
Abstract
Seabird species that breed on remote islands in the southern and northern hemispheres are occasionally seen in the continental shelf waters of South Africa. Most are only seen at sea; however, weak, oiled, or injured individuals found on land or on fishing vessels are occasionally admitted to rehabilitation centres. From 1993 to 2017 (25 years), the Southern African Foundation for the Conservation of Coastal Birds (SANCCOB) admitted 297 southern and northern hemisphere remotely-breeding seabirds from 35 species. This represents an average of 12 birds per year, ranging from 0 to 32. The most frequently recorded families were: Procellariidae (198 individuals, 67%), Spheniscidae (29 individuals, 10%), Stercorariidae (22 individuals, 7%), and Diomedeidae (21 individuals, 7%). The three most common species corresponded to the largest Procellariidae species: southern giant petrel (Macronectes giganteus, 49 individuals, 16%), northern giant petrel (Macronectes halli, 34 individuals, 11%), and white-chinned petrel (Procellaria aequinoctialis, 34 individuals, 11%). The majority of birds were admitted due to debilitation (61%) or injury (21%). Of the 185 birds for which the outcome of rehabilitation was recorded, 39% survived to be released back into the wild.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Nearly, half of all seabird species are experiencing population declines (Croxall et al. 2012), and the world’s monitored population of seabirds has declined by 70% from 1950 to 2010 (Paleczny et al. 2015). The population declines are more pronounced in families of wide-ranging, pelagic species, suggesting that pan-global populations may be more at risk than shorter-ranging, coastal populations (Croxall et al. 2012; Paleczny et al. 2015). Pelagic species tend to nest on oceanic islands, and their populations are often difficult to monitor consistently, so that threats to these species might only be detected once population numbers have reached dramatic lows (Oppel et al. 2022). Information on the occurrence, abundance, and health status of these pelagic seabirds throughout their non-breeding areas can therefore provide clues to identify their conservation threats, and improve their protection.
At the breeding grounds, the main threats to seabird populations comprise of invasive species (e.g. rats, cats), disturbance, direct exploitation (of eggs, birds, and guano), and habitat degradation (Rand 1954; Nogales et al. 2004; Russell 2011; Croxall et al. 2012; Dilley et al. 2017; Dias et al. 2019). However, in broader terms, the population declines in seabird species are largely related to human impacts throughout their non-breeding distribution, such as entanglement with fishing gear, overfishing, climate change, marine pollution, and wind energy production (Votier et al. 2005; Croxall et al. 2012; Maree et al. 2014; Trathan et al. 2015; Crawford et al 2017; Dias et al. 2019).
Some conservation efforts have successfully reduced seabird mortality (Paleczny et al. 2015). In southern Africa, BirdLife South Africa's Albatross Task Force has had great success by introducing mitigation methods to reduce albatross deaths by 95% in demersal trawl fisheries (Maree et al. 2014). The establishment of marine protected areas and fishery closures have also been sought in order to benefit seabird conservation in the region (Pichegru et al. 2009, 2012; Sherley et al. 2018). The Southern African Foundation for the Conservation of Coastal Birds (SANCCOB) has contributed to reduced mortality of seabirds caused by oil spills, trauma, and nest desertion, especially for locally-breeding species such as the African penguin (Spheniscus demersus) by rehabilitating and releasing birds back into the wild (Parsons and Underhill 2005; Klusener et al. 2018; Parsons et al. 2018).
Regular observations at sea of birds in South African waters were conducted in the 1980s and showed that many of the species breeding on remote Subantarctic islands are regularly seen in the region (Ryan and Moloney 1988; Ryan et al. 1989). Regular beach patrols were conducted on the coast of Namibia and the western coast of South Africa in the late 1970s and early to mid 1980s, providing a wealth of information on the seabirds that occur in the region (Avery 1980, 1981, 1982, 1984, 1989). Unfortunately these beach patrols are no longer conducted and majority of stranding’s data available are from birds admitted to rehabilitation facilities. Since 2009, BirdLife South Africa has collated at-sea observations in their Atlas of Seabirds at Sea (AS@S) database (http://seabirds.saeon.ac.za/smap.aspx.) In the absence of current coordinated beach patrols, data from rehabilitation admissions can also provide information on some of the threats faced by these birds in South African waters. In this study, we present data on visitor and vagrant seabird species admitted at SANCCOB’s rehabilitation facility in Cape Town from 1993 to 2017 (25 years).
Methods
SANCCOB is a non-profit organization that rehabilitates oiled, sick, injured, and abandoned marine and coastal birds rescued by local authorities and the general public. Given the lack of information of carcasses admitted to the facility this study explicitly considers the admission and rehabilitation records of only live pelagic seabirds that do not breed on mainland South Africa that were admitted to the Cape Town facility from 1993 to 2017 (inclusive). Lastly this study did not include any seabirds affected by widespread oiling events during this period.
The following variables were recorded for each individual: species, date of admission, age group, location of rescue, reason for admission, outcome, and date of outcome. Unfortunately, some of the rehabilitation records were incomplete (especially prior to 2001), and therefore, some of the analyses had relatively small sample sizes. Furthermore, some individuals could not be identified to the species level because their plumage was immature, oiled, or damaged, or they were not considered a separate species at the time (e.g.northern and southern rockhopper penguins).
Species were classified in relation to their taxonomic family, their status in southern Africa (common visitor, vagrant, rare visitor), and their origin in relation to South Africa (North, South) based on Ryan (2017). The conservation status of each species (Least Concern, Near Threatened, Vulnerable, Endangered) was obtained from the IUCN (2022). Species were classified based on their body size (very small < 120 g, small 120 g to 1 kg, medium 1 to 2 kg, large > 2 kg; derived from average body mass given by Ryan (2017)) and their main prey (cephalopod, crustacean, fish, other; derived from Schramm 1986; Shealer 2001; Connan et al. 2014; Ryan 2017).
The season of admission was categorized as follows: spring (September equinox–December solstice), summer (December solstice–March equinox), autumn (March equinox–June solstice), and winter (June solstice–September equinox). Location of admission was categorized into seven regions using select landmarks along the South African coast (Fig. 1; following Parsons et al. 2018). SANCCOB has first responders based all over the South African coastline and will routinely rescue and stabilize birds until they are transported to the Cape Town facility therefore this study could include birds rescued from regions outside of the Western Cape of South Africa. A separate category was also established for birds brought to the facility from oceanic fishing vessels. The age group (juvenile, adult) of each bird was classified upon admission based on their plumage (Ryan 2017). Reason for admission was classified into five categories (following Parsons et al. 2018): oiling, debilitation, injury, molt (penguins), and other. The ‘debilitation’ category comprised of individuals that were prostrate, lethargic, dehydrated, emaciated, or otherwise weakened but not exhibiting signs of molt, external injury, or oiling. The ‘other’ category included miscellaneous uncommon reasons for admittance (1 case each): respiratory distress, neurological signs, peritonitis, and a healthy individual found on a boat. It is believed that this individual might have just been tired and when landing on deck of the vessel was kept on board and brought to SANCCOB on arrival to the harbour.

Map of the study area and select localities. a African continent and oceanic islands and archipelagos of the Atlantic and Indian Oceans from where the vagrant and visitor seabird species in South Africa may have originated and b Southern African Foundation for the Conservation of Coastal Birds (SANCCOB) and landmarks used to subdivide the South African coast
The outcome was classified as ‘released’ or ‘not released’, the latter indicating that individuals had died in captivity, were euthanized, or were kept permanently captive because they were considered unfit for release. The release rate was calculated as the number of birds released during the period of 1993–2017 divided by the number of birds for which the outcome was known. The rehabilitation period (only for released birds), in days, was calculated as the interval between admission to the facility and release dates. Seasonal distribution was evaluated only for species for which the month of admission had been recorded for at least 10 individuals.
The Atlas of Seabirds at Sea (AS@S) is an open-access database that compiles at-sea observations by seabird experts and volunteers, collected according to a standard protocol and submitted via a website (http://seabirds.saeon.ac.za/smap.aspx.). As of 10 October 2018, AS@S had data from 88 observers on 111 cruises, resulting in 17,597 data transects and 112,485 bird sightings; the vast majority of the information was obtained on vessels traveling to and from South Africa (data contributed on a voluntary basis since 2009). The total number of AS@S records was obtained for each species recorded in this study. Additionally, we used QGIS 3.8.3 (https://qgis.org/) to obtain the number of AS@S records that were within South African waters (i.e. 200 nm from the South African coastline, excluding the Prince Edward Islands).
A stepwise binary logistic regression was used to evaluate which species and individual parameters (species family, species’ breeding distribution(according to Birdlife International (2022)), species conservation status, species visitor status, species size, species’ main prey item, age at admission, region of capture, reason for admission, year of admission and season of admission could best predict the outcome (released vs. not released) based on a subset of 157 individuals for which the outcome had been recorded. Significance level for inclusion or exclusion in the model was 0.15. Lastly, linear regression analysis was used to evaluate if there was a correlation between the number of recorded individuals for each species in this study and the number of AS@S records.
Results
A total of 297 pelagic seabirds of 35 species were admitted for rehabilitation from 1993 to 2017, averaging 12 ± 9 individuals per year (mean ± SD; range: 0–32) (Table 1). The most frequently recorded families were: Procellariidae (198 individuals, 67%), Spheniscidae (29 individuals, 10%), Stercorariidae (22 individuals, 7%), and Diomedeidae (21 individuals, 7%). Online Resource 1 provides an overview of the breeding distribution of these seabird species relative to the oceanic islands shown in Fig. 1.
For individuals whose age group was recorded, juveniles (98 individuals, 59%) were more frequently admitted than adults (67 individuals, 41%). For individuals whose location of collection was recorded, the majority were found along the coast from Lambert’s Bay to Cape Point (75 individuals, 45%) and from Cape Point to Cape Agulhas (77 individuals, 46%), with fewer individuals gathered in the other regions: Cape Agulhas to Mossel Bay (2 individuals), Cape St. Francis to Port Alfred (4), and East of Port Alfred (4). No individuals were found north of Lambert’s Bay or between Mossel Bay and Cape St. Francis. An additional six individuals were rescued at sea and brought ashore by fishing vessels. The annual and monthly distributions of the admissions are presented in Fig. 2 and 3. The reasons for admission were: debilitation (113 individuals, 61%), injury (39 individuals, 21%), oiling (19 individuals, 10%), molt (10 individuals, 5%), and other (4 individuals, 2%) (Table 2); the reason for admission was not recorded for 112 individuals.

Annual number of visitor and vagrant seabirds admitted for rehabilitation

Monthly number of visitor and vagrant seabirds most frequently admitted for rehabilitation
Thirty-nine percent of the individuals with a recorded rehabilitation outcome were successfully released. The average release rate varied considerably among the reasons for admission: debilitation (release rate = 42%), injury (31%), molt (40%), oiling (89%), and other (25%). Binary logistic regression analysis (AIC = 198.79, r2 = 0.121) revealed that reason for admission (p = 0.002) and species breeding distribution (p = 0.035) were the most significant factors determining whether a bird was released, whereas the year of admission (p = 0.051) and age at admission (p = 0.093) approached significance and were also included in the final model. Birds admitted due to 'injury' (z = − 3.11), 'debilitation' (z = − 2.76) and 'molt' (z = − 2.68) were less likely to be released than those admitted due to oiling. Birds belonging to species originating from the North (in relation to South Africa) (z = − 1.93) had a lower probability of being released than those belonging to species originating from the South. Year of admission had a negative effect (z = − 1.93); birds admitted in recent years had a lower probability of being released than those admitted in the earlier years of the study period. Juvenile birds (z = − 1.68) were less likely to be released than adult birds. The remaining variables were not included in the final model due to lack of statistical significance or to quasi-separation of data points.
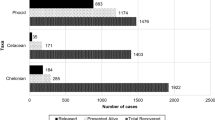
There was sufficient data to calculate the rehabilitation period (the interval from admission to release) for 80 individuals. For all species combined, the rehabilitation period was 14 ± 17 days (mean ± SD) (n = 80), ranging from 2 to 147 days (see Table 2). Data on the rehabilitation outcome and the outcome date (date released, euthanised or died) were available for a subset of individuals depending on the reason for their admission: debilitation (98 individuals), injury (36), oiling (17), molt (9), and other (3). The distribution of these outcomes in relation to the date of admission is presented in Fig. 4; the ‘other’ category was omitted due to the small sample size.

Rehabilitation outcomes of visitor and vagrant seabirds during the first 30 days following admission, in relation to the reason for admission. a Debilitation, b Injury, c Oiling and d Molt
There was sufficient temporal overlap between this study and the AS@S as well as the area covered. Since the AS@S records was broadly distributed in the South African Exclusive Economic Zone (EEZ) therefore matching to a reasonable extent with the data from this study. Therefore for each species, we could compare the number of individuals recorded in this study with the number of records on the AS@S database (Online Resource 2). A very weak correlation existed between the number of individuals recorded in this study and the total number of AS@S records (r2 = 0.010), the number of AS@S records within SA waters (r2 = 0.008), and the proportion of AS@S records in SA waters (r2 = 0.022) (Online Resource 2).
Discussion
Species composition
Although there are differences in the period as well as methodologies used between this study, Avery’s beach surveys (1980’s) and coastal cruises (2008–2018) they can still add value in the absence of more recent coordinated beach cast surveys, but care should still be taken when comparing these different data sets. The species composition of live seabirds admitted to SANCCOB was generally consistent with that observed during coastal cruises from 2009 and beach surveys along the western coast of South Africa in the 1980s; however, there are some relevant quantitative differences. The greatest discrepancy was noted for prions (Pachyptila spp.), which made up 41% of the beach-cast seabirds recorded by Avery (1980, 1981, 1982, 1989), averaging 130 individuals per year, whereas only three individuals (1%; 0.12 individuals per year) were recorded in this study. Other small Procellariidae were also less common in this study; for example sooty shearwaters (Ardenna grisea) were more frequent in Avery’s reports (19%; 61 individuals per year) than in this study (6%; 1 individual per year). In contrast, the medium-sized white-chinned petrel (Procellaria aequinoctialis) occurred in relatively similar proportions in our study and in Avery’s reports (respectively, 11% and 16%), whereas the larger giant petrels (Macronectes spp.) were the most frequently recorded species in this study (32%) but were comparatively uncommon in Avery’s surveys (1%). These findings suggest that our sampling strategy, which relies on the public and local authorities bringing birds requiring rehabilitation to SANCCOB, has a tendency to produce a significant bias in the observed species composition, as larger species are more likely to be rescued and recorded. An additional factor may be that larger birds have a lower surface-to-volume ratio, rendering them less susceptible to hypothermia (Nuckton et al. 2002). Additionally, their greater size might protect them from smaller predators. As a result, debilitated individuals of larger species may have a greater likelihood of surviving to be rescued. It should be noted that although Avery (1980, 1981, 1982, 1984, 1989) did not report visitor or vagrant penguins, other publications provided records of these species in South Africa during the 1980s (Cooper 1992a; Parsons et al. 2017).
Increase in admissions
The sudden increase in the number of visitor and vagrant seabirds admitted to SANCCOB in 2001 likely does not reflect an actual abrupt change in the presence of these species on the South African coast, but rather a shift in the probability of stranded seabirds being rescued, brought to, and recorded by SANCCOB. The MS Treasure oil spill occurred in June 2000 when approximately 19,000 oiled African penguins were admitted for rehabilitation (Crawford et al. 2000). This led to increased public awareness of SANCCOB’s efforts to rescue and rehabilitate seabirds in the region, and also led to the adoption of stricter record keeping procedures. From 2001 to 2017, SANCCOB’s operations were relatively stable (no single events with admissions of > 1,000 birds) and fluctuations in the number of birds admitted were more likely to represent regional ecological dynamics. Although the proportions of different seabird families relatively consistent during this period, the total number of individuals admitted varied substantially, ranging from seven individuals each in 2014 and 2016, to 32 individuals in 2002 (Fig. 2). Previous studies reported increased numbers of seabirds stranded and sighted at sea along the South African coast following the strong 1982/1983 El Niño (La Cock 1986; Avery 1989; Ryan et al. 1989). From 2001 to 2017, there were two moderate El Niño events (2002/03 and 2009/10), a moderate La Niña event (2011/12), and a strong La Niña event (2010/11) (Null 2019). During the years of these El Niño and La Niña events, visitor or vagrant seabirds occurred relatively more frequently than in average years corroborating that the El Niño Southern Oscillation (ENSO) plays a role in determining the number of seabirds that strand on the South African coast (Fig. 2). However, because it is known that the populations of seabirds may present distinct responses to ENSO (Duffy 1990; Crawford et al. 2003), future studies will be necessary to clarify how different species are affected by these fluctuations off the southern African coast relative to their feeding preferences and foraging strategies.
Reason for admission
Debilitation was the most frequent reason for admission (61%), and it also had a significant negative effect on an individual’s likelihood to survive to be released. The amount of tissue damage caused by malnutrition (depending on the severity and duration) may accumulate to irreversible levels and continue to manifest beyond the first days of rehabilitation (Parsons et al. 2018), explaining the gradual ascent in the mortality rate (Fig. 4a) and the poor release rate (42%) observed in debilitated birds. Hypothermia and starvation are frequent causes of death of stranded seabirds, and may result from decreased prey availability due to overfishing or oceanographic and climatic fluctuations (La Cock 1986; Cury et al. 2011; Tavares et al. 2016). Furthermore, seabirds stranded on the southern African coast due to winter storms are often in poor body condition, which suggests they were already struggling to obtain food (Avery 1980; Ryan et al 1989). Overfishing and climate change have markedly decreased prey availability and distribution along the South African coast in recent decades, with profound impacts on the local seabird community (Crawford and Dyer 1995; Crawford et al. 2008, 2011). Further studies will be needed to evaluate if and how visitor and vagrant seabirds are also affected.
Injury was the second most frequent reason for admission (21%) and had the strongest negative effect on an individual’s likelihood of being released. Potential causes of injury include predation, collision (e.g. watercrafts and buildings) and entanglement in fishing gear or marine debris (Hocken 2000, 2005; Cannell et al. 2016). Seabirds admitted due to injuries or trauma were often euthanized due to the poor prognosis, which explains their poor release rate (31%) and the high proportion of birds dying within the first three days (Fig. 4b).
Molt was the most frequent reason for admission for penguins (63%). Unlike flying birds that molt their plumage progressively, penguins undergo a catastrophic molt process in which they regrow their entire plumage at once. This is a physiologically and energetically demanding process that places a critical constrain on the life cycle of these birds (Adams and Brown 1990; Kemper et al. 2008). A penguin’s inability to gain weight before molting may result in an arrested molt, where the molt is incomplete and the plumage lacks adequate waterproofing (Cooper 1978; Webster et al. 2016; Parsons et al. 2018). Recovering from arrested molt requires an extended stay at the rehabilitation center and the penguins may develop captivity-associated health problems (e.g. aspergillosis, pododermatitis), which explains the poor release rate (40%) and delayed mortality (Fig. 4c) observed in this group.
Oiling was the reason for admission of 19 (10%) birds in this study. It is worth noting that six of these oiled birds (32%) were pintado petrels brought in by a fishing vessel on 17 June 2010, presumably due to a unidentified oil spill (i.e. an oil spill that was not reported to authorities and for which the source is not known). The swift rehabilitation (Fig. 4d) and high release rate (89%) are consistent with the good body condition and health status that is usually displayed by oiled seabirds admitted to SANCCOB (Parsons and Underhill 2005; Parsons et al. 2018). Furthermore, Vanstreels et al. (2023) also found that the timing of rescue in oiled African penguins played a vital role in the general health status and higher rehabilitation success of birds admitted earlier after an oil spill.
Location of rescue
The majority (90%) of seabirds in this study was rescued from Lambert’s Bay to Cape Agulhas. This is likely related to the high biological productivity associated with the Benguela upwelling system (Shannon 1985; Brown 1992; Phillips et al. 2005), but may also be due to the higher probability that stranded seabirds will be detected, rescued, and brought in for rehabilitation due to the higher human population density in the region, and the proximity to SANCCOB’s facility.
Most species of seabirds are subjected to stranding, but the degree to which different species are likely to be found and rescued varies (Camphuysen et al. 1999). For instance, swimming and diving seabirds such as penguins are highly vulnerable to oiling as traveling from colony to foraging grounds means that both the area covered and time spent in water are going to be greater, with both increasing the likelihood of being oiled. (Williams et al. 1994; Camphuysen et al. 1999; García-Borboroglu et al. 2006), whereas small flying species, such as storm petrels, shearwaters, and fulmars, may be more susceptible to storms (Pashby and Cudworth 1969; Threlfall et al. 1974; Miskelly et al. 2011; Tavares et al. 2016). Other factors, such as prey availability and distribution, interaction with fisheries, and prevailing wind speed and direction, are also likely to determine the stranding of seabirds (Camphuysen et al. 1999; Tavares et al. 2016).
Seasonality
The Benguela upwelling has a marked seasonality; the South African coast experiences a peak in primary production during the austral spring and summer, while the primary production in other regions is almost entirely restricted to the spring (Brown 1992). There was marked seasonal variation in the number of visitor and vagrant seabirds admitted to SANCCOB (Fig. 3); however, there was no clear pattern that could be unequivocally attributed to the seasonality of the Benguela upwelling system. Rockhopper penguins (Eudyptes spp.) were most frequently admitted during summer months, which is consistent with previous studies in the region (Cooper 1978; 1992b) and due to mostly molting individuals. In contrast, pintado petrels (Daption capense) and brown skuas (Stercorarius antarcticus) were predominantly admitted during winter months, suggesting they were overwintering in South African waters. The admissions of sooty shearwaters, white-chinned petrels, and giant petrels were relatively evenly distributed over the year, presumably because these are predominantly non-breeding individuals that do not have to return to their colonies for the breeding season.
Age group
With regards to the age groups, juvenile birds were more frequently admitted than adults, and also tended to experience a marginally lower release rate than adults, suggesting a poorer health status when admitted. Since fledgling/juvenile birds are less experienced than adults in locating productive feeding grounds and capturing prey, wrecks of young seabirds are not uncommon (Avery 1989). Also, juvenile individuals may be attempting to disperse beyond their natal breeding site, or simply take advantage of not having to return to oceanic islands to breed, and are therefore able to spend a greater proportion of time foraging off southern Africa.
Outcome
The average release rate was 39%, contrasting with the 83% rate for African penguins at the same facility (Parsons et al. 2018). However, when the reason for admission is considered, it becomes clear that the release rates of visitor and vagrant seabirds are generally comparable to those of African penguins (53% release rate for debilitation, 60% for injury, 55% for molt, and 93% for oiling; Parsons et al. 2018). It is therefore clear that the poorer release rates observed in this study are largely related to the different reasons for admission. On the other hand, the rehabilitation period in this study was relatively short, with an average of 14 days compared to the average of 30 days for African penguins at the same facility (Parsons et al. 2018). This is likely due to the smaller body size of the species in this study, which require a shorter period in order to gain weight and achieve an adequate body condition to be released. It is also worth noting that these species are usually released sooner in order to improve their chances of survival as they are more prone to health issues, such as respiratory infections and pododermatitis.
At sea observations
We did not find a significant correlation between the results of this study and the AS@S database, not even when we only considered the records made within South African waters. This disagreement might be related to a number of factors: (a) species with small body size and subtle differences in morphological characteristics, such as prions and shearwaters, may be less likely to be detected and identified at sea, and would therefore tend to be underestimated by AS@S; (b) the trajectories of cruises upon which the AS@S database relies are determined for other reasons (e.g. operational needs of the South African National Antarctic Programme), and this may lead to a bias in the geographical distribution of the sampling effort that unevenly represents different species depending on their habitat use; and (c) depending on their body size, physiological and ecological characteristics, different species visiting South African waters are likely to experience dissimilar odds of becoming beach-cast, rescued and brought to the rehabilitation center. It is therefore clear that the two approaches (at-sea surveys and recording of beach-cast individuals) are complementary in the development of knowledge about the diversity and density of seabirds that occur in coastal waters.
Species origin
South Africa’s coast lies in the subtropical zone. Therefore, it would be reasonable to expect the presence of vagrant and visitor seabirds from both tropical and temperate regions. However, the findings of this study agree with previous data showing that the majority of the vagrant and visitor seabirds that come to South African waters originate from temperate, not tropical regions (Liversidge 1959; Morant et al. 1983; Ryan and Moloney 1988; Ryan et al. 1989). The species recorded in this study can be broadly classified as:group a: species that breed in the northern temperate zone (north of 35°N), including four species Corry’s shearwater Calonectris borealis, European storm petrel Hydrobates pelagicus, arctic tern Sterna paradisaea and parasitic jaeger Stercorarius parasiticus); group b: species that breed in the tropical and subtropical zones (35°N to 35°S)(two species: red-billed tropic bird Phaethon aethereus and red-footed booby Sula sula); and group c: species that breed in the southern temperate zone (south of 35°S) (29 species, see Table 1). Interestingly, birds originating from the north (i.e. groups a and b) were less likely to be released, which may suggest a poorer health status when admitted.
Additionally, it is worth noting that the vast majority of group c species breed on oceanic islands in the southwest Indian (Prince Edward, Crozet and Kerguelen islands) or southeast Atlantic oceans (Tristan da Cunha and Gough islands) (Online Resource 1). Based on large-scale ring recovery studies conducted from the 1950s to the 1980s, these island groups are indeed the main origin of seabirds visiting the South African coast, along with a substantial number of individuals from South Georgia and the South Orkney Islands (Morant et al. 1983). However, there were two noteworthy exceptions in our study: shy albatrosses (Thalassarche cauta), which breed on three small islands off the coast of Tasmania, Australia, and sooty shearwaters, which breed on islands off New Zealand, Australia and Chile, and on the Falkland/Malvinas Islands (Ryan 2017).
Conclusion
Lastly, it is worth noting that there is an on-going debate on whether Antarctic and Subantarctic species that stray to southern continents should be released back to the wild. In 1996, the Scientific Committee on Antarctic Research (SCAR) issued a recommendation that the re-introduction of indigenous seals and seabirds that have been held in captivity to subantarctic islands and the Antarctic continent should be discouraged out of concern that it would risk introducing pathogens to native populations (SCAR Recommendation XXIV-3). However, the SCAR secretariat later clarified that SCAR Recommendation XXIV-3 only referred to the transport and release of rehabilitated wildlife within the SCAR area of interest (Antarctica, its offshore islands, and the surrounding Southern Ocean), and was not intended to apply to releases at other continents (Agreement on the Conservation of Albatrosses and Petrels 2014). In South Africa, the decision of whether or not visitor and vagrant Antarctic species (including seabirds) should be released, and what disease screening and release criteria should be employed, are still being discussed with various stakeholders. Comprehensive disease risk assessments have yet to be conducted both at national and international levels to adequately evaluate the different pathogens, host species potentially involved, and their epidemiology and ecology in order to reach a clearer set of criteria and recommendations.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Avery G (1980) Results of beach patrols conducted in southern Africa in 1979. Cormorant 8:65–72
Avery G (1981) Results of beach patrols conducted in southern Africa in 1980. Cormorant 9:113–122
Avery G (1982) Results of beach patrols conducted in southern Africa in 1981. Cormorant 10:87–96
Avery G (1984) Results of beach patrols conducted in southern Africa in 1982. Cormorant 12:29–43
Avery G (1989) Results of patrols for beached seabirds conducted in southern Africa in 1984 and 1985. Cormorant 17:57–71
Adams NJ, Brown CR (1990) Energetics of molt in penguins. In: Davis LS, Darby JT (eds) Penguin biology. Academic Press, San Diego, pp 297–315
Agreement on the Conservation of Albatrosses and Petrels (2014) Report of the Eighth Meeting of the Advisory Committee, Punta del Este, Uruguay, 15–19 September 2014
Brown P (1992) Spatial and seasonal variation in chlorophyll distribution in the upper 30 m of the photic zone in the southern Benguela/Agulhas ecosystem. Afr J Mar Sci 12:515–525. https://doi.org/10.2989/02577619209504722
BirdLife International (2022) The IUCN Red List of Threatened Species. Version 2022-2. https://www.iucnredlist.org. Accessed 11 Nov 2022
Cooper J (1978) Results of beach patrols conducted in 1977. Cormorant 4:4–9
Cooper J (1992a) First record of the Gentoo Penguin Pygoscelis papua for Africa. Mar Ornithol 20:80–81
Cooper J (1992b) Most northerly record of the Rockhopper Penguin Eudyptes chrysocome for Africa. Mar Ornithol 20:82–83
Crawford RJM, Dyer BM (1995) Responses by four seabirds to a fluctuating availability of Cape Anchovy Engraulis capensis off South Africa. Ibis 137:329–339. https://doi.org/10.1111/j.1474-919X.1995.tb08029.x
Camphuysen CJ, Wright PJ, Leopold M, Hüppop O, Reid JB (1999) A review of the causes, and consequences at the population level, of mass mortalities of seabirds. ICES Cooperative Research Reports 232:51–66
Crawford RJM, Davis SA, Harding RT, Jackson LF, Leshoro TM, Meyer MA, Randal RM, Underhill LG, Upfold L, Van Daslen AP et al (2000) Initial impact of the treasure oil spill on seabirds off western South Africa. Afr J Mar Sci 22:157–176. https://doi.org/10.2989/025776100784125645
Crawford RJM, Rae CMD, Nel DC, Cooper J (2003) Unusual breeding by seabirds at Marion Island during 1997/98. Afr J Mar Sci 25:453–462
Crawford RJM, Sabarros P, Fairweather T, Underhill LG, Wolfaardt A (2008) Implications for seabirds off South Africa of a long-term change in the distribution of sardine. Afr J Mar Sci 30:177–184. https://doi.org/10.2989/AJMS.2008.30.1.18.468
Cury PM, Boyd IL, Bonhommeau S, Anker-Nilson T, Crawford RJM, Furness RW, Mills JA, Murphy EJ, Osterblom H, Paleczny M, Piatt JF, Roux JP, Shannon L, Sydeman WJ (2011) Global seabird response to forage fish depletion: One-third for the birds. Sci 334:1703–1706
Crawford RJM, Altwegg R, Barham BJ, Barham PJ, Durant JM, Dyer BM, Geldenhuys D, Makhado AB, Pichegru L, Ryan PG et al (2011) Collapse of South Africa’s penguins in the early 21st century. Afr J Mar Sci 33:139–156. https://doi.org/10.2989/1814232X.2011.572377
Croxall JP, Stuart H, Butchart M, Lascelles B, Stattersfield AJ, Sullivan B, Symes A, Taylor P (2012) Seabird conservation status, threats and priority actions: a global assessment. Bird Conserv Int 22:1–34. https://doi.org/10.1017/S0959270912000020
Connan M, McQuaid C, Bonnevie B, Smale M, Cherel Y (2014) Combined stomach content, lipid and stable isotope analyses reveal spatial and trophic partitioning among three sympatric albatrosses from the Southern Ocean. Mar Ecol Prog Ser 497:259–272. https://doi.org/10.3354/meps10606
Cannell BL, Campbell K, Fitzgerald L, Lewis JA, Baran IJ, Stephens NS (2016) Anthropogenic trauma is the most prevalent cause of mortality in Little Penguins (Eudyptula minor) in Perth, Western Australia. Emu 116:52–61. https://doi.org/10.1071/MU15039
Crawford R, Ellenberg U, Frere E, Hagen C, Baird K, Brewin P, Crofts S, Glass J, Mattern T, Pompert J, Ross K, Kemper J, Ludynia K, Sherley R, Steinfurth A, Suazo C, Yorio P, Tamini L, Mangel J, Bugoni L, Jiménez Uzcátegui G, Simeone A, Luna-Jorquera G, Gandini P, Woehler E, Pütz K, Dann P, Chiaradia A, Small C (2017) Tangled and drowned: a global review of penguin bycatch in fisheries. Endanger Species Res 34:373–396. https://doi.org/10.3354/esr00869
Duffy D (1990) Seabirds and the 1982–1984 El Nino-Southern Oscillation: global ecological consequences of the 1982–83 El Nino-Southern Oscillation. Elsevier Oceanogr Ser 52:395–415. https://doi.org/10.1016/S0422-9894(08)70043-6
Dilley BJ, Schoombie S, Stevens K, Davies D, Perold V, Osborne A, Schoombie J, Brink CW, Carpenter-Kling T, Ryan PG (2017) Mouse predation affects breeding success of burrow-nesting petrels at sub-Antarctic Marion Island. Antarct Sci 30:93–104. https://doi.org/10.1017/S0954102017000487
Dias MP, Martin R, Pearmain EJ, Burfield IJ, Small C, Phillips RA, Yates O, Lascelles B, Garcia Borboroglu P, Croxall JP (2019) Threats to seabirds: A global assessment. Biol Conserv 237:525–537. https://doi.org/10.1016/j.biocon.2019.06.033
García-Borboroglu P, Boersma P, Ruoppolo V, Reyes L, Rebstock G, Griot K, Heredia S, Adornes A, da Silva R (2006) Chronic oil pollution harms Magellanic penguins in the Southwest Atlantic. Mar Pollut Bull 52:193–198. https://doi.org/10.1016/j.marpolbul.2005.11.004
Hocken AG (2000) Cause of death in blue penguins (Eudyptula m. minor) in North Otago, New Zealand. N Z J Zool 27:305–309. https://doi.org/10.1080/03014223.2000.9518239
Hocken AG (2005) Necropsy findings in yellow-eyed penguins (Megadyptes antipodes) from Otago, New Zealand. N Z J Zool 32:1–8. https://doi.org/10.1080/03014223.2005.9518391
IUCN (2022) The IUCN Red List of Threatened Species. Version 2022-2. https://www.iucnredlist.org. Accessed 11 Nov 2022
Kemper J, Roux J, Underhill L (2008) Effect of age and breeding status on molt phenology of adult African penguins Spheniscus demersus in Namibia. Auk 125:809–819. https://doi.org/10.1525/auk.2008.06262
Klusener R, Hurtado R, Parsons N, Vanstreels R, Stander N, van der Spuy S, Ludynia K (2018) From incubation to release: Hand-rearing as a tool for the conservation of the endangered African penguin. PLoS ONE 13:e0205126. https://doi.org/10.1371/journal.pone.0205126
Liversidge R (1959) Tropical mountain birds south of the Zambezi. Ostrich 30:68–78. https://doi.org/10.1080/00306525.1959.9633314
La Cock GD (1986) The Southern Oscillation, environmental anomalies, and mortality of two Southern African seabirds. Clim Change 8:173–184. https://doi.org/10.1007/BF00139753
Morant P, Brooke RK, Abrams RW (1983) Recoveries in southern Africa of seabirds breeding elsewhere. Ring Migr 4:257–268. https://doi.org/10.1080/03078698.1983.9673818
Miskelly CM, Scofield RP, Sagar PM, Tennyson AJD, Bell BD, Bell EA (2011) Vagrant and extra-limital bird records accepted by the OSNZ Records Appraisal Committee 2008–2010. Notornis 58:64–70
Maree B, Wanless R, Fairweather T, Sullivan B, Yates O (2014) Significant reductions in mortality of threatened seabirds in a South African trawl fishery. Anim Conserv 17:520–529. https://doi.org/10.1111/acv.12126
Nuckton TJ, Goldreich D, Rogaski KD, Lessani TM, Higgins PJ, Claman DM (2002) Hypothermia from prolonged immersion: biophysical parameters of a survivor. J Emerg Med 22:371–374
Nogales M, Martin A, Tershy BR. Donlan CJ, Veitch D, Puerta N, Wood, Alonso J (2004) A review of feral cat eradication on islands. Conserv Biol 18:310–319
Null I (2019) El Niño and La Niña Years and Intensities - Based on Oceanic Niño Index (ONI). http://ggweather.com/enso/oni.htm. Accessed 8 Apr 2021
Oppel S, Clark BL, Risi MM, Horswill C, Converse SJ, Jones CW, Osborne AM, Stevens K, Perold V, Bond AL, Wanless RM, Cuthbert R, Cooper J, Ryan PG (2022) Cryptic population decrease due to invasive species predation in a long-lived seabird supports need for eradication. J Appl Ecol. https://doi.org/10.1111/1365-2664.14218
Pashby BS, Cudworth J (1969) The fulmar ‘wreck’ of 1962. Br Birds 62:97–109
Parsons N, Underhill L (2005) Oiled and injured African penguins Spheniscus demersus and other seabirds admitted for rehabilitation in the Western Cape, South Africa, 2001 and 2002. Afr J Mar Sci 27:289–296. https://doi.org/10.2989/18142320509504087
Phillips R, Silk J, Croxall J, Afanasyev V, Bennett V (2005) Summer distribution and migration of nonbreeding Albatrosses. Individual consistencies and implications for conservation. Ecol 86:2386–2396. https://doi.org/10.1890/04-1885
Pichegru L, Ryan P, Le Bohec C, van der Lingen C, Navarro R, Petersen S, Lewis S, van der Westhuizen J, Grémillet D (2009) Overlap between vulnerable top predators and fisheries in the Benguela upwelling system: implications for marine protected areas. Mar Ecol Prog Ser 391:199–208. https://doi.org/10.3354/meps08283
Pichegru L, Ryan P, van Eeden R, Reid T, Grémillet D, Wanless R (2012) Industrial fishing, no-take zones and endangered penguins. Biol Conserv 156:117–125. https://doi.org/10.1016/j.biocon.2011.12.013
Paleczny M, Hammill E, Karpouzi V, Pauly D (2015) Population Trend of the World’s Monitored Seabirds, 1950–2010. PLoS ONE 10:e0129342. https://doi.org/10.1371/journal.pone.0129342
Parsons NJ, Gous TA, Cranfield MR, Cheng LI, Schultz A, Horne E, Last RP, Lampen F, Ludynia K, Bousfield B, Strauss V, Peirce MA, Vanstreels RET (2017) Novel vagrant records and occurrence of vector-borne pathogens in King Penguins (Aptenodytes patagonicus) in South Africa. Polar Biol 41:625–637. https://doi.org/10.1007/s00300-017-2171-7
Parsons NJ, Vanstreels RET, Schaefer AM (2018) Prognostic indicators of rehabilitation outcomes for adult African Penguins (Spheniscus demersus). J Wildl Dis 54:54–65. https://doi.org/10.7589/2017-06-146
Rand R (1954) Notes on the birds of Marion Island. Ibis 96:173–205. https://doi.org/10.1111/j.1474-919X.1954.tb04125.x
Ryan P, Moloney C (1988) Effect of trawling on bird and seal distributions in the southern Benguela region. Mar Ecol Prog Seri 45:1–11
Ryan PG, Avery G, Rose B, Ross GJB, Sinclair JC, Vernon CJ (1989) The Southern Ocean seabird irruption to South African waters during winter 1984. Cormorant 17:41–55
Russell JC (2011) Indirect effects of introduced predators on seabird islands. In: Mulder CP, Anderson WB, Towns DR, Bellingham PJ (eds) Seabird islands: ecology, invasion, and restoration. Oxford University Press, New York, pp 261–279
Ryan P (2017) Guide to seabirds of Southern Africa. Struik Nature, Century City
Shannon LV (1985) The Benguela ecosystem. I: evolution of the Benguela physical features and processes. Oceanog Mar Biol 23:105–182
Schramm M (1986) The diet of chicks of Great-winged, Kerguelen and Soft-plumaged Petrels at the Prince Edward Islands. Ostrich 57:9–15. https://doi.org/10.1080/00306525.1986.9633632
Shealer DA (2001) Foraging behavior and food of seabirds. In: Shreiber EA, Burger J (eds) Biology of marine birds. CRC Press, Boca Raton, pp 137–178
Sherley R, Barham B, Barham P, Campbell K, Crawford R, Grigg J, Horswill C, McInnes A, Morris T, Pichegru L, Steinfurth A, Weller F, Winker H, Votier S (2018) Bayesian inference reveals positive but subtle effects of experimental fishery closures on marine predator demographics. Proc R Soc B 285:20172443. https://doi.org/10.1098/rspb.2017.2443
Threlfall W, Eveleigh E, Maunder JE (1974) Seabird mortality in a storm. Auk 91:846–849. https://doi.org/10.2307/4084756
Trathan P, García-Borboroglu P, Boersma D, Bost C, Crawford R, Crossin G, Cuthbert R, Dann P, Davis L, De La Puente S, Ellenberg U, Lynch H, Mattern T, Pütz K, Seddon P, Trivelpiece W, Wienecke B (2015) Pollution, habitat loss, fishing, and climate change as critical threats to penguins. Conserv Biol 29:31–41. https://doi.org/10.1111/cobi.12349
Tavares D, Fulgencio de Moura J, Siciliano S (2016) Environmental predictors of seabird wrecks in a tropical coastal area. PLoS ONE 11:e0168717. https://doi.org/10.1371/journal.pone.0168717
Vanstreels RET, Parson NJ, Sherley RB, Stander N, Strauss V, Kemper J, Waller L, Barham BJ, Ludynia K (2023) Factors determining the number of seabirds impacted by oil spills and the success of their rehabilitation: Lessons learned from Namibia and South Africa. Mar Pollut Bull 188:114708. https://doi.org/10.1016/j.marpolbul.2023.114708
Votier S, Hatchwell B, Beckerman A, McCleery R, Hunter F, Pellatt J, Trinder M, Birkhead T (2005) Oil pollution and climate have wide-scale impacts on seabird demographics. Ecol Lett 8:1157–1164. https://doi.org/10.1111/j.1461-0248.2005.00818.x
Williams JM, Tasker ML, Carter IC, Webb A (1994) A method of assessing seabird vulnerability to surface pollutants. Ibis 137:147–152. https://doi.org/10.1111/j.1474-919X.1995.tb08435.x
Webster R, Aguilar R, Argandona-Gonzalez A, Conayne P, De Sousa D, Sriram A, Svensson C, Gartrell B (2016) Forced molt in four juvenile Yellow-eyed penguins Megadyptes antipodes. J Wildl Dis 52:809–816. https://doi.org/10.7589/2015-11-305
Acknowledgements
The authors thank the staff and volunteers working at the Southern African Foundation for the Conservation of Coastal Birds (SANCCOB) who performed all the rehabilitation work and generated the data for this study. SANCCOB is supported by a wide range of donors, including the International Fund for Animal Welfare, Hans Hoheisen Charitable Trust, and the National Lottery Distribution Trust Fund. SANCCOB worked under annual permits for the rehabilitation of seabirds from the Department of Forestry, Fisheries and the Environment (South Africa) for this study. Lastly we also thank Dr. Dieter Piepenburg and two anonymous reviewers for their constructive and helpful comments.
Author information
Authors and Affiliations
Contributions
AS, RETV and KL contributed to the study conception and design. Material preparation, data collection and analysis were performed by AS, RETV and NJP. The first draft of the manuscript was written by AS and RETV. All authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
The authors have no competing interests to declare that are relevant to the content of this article. All authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest or non-financial interest in the subject matter or materials discussed in this manuscript. No ethical approval was required as this is descriptive account and data were collected as part of official rehabilitation activities. No funding was received to assist with the preparation of this manuscript.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Snyman, A., Vanstreels, R.E.T., Parsons, N.J. et al. Visitor and vagrant seabirds admitted for rehabilitation in South Africa, 1993–2017. Polar Biol 46, 1097–1110 (2023). https://doi.org/10.1007/s00300-023-03186-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-023-03186-w