd accumulation and the pH of the medium in seedlings of the two cultivars, Zhonjiazao17 (ZJZ17) and Xiangzhengyu (XZY). The uptakes of Cd by the roots of both cultivars and its translocation to the shoots were the highest from a medium of pH 6.0. XZY plants accumulated more Cd than ZJZ17 ones. Transcription profiling indicated that the genes, OsNRAMP1 and OsHMA2, were actively involved in Cd uptake and transport, as they were both strongly upregulated at pH 6.0. Both the Cd concentration of the medium and its pH exerted significant effects on the seedling growth. At pH 6.0, zinc was efficiently transported, but the translocation of iron was suppressed in shoot. The genes, OsZIP5 and OsYSL15, are the most likely responsible for the uptake and translocation of both these elements in rice seedlings.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The contamination of ground water, air and soil by heavy metals is of major concern worldwide. Rapid industrial development and population expansion, coupled with less efficient controls over pollution in many parts of the world, has resulted in a widespread occurrence of heavy metal contaminated agricultural soils. Cadmium (Cd) is the most mobile and the most toxic of the heavy metals, and has been implicated as responsible for a range of human health issues (Godt et al. 2006; Xue et al. 2014). It has been estimated that ~ 1,333,333 hm2 of agricultural soils have become polluted with Cd, and that annually some 0.15 Mt of agricultural products contain an excessive accumulation of Cd. The annual loss in value of Chinese primary produce resulting from Cd contamination is at least 20 Bn RMB (US$3 Bn) (Chen et al. 2013).
A number of soil factors have been shown to govern the uptake of Cd by plants, but of these, pH is probably the most important (Grant et al. 1999). The Cd adsorptive capacity of soils approximately triples for each unit increase in pH over the interval of pH 4–7 (Christensen 1984). The uptake of Cd into hydroponically grown Elodea canadensis shoots increases as the solution’s pH increases (Javed and Greger 2011), while less Cd accumulates in the roots and shoots of rice plants grown in a pH 5.5 solution than in those of plants grown at both a lower and a higher pH (Liu et al. 2013). In hydroponically grown transgenic lines of poplar (Populus tremula × P. alba), the net influx of Cd is highest at pH 5.0 (He et al. 2015), while in potato, more Cd accumulates in plants grown at a pH of 6.5 than at either pH 4.5 or 5.5 (Larsson and Asp 2013). In soil-grown rice, the Cd content of the grain is negatively correlated with the soil’s pH (Rafiq et al. 2014), while in maize, increasing the soil’s pH from 4.9 to 6.0 has the effect of reducing the shoot’s Cd concentration (Cunha et al. 2008). Cd transporter proteins are important determinants of Cd uptake and translocation. In rice, OsHMA2 is known to be a major transporter of both Cd and zinc (Zn) from the root to the shoot, localized in the plasma membrane of root pericycle cells at the vegetative growth stage. Since loss-of-function mutants accumulate much less Cd and Zn than do wild type plants in their root tips and in the shoot elongation zone, the suggestion is that OsHMA2 functions to transport Zn and Cd from the apoplast to the symplast via the phloem (Yamaji et al. 2013). OsNRAMP1, a member of natural resistance-associated macrophage protein family, participates in the uptake of both Cd and iron (Fe): the overexpression of OsNRAMP1 resulted in increased Cd content of wild type rice leaves (Takahashi et al. 2011b). OsNRAM5 has been reported to encode a major transporter of Cd (Sasaki et al. 2012), while the Fe-regulated transporters encoded by OsIRT1 and OsIRT2 are also thought to participate in Cd uptake (Uraguchi and Fujiwara 2012). Both OsIRT1 and OsNRAM5 were upregulated in rice cultivars grown under limed soil conditions (Yang et al. 2018).
The present study was designed to investigate the effect of pH on Cd accumulation in hydroponically-grown rice, to characterize the effect of Cd stress on plant growth and to monitor the transcription of key genes related to Cd uptake and transport at various pH levels. Whereas another objective was to establish at what pH level(s) the uptake of Cd by rice seedlings is at a minimum, in order to better manage crops grown under Cd contaminated soil.
Materials and Methods
Plant Materials and Growing Conditions
Grains of the two indica cultivars Zhonjiazao17 (ZJZ17, a low Cd accumulator) and Xiangzhengyu (XZY, high accumulator) were surface sterilized by immersion in 5% w/v sodium hypochlorite for 15 min, followed by an extensive rinse in sterile water. Germination proceeded at 30 °C for 48 h in the dark, followed by an additional 24 h at 35 °C. The germinating seedlings were allowed to grow in deionized H2O for seven days, after which they were transferred into one quarter strength Yoshida’s nutrient solution (Yoshida et al. 1976) for further seven days. On the day 14, the seedlings were moved into a full-strength nutrient solution adjusted to a pH of either 3.5, 5.0, 6.0, 7.0, or 8.0 by the addition of NaOH or HCl, and containing one of 0, 0.5, or 2.0 µM CdCl2. A 5 mM MES buffer was added to keep the pH constant and was checked every day. Mettler Toledo™ FiveEasy Plus™ FEP20 pH Meter was used in the whole experiment and the pH was monitored on daily basis. The nutrient solution was replaced every fourth day. The entire experiment was run in triplicate.
Trait Measurement
One day before harvest (on 27th day), the chlorophyll content (SPAD value) of the third expanded leaf was measured using a SPAD-5 chlorophyll meter (Konika Minolta, Tokyo, Japan). The readings were taken at the center of the leaf beside the midrib. The following day, the roots were rinsed three times in deionized water, then immersed for 2 min in 20 mM disodium ethylene diamine tetraacetate in order to remove any surface-adsorbed Cd. After a further three rinses in deionized water, the root and shoot were separated and fresh weight was measured, along with the length of the root and shoot. The root and shoot material was oven dried at 105 °C for 48 h, weighed and ground to a powder. A 100 mg aliquot of the powdered tissue was digested in 5 mL 65% HNO3, held at 150 °C to reduce the volume of the solution to 1 mL, then made up to 20 mL with Milli-Q water. The concentration of Cd, Fe and Zn (in mg per kg dry weight) was determined from these samples using an AA-6800 atomic absorption spectrophotometer (Shimadzu, Tokyo, Japan). Certified standards were used to avoid the errors.
RNA Extraction and Quantitative Real Time PCR (qRT-PCR) Analyses
RNA was extracted from shoots and roots of 28 days old rice seedlings using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA), then treated with DNase I (Promega, Madison, WI, USA) to remove contaminating genomic DNA. A 2 µL aliquot of the resulting RNA was reverse transcribed to cDNA using a PrimeScript® RT reagent kit (Takara, Tokyo, Japan). The primer sequences used for the subsequent qRT-PCR analyses (Table S1) were designed using Primer 5.0 software (PREMIER Biosoft International, Palo Alto, CA, USA). The Ubiquitin gene (LOC_Os03g13170) was chosen as the reference. Each 20 µL qRT-PCR contained 1 µL cDNA, 2.0 µL of each primer (10 µM), 10 µL 2.5 × Real Master Mix/20 × SYBR solution and 7 µL ddH2O. The reactions were given an initial denaturation (95 °C/3 min), then subjected to 40 cycles of 95 °C/20 s, 55 °C/10 s, 72 °C/20 s. At the end of the cycling process, a melting curve (65–95 °C) analysis was applied to check amplification specificity. Relative transcript abundances were derived using 2−∆∆CT method (Livak and Schmittgen 2001).
Statistical Analyses
The results are presented in means ± standard deviations (SD) of three independent biological replicates. Differences among treatments were assessed using three-way Analysis of Variance (ANOVA, p < 0.05), followed by LSD test in SPSS Statistics for Windows, v21.0 (IBM Corp., Armonk, NY, USA).
Results
The Effects on Seedling Growth of the Medium’s pH and Cd Stress
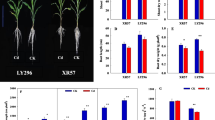
At each level of pH, the greater the concentration of Cd in the nutrient medium, the more inhibited was the growth of the seedlings (of both cultivars); the growth suppression effect of Cd stress was greatest when the nutrient medium was maintained at pH 6.0 (Fig. 1a, b). The effect of the stress on shoot and root fresh and dry weight, shoot and root lengths and leaf chlorophyll content (SPAD) are shown in Fig. 2. Increasing the concentration of CdCl2 significantly reduced (p < 0.05, Table S2) the seedling growth of both cultivars (Fig. 2a–f). The stress induced by the presence of either 0.5 µM or 2.0 µM CdCl2 had only a small (but nevertheless significant) effect on the leaf chlorophyll content (SPAD) (Fig. 2g). Instead of the concentration of CdCl2, seedling growth was reduced as the medium’s pH was raised from 3.5 to 6.0, then rose when the pH was increased to 7.0 and to 8.0. The poorest performance for each of the traits occurred at pH 6.0, irrespective of the CdCl2 concentration. XZY seedlings showed high in growth traits than ZJZ17 seedlings, except with respect to leaf chlorophyll content (SPAD).

The growth response of ZJZ17 (V1) and XZY (V2) grown in a medium held at a range of pH levels and containing a range of Cd concentrations. a Seedlings grown at pH (i) 3.5 (ii) 5.0 (iii) 6.0 (iv) 7.0 (v) 8.0. b Seedlings grown in the presence of (i) 0 µM Cd (ii) 0.5 µM Cd (iii) 2.0 µM Cd

The effect of different pH levels on rice seedlings grown in hydroponics solution containing three different concentrations (0, 0.5, 2.0 µM) of CdCl2. a Shoot fresh weight, b root fresh weight, c shoot dry weight, d root dry weight, e shoot length, f root length, g SPAD chlorophyll values of 28 days old seedlings. Black bars refer to ZJZ17 and white ones to XZY. Mean values were derived from the performance of three biological replicates. Different letters indicate significant differences (p < 0.05) between treatments and error bars indicate standard deviations
Cd Uptake and Transport are Affected by Both the Medium’s pH and Its Cd Concentration
The Cd content of both the shoot and root of both cultivars significantly increased (p < 0.05, Table S2) as the Cd content of the medium was increased: the highest concentration of Cd was accumulated when the plants were exposed to 2.0 µM CdCl2 (Fig. 3a, b). More Cd entered and translocated to the shoot of the plants as the medium’s pH was raised from 3.5 to 6.0, reducing from its peak level as the pH was raised beyond this to 7.0 and to 8.0. Peak Cd accumulation occurred at pH 6.0 at each of the CdCl2 levels in both cultivars, with the one exception of the roots of XZY seedlings exposed to 2.0 µM CdCl2, where peak Cd accumulation occurred at pH 7.0. More Cd accumulated in root as compared to the shoot of both cultivars at a given level of Cd stress. ZJZ17 seedlings tended to accumulate less Cd than XZY seedlings, both in the shoot and in the root.

The effects of varying the medium’s pH on the Cd content of seedlings exposed to a range of concentrations (0, 0.5, 2.0 µM) of CdCl2. The Cd content of the a shoot and b roots of 28 days old seedlings. Black bars refer to ZJZ17 and white ones to XZY. Mean values were derived from the performance of three biological replicates. Different letters indicate significant differences (p < 0.05) between treatments and error bars indicate standard deviations
The Impacts of pH of the Growth Medium and Cd Stress on Zn and Fe Accumulation
Zn content was largely unaffected by the medium’s Cd concentration (Fig. 4a, b). With respect to the response of medium’s pH, the ZJZ17 shoot Zn content was significantly higher (p < 0.05, Table S2) at pH 7.0, 6.0, and 6.0 under 0, 0.5 and 2.0 µM CdCl2, respectively. While in XZY, Zn was highly accumulated at pH 6.0 and 8.0 under 0 and 0.5 µM and pH 6.0 and 7.0 under 2.0 µM CdCl2 (Fig. 4a). The Zn content of the roots significantly increased (p < 0.05, Table S2) as the pH of the medium was raised from 3.5 to 7.0, but decreased upon its further rise to 8.0 with the exceptions the roots of XZY plants exposed to 0.5 µM CdCl2 and in the roots of ZJZ17 plants exposed to 2.0 µM CdCl2 (Fig. 4b). Fe translocation to shoot was suppressed by the outer CdCl2 concentration especially in 2.0 µM. In shoot, a V-type pattern was formed under 0.5 and 2.0 µM Cd where shoot Fe content was lowest in plants grown at pH 6.0 under 0.5 and 2.0 µM CdCl2 in both cultivars (Fig. 4c). The ZJZ17 root Fe content was highest in seedlings grown at pH 7.0 either in 0 µM or 0.5 µM CdCl2, and in those grown at pH 8.0 in the 2.0 µM Cd, but that of XZY was highest at pH 6.0 at each level of Cd stress (Fig. 4d).

The effects of varying the medium’s pH on the accumulation of Zn and Fe content in rice seedlings exposed to three different concentrations (0, 0.5, 2.0 µM) of CdCl2. a and b Zn content, c and d Fe content in a and c shoots, b and d roots of 28 days old seedlings. Black bars refer to ZJZ17 and white ones to XZY. Mean values were derived from the performance of three biological replicates. Different letters indicate significant differences (p < 0.05) between treatments and error bars indicate standard deviations
The Transcriptional Response to Varying Medium’s pH and Cd Contents
The transcriptional response of OsNRAMP1, OsNRAMP5, OsHMA2, OsIRT1, OsYSL15 and OsZIP5 in the shoots and roots of both cultivars was monitored using qRT-PCR and their statistical analysis is shown in (p < 0.05, Table S2). The abundance of OsNRAMP1 and OsHMA2 transcript responded positively to exposure to Cd stress in both the shoot (Fig. 5a, c) and the root (Fig. 5b, d) of both cultivars. The effect of raising the medium’s pH from 3.5 to 6.0 was to upregulate OsNRAMP1, while a further rise to pH 7.0 and 8.0 reduced the abundance of this transcript (the sole exception to this behavior was in the roots of XZY seedlings exposed to 2.0 µM CdCl2, where the peak level of OsNRAMP1 transcription occurred at pH 7.0 rather than at pH 6.0) (Fig. 5a, b). The transcriptional response of OsHMA2 was similar to that of OsNRAMP1, although in this case the abundance of transcript in the roots of XZY seedlings exposed to either 0.5 or 2.0 µM CdCl2 continued to increase up to a pH of 7.0. Generally, the abundance of both OsNRAMP1 and OsHMA2 transcript was higher in XZY than in ZJZ17 seedlings for a given pH/CdCl2 environment. Increasing the concentration of CdCl2 significantly suppressed the transcription of OsIRT1 in the shoots of both cultivars (Fig. 5e). For ZJZ17 plants exposed to either 0 or 0.5 µM CdCl2, the strongest transcription occurred when the medium’s pH was 6.0; however, when the external CdCl2 concentration was raised to 2.0 µM, transcript abundance occurred higher in the shoot of plants grown in a pH 8.0 medium. For XZY seedlings, an ascending order of OsIRT1 transcription in shoot was observed as pH increased under 0 or 0.5 CdCl2, but when seedlings were exposed to 2.0 µM CdCl2, the transcription level was low around pH 6.0 and was higher at the extreme pH (Fig. 5e); interestingly in the roots, the abundance of OsIRT1 transcript responded positively both to the concentration of CdCl2 in the medium and to its pH (Fig. 5f). In shoots of both cultivars, Transcript abundance of OsNRAMP5 was low and there was not much of an evidence of a response to pH under 0 and 0.5 µM CdCl2, but for 2.0 µM CdCl2, it seemed that the gene was induced by low pH in ZJZ17 and high pH in XZY (Fig. 6a). In roots, the highest abundance of OsNRAMP5 took place at pH 8.0, 7.0, and 6.0 in ZJZ17 and pH 6.0, 3.5, and 6.0 in XZY under 0.0 or 0.5 and 2.0 µM CdCl2, respectively (Fig. 6b). The peak transcription of OsZIP5 in the shoot occurred at pH 6.0 irrespective of the concentration in the medium of CdCl2 (Fig. 6c). Amazingly, OsYSL15 was transcribed at a low level at pH 6.0 in the shoot of plants exposed to both 0.5 and 2.0 µM CdCl2 stress (Fig. 6e). In the root, transcription of OsZIP5 was seemed low under 0 and 0.5 µM CdCl2 in both cultivars, however it showed maximum transcript levels at pH 6.0 in ZJZ17 and pH 7.0 in XZY cultivar under 2.0 µM CdCl2 (Fig. 6d). The transcriptional pattern of OsYSL15 (Fig. 6f) was consistent with that of the uptake of Fe in root of both cultivars.

The effects of varying the medium’s pH on the transcriptome of rice seedlings challenged with Cd stress. Relative transcript abundances of a and bOsNRAMP1, c and dOsHMA2, e and fOsIRT1 in a, c, and e the shoot and b, d, and f the root of 28 days old seedlings. Black bars refer to ZJZ17 and white ones to XZY. Mean values were derived from the performance of three biological replicates. Different letters indicate significant differences (p < 0.05) between treatments and error bars indicate standard deviations

The effects of varying the medium’s pH on the transcriptome of rice seedlings challenged with Cd stress. Transcript abundances in a, c, and e the shoot and b, d, and f the root of a and bOsNRAMP5, c and dOsZIP5, e and fOsYSL15 of 28 days old seedlings. Black bars refer to ZJZ17 and white ones to XZY. Mean values were derived from the performance of three biological replicates. Different letters indicate significant differences (p < 0.05) between treatments and error bars indicate standard deviations
Discussion
The Effects of Cd Stress and the Medium’s pH on the Growth of Rice Seedlings
The inhibition of root growth induced by Cd results in a reduction in the plant’s ability to take up both water and nutrients, thereby slowing the growth of the shoot (Chen et al. 2003). The growth response of the rice seedlings presented here was therefore as expected: for both cultivars, both the length and weight of the shoots and roots were compromised by the addition to the hydroponics medium of CdCl2 (Figs. 1a, b and 2a–f). Soil pH is well understood to be a major determinant of the growth of most crops (Spain et al. 1975), as it strongly affects the availability of most nutrients (Juo and Uzu 1977). The dry matter yield of rice tops and roots were greatest at pH values 5.5 (Alam 1981). In the present study, biomass accumulation was most favored when the pH of the medium was 5.0 or 7.0 (Fig. 2). The loss of photosynthetic pigments has been observed to be one of the earliest responses to Cd toxicity (Chen et al. 2019; Goel et al. 2012; Vassilev and Lidon 2011), a phenomenon which may be due to either the degradation of extant chlorophyll and/or to the inhibition of its de novo synthesis (Jain et al. 2007). Zhang et al. (2015) conducted an experiment on rice seedlings with 0, 50, 100 and 200 μM Cd and found that the increasing concentration of Cd significantly decreased the chlorophyll content of both W7 and M63 cultivars. The lowest chlorophyll content was observed when plants were treated with 200 μM Cd. Furthermore, Wang et al. (2018) reported a major QTL qGLR-3 responsible for the leaf green ratio under Cd stress condition in rice seedlings in a DH population. However, there was little evidence for chlorophyll loss in rice seedlings in the present experiments (Fig. 2g), probably because the CdCl2 concentrations employed were much lower than those used in other experiments (Chen et al. 2019; Zhang et al. 2015; Goel et al. 2012). The loss of growth and less SPAD chlorophyll values at pH 6.0 may have been due to the effect of Cd under this pH and/or independently, pH 6.0 inhibited the uptake of essential nutrients which are required for plant growth.
The Medium’s pH and Cd Content Affect the Uptake and Transport of Metal Ions
Experiments using soil or hydroponics have shown that plants absorb increasing quantities of Cd as the concentration of soluble Cd in the growth medium is increased (Sui et al. 2018; Yang et al. 2018; Song et al. 2015). The same result was observed in the present experiment where Cd concentration was increased in rice seedlings when solution Cd was enhanced by 0.5 and 2.0 µM CdCl2 (Fig. 3a, b). The pH of the medium affects both the solubility of Cd and its uptake (Wallace et al. 1977). In the root of poplar plants, Cd uptake has been shown to be low from either low or high pH media (He et al. 2015). Cd accumulation was greatest in plant parts of Dactylis glomerata when pH was enhanced up to 6.0 but decreased again at pH 7.0 when the plants were grown for a month in hydroponic Cd containing solution. Although same study also reported opposite results for Lolium perenne, Lactuca sativa, and Rorippa nasturstium–aquaticum where high Cd accumulation was observed at pH 7.0 rather than at pH 6.0 (Hatch et al. 1988). In potato, Cd was accumulated at a higher rate from a medium held at pH 6.0 than from one held at either pH 4.5 or 5.5 (Larsson and Asp 2013). According to a study of Cd uptake by Chrysophyllum albidium from soil, more Cd was accumulated ranging in pH from 2.0–6.0, but not from a soil of pH 8.0 (Onwu and Ogah 2010); the authors described that decline in Cd uptake at higher pH is due to the formation of CdOH+ and CdOHCl and at lower pH levels to increased competition from H+ ions with Cd for active sites on the adsorbent surface. Here, the Cd content of both the rice root and shoot was markedly lower in plants raised under the more extreme (both high and low) pH regimes than at either pH 6.0 or 7.0 (Fig. 3a, b). The possible reason for that could be either the expressions of genes related to Cd and/or the formation of cadmium species other than Cd2+.
The structural similarity between the cations Zn2+ and Cd2+ results in their being taken up by plants and then translocated using the same channels and/or carriers. In tomato, the Zn content of the shoot generally rises as the level of Cd is raised, although the opposite was in the case of both Capsicum annuum and Solanum melongena (Cikili et al. 2016). On the other hand, the shoot Fe content of Physalis peruviana and Capsicum annuum responds positively to Cd stress, but that of tomato responds negatively. In both Romaine Lettuce and Thlaspi caerulescens plants, Zn uptake appears to be at its highest at intermediate levels of pH (Chaney et al. 2009; Wang et al. 2006). The present experiments showed that the rice shoot Zn content was unaffected by the presence of CdCl2 in the medium, but that of shoot Fe content dropped as the external concentration of CdCl2 was increased (Fig. 4a–c). At the same time, the Fe content of the roots was unresponsive to Cd stress (Fig. 4d). In light of previous studies and focusing on our results, we can assume that the response of Zn and Fe vary among different plant parts of different crops under Cd stress.
The Medium’s pH Affects the Transcriptomes of Both the Shoot and the Root
The transcriptional response of genes encoding Cd transporters has been suggested as the most important determinant of Cd accumulation in rice (Gao et al. 2016). The OsNRAMP1 is found in the plasma membrane and participates in both the uptake and transport of Cd. The upregulation of OsNRAMP1 in the rice root has been correlated with a higher level of Cd accumulation in the shoot (Takahashi et al. 2011a). The OsHMA2 is found in the root pericycle cell plasma membrane, and is held to be responsible for the translocation of both Cd and Zn from the root to the shoot (Yamaji et al. 2013; Takahashi et al. 2012). The abundance of OsHMA2 transcript is known to be higher in plants exposed to Cd stress than in nonstressed ones (Nocito et al. 2011). Presently, the abundance in both test cultivars of both OsNRAMP1 (Fig. 5a, b) and OsHMA2 (Fig. 5c, d) transcript rose as the external concentration of CdCl2 was increased from 0 to 2.0 µM. Raising the growth medium’s pH has been shown to increase Cd entry into the cells of the green alga Chlamydomonas reinhardtii, a process driven by pH-induced conformational changes to specific Cd transport proteins (Francois et al. 2007), but as yet the mechanistic basis via which the medium’s pH affects Cd accumulation in rice has not been revealed. According to the present qRT-PCR analyses of the transcriptional consequences of altering the external medium’s pH, both OsNRAMP1 (Fig. 5a, b) and OsHMA2 (Fig. 5c, d) were abundantly transcribed at neutral and near-neutral levels of pH in both shoot and root, which implies that both of them may well be involved in Cd uptake and translocation. It has been shown that Fe-deficient rice plants take up larger amounts of Cd than do Fe-sufficient ones, an observation which has been taken to suggest that Fe transporters encoded by genes upregulated by Fe deficiency also contribute to the uptake of Cd (Nakanishi et al. 2006). In the present experiments, the level of Fe accumulation in the shoot of both cultivars exposed to Cd stress was lowest at pH 6.0 (Fig. 4c), a pH which favored the accumulation of Cd (Fig. 3a); the possibility is therefore that OsNRAMP1 was active in transporting Cd rather than Fe into the shoot. A greater abundance of OsNRAMP1 transcript has been identified in the Cd accumulator rice cultivar Habataki than in the non-accumulator cultivar Sasanishiki, although the two cultivars encode an identical OsNRAMP1 protein (Takahashi et al. 2011b). Similarly, the two cultivars compared here were selected on the basis that they contrasted for their capacity to accumulate Cd; the level of OsNRAMP1 transcription was higher in the (high Cd accumulator) XZY than in (low Cd accumulator) ZJZ17 (Fig. 5a, b).
The proteins OsIRT1 (Fe transporter) and OsNRAMP5 (manganese transporter) have also been identified as being important for Cd accumulation in a number of crop species. However, in the present study, the transcriptional response of neither OsIRT1 (Fig. 5e, f) nor OsNRAMP5 (Fig. 6a, b) was consistent with their involvement in Cd uptake. But the transcription levels of OsIRT1 in the roots of both cultivars increased continuously with increasing pH and CdCl2 concentrations (Fig. 5f). Similar results were reported by Yang et al. (2018), where he exhibited that the abundance transcription of OsIRT1 were sharply upregulated with increased lime concentrations and Cd concentrations. Along with that we also checked the abundance transcriptional response of OsCOPT1, OsMGT1, OsNAAT1, OsPT2 (Fig. S1), OsIRO2, and OsYSL2 (Fig. S2) listed in supplementary material but we could not find any strong relation of these genes with pH or Cd stress. Research has suggested that most of these genes have ability to transport multiple elements in plants but this was not our case of study. So, further study is needed to check that which element(s) was regulated by these genes when dealing with pH levels. ZIP proteins are thought to control the entry of Zn into root cells (Olsen and Palmgren 2014), while the barley ZIP5 protein is known to be involved in both Zn uptake and the root-to-shoot translocation of Zn (Tiong et al. 2015). Here, the abundance of OsZIP5 transcript in the shoot was particularly high in plants grown in a medium held at pH 6.0 (Fig. 6c), thereby potentially enhancing the root-to-shoot translocation of Zn in the presence of Cd stress. Finally, OsYSL15 is thought to be a key determinant of Fe homeostasis during early growth stages of the plant (Lee et al. 2009). Here, the transcriptional behavior of OsYSL15 was consistent with the accumulation of Fe in the root and shoot (Fig. 4c, d).
Conclusions
Tracking the performance of seedlings challenged with three concentrations of CdCl2 in the medium and five pH levels confirmed that growth of two rice cultivars was impacted by both factors. Increasing the external CdCl2 concentration resulted in an increased accumulation of Cd into the plants. The Cd accumulation was the highest from a near-neutral (pH 6.0) medium, driven by the upregulation of the genes—OsNRAMP1 and OsHMA2. The cultivar ZJZ17 accumulated less Cd than did XZY.
References
Alam SM (1981) Effects of solution pH on the growth and chemical composition of rice plants. J Plant Nutr 4:247–260
Chaney RL, Green CE, Ajwa HA, Smith RF (2009) Zinc fertilization plus liming to reduce cadmium uptake by Romaine lettuce on Cd-mineralized Lockwood soil. In: Paper presented at the Proceedings of the International Plant Nutrition Colloquium XVI, California, USA
Chen YX, He YF, Luo YM, Yu YL, Lin Q, Wong MH (2003) Physiological mechanism of plant roots exposed to cadmium. Chemosphere 50:789–793
Chen A, Wang M, Liu X (2013) Research progress on the effect of cadmium on rice and its absorption and tolerance mechanisms. Ecol Sci 32:514–522
Chen D, Chen D, Xue R, Long J, Lin X, Lin Y, Jia L, Zeng R, Song Y (2019) Effects of boron, silicon and their interactions on cadmium accumulation and toxicity in rice plants. J Hazard Mater 367:447–455
Christensen TH (1984) Cadmium soil sorption at low concentrations: I. Effect of time, cadmium load, pH, and calcium. Water Air Soil Poll 21:105–114
Cikili Y, Samet H, Dursun S (2016) Cadmium toxicity and its effects on growth and metal nutrient ion accumulation in Solanaceae plants. J Agr Sci 22:576–587
Cunha KP, do Nascimento CW, Pimentel RM, Ferreira CP (2008) Cellular localization of cadmium and structural changes in maize plants grown on a cadmium contaminated soil with and without liming. J Hazard Mater 160:228–234
Francois L, Fortin C, Campbell PG (2007) pH modulates transport rates of manganese and cadmium in the green alga chlamydomonas reinhardtii through non-competitive interactions: implications for an algal BLM. Aquat Toxicol 84:123–132
Gao L, Chang J, Chen R, Li H, Lu H, Tao L, Xiong J (2016) Comparison on cellular mechanisms of iron and cadmium accumulation in rice: prospects for cultivating Fe-rich but Cd-free rice. Rice 9:39
Godt J, Scheidig F, Grosse-Siestrup C, Esche V, Brandenburg P, Reich A, Groneberg DA (2006) The toxicity of cadmium and resulting hazards for human health. J Occup Me Toxicol 1:22
Goel S, Malik JA, Awasthi R, Sandhir R, Nayyar H (2012) Growth and metabolic responses of maize (C4 species) and rice (C3 species) genotypes to cadmium toxicity. Cereal Res Commun 40:225–234
Grant C, Bailey L, Mclaughlin M, Singh B (1999) Management Factors which influence cadmium concentrations in crops. Springer, Netherlands, pp 151–198
Hatch D, Jones L, Burau R (1988) The effect of pH on the uptake of cadmium by four plant species grown in flowing solution culture. Plant Soil 105:121–126
He J, Li H, Ma C, Zhang Y, Polle A, Rennenberg H, Cheng X, Luo Z (2015) Overexpression of bacterial gamma-glutamylcysteine synthetase mediates changes in cadmium influx, allocation and detoxification in poplar. New Phytol 205:240–254
Jain M, Pal M, Gupta P, Gadre R (2007) Effect of cadmium on chlorophyll biosynthesis and enzymes of nitrogen assimilation in greening maize leaf segments: role of 2-oxoglutarate. Indian J Exp Biol 45:385–389
Javed MT, Greger M (2011) Cadmium triggers elodea canadensis to change the surrounding water pH and thereby Cd uptake. Int J Phytoremediat 13:95–106
Juo A, Uzu F (1977) Liming and nutrient interactions in two ultisols from southern Nigeria. Plant Soil 47:419–430
Larsson JEH, Asp H (2013) Effects of pH and nitrogen on cadmium uptake in potato. Biol Plantarum 57:788–792
Lee S, Chiecko JC, Kim SA, Walker EL, Lee Y, Guerinot ML, An G (2009) Disruption of OsYSL15 leads to iron inefficiency in rice plants. Plant physiol 150:786–800
Liu D, Zhang C, Chen X, Yang Y, Wang S, Li Y, Hu H, Ge Y, Cheng W (2013) Effects of pH, Fe, and Cd on the uptake of Fe2+ and Cd2+ by rice. Environ Sci and Pollut Res Int 20:8947–8954
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25:402–408
Nakanishi H, Ogawa I, Ishimaru Y, Mori S, Nishizawa NK (2006) Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci Plant Nutr 52:464–469
Nocito FF, Lancilli C, Dendena B, Lucchini G, Sacchi GA (2011) Cadmium retention in rice roots is influenced by cadmium availability, chelation and translocation. Plant Cell Environ 34:994–1008
Olsen LI, Palmgren MG (2014) Many rivers to cross: the journey of zinc from soil to seed. Front Plant Sci 5:30
Onwu F, Ogah S (2010) Studies on the effect of pH on the sorption of cadmium (ll), nickel (II), lead (III) and chromium (VI) from aqueous solutions by African white star apple (Chrysophyllum albidium) shell. Afr J Biotechnol 9:7086–7093
Rafiq MT, Aziz R, Yang X, Xiao W, Rafiq MK, Ali B, Li T (2014) Cadmium phytoavailability to rice (Oryza sativa L.) grown in representative Chinese soils. A model to improve soil environmental quality guidelines for food safety. Ecotox Environ Saf 103:101–107
Sasaki A, Yamaji N, Yokosho K, Ma JF (2012) Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 24:2155–2167
Song W, Chen S, Liu J, Chen L, Song N, Li N, Liu B (2015) Variation of Cd concentration in various rice cultivars and derivation of cadmium toxicity thresholds for paddy soil by species-sensitivity distribution. J Integr Agr 14:1845–1854
Spain JM, Francis CA, Howeler RH, Calvo F (1975) Differential species and varietal tolerance to soil acidity in tropical crops and pastures. North Carolina State Univiversity Raleigh, NC, USA, pp 308–329
Sui FQ, Chang JD, Tang Z, Liu WJ, Huang XY, Zhao FJ (2018) Nramp5 expression and functionality likely explain higher cadmium uptake in rice than in wheat and maize. Plant Soil 433:377–389
Takahashi R, Ishimaru Y, Nakanishi H, Nishizawa NK (2011a) Role of the iron transporter OsNRAMP1 in cadmium uptake and accumulation in rice. Plant Signal Behav 6:1813–1816
Takahashi R et al (2011b) The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. J Exp Bot 62:4843–4850
Takahashi R, Ishimaru Y, Shimo H, Ogo Y, Senoura T, Nishizawa NK, Nakanishi H (2012) The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ 35:1948–1957
Tiong J, McDonald G, Genc Y, Shirley N, Langridge P, Huang CY (2015) Increased expression of six ZIP family genes by zinc (Zn) deficiency is associated with enhanced uptake and root-to-shoot translocation of Zn in barley (Hordeum vulgare). New Phytol 207:1097–1109
Uraguchi S, Fujiwara T (2012) Cadmium transport and tolerance in rice: perspectives for reducing grain cadmium accumulation. Rice 5:5
Vassilev A, Lidon F (2011) Cd-induced membrane damages and changes in soluble protein and free amino acid contents in young barley plants. Emir J Food Agr 23:130–136
Wallace A, Romney E, Alexander G, Soufi S, Patel P (1977) Some interactions in plants among cadmium, other heavy metals, and chelating agents. Agron J 69:18–20
Wang AS, Angle JS, Chaney RL, Delorme TA, Reeves RD (2006) Soil pH effects on uptake of Cd and Zn by Thlaspi caerulescens. Plant Soil 281:325–337
Wang J, Fang Y, Tian B, Zhang X, Zeng D, Guo L, Hu J, Xue D (2018) New QTLs identified for leaf correlative traits in rice seedlings under cadmium stress. Plant Grow Reg 85:329–335
Xue D, Jiang H, Deng X, Zhang X, Wang H, Xu X, Hu J, Zeng D, Guo L, Qian Q (2014) Comparative proteomic analysis provides new insights into cadmium accumulation in rice grain under cadmium stress. J Hazard Mater 280:269–278
Yamaji N, Xia J, Mitani-Ueno N, Yokosho K, Feng Ma J (2013) Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2. Plant Physiol 162:927–939
Yang Y, Chen J, Huang Q, Tang S, Wang J, Hu P, Shao G (2018) Can liming reduce cadmium (Cd) accumulation in rice (Oryza sativa) in slightly acidic soils? A contradictory dynamic equilibrium between Cd uptake capacity of roots and Cd immobilisation in soils. Chemosphere 193:547–556
Yoshida S, Forno DA, Cook JH, Gomez KA (1976) Laboratory manual for physiological studies of rice. International Rice Research Institute, Los Baños, Philippines, pp 61–66
Zhang X, Chen H, Jiang H, Lu W, Pan J, Qian Q, Xue D (2015) Measuring the damage of heavy metal cadmium in rice seedlings by SRAP analysis combined with physiological and biochemical parameters. J Sci Food Agric 95:2292–2298
Acknowledgements
This work was supported by the National Key Research and Development Program of China (2017YFD0100300; 2016YFD0101801), Agricultural Sciences and Technologies Innovation Program of the Chinese Academy of Agricultural Sciences (CAAS), and by the Central Level, Non-Profit, Scientific Research Institutes Basic R & D Operations Special Fund (Y2017PT46; 2017RG002-1).
Author information
Authors and Affiliations
Contributions
PH, XW, and ST conceived and designed experiment. UA performed the experiment. MZ, TS, and LX helped in hydroponics-based solution preparation and metal determination. MZ, TS, and SF helped RNA extraction. SA made the graphs and figures. UA, ZS, XW, and GJ analyzed the data and drafted the manuscript which was critically revised by PH and XW, All the authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Conflicts of interest
The authors have no conflicts of interest to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Ali, U., Zhong, M., Shar, T. et al. The Influence of pH on Cadmium Accumulation in Seedlings of Rice (Oryza sativa L.). J Plant Growth Regul 39, 930–940 (2020). https://doi.org/10.1007/s00344-019-10034-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00344-019-10034-x