
Abstract
Rapid climate change due to anthropogenic greenhouse gas emissions is pushing corals to and past their physiological limits, while their microbiome is being pressed towards dysbiosis. Microbes greatly influence the health and functioning of corals, but thermal anomalies that cause bleaching can affect certain taxa of the host-associated prokaryote and Symbiodiniaceae communities, leading corals towards a disease-prone state. In some coral species, however, even significant thermal stress may not result in visible signs of bleaching. Whether changes in the coral microbiome occur in these resilient species during temperature anomalies is not well described. In the present study, we tagged, visually assessed and sampled 10 colonies of the branching coral Pocillopora acuta on a fringing reef off Orpheus Island in the central Great Barrier Reef for 1 yr, of which the summer coincided with the 2016 mass-bleaching event. No visible signs of bleaching were observed in any of the 10 colonies throughout the study period, despite experiencing two degree heating weeks of thermal stress and observations of bleaching in other coral species on the same reef. Metabarcoding based on the Symbiodiniaceae ITS2 rDNA spacer and the bacterial 16S rRNA gene provided evidence for stability of the overall microbial community structure, although the bacterial community showed increases in a number of potentially beneficial taxa, such as diazotrophs, during the thermal stress event. These findings suggest some flexibility in the microbiome to adjust to higher than average temperatures without disrupting microbiome stability, perhaps contributing to the thermal resilience of P. acuta.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Climate change is causing global coral decline as a result of increasing sea surface temperatures and changes in ocean chemistry. Specifically, mass-bleaching as a result of thermal stress has decimated coral reefs in the past two decades (Heron et al. 2016; Oliver et al. 2018), exemplified by the 2016 bleaching event that resulted in ~ 30% mortality of coral on the Great Barrier Reef (GBR) alone (Hughes et al. 2017). The health and functioning of corals are governed in part by their associated micro-organisms. These include their photosynthetic algal endosymbiont of the family Symbiodiniaceae, as well as other single-celled and filamentous eukaryotes (e.g. fungi or algae), prokaryotes (i.e. bacteria and archaea), and acellular viruses, many of which play important roles in a range of biological processes. Symbiodiniaceae algae contribute to both carbon fixation and translocation that meet a large portion of the coral host’s respiratory requirements (Muscatine and Porter 1977). Prokaryotes contribute to cycling and metabolism of various nutrients within the coral holobiont, such as nitrogen and sulphur. These nutrients are crucial for the growth and health of the coral host as well as its algal endosymbionts (Raina et al. 2009; Lema et al. 2014; Rädecker et al. 2015), and can aid in host immunity (Ritchie 2006). Thermal stress, however, can significantly alter coral-associated microbial communities. These changes may allow for rapid acclimatisation or adaptation to environmental change (e.g. Reshef et al. 2006; Torda et al. 2017), but may also drive the coral holobiont towards a disease-prone state (e.g. Ben-Haim and Rosenberg 2002; Rosenberg et al. 2007; Vega Thurber et al. 2009).
Bleaching, by definition, is the loss of Symbiodiniaceae from the coral tissue (Glynn 1984), where certain Symbiodiniaceae types may be preferentially removed by the host (Jones et al. 2008). Some corals exhibit a shift to dominance by a more heat-tolerant Symbiodiniaceae type following bleaching (Berkelmans and van Oppen 2006) or acquire novel strains that are more thermally tolerant from the environment (Boulotte et al. 2016). Major shifts in the composition of bacterial communities have been previously observed in stressed corals (Bourne et al. 2008; Littman et al. 2011) that may include increases in pathogenic bacteria (e.g. Ben-Haim and Rosenberg 2002; Rosenberg et al. 2007; Bourne et al. 2008; Vega Thurber et al. 2009). Conversely, some coral species have been found to harbour stable communities during thermal stress (e.g. Webster et al. 2016; Hadaidi et al. 2017; Grottoli et al. 2018), and others have exhibited increases in abundances of beneficial bacteria, such as nitrogen-fixing diazotrophs (Santos et al. 2014). While these variations in response of the bacterial community to thermal stress may be specific to the coral host, characterising the response of the coral microbiome to thermal stress in resilient species may provide insight into the drivers of coral health and potential mechanisms of slowing, preventing or even reversing coral decline (i.e. “Assisted Evolution”; van Oppen et al. 2015). With the increased risk of thermal stress worldwide, investigation into the contribution of microbial communities to either coral decline or coral resilience is imperative.
The majority of literature on the effects of thermal stress on the coral microbiome is focused on changes that occur during and after bleaching. However, effects of temperature anomalies may result in changes in microbial communities that occur prior to or without visible signs of deteriorating health or bleaching. In fact, Glasl et al. (2017) propose the use of microbial communities as a diagnostic tool, speculating that changes to coral microbial communities can provide early detection of ecosystem stress. The present study followed tagged colonies of the common branching coral Pocillopora acuta (chunky morphology) for 1 yr of which the summer spanned the 2016 mass-bleaching event on the GBR (Hughes et al. 2017). Despite experiencing heat stress, the tagged colonies showed no visible signs of bleaching. This presented an opportunity to examine the effects of a thermal stress event on the long-term stability of the coral microbiome from a thermally resilient species.
Methods
Sample environment, collection and fixation
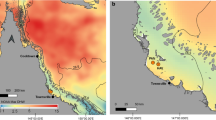
Ten colonies of P. acuta were tagged, visually examined and sampled for 1 yr spanning from November 2015 through November 2016 at Little Pioneer Bay (Orpheus Island) in the central Great Barrier Reef, Australia (Fig. 1). In the time since sampling occurred for the present study, P. acuta has been found to show inter-colony variation in bleaching susceptibility depending on colony morphology. It is now recognised that this branching coral species appears in two morphologies; a fine-branching morph and a chunky morph, where the former tends to bleach more readily (Smith et al. 2017). All colonies in the present study were of the chunky morphology and were located at 2–3 m depth along a 50 m stretch of reef parallel to the shoreline. Sampling took place every 2 months (November, January, March (a and b), May, July, September, November). Colonies were sampled twice in March, due to the onset of the mass-bleaching event across the central and northern GBR (Hughes et al. 2017). The March (a) time point occurred when bleaching was prominent at the sampling site. At each time point, a small nubbin (2–5 cm in length) of each colony was collected, immediately snap-frozen in liquid nitrogen (LN2) and stored at − 80 °C.

Map of study site at Little Pioneer Bay on Orpheus Island in the central Great Barrier Reef, Australia, located at approximately 18° 36′3.6″S 146° 29′20.4″E
A HOBO UA-001064 Pendant temperature logger (Onset Computer Corp.) was placed at the study site at the same depth and within the vicinity of the tagged colonies throughout the study period, taking a temperature reading every 30 min.
DNA extraction, PCR and sequencing
Nubbins were crushed on LN2 using a hydraulic bench top laboratory press prior to DNA extraction. DNA was extracted from whole coral homogenate using the method outlined in Wilson et al. (2002), modified to include an additional lysozyme digestion and bead-beating step. Amplification of double-stranded products from the 16S rRNA gene for bacteria and the rDNA internal transcribed spacer region 2 (ITS2) for Symbiodiniaceae was achieved through polymerase chain reaction (PCR) using gene-specific primers. The V5-V6 region of 16S was targeted using the primers 784F 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGAGGATTAGATACCCTGGTA -3′ and 1061R 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGCRRCACGAGCTGACGAC -3′ (Andersson et al. 2008). ITS2 was targeted using the primers ITS2F 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGGTGAATTGCAGAACTCCGTG-3′ (Pochon et al. 2001) and ITS2R 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGCCTCCGCTTACTTATATGCTT-3′ (Stat et al. 2009a). Both sets of primers contained the Illumina adapter overhangs (underlined above). The 16S PCR was performed in triplicate 10 µL reactions. Each reaction consisted of: 5 µL of AmpliTaq Gold MasterMix (Applied Biosystems), 2 µL of each primer (2 µM stock), and 1 µL of DNA template. The following PCR protocol was used: initial denaturation at 95 °C for 10 min, 30 cycles each of 95 °C for 30 s, 57 °C for 60 s and 72 °C for 60 s, followed by a final extension at 72 °C for 7 min. The ITS2 PCR was also performed in triplicate 10 µL reactions. Each reaction consisted of 5 µL of Qiagen Mulitplex MasterMix (Qiagen), 3 µL Milli Q water, 0.5 µL of each primer (4 µM stock), and 1 µL of DNA template. The following PCR protocol was used: 95 °C for 5 min, 31 cycles each of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s, followed by a final extension at 72 °C for 5 min. PCR clean-up, indexing and sequencing were carried out at the Ramaciotti Centre for Genomics at the University of New South Wales on a 2 × 300 bp Illumina MiSeq run. Negative PCR controls were also run alongside samples for sequencing to check for contamination.
Sequence assembly, quality control and taxonomic assignment
Demultiplexed sequences for both 16S and ITS2 were assembled, checked for quality and assigned taxonomic classification using QIIME2 v 2017.10 (QIIME 2 Development Team 2017). The plug-in demux (QIIME 2 Development Team 2017) was used for visualising read quality and setting quality filtering guidelines. Quality filtering, trimming of poor quality bases, de-replication, chimera filtering, merging of paired-end reads and identifying fine-scale single nucleotide variation among sequences were performed using the DADA2 plug-in (Callahan et al. 2016). For 16S, mitochondrial and chloroplast sequences were removed and taxonomic assignment was done by training a naïve-Bayes classifier on the V5-V6 region of the 16S gene in the SILVA 128 database (Quast et al. 2013) to match the primers used. Due to the high number of single-variants found for ITS2 and the subsequent small taxonomic database, it was not useful to use a classifier as above due to low resolution. Therefore, these single-variants for ITS2 were clustered by 97% similarity using a vsearch plug-in (Rognes et al. 2016). Taxonomic assignment was done according to the database from Arif et al. (2014), allowing assignment down to the sub-type level for Symbiodiniaceae. At the end of the pipelines for both 16S and ITS2, the taxa plug-in (QIIME2 Development Team 2017) was used to create a taxonomy table and an OTU or sequence variant biom table of raw sequence counts that could then be used for further downstream analyses.
Statistical analyses
All statistical analyses were performed in R v. 3.5.0 (The R Core Team 2018), and all graphics done using the R package ggplot2 (Wickham 2009) (See Supplementary Materials 1). Temperature data from the logger were collated and presented as daily temperatures. Degree heating weeks (DHWs) were calculated as the sum of the daily temperature anomalies (i.e. ≥ 1 degree higher than the maximum monthly mean) over a 90-d window, divided by 7 (Liu et al. 2013). Maximum monthly mean was derived from long-term temperature data (1993–2017) at Orpheus Island that is publicly available from the Australian Institute of Marine Science (AIMS) (AIMS Historical Data Tool: http://data.aims.gov.au/aimsrtds/datatool.xhtml) and the daily temperature was taken from the HOBO logger at the study site. It was assumed there were no temperature anomalies beyond the summer maximum temperatures in the 90 d prior to the first temperature data point taken in November 2015 (see AIMS Historical Data Tool: http://data.aims.gov.au/aimsrtds/datatool.xhtml), which allowed for complete profiling of DHWs during the summer period.
Alpha diversity, richness and evenness of 16S and ITS2 sequence data were examined using the package vegan (Oksanen et al. 2018). Significant differences in these three measures among the eight time points were assessed by analysis of variance (ANOVA) using either a linear mixed effects model fit by restricted maximum likelihood (REML) for repeated measures with an added autoregressive first-order (AR1) correlation structure to account for the autocorrelation, or a generalised least squares model fit by REML for repeated measures using an added autoregressive moving average (ARMA) correlation structure using nlme (Pinheiro et al. 2018) and car (Fox and Weisberg 2011). The Tukey’s test for pairwise comparisons was run on the model using multcompView (Graves et al. 2015) and lsmeans (Lenth 2016). To assess beta-diversity, repeated measures permutational multivariate analyses of variance (PERMANOVA, strata = colony) were performed and homogeneity of dispersions (PERMDISP) was checked to examine variations in beta-diversity and dispersion between and among time points in vegan (Oksanen et al. 2018); both permutational tests used 999 permutations. Post-hoc pairwise PERMANOVA was performed in pairwiseADONIS (Arbizu 2017) to assess between time point significance. Differences among time points were visualised using nMDS, line plots and stacked bar plots. Intra- and inter-colony dispersions were also quantified using betadisper in vegan (Oksanen et al. 2018) and tested against each other using a two-sample t test. Indicator species were checked among time points using a multi-level pattern analysis with 999 permutations using indicspecies (De Cáceres and Legendre 2009).
Using the package phyloseq (McMurdie and Holmes 2013), bacterial taxa driving the temporal patterns were identified by examining the relative abundances of bacterial taxa among samples and time points as well as identifying the taxa present in 100% of the colonies within a given time point, visualised using bubble plots and heatmaps. Further exploration of individual taxon abundances through time were also analysed by ANOVA, again using a mixed effects model fit by REML for repeated measures with an added autocorrelation structure in nlme (Pinheiro et al. 2018), car (Fox and Weisberg 2011), multcompView (Graves et al. 2015) and lsmeans (Lenth 2016).
Results
Orpheus Island seawater temperatures and thermal stress
There were no visible signs of bleaching in any of the tagged colonies throughout the study period, despite high temperatures occurring in Little Pioneer Bay during Feb-May 2016 (Fig. 2). This time period coincided with a mass-bleaching event on the Great Barrier Reef, and temperatures reached a high of 32.6 °C on 20 February 2016, exceeding 32 °C (daytime temperature) for 5 d in a row. February experienced a total of 8 d of temperatures reaching above 32 °C, and 19 d exceeding 31 °C. Little Pioneer Bay reached 2 DHWs at the end of February and remained at 2 DHWs through April (Fig. 2). The reefs around Orpheus Island were reported between 10 and 30% bleached at this time (Hughes et al. 2017) and within the study area, many coral species, such as the less robust acroporids and pocilloporids that included colonies of P. acuta of the fine-branching type, were found bleached in early March (Smith et al. 2017; Hughes et al. 2017; pers. obsv).

Daily temperature profile (red line is the daily average) and degree heating weeks for Little Pioneer Bay from November 2015–November 2016. The black arrows indicate sampling time points. Photographs of a representative tagged colony (X1) show no visible signs of bleaching throughout the study period
Amplicon sequencing
ITS2 Symbiodiniaceae Community: A total of 2,660,251 sequences from 79 whole coral homogenate samples were recovered to describe the Symbiodiniaceae community within P. acuta samples. Clustering at the 97% sequence similarity threshold identified 16 unique OTUs at the sub-type taxonomic level. Negative controls were checked visually using nMDS for overlap and removed from the data set.
16S rRNA Bacterial Community: A total of 2,582,381 sequences from 79 whole coral homogenate samples were recovered for bacterial community analysis. Single variant methods identified 3,358 unique bacterial sequence variants (putative OTUs); no archaeal sequences were retrieved. Singletons were removed from the data set, and negative controls were checked visually using nMDS for overlap. Sequence variants that were present in negative controls were considered contaminants if they contained high relative abundances in negative controls and low relative abundances in samples (Lee et al. 2015). Despite no visual overlap, negative controls contained four sequence variants of the taxa Bradyrhizobium, Ralstonia, and Oxalobacteraceae that followed this pattern, and were subsequently removed from all samples of the data set for further analysis.
Raw sequence data for this study are available on the NCBI Sequence Read Archive under the accession SRP150755.
Characterisation of the Symbiodiniaceae community structure across time
The Symbiodiniaceae alpha diversity and community structure (beta-diversity) within P. acuta did not differ significantly among time points (Fig. 3a). At each time point, the Symbiodiniaceae communities were dominated by Cladocopium goreaui (type C1), making up on average a relative abundance of 88.9 ± 0.3% (mean ± SE). Type C1 was largely comprised of the sub-type C1d. Cladocopium type C42 on average accounted for 8.2 ± 0.2% (mean ± SE) of the Symbiodiniaceae community. Durusdinium type D1 was present in low relative abundances (average < 1%) (Fig. 3b). However, this was fully driven by a single colony, which gained a relative abundance of 13.5% of type D1 in July but was completely absent at all other time points and in the other nine colonies.

a Alpha diversity (Shannon diversity index) of Symbiodiniaceae communities and b mean relative abundance of Symbiodiniaceae types among time points ranging from November 2015 through November 2016
Characterisation of the bacterial community structure over time
The most abundant bacterial phylum in P. acuta was Proteobacteria, with Gammaproteobacteria, Cytophagia, Alphaproteobacteria, Betaproteobacteria, Deltaproteobacteria, Flavobacteriia, Sphingobacteriia, Spirochaetes and Acidimicrobiia and Bacilli being the top ten most abundant classes (Figure S1). The top four most abundant classes were further broken down into orders to examine the structure of the P. acuta microbiome among time points (Fig. 4). The order Oceanospirillales occurred at a relative abundance between 15 and 42% among time points, where the genus Endozoicomonas made up over 61% of all Oceanospirillales and varied between 44.5 and 78% on average per time point (Fig. 4a). Cytophagia was fully comprised of the order Cytophagales, in which a single candidate genus, “Candidatus Amoebophilus”, accounted on average for over 84% of all Cytophagales (Fig. 4b). Alphaproteobacteria was dominated by the order Rhodobacterales, which was comprised mostly of the genera Roseovarius, Thalassobius and other unknown or uncultured Rhodobacterales (Fig. 4c). Betaproteobacteria consisted almost entirely of the order Burkholderiales, which was dominated by the genera Burkholderia-Paraburkholderia (97.8%) (Fig. 4d).

Relative abundances of the top four most abundant classes a Gammaproteobacteria, b Cytophagia, c Alphaproteobacteria and d Betaproteobacteria broken down into order across time points ranging from November 2015 to November 2016. Pie charts show a breakdown of genera in the most abundant order from each class
Alpha diversity, richness and evenness in P. acuta microbial communities were variable over time. At the sequence variant level, alpha diversity (Shannon-Weiner diversity index) differed significantly between time points (ANOVAdiv: df = 7, F = 5.168, p < 0.001), as did species richness (ANOVArich: df = 7, F = 3.69, p < 0.01) and evenness (ANOVAev: df = 7, F = 4.29, p < 0.001). Both alpha diversity and evenness showed significant variation between May 2016 and both March (a) 2016 and November 2015. Richness showed significant differences between May and November 2015 (Fig. 5a–c). The bacterial community composition (beta-diversity) at the sequence variant level also significantly differed among time points (PERMANOVA: df = 7, MS = 0.47, p = 0.001; Fig. 5d), with no significant variation in dispersion (PERMDISP: df = 7, MS = 0.02, p = 0.23). The ordination plot showed some separation between January and March (a) from the rest of the time points (Fig. 5d); however, further pairwise comparisons found no significant differences in beta-diversity between time points (Table S1). In addition, intra-colony dispersion was significantly lower than inter-colony dispersion among time points (t = 2.18, df = 14.099, p = 0.047; Figure S2). Relative abundances of the top 50 families showed some individual variation but little general variation among time points (Fig. 6). Each time point contained a number of significant indicator species that ranged between 1 and 26 (Table S2). March (a) 2016 had the highest number of indicators, followed by November 2015, January 2016 and March (b) 2016, respectively.

The a alpha diversity (Shannon Diversity index), b evenness, c species richness, and d beta-diversity (data clouds visualised using nMDS) among time points, ranging from November 2015 through November 2016. Ellipses represent 95% confidence intervals

Relative abundance of the top 50 most abundant families among time points, ranging from November 2015 through November 2016. Families in bold are members of the coral microbiome that are further discussed below
Specific individual taxa were further examined across time points. Despite visual trends of relative abundance through time of the top 10 most abundant classes (Fig. 7a), there was little evidence of significant variations. Relative abundances of the classes Gammaproteobacteria, Alphaproteobacteria and Betaproteobacteria did not vary significantly through time. The relative abundance of Cytophagia differed significantly among time points (ANOVA: df = 7, F = 2.21, p < 0.05), but was driven by significant differences only between November 2015 and November 2016. Relative abundances of the genera Endozoicomonas and Vibrio also showed no significant variation in relative abundances through time; however, November 2016 displayed unusually high variance for Vibrio (Fig. 7b). There was no relationship between Vibrio and Endozoicomonas (R2 = 0.004, data not shown).

Mean relative abundance of a top most abundant bacterial classes and b the two genera Endozoicomonas and Vibrio throughout the study period from November 2015 to November 2016
The temporal patterns of specific taxa were analysed by examining those present in all colonies within a given time point. At each time point, only a few members of the bacterial microbiome were present in all colonies, where Burkholderia-Paraburkholderia was the only taxon present in all colonies across time points. Most time points displayed differing combinations of bacteria that were present in every sample. Over 94% of samples included at least one or more sequence variants of Endozoicomonas, and while some colonies contained no Endozoicomonas at some time points, six of the 10 colonies had at least one or more sequence variants of Endozoicomonas at every time point. “Candidatus Amoebophilus” was present in high abundances throughout the study period in every colony aside from one, in which it was absent during seven of the eight time points.
Discussion
This study showed that thermal stress during the 2016 summer had a minimal effect on the microbiome composition of P. acuta in Little Pioneer Bay. The Symbiodiniaceae community remained stable throughout the sampling period and while the bacterial community showed some evidence of variation through time, overall stability was maintained, including during the months that coincided with the mass-bleaching event on the GBR. According to Hughes et al. (2017), two DHWs elicited a widespread bleaching response across the GBR at the end February 2016, and their aerial surveys found the reefs around Orpheus Island were between 10-30% bleached during this time period. The temperature data from the present study corroborate these findings and suggest that this was a period of thermal stress for corals in Little Pioneer Bay, which experienced two DHWs from the end of February through April. While many corals were bleached at the study site, the targeted colonies in this study remained unbleached.
The dominant Symbiodiniaceae type from the whole coral homogenate samples was Cladocopium goreaui (type C1) and remained the same for all 10 colonies throughout the 1-yr study period. Pocilloporid Symbiodiniaceae communities are commonly dominated by members of the genus Cladocopium (formerly clade C, LaJeunesse et al. 2018), sometimes by type C1 (Magalon et al. 2007) and at other times types C42 and C33 (Sampayo et al. 2009; Tonk et al. 2013). Sequence types C42 and C1 are likely intragenomic variants of the same Symbiodiniaceae species (Sampayo et al. 2009). Symbiodiniaceae is a key player in the thermal acclimatisation of coral hosts, and some corals have previously been found to either shuffle their dominant types or acquire novel Symbiodiniaceae types following severe bleaching (Berkelmans and van Oppen 2006; Boulotte et al. 2016; Cunning et al. 2018). However, the Symbiodiniaceae community in P. acuta in the present study remained unchanged throughout the entire study period. These results suggest a high degree of temporal stability in the Symbiodiniaceae community of P. acuta in the absence of bleaching, which is consistent with previous studies showing minimal community changes through both time (LaJeunesse et al. 2005; Thornhill et al. 2006a, b; Klepac et al. 2015; Cai et al. 2018) and during a similar sub-bleaching thermal stress event (Stat et al. 2009b). It is possible that sub-bleaching thermal stress does not generally affect the stability and maintenance of the Symbiodiniaceae communities. Alternatively, the ability of the coral host to shift this community in response to thermal pressure may be species-specific and reliant on the degree or severity of thermal stress (Cunning et al. 2018). Symbiodiniaceae types in the genus Durusdinium (formerly clade D, LaJeunesse et al. 2018) have often been associated with high thermal tolerance of their coral host, and corals have been found to shift their dominant Symbiodiniaceae from Cladocopium to Durusdinium following bleaching events (Berkelmans and van Oppen 2006; Jones et al. 2008). In contrast, the present study found only one colony that contained type D1, and not only was it never the dominant type, its acquisition in the colony did not occur directly after the thermal stress event and was instead present only in July. It could, however, be argued that this is still a delayed result of the heat stress. If some of the Cladocopium types were impaired in the months following the heat stress, this could have allowed the Durusdinium type to outcompete the Cladocopium types for a short period of time. Regardless, the shifting of dominant Symbiodiniaceae from Cladocopium to Durusdinium types was not observed within P. acuta during this study and suggests that the tolerance of this P. acuta morphology is not necessarily reliant on its association with a thermally tolerant symbiont such as those in the genus Durusdinium.
Gammaproteobacteria, Alphaproteobacteria, Cytophagia and Betaproteobacteria dominated the bacterial community of P. acuta whole coral homogenate samples throughout the study period. The only bacterium that was present in 100% of samples at all time points was a strain in the genus Burkholderia-Paraburkholderia, of the family Burkholderiaceae. Burkholderia spp. have previously been found to be abundant members of the early life stages of corals (Leite et al. 2017) and are suggested to play a role in nitrogen fixation as has been demonstrated for plants (reviewed by Coenye and Vandamme 2003). One of the most abundant genera in this data set was the candidate genus “Candidatus Amoebophilus” of the family Flammeovirgaceae, and one sequence variant was present in nine of the 10 colonies in high abundances throughout the study period. It is a recognised intracellular symbiont of amoebae (Horn et al. 2001) and has the potential for establishing a symbiotic relationship with other eukaryotes (Horn et al. 2001; Schmitz-Esser et al. 2010). Recently, it has been found to consistently associate with numerous coral species and is thought to associate with other eukaryotic hosts within the coral microbiome, such as the Symbiodiniaceae, or unidentified amoebae within coral tissue (Apprill et al. 2016; Huggett and Apprill 2018). The genus Endozoicomonas, of the family Hahellaceae, was present in the majority of samples, likely representing an important member of the P. acuta microbiome. This genus has been found in a variety of coral species, including those in the families Pocilloporidae (Ziegler et al. 2016; Neave et al. 2017), Acroporidae (Ziegler et al. 2016), Poritidae (Apprill et al. 2016), and Fungiidae (Roder et al. 2015), and their abundances are often linked with coral health (e.g. Bourne et al. 2008; Morrow et al. 2015; Ziegler et al. 2016). Endozoicomonas typically resides within coral tissues as aggregates, and some strains have been suggested to be responsible for carbohydrate upcycling (e.g. assisting the coral host with carbohydrate metabolism and transport) and provision of amino acids to the coral host (Neave et al. 2016, 2017), as well as protection from thermal or bleaching stress (Pantos et al. 2015).
Over the study period, there was a significant difference in bacterial beta-diversity. However, further statistical tests did not resolve which time points drove this significance, instead suggesting overall bacterial stability through time and during the thermal stress event. This is further supported by the significantly lower intra-colony variation among time points in comparison with inter-colony variation, implying that while there may be differences among colonies, bacterial communities are relatively stable within individual colonies throughout the thermal stress event. Multi-level pattern analysis, which identifies the microbes that are indicative of a specific time point, found twenty-six significant indicator species during the March (a) 2016 time point, coinciding with the mass-bleaching event. Understanding these fine-scale microbiome changes in bleaching-resistant corals may provide an opportunity to identify beneficial microbes for coral resilience. At this time point, there were four indicator taxa of the phylum Actinobacteria, which have been found to have antimicrobial, antioxidant and antiparasitic properties (Valliappan et al. 2014). Although these bacteria may provide some level of protection to the coral host during thermal stress, other Actinobacteria species were also found as indicators during the January 2016 time point, instead suggesting that the presence of Actinobacteria may be following a seasonal pattern, as has been found previously (Cai et al. 2018). Three nitrogen-fixers (diazotrophs) from the order Rhizobiales were also found as indicators of the March (a) time point, which supports previous findings by Santos et al. (2014) who propose increases in diazotroph abundance are indicative of thermal stress in corals. This increase may represent a beneficial mechanism for maintaining homoeostasis by providing otherwise limited nitrogen to the Symbiodiniaceae (Olsen et al. 2009; Lema et al. 2012; Santos et al. 2014). These fine-scale differences among time points, however, did not affect the overall stability of the microbiome, instead suggesting a low level of flexibility to adjust to environmental changes without affecting microbiome structure.
Previous studies on the coral microbiome suggest that changes in the abundances of certain taxa are common when corals are exposed to environmental stress (e.g. high temperature, Bourne et al. 2008; low pH, Webster et al. 2016; or pollution, Ziegler et al. 2016). During bleaching, the abundance of Endozoicomonas has been found to decrease significantly at the same time as the abundance of Vibrio increases (Ben-Haim and Rosenberg 2002; Bourne et al. 2008). Some suggest certain Vibrio species may be a causative agent of bleaching (e.g. Vibrio shiloi in Oculina patagonica, Kushmaro et al. 1996, 1997, 2001 and Vibrio corallilyticus in Pocillopora damicornis, Ben-Haim and Rosenberg 2002). Further, Bourne et al. (2008) found that the shift to a Vibrio-dominated microbiome occurred prior to visual signs of bleaching. In the present study, there was no evidence of significant increases in the abundance of Vibrio or decreases in the abundance of Endozoicomonas throughout the study period, nor was there any relationship between the abundances of these two genera. Similar stability in Endozoicomonas and Vibrio has been found previously under nutrient-induced bleaching conditions (Pogoreutz et al. 2017). Note that it is possible that the functional response of a microbial community could change without considerable variations in abundance of these taxa. For instance, recent metagenomic work has found that even with minimal changes in abundance of Vibrio species, increases in virulence genes from the Vibrio family were observed during bleaching, suggesting that the group may contribute disproportionately to coral microbiome function under temperature stress (Vega Thurber et al. 2009; Littman et al. 2011).
Host species is perhaps a key factor in maintaining a stable microbiome during periods of stress (e.g. Meistertzheim et al. 2016; Grottoli et al. 2018; Pogoreutz et al. 2018). Smith et al. (2017) found that colony morphology of P. acuta may play a role in its bleaching tolerance where the chunky morph of P. acuta was more resistant to bleaching than its fine morph counterpart. It has been well documented that skeletal morphology controls intra-colonial light amplification and scattering, where fine branching can increase irradiance (Marcelino et al. 2013). While highly beneficial for corals at depth for improving light capture, these morphologies may accelerate bleaching in shallow water corals (Marcelino et al. 2013; Swain et al. 2016). Smith et al. (2017) therefore suggest that the closer branching of the chunky morph of P. acuta reduces irradiance within the coral colony and minimises the potential for bleaching to occur, resulting in a stable Symbiodiniaceae community as seen in the present study. We hypothesise this morphology may also play a role in maintaining a stable bacterial community by reducing the stress within the colony. Future experimental work on the two morphologies of P. acuta could provide valuable insight into the drivers of both microbial community composition and coral resilience.
Microbiome stability could represent an adaptive advantage or disadvantage to the coral’s ability to respond to environmental pressures. In an experiment by Grottoli et al. (2018), two differentially tolerant coral species (one thermally sensitive and one thermally tolerant) were subjected to temperature stress. The microbiome was found to remain stable in the thermally tolerant species, and the authors suggest that corals with stable microbiomes are more likely to be resilient to adverse environmental conditions. Alternatively, if corals are affected by severe stress, the stability, or rather inflexibility, of the microbiome could facilitate a breakdown of the relationship between host and microbiome resulting in disease and/or mortality. For instance, Pogoreutz et al. (2018) exposed corals to extreme nutrient levels that resulted in tissue loss and mortality within 14 d. During this time, the microbiome remained stable through to coral death (Pogoreutz et al. 2018). Therefore, the ability of a stable microbiome to confer an adaptive advantage is likely highly dependent on both host factors and the severity of the stress event. However, given the minimal evidence available in the literature at present, it is difficult to reliably characterise the benefits and drawbacks of stability versus flexibility in the microbiome without manipulative experiments. Further, it is possible that other coral-associated microbes (e.g. fungal taxa) that were not assessed in the present study could play important roles in stress tolerance. Here, the stability of the microbiome of P. acuta may be an advantage to host resilience during the 2016 mass-bleaching event as the corals remained pigmented and there was no evidence of increases in potentially pathogenic bacterial taxa. Future experimental work exposing P. acuta to more severe stress could help identify whether this seemingly beneficial relationship between host and microbiome found in the present study is maintained or breaks down.
The present study provided a temporal view of the P. acuta microbiome in response to the 2016 thermal anomaly that resulted in mass-bleaching across the GBR. Despite significant thermal stress, the tagged colonies of P. acuta did not bleach and Symbiodiniaceae communities were stable through time. Small differences in relative abundance of individual bacterial taxa suggest some level of flexibility in the microbiome to respond to thermal stress, but these differences did not affect overall bacterial community composition throughout the sampling period. Further, expected changes in the taxa Endozoicomonas and Vibrio, often used as indicators of thermal stress, were not observed. Microbial community changes in response to thermal stress, and consequently their ability to confer host resilience, are likely reliant on host species, location, and severity of the stress event, demanding caution when making generalisations across corals as a group.
References
Andersson AF, Lindberg M, Jakobsson H, Bäckhed F, Nyrén P, Engstrand L (2008) Comparative analysis of human gut microbiota by barcoded pyrosequencing. PLoS One 3:
Apprill A, Weber LG, Santoro AE (2016) Distinguishing between Microbial Habitats Unravels Ecological Complexity in Coral Microbiomes. mSystems 1:e00143-16
Arbizu PM (2017) pairwiseAdonis: Pairwise multilevel comparison using Adonis
Arif C, Daniels C, Bayer T, Banguera-Hinestroza E, Barbrook A, Howe CJ, Lajeunesse TC, Voolstra CR (2014) Assessing Symbiodinium diversity in scleractinian corals via next-generation sequencing-based genotyping of the ITS2 rDNA region. Mol Ecol 23:4418–4433
Ben-Haim Y, Rosenberg E (2002) A novel Vibrio sp. pathogen of the coral Pocillopora damicornis. Mar Biol 141:47–55
Berkelmans R, van Oppen MJ (2006) The role of zooxanthellae in the thermal tolerance of corals: a “nugget of hope” for coral reefs in an era of climate change. Proc R Soc B Biol Sci 273:2305–2312
Boulotte NM, Dalton SJ, Carroll AG, Harrison PL, Putnam HM, Peplow LM, Oppen MJH Van (2016) Exploring the Symbiodinium rare biosphere provides evidence for symbiont switching in reef-building corals. Nature 1–9
Bourne D, Iida Y, Uthicke S, Smith-Keune C (2008) Changes in coral-associated microbial communities during a bleaching event. ISME J 2:350–363
Cai L, Zhou G, Tong H, Tian R, Zhang W, Ding W, Liu S (2018) Season structures prokaryotic partners but not algal symbionts in subtropical hard corals. Appl Microbiol Biotechnol 102:4963–4973
Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJA, Holmes SP (2016) DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods 13:581
Coenye T, Vandamme P (2003) Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ Microbiol 5:719–729
Cunning R, Silverstein RN, Baker AC (2018) Symbiont shuffling linked to differential photochemical dynamics of Symbiodinium in three Caribbean reef corals. Coral Reefs 37:145–152
De Cáceres M, Legendre P (2009) Associations between species and groups of sites: indices and statistical inference. Ecology 90:3566–3574
Fox J, Weisberg S (2011) An R companion to applied regression, second edition
Glasl B, Webster NS, Bourne DG (2017) Microbial indicators as a diagnostic tool for assessing water quality and climate stress in coral reef ecosystems. Mar Biol 164:1–18
Glynn PW (1984) Widespread Coral Mortality and the 1982–83 El Niño Warming Event. Environ Conserv 11:133
Graves S, Piepho H-P, Selzer L, Dorai-Raj S (2015) multcompView: Visualizations of paired comparisons
Grottoli AG, Martins PD, Wilkins MJ, Johnston MD, Warner ME, Cai WJ, Melman TF, Hoadley KD, Pettay DT, Levas S, Schoepf V (2018) Coral physiology and microbiome dynamics under combined warming and ocean acidification. PLoS One 13:1–22
Hadaidi G, Röthig T, Yum LK, Ziegler M, Arif C, Roder C, Burt J, Voolstra CR (2017) Stable mucus-associated bacterial communities in bleached and healthy corals of Porites lobata from the Arabian Seas. Sci Rep 7:1–11
Heron SF, Maynard JA, van Hooidonk R, Eakin CM (2016) Warming Trends and Bleaching Stress of the World’s Coral Reefs 1985–2012. Sci Rep 6:38402
Horn M, Harzenetter MD, Linner T, Schmid EN, Muller K-D, Michel R, Wagner M (2001) Members of the Cytophaga-Flavobacterium-Bacteroides phylum as intracellular bacteria of acanthamoebae: proposal of “Candidatus Amoebophilus asiaticus”. Environ Microbiol 3:440–449
Hughes TP, Kerry J, Álvarez-Noriega M, Álvarez-Romero J, Anderson K, Baird A, Babcock R, Beger M, Bellwood D, Berkelmans R, Bridge T, Butler I, Byrne M, Cantin N, Comeau S, Connolly S, Cumming G, Dalton S, Diaz-Pulido G, Eakin CM, Figueira W, Gilmour J, Harrison H, Heron S, Hoey AS, Hobbs J-P, Hoogenboom M, Kennedy E, Kuo C-Y, Lough J, Lowe R, Liu G, Malcolm McCulloch HM, McWilliam M, Pandolfi J, Pears R, Pratchett M, Schoepf V, Simpson T, Skirving W, Sommer B, Torda G, Wachenfeld D, Willis B, Wilson S (2017) Global warming and recurrent mass bleaching of corals. Nature 453:373–377
Huggett MJ, Apprill A (2018) Coral Microbiome Database: Integration of sequences reveals high diversity and relatedness of coral-associated microbes. Environ Microbiol Rep. https://doi.org/10.1111/1758-2229.12686
Jones A, Berkelmans R, van Oppen MJ, Mieog J, Sinclair W (2008) A community change in the algal endosymbionts of a scleractinian coral following a natural bleaching event: field evidence of acclimatization. Proc R Soc B Biol Sci 275:1359–1365
Klepac C, Beal J, Kenkel C, Sproles A, Polinski J, Williams M, Matz M, Voss J (2015) Seasonal stability of coral-Symbiodinium associations in the subtropical coral habitat of St. Lucie Reef. Florida. Mar Ecol Prog Ser 532:137–151
Kushmaro A, Banin E, Loya Y, Stackebrandt E, Rosenberg E (2001) Vibrio shiloi sp. nov., the causative agent of bleaching of the coral Oculina patagonica. Int J Syst Evol Microbiol 51:1383–1388
Kushmaro A, Loya Y, Fine M, Rosenberg E (1996) Bacterial infection and coral bleaching. Nature 380:396
Kushmaro A, Rosenberg E, Fine M, Loya Y (1997) Bleaching of the coral Oculina patagonica by Vibrio AK-1. Mar Ecol Prog Ser 147:159–165
LaJeunesse TC, Lee S, Bush S, Bruno JF (2005) Persistence of non-caribbean algal symbionts in Indo-Pacific mushroom corals released to Jamaica 35 years ago. Coral Reefs 24:157–159
LaJeunesse TC, Parkinson JE, Gabrielson PW, Jeong HJ, Reimer JD, Voolstra CR, Santos SR (2018) Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr Biol 1–11
Lee MD, Walworth NG, Sylvan JB, Edwards KJ, Orcutt BN (2015) Microbial communities on seafloor basalts at Dorado Outcrop reflect level of alteration and highlight global lithic clades. Front Microbiol 6:1–20
Leite DCA, Leão P, Garrido AG, Lins U, Santos HF, Pires DO, Castro CB, van Elsas JD, Zilberberg C, Rosado AS, Peixoto RS (2017) Broadcast spawning coral Mussismilia Hispida can vertically transfer its associated bacterial core. Front Microbiol 8:1–12
Lema KA, Bourne DG, Willis BL (2014) Onset and establishment of diazotrophs and other bacterial associates in the early life history stages of the coral Acropora millepora. Mol Ecol 23:4682–4695
Lema KA, Willis BL, Bourneb DG (2012) Corals form characteristic associations with symbiotic nitrogen-fixing bacteria. Appl Environ Microbiol 78:3136–3144
Lenth RV (2016) Least-Squares Means: The R Package lsmeans. J Stat Softw 69:1–33
Littman R, Willis BL, Bourne DG (2011) Metagenomic analysis of the coral holobiont during a natural bleaching event on the Great Barrier Reef. Environ Microbiol Rep 3:651–660
Liu G, Rauenzahn J, Heron S, Eakin C, Skirving W, Christensen T, Strong A, Li J (2013) NOAA Technical Report NESDIS 143 NOAA Coral Reef Watch 50 km Satellite Sea Surface Temperature-Based Decision Support System for Coral Bleaching Management. 41
Magalon H, Flot J-F, Baudry E (2007) Molecular identification of symbiotic dinoflagellates in Pacific corals in the genus Pocillopora. Coral Reefs 26:551–558
Marcelino LA, Westneat MW, Stoyneva V, Henss J, Rogers JD, Radosevich A, Turzhitsky V, Siple M, Fang A, Swain TD, Fung J, Backman V (2013) Modulation of Light-Enhancement to Symbiotic Algae by Light-Scattering in Corals and Evolutionary Trends in Bleaching. PLoS One 8:e61492
McMurdie PJ, Holmes S (2013) phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217
Meistertzheim AL, Lartaud F, Arnaud-Haond S, Kalenitchenko D, Bessalam M, Le Bris N, Galand PE (2016) Patterns of bacteria-host associations suggest different ecological strategies between two reef building cold-water coral species. Deep Res Part I Oceanogr Res Pap 114:12–22
Morrow KM, Bourne DG, Humphrey C, Botté ES, Laffy P, Zaneveld J, Uthicke S, Fabricius KE, Webster NS (2015) Natural volcanic CO2 seeps reveal future trajectories for host-microbial associations in corals and sponges. ISME J 9:894–908
Muscatine L, Porter JW (1977) Reef Corals: Mutualistic Symbioses Adapted to Nutrient-Poor Environments. Bioscience 27:454–460
Neave MJ, Apprill A, Ferrier-Pagès C, Voolstra CR (2016) Diversity and function of prevalent symbiotic marine bacteria in the genus Endozoicomonas. Appl Microbiol Biotechnol 100:8315–8324
Neave MJ, Michell CT, Apprill A, Voolstra CR (2017) Endozoicomonas genomes reveal functional adaptation and plasticity in bacterial strains symbiotically associated with diverse marine hosts. Sci Rep 1–12
Oksanen J, Blanchet G, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin P, O’Hara R, Simpson G, Solymos P, Stevens H, Szoecs E, Wagner H (2018) vegan: Community ecology package
Oliver JK, Berkelmans R, Eakin CM, Coral bleaching in space and time, (2018) In: van Oppen MJH, Lough JM (eds) Coral bleaching: Patterns, processes, causes and consequences. Springer-Verlag, Berlin, Heidelberg, pp 21–39
Olson ND, Ainsworth TD, Gates RD, Takabayashi M (2009) Diazotrophic bacteria associated with Hawaiian Montipora corals: Diversity and abundance in correlation with symbiotic dinoflagellates. J Exp Mar Bio Ecol 371:140–146
Pantos O, Bongaerts P, Dennis PG, Tyson GW, Hoegh-Guldberg O (2015) Habitat-specific environmental conditions primarily control the microbiomes of the coral Seriatopora hystrix. ISME J 9:1916–1927
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team (2018) nlme: Linear and nonlinear mixed effects models
Pochon X, Pawlowski J, Zaninetti L, Rowan R (2001) High genetic diversity and relative specificity among Symbiodinium-like endosymbiotic dinoflagellates in soritid foraminiferans. Mar Biol 139:1069–1078
Pogoreutz C, Rädecker N, Cárdenas A, Gärdes A, Meyer JL, Wild C, Voolstra CR (2017) Nitrogen fixation aligns with nifH abundance and expression in two coral trophic functional groups. Front Microbiol. 8:1187
Pogoreutz C, Rädecker N, Cárdenas A, Gärdes A, Wild C, Voolstra CR (2018) Dominance of Endozoicomonas bacteria throughout coral bleaching and mortality suggests structural inflexibility of the Pocillopora verrucosa microbiome. Ecol Evol 8:2240–2252
QIIME2 Development Team (2017) QIIME2. < URL: http://qiime2.org>
Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner FO (2013) The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res 41:590–596
R Core Team (2018) R: A language and environmental for statistical computing
Rädecker N, Pogoreutz C, Voolstra CR, Wiedenmann J, Wild C (2015) Nitrogen cycling in corals: The key to understanding holobiont functioning? Trends Microbiol 23:490–497
Raina J-B, Tapiolas D, Willis BL, Bourne DG (2009) Coral-associated bacteria and their role in the biogeochemical cycling of sulfur. Appl Environ Microbiol 75:3492–3501
Reshef L, Koren O, Loya Y, Zilber-Rosenberg I, Rosenberg E (2006) The Coral Probiotic Hypothesis. Environ Microbiol 8:2068–2073
Ritchie KB (2006) Regulation of microbial population by coral surface mucus and mucus-associated bacteria. Mar Ecol Prog Ser 322:1–14
Roder C, Bayer T, Aranda M, Kruse M, Voolstra CR (2015) Microbiome structure of the fungid coral Ctenactis echinata aligns with environmental differences. Mol Ecol 3501–3511
Rognes T, Flouri T, Nichols B, Quince C, Mahé F (2016) VSEARCH: a versatile open source tool for metagenomics. PeerJ Prepr 4:e2409v1
Rosenberg E, Koren O, Reshef L, Efrony R, Zilber-Rosenberg I (2007) The role of microorganisms in coral health, disease and evolution. Nat Rev Microbiol 5:355–362
Sampayo EM, Dove S, LaJeunesse TC (2009) Cohesive molecular genetic data delineate species diversity in the dinoflagellate genus Symbiodinium. Mol Ecol 18:500–519
Santos HF, Carmo FL, Duarte G, Dini-Andreote F, Castro CB, Rosado AS, van Elsas JD, Peixoto RS (2014) Climate change affects key nitrogen-fixing bacterial populations on coral reefs. ISME J 8:2272–2279
Schmitz-Esser S, Tischler P, Arnold R, Montanaro J, Wagner M, Rattei T, Horn M (2010) The genome of the amoeba symbiont “Candidatus Amoebophilus asiaticus” reveals common mechanisms for host cell interaction among amoeba-associated bacteria. J Bacteriol 192:1045–1057
Smith H, Epstein H, Torda G (2017) The molecular basis of differential morphology and bleaching thresholds in two morphs of the coral Pocillopora acuta. Sci Rep 7:1–12
Stat M, Pochon X, Cowie ROM, Gates RD (2009a) Specificity in communities of Symbiodinium in corals from Johnston Atoll. Mar Ecol Prog Ser 386:83–96
Stat M, Loh WKW, LaJeunesse TC, Hoegh-Guldberg O, Carter DA (2009b) Stability of coral–endosymbiont associations during and after a thermal stress event in the southern Great Barrier Reef. Coral Reefs 28:709–713
Swain TD, DuBois E, Gomes A, Stoyneva VP, Radosevich AJ, Henss J, Wagner ME, Derbas J, Grooms HW, Velazquez EM, Traub J, Kennedy BJ, Grigorescu AA, Westneat MW, Sanborn K, Levine S, Schick M, Parsons G, Biggs BC, Rogers JD, Backman V, Marcelino LA (2016) Skeletal light-scattering accelerates bleaching response in reef-building corals. BMC Ecol 16:15–19
Thornhill DJ, Fitt WK, Schmidt GW (2006a) Highly stable symbioses among western Atlantic brooding corals. Coral Reefs 25:515–519
Thornhill DJ, Fitt WK, Schmidt GW (2006b) Highly stable symbioses among western Atlantic brooding corals. Coral Reefs 25:515–519
Tonk L, Bongaerts P, Sampayo EM, Hoegh-Guldberg O (2013) SymbioGBR: a web-based database of Symbiodinium associated with cnidarian hosts on the Great Barrier Reef. BMC Ecol 13:7
Torda G, Donelson JM, Aranda M, Barshis DJ, Bay L, Berumen ML, Bourne DG, Cantin N, Foret S, Matz M, Miller DJ, Moya A, Putnam HM, Ravasi T, Van Oppen MJH, Thurber RV, Vidal-Dupiol J, Voolstra CR, Watson SA, Whitelaw E, Willis BL, Munday PL (2017) Rapid adaptive responses to climate change in corals. Nat Clim Chang 7:627–636
Valliappan K, Sun W, Li Z (2014) Marine actinobacteria associated with marine organisms and their potentials in producing pharmaceutical natural products. Appl Microbiol Biotechnol 98:7365–7377
van Oppen MJH, Oliver JK, Putnam HM, Gates RD (2015) Building coral reef resilience through assisted evolution. Proc Natl Acad Sci 1–7
Vega Thurber R, Willner-Hall D, Rodriguez-Mueller B, Desnues C, Edwards RA, Angly F, Dinsdale E, Kelly L, Rohwer F (2009) Metagenomic analysis of stressed coral holobionts. Environ Microbiol 11:2148–2163
Webster NS, Negri AP, Botté ES, Laffy P, Flores F, Noonan S, Schmidt C, Uthicke S (2016) Host-associated coral reef microbes respond to the cumulative pressures of ocean warming and ocean acidification. Sci Rep 19324
Wickham H (2009) ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag, New York
Wilson K, Li Y, Whan V, Lehnert S, Byrne K, Moore S, Ballment E, Fayazi Z, Swan J, Kenway M, Benzie J (2002) Genetic mapping of the black tiger shrimp Penaeus monodon with amplified fragment length polymorphism. Aquaculture 204:297–309
Ziegler M, Roik A, Porter A, Zubier K, Mudarris MS, Ormond R, Voolstra CR (2016) Coral microbial community dynamics in response to anthropogenic impacts near a major city in the central Red Sea. Mar Pollut Bull 105:629–640
Acknowledgements
We would like to thank the Coral Health Group at the Australian Institute of Marine Science for initial coral tagging, and K. Damjanovic, A. Hernandez-Agreda and H. Smith for statistical advice. This study was funded by Paul G. Allen Philanthropies and AIMS@JCU, the joint venture between James Cook University and the Australian Institute of Marine Science, as part of H.E.E’s PhD research. H.E.E acknowledges receipt of an AIMS@JCU Postgraduate Scholarship.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Topic Editor Morgan S. Pratchett
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Epstein, H.E., Torda, G. & van Oppen, M.J.H. Relative stability of the Pocillopora acuta microbiome throughout a thermal stress event. Coral Reefs 38, 373–386 (2019). https://doi.org/10.1007/s00338-019-01783-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-019-01783-y