
Abstract
Diet compositions of Patagonotothen cornucola and Harpagifer bispinis were studied from two localities, one with continental influence in a wind-protected area (Puerto Riquelme) and another with oceanic influence in a wind-exposed area (Puerto Williams). The dietary analysis performed showed intra-specific differences among contrasting intertidal areas, with greater richness and diversity of prey types in fish from Puerto Riquelme than in Puerto Williams. Gammarid amphipods and Tanaidae dominated the diet of P. cornucola and H. bispinis in Puerto Riquelme, whereas the diet of both fish species in Puerto Williams was characterized by the crustacean isopod Exosphaeroma gigas and gammarids. The dietary overlap was quantified using two similarity indexes (Sorensen and Morisita-Horn), and the results obtained revealed a high diet overlap among the species in each locality. Diet composition did not show significant difference between sexes in both species, while P. cornucola from Puerto Riquelme presented significant differences in fish size in the diet, supporting the existence of some food partitioning. Dissolved oxygen and salinity were important factors in the diet variability of P. cornucola and H. bispinis, since both species showed greater richness and diversity of prey types in intertidal areas with higher dissolved oxygen and lower salinity levels (Puerto Riquelme). In conclusion, the dietary patterns of these two sympatric rocky intertidal species in the Patagonian channels and fjords appear to be regulated by the different environmental conditions that influence the availability and abundance of potential prey in the diet.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The channel and fjord ecosystems of southern Chile are characterized by great diversity and heterogeneity of coastal marine habitats, mainly due to the high degree of geomorphological and hydrographic complexity that have been molded by ice expansion and contraction during the Quaternary glacial period (Hulton et al. 2002; Sudgen et al. 2002). Habitat heterogeneity is also influenced both by freshwater discharge from the melting of the Southern Patagonia Ice Field (48°20′–51°30′S) and by oceanographic factors related to sub-Antarctic Pacific waters that mix with Atlantic waters through the Strait of Magellan and Beagle Channel (Andrade 1991; Antezana 1999; Rignot et al. 2003; Porter and Santana 2003; Sievers and Silva 2006). The rocky intertidal area is a highly variable habitat because of physical factors associated with wave exposure, wind and tidal fluctuations, affecting temperature and nutrient concentration (Menge and Branch 2001). For example, rocky intertidal communities in channels and fjords of southern Chile are subjected to intense physical stress due to severe weather conditions, characterized by westerly winds between 20 and 40 km h−1 and annual rainfall ranging from 2000 to 8000 mm (Zamora and Santana 1979; Andrade 1991). The climatic conditions in these rocky intertidal areas cause variations in salinity related to the drying effects of wind and freshwater runoff caused by heavy rainfall. Despite the adverse climatic conditions of the rocky intertidal environment, the abundance of fish that live in this area is striking, primarily species of the family Nototheniidae (Lloris and Rucabado 1991; Pequeño et al. 1995; Hüne and Ojeda 2012). The nototheniid fish inhabiting high latitudes of the Southern Hemisphere play important ecological roles in the marine ecosystem, utilizing a broad variety of trophic niches that in other regions are used by fish belonging to several taxa (Eastman 1993; Klingenberg and Ekau 1996). Many authors outlined the importance of nototheniid fish as predators on benthic, zooplanktonic and nektonic organisms, while as prey they are a key food resource for most top predators (Birt et al. 1987; Leopold et al. 1998; Barrera-Oro 2002; La Mesa et al. 2004). In the channels and fjords south of the Gulf of Penas (48°S), the rocky intertidal ichthyofauna is dominated by the nototheniids Harpagifer bispinis and Patagonotothen cornucola (Moreno and Jara 1984; Lloris and Rucabado 1991; Hüne and Ojeda 2012). They are benthic species inhabiting shallow subtidal areas (<15 m depth) along the southern extreme of South America (Lloris and Rucabado 1991; Hüne and Ojeda 2012; Reyes and Hüne 2012). Adults of H. bispinis reach 10 cm total length (TL), although most are < 7 cm, while the size range of P. cornucola is from 5 to 13 cm TL (Lloris and Rucabado 1991; Reyes and Hüne 2012). Both species have two spawning periods per year in the Beagle Channel, during the austral winter and summer (Rae and Calvo 1995, 1996). The diet of these fishes consists mainly of benthic invertebrates such as isopods and polychaetes (Moreno and Jara 1984; Hidalgo et al. 2007; Hüne and Rivera 2010). However, there is no information about the factors that influence their feeding behaviors, changes in their diet between size groups or trophic overlap among these species.
The partitioning of resources (e.g., food and habitat) may be viewed as one of the key factors in the diversifying process, which promotes the coexistence of closely related and ecologically equivalent species (Colwell and Fuentes 1975). Partitioning of resources results in maximization of habitat availability and facilitation of species coexistence, which contribute to determine the community structure (Pianka 1974). On the other hand, as proposed by Zander and Berg (1984), coexistence of dissimilar species in the ecosystem can be explained based on the separation of the species into different ecological niches. Niche overlap may cause the competitive exclusion of a species, or species may avoid exclusion by employing isolation mechanisms (Diamond 1978). In the case of food resources, this separation avoids possible competition among consumers on the same trophic levels, with coexisting fish species presenting differences in their diet composition (Morton et al. 2008). The analysis of dietary overlap provides key information on the similarity in the use of resources and the degree of trophic competition among the species (Colwell and Futuyma 1971; Mayr and Berger 1992). Therefore, the degree of species overlap in the utilization of resources has become a valuable approach in studying both community structure and species coexistence.
Considering that H. bispinis and P. cornucola share the same intertidal habitats, exhibiting similar feeding behavior and reproductive activity (Moreno and Jara 1984; Lloris and Rucabado 1991; Rae and Calvo 1995; Rae and Calvo 1996; Hidalgo et al. 2007; Hüne and Rivera 2010; Hüne and Ojeda 2012; Reyes and Hüne 2012), in this study we attempt to (1) determine whether the dietary composition of these two sympatric species exhibits variations between two rocky intertidal areas with different environmental conditions, identifying those environmental variables influencing feeding behavior of both species; (2) assess the degree of diet overlap between H. bispinis and P. cornucola; and (3) determine the influence of body size and sex in the composition of the diet. The analysis of prey diversity and the degree of dietary overlap among the species will help us to better understand the different interspecific patterns in the use of resources depending on fish size, sex and type of environment.
Materials and methods
Study area and sampling
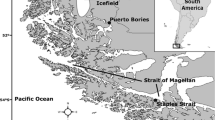
The study was performed in two localities of the Magellan Province: (1) Puerto Riquelme (PR) (51°49′S, 72°38′W) in the Almirante Montt Gulf, close to the Última Esperanza fjord, an area influenced by the freshwater discharge from the melting of the Southern Patagonia Ice Field and protected from waves and westerly winds and (2) Puerto Williams (PW) (54°56′S, 67°35′W) on Navarino Island, an area associated with the Beagle Channel with oceanic water influence and exposed to waves caused by westerly winds (Fig. 1). The samples of H. bispinis and P. cornucola were collected between September 5, 2008, and September 20, 2008. At each locality, the main oceanographic variables of the water column were measured with a multiparameter instrument (YSI model 6600 V2), obtaining measurements every 3 s between 0 and 10 m depth. During the study, the tidal range varied between 0.5 m (PR) and 1.3 m (PW) (SHOA 2008). All fish were captured by hand and with hand nets during diurnal low-tide periods. All individuals caught were measured for total length (TL, cm) and total weight (g) and sexed. After dissection, the stomach was removed, labeled and preserved in a solution of 80 % ethanol for subsequent analysis. In the laboratory, all stomachs were put in Petri plates and washed and drained, then weighed (g) with and without contents. Stomach contents were identified to the lowest possible prey taxon, counted and weighed.

Study area and location of samples in black circles: PR Puerto Riquelme and PW Puerto Williams
Diet composition
In order to assess sample sufficiency, randomized cumulative prey curves were constructed for each species in each locality. Stomach contents of H. bispinis and P. cornucola were randomized 1000 times, using EcoSim version 7.72 (Gotelli and Entsminger 2008). The mean cumulative number of prey types and Shannon diversity index were plotted as a function of the number of non-empty stomachs (Gotelli and Colwell 2001; Magurran 2004). According to Hurtubia (1973), sample size is considered sufficient to describe diet if the cumulative Shannon diversity index reaches an asymptote. To compare the Shannon diversity index and prey richness among fish species and localities, we calculated a rarefied estimate of this index, i.e., the mean expected number of prey types and diversity observed given a fixed number of stomachs (Gotelli and Colwell 2001). For each species at each location, we rarefied all samples to the least number of stomachs analyzed using EcoSim. The contribution of different prey items to the diet of H. bispinis and P. cornucola in each locality was determined by the frequency of occurrence (%F), numerical importance (%N) and mass (%W) (Cortés 1997). The index of relative importance (IRI) was calculated as IRI = %F (%N + %W) and expressed as a percentage (%IRI) (Hyslop 1980).
Diet overlap and feeding habits
Diet composition was first evaluated using univariate measures of diet similarity. Two similarity indices were selected, the Sorensen similarity index (S S ) and the Morisita-Horn index (C H ). The Sorensen similarity index measures overlap in prey taxa between pairs of consumers (A and B), irrespective of the number of prey items of each taxon consumed (Magurran 2004):
where a = number of taxa in the diets of both species A and B; b = number of taxa in the diet of species B but not in the diet of species A; c = number of taxa in the diet of species A but not in the diet of species B. In contrast, the Morisita-Horn index (C H ) quantitatively assesses diet overlap in prey numbers between two species (Magurran 2004):
where p iA ; p iA = proportion consumed of taxon i out of the total number of prey consumed by fish species A and B, respectively; n = total number of prey taxa. EstimateS version 8.2 (Colwell 2009) was used to calculate both indexes of diet overlap. Differences in diet composition were additionally investigated using multivariate ordination analysis. A Bray–Curtis coefficient similarity matrix was obtained from the square root-transformed data set. A cluster analysis was then applied to determine groupings among species and localities. The significance of these groupings was assessed using a similarity profile test (SIMPROF) implemented in PRIMER version 6 (Clarke and Warwick 2001; Clarke and Gorley 2006; Clarke et al. 2008).
To determine whether species, localities, body size and sex affect significantly the diet of H. bispinis and P. cornucola, we used PERMANOVA, a linear permutation test based on 9999 permutations of the matrix of distances calculated with the Bray–Curtis similarity index (Anderson 2001), implemented in the VEGAN R-package in R version 3.0.2 (R Core Team 2013). Regarding their length composition (total length = TL), we chose arbitrarily two group sizes depending on the total range observed in each species: H. bispinis, small (3.5–5.5 cm) and large (5.6–7.6 cm); P. cornucola, small (5.1–9.0 cm) and large (9.1–13.0 cm). Group sizes of H. bispinis and P. cornucola were expressed in 2.1- and 4.0-cm intervals, respectively. A dissimilarity analysis was performed to determine the relevant contribution of each prey species to the diet composition between species and localities using the SIMPER routine of the PRIMER software. Finally, in order to analyze the relationship between variations in diet composition and environmental factors, a non-metric multidimensional scaling (nMDS) was applied. The similarity matrix of fish species and localities was analyzed in a two-dimensional plane according to their diet composition. Environmental variables (salinity, temperature, dissolved oxygen) were also superimposed with the ordisurf function of the VEGAN R-package, to fit a generalized additive model (GAM) surface to the ordination plot (Oksanen et al 2010). The correlations of the environmental variables with the nMDS plot were projected as vectors on the ordination to visualize their relationship with diet variability. The vectors point in the direction of the most rapid change in the environmental variable, and their length is proportional to the correlation between diet composition and the environmental variable. The significance of correlations was tested with a randomized permutation test using 10,000 permutations.
Results
Diet composition
Of the total 80 and 24 specimens of P. cornucola examined from PR and from PW, 29 (36 %) and 2 (8 %) had empty stomachs, respectively. New types of prey continued to be identified with increasing sample size in PW (Fig. 2a). However, the diversity curve reached an asymptote, indicating that the sample was large enough to describe the diversity of the diet (Fig. 2b). The sample size was large enough to describe the diet adequately in PR, as the cumulative number of prey types and diversity curves reached an asymptote (Fig. 2a, b). Fish size of P. cornucola with stomach contents was composed of 28 females ranged between 6.8 and 12.9 cm TL, with an average (±SD) of 9.0 ± 1.6 cm, and 45 males ranged between 5.1 and 12.9 cm TL, with an average of 9.2 ± 2.0 cm (Fig. 3a) (Online Resource 1).

Cumulative curves of non-empty stomachs of P. cornucola (a, b) and H. bispinis (c, d). a–c Mean cumulative number of prey types identified and b–d, mean cumulative diversity of prey categories (using the Shannon H′ index of diversity). Broken lines indicate the 95% confidence intervals

Length–frequency distributions of P. cornucola (a) and H. bispinis (b). Samples locations: PW (black) and PR (gray)
Of the total 42 and 75 specimens of H. bispinis examined from PR and from PW, 3 (7 %) and 10 (13 %) had empty stomachs, respectively. In both localities, the cumulative number of prey types and diversity curves reached an asymptote, showing that the sample size was large enough to describe the diet (Fig. 2c, d). Fish size of H. bispinis with stomach contents was composed of 47 females ranged between 3.8 and 7.6 cm TL, with an average (±SD) of 5.6 ± 0.8 cm, and 57 males ranged between 4.3 and 7.2 cm TL, with an average of 5.5 ± 0.8 cm (Fig. 3b) (Online Resource 1). We rarefied all samples to 22 stomachs, the number of stomachs observed in the least abundant location. Rarefied estimates of prey richness ranged between 20.79 (P. cornucola in PR) and 7.58 (H. bispinis in PW), while estimates of the Shannon diversity index varied between 2.06 and 0.99 in the diet of P. cornucola in PR and H. bispinis in PW, respectively (Online Resource 2 and 3). The diet of P. cornucola in PR was characterized by the dominance of gammarid amphipods (IRI = 35.25 %) and Tanaidae (IRI = 25.30 %). In contrast, the main prey in PW were the crustacean isopod Exosphaeroma gigas (IRI = 72.82 %) and gammarids (IRI = 26.46 %) (Online Resource 2). The main prey in the diet of H. bispinis in PR were Tanaidae crustaceans (IRI = 44.66 %) and gammarid amphipods (IRI = 34.66 %), while in PW the diet was characterized by the dominance of the isopod E. gigas (IRI = 75.67 %) and gammarids (IRI = 13.26 %) (Online Resource 3).
Diet overlap and feeding habits
Sorensen similarity results suggested that P. cornucola and H. bispinis diets from PR were most similar, whereas the P. cornucola diet between localities was least similar (Table 1). Morisita-Horn similarity results, on the other hand, suggested that greatest overlap was between P. cornucola and H. bispinis diets of PW, followed by the diets of both species from PR. The diet of H. bispinis between localities was the least similar (Table 1).
Cluster analysis (using SIMPROF) revealed the presence of two spatial clusters within which the diet compositions were similar and significantly different between the groups (P < 0.05, Fig. 4). Diet composition between species from PR showed the highest similarity, followed by the diets of P. cornucola and H. bispinis from PW (Fig. 4). Diet composition showed significant differences among localities in both species (PERMANOVA P. cornucola, F = 17.76, P < 0.001; H. bispinis, F = 38.85, P < 0.001). Diet composition showed significant differences between the species in PR (PERMANOVA, F = 8.65, P = 0.001), but not in PW (PERMANOVA, F = 2.32, P = 0.09). Regarding fish size, the composition of the diet showed significant differences between size groups of P. cornucola in PR, whereas PW did not show significant differences (PERMANOVA PR, F = 3.87, P < 0.01; PW, F = 0.56, P = 0.55). The comparison between small and large size groups of H. bispinis in each locality did not show a significant difference in diet composition (PERMANOVA PR, F = 1.23, P = 0.31; PW, F = 0.68, P = 0.55). In addition, there was no significant difference between sexes in diet composition of P. cornucola (PERMANOVA PR, F = 2.14, P = 0.06; PW, F = 0.31, P = 0.73) or H. bispinis (PERMANOVA PR, F = 0.88, P = 0.52; PW, F = 0.38, P = 0.73).

Dendrogram resulting from the cluster analysis performed on stomach content data in the different species and sites. Each dashed line represents a significant cluster (SIMPROF P < 0.05)
The SIMPER analysis showed greatest dissimilarity between P. cornucola from PR and H. bispinis from PW, followed by the diet of P. cornucola among localities, with an average dissimilarity of 89.27 and 89.07, respectively (Table 2). In contrast, the diet of the two species in PW showed low level of dissimilarity, with an average of 49.87. Crustaceans such as isopods (E. gigas) and gammarid amphipods were the main prey responsible for dissimilarities in diet composition, followed by ostracods, Tanaidae and polychaetes (Nereidae) (Table 2).
The non-metric multidimensional scaling (nMDS) showed that the composition of the diet was correlated with environmental factors and the differences in the diet among localities, with a stress level of 0.12 (Fig. 5). Ordination was significantly correlated with dissolved oxygen, salinity and temperature (P < 0.001), with correlation coefficients (r 2) of 0.68, 0.54 and 0.31, respectively (Fig. 5). A nonlinear influence of environmental variables associated with diet composition was fitted to the ordinations, reflecting a greater level of dissolved oxygen and lower salinity in PR than in PW (Fig. 5).

Plot of diet composition in ordination space (1st 2 NMDS) overlaid on a fitted environmental surface (black contour lines). Sample locations are marked with circles: Harpagifer bispinis from PW (black); Patagonotothen cornucola from PW (dark gray); H. bispinis from PR (gray); P. cornucola from PR (light gray). The arrow points to the direction of most rapid change in the variable: dissolved oxygen (a), temperature (b) and salinity (c)
Discussion
Our study indicated that P. cornucola and H. bispinis exhibited spatial differences in diet composition among different environmental localities and showed greater richness and diversity of prey types in PR than in PW. Rarefied estimates of prey richness and diversity were higher in PR in the diet of P. cornucola (20.79 prey items and H′ = 2.06) and H. bispinis (18 prey items and H′ = 1.77). In contrast, the level estimates of richness and diversity of prey types were surprisingly low in PW in the diet of P. cornucola (9 prey items and H′ = 1.15) and H. bispinis (7.58 prey items and H′ = 0.99). A plausible explanation for this low level of richness and diversity of prey types in the diet of these fish species from the rocky intertidal of PW could be reduced availability of potential prey species conditioned by the desiccation stress during low tides due to westerly winds in the Beagle Channel (Santana et al. 2006). These severe climatic conditions are not present in PR, owing to the position of the rocky intertidal in a wind-protected area, while the PW study site is a wind-exposed area.
Our results showed that the dominant prey species in the diet of P. cornucola and H. bispinis in PR were crustacean gammarids and Tanaidae. In contrast, the dominant prey in PW was the crustacean isopod Exosphaeroma gigas, with 72.82 and 75.67 % of the IRI in the diet of P. cornucola and H. bispinis, respectively. These important levels of the IRI suggest the lack of other potential prey or high abundance of E. gigas. In this regard, Moreno and Jara (1984) found isopods such as E. gigas in the diet of P. cornucola and H. bispinis in the belt of kelp forest (Macrocystis pyrifera) on the east coast of Navarino Island. According to these authors, the high abundances of detritivorous crustaceans (i.e., isopods and amphipods) consuming algal detritus could be linked to the importance of these prey in the diet of both fish species in the kelp forest. Also, the isopod E. gigas represents an important food for other fish species that frequent the intertidal zone of Navarino Island such as Paranotothenia magellanica and Patagonotothen tessellata (Moreno and Jara 1984). The benthic isopod E. gigas has been found in the wind-exposed shoreline of the Strait of Magellan in Tierra Del Fuego Island, an area mainly dominated by mussel beds (Mytilus edulis platensis and Perumytilus purpuratus) (Ríos and Mutschke 1999). According to Bertness et al. (2006), P. purpuratus beds provide shelter from lethal desiccation stress with associated organisms such as E. gigas and gammarid amphipods. These sessile mollusks (M. edulis platensis and P. purpuratus) exhibit high abundance in the intertidal zones of Navarino Island over the whole year (Ojeda et al. 2014). In addition, the isopod E. gigas has been reported in high abundance associated with these mollusks in a wind-exposed intertidal area of the Beagle Channel (Navarino Island) (Jaime Ojeda, pers. comm.). Mussel beds of P. purpuratus are highly abundant in the rocky shoreline of the Atlantic Patagonia, a system extremely exposed to desiccation by wind, where it has been documented that the diet of P. cornucola is similar to that at PW (i.e., small isopods, amphipods and polychaetes) (Hidalgo et al. 2007). In habitats such as the rocky intertidal of PW with intense climatic stress, theory predicts that predation should be less important in structuring communities (Menge and Sutherland 1987). Physical environment factors appear to be the dominating structuring force regulating community structure and prey availability in rocky shorelines that are subjected to severe climatic conditions (Bertness et al. 2006; Hidalgo et al. 2007).
High levels of dietary overlap between intertidal fish species were found in both localities using two similarity indices (Table 1). However, these degrees of diet overlap may include some uncertainty, since some proportions of prey types could not be identified to lower taxonomic levels and might be taken as a potential overestimation of diet overlap between sympatric species. Differences were found between the indices (i.e., greater similarity in PR according to the Sorensen index and a higher level of diet overlap in PW according to the Morisita-Horn index). These differences could be explained because the Sorensen index considers similarity in prey taxa, whereas the Morisita-Horn index considers similarity in number of prey of a given taxon (Magurran 2004). Our cluster analysis was consistent with the similarity indices and indicated an interspecific dietary overlap between P. cornucola and H. bispinis in each locality. In addition, there was an intra-specific difference in diet composition between localities in both intertidal fish species. These results were confirmed with the PERMANOVA analysis and indicated significant spatial differences in diet composition of both species. The PERMANOVA analysis did not show significant differences in fish size in the diet of H. bispinis in either locality, whereas P. cornucola presented significant differences in the diet among size groups in PR, supporting the existence of some food partitioning in PR. In this regard, gammarid amphipods were primarily preyed on by small fish, while the decapod Acanthocyclus albatrossis and fish eggs were eaten preferentially by large fish. This dietary change between fish group sizes has been recorded in Patagonotothen longipes from Chiloé Island in southern Chile, where small fish feed on euphausiids and ostracods, while large fish feed mainly on decapods and mollusks (Murillo and Oyarzún 2002).
According to the SIMPER analysis, dissimilarity in the diet between P. cornucola and H. bispinis was higher in PR than in PW (Table 2). Several authors have proposed that segregation of microhabitats is an important factor for reducing the effects of the competition for trophic resources (Ross 1986; Grossman et al. 1987; Rincón and Lobón-Cerviá 1993). In our case, the differences among fish species could be related to the intertidal zone where they inhabit. For example, we found that P. cornucola mainly occupy the lower level of the intertidal zone, while H. bispinis are abundant at the lower and medium levels. This might be attributed to the high tolerance to variation in salinity and temperature of H. bispinis due to morphological modification in their kidneys and the presence of antifreeze activity in their blood (Pérez et al. 2003; Hüne et al. 2015). Thus H. bispinis could occupy the medium level of the intertidal zone, an area often frozen during winter and exposed to variation in salinity caused by heavy rainfall and desiccation by wind, especially during spring and summer. Another factor that might reduce the effects of the competition is related to differences in the diel activity patterns of fish (Hansen and Closs 2005; David et al. 2007). In our case, we did not observe feeding preferences in certain periods of the day to reduce interspecific competition, mainly because the intertidal fish remain under rocks during low tide. However, this feeding strategy has been observed in other nototheniid species such as P. tessellata, which showed nocturnal activity patterns in the subtidal waters of Patagonian channels (Salas-Berrios et al. 2013; Hüne and Vega 2015).
We also identified with the nMDS analysis visual differences in diet composition between different environments (PR and PW), indicating dissolved oxygen, salinity and temperature as important factors in the diet variability of P. cornucola and H. bispinis (Fig. 5). During our study, PR exhibited higher dissolved oxygen and lower salinity than PW. Consequently, P. cornucola and H. bispinis showed greater richness and diversity of prey types in environmental conditions with higher dissolved oxygen and less salinity. This could be attributed to higher availability of prey in the wind-protected intertidal of PR, or alternatively, high abundance of few potential prey in the wind-exposed intertidal of PW. According to Prince and Goodyear (2006), low levels of dissolved oxygen, as recorded in PW, could also restrict the movement of predators, implying a lower catch of mobile prey. However, we did not observe such restriction in PW, as both fish species feed mainly on E. gigas, a highly mobile isopod. On the other hand, the estimated higher level of richness and diversity of prey types in P. cornucola of PR could be related to a size factor, because larger specimens were recorded in this locality. In this regard, numerous studies indicate that many fish species change their diet from smaller, easier to access prey, to larger prey or to prey more difficult to catch and with higher nutritive value (Hanson and Chouinard 2002; Nakamura et al. 2003).
Finally, considering the diet composition of P. cornucola and H. bispinis from two rocky intertidal areas in the channels and fjords of southern Chile, our results suggest intra-specific difference in trophic dynamics among contrasting environmental localities and interspecific dietary overlap between these species in each rocky intertidal area. The dietary patterns of these two sympatric species appear to be regulated by the different environmental conditions that influence the availability and abundance of potential prey in the diet. However, this preliminary result should be confirmed with seasonal sampling effort in other localities of Patagonia to understand better the trophic ecology of these important intertidal fish.
References
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecol 26:32–46. doi:10.1111/j.1442-9993.2001.01070
Andrade S (1991) Geomorfología costera y antecedentes oceanográficos físicos de la región de Magallanes, Chile (48°–56°S). An Inst Pat Ser Cienc Nat 20:135–151
Antezana T (1999) Hydrographic features of Magellan and Fuegian inland passages and adjacent Subantarctic waters. Sci Mar 63:23–34
Barrera-Oro E (2002) Review: the role of fish in the Antarctic marine food web: differences between inshore and offshore waters in the southern Scotia Arc and west Antarctic Peninsula. Antarct Sci 14:293–309
Bertness MD, Crain CM, Silliman BR, Bazterrica MC, Reyna MV, Hildago F, Farina JK (2006) The community structure of western Atlantic Patagonian rocky shores. Ecol Monogr 76:439–460
Birt V, Birt T, Goulet D, Lairus D, Montevecchi W (1987) Ashmole’s halo: direct evidence for prey depletion by a seabird. Mar Ecol Prog Ser 40:205–208
Clarke KR, Gorley RN (2006) Primer v6: user manual/tutorial
Clarke KR, Warwick RM (2001) Changes in marine communities: an approach to statistical analysis and interpretation. Plymouth Marine Laboratory
Clarke KR, Somerfield P, Gorley R (2008) Testing of null hypotheses in exploratory community analyses: similarity profiles and biota-environment linkage. J Exp Mar Biol Ecol 366:56–69
Colwell RK (2009) EstimateS: statistical estimation of species richness and shared species from samples. Version 8.2. User’s Guide and application published at: http://purl.oclc.org/estimates
Colwell RK, Fuentes E (1975) Experimental studies of the niche. Annu Rev Ecol Syst 6:281–310
Colwell RK, Futuyma DJ (1971) On the measurement of niche breadth and overlap. Ecology 52:567–576
Cortés E (1997) A critical review of methods of studying fish feeding based on analysis of stomach contents: application to the elasmobranch fishes. Can J Fish Aquat Sci 54:726–738
David B, Closs G, Crow S, Hansen E (2007) Is diel activity determined by social rank in a drift-feeding stream fish dominance hierarchy? Anim Behav 74:259–263
Diamond JM (1978) Niche shifts and the rediscovery of interspecific competition. Am Sci 66:322–331
Eastman JT (1993) Antarctic fish biology: evolution in a unique environment. Academic Press, San Diego
Gotelli N, Colwell R (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4:379–391
Gotelli N, Entsminger G (2008) EcoSim: null models software for ecology. Version 7.72 Acquire Intelligence & Kesey-Bear
Grossman G, de Sostoa A, Freeman M, Lobón-Cerviá J (1987) Microhabitat use in a Mediterranean riverine fish assemblage. Fishes of the Upper Matarraña. Oecologia 73:501–512
Hansen E, Closs G (2005) Diel activity and home range size in relation to food supply in a drift-feeding stream fish. Behav Ecol 16:640–648
Hanson J, Chouinard G (2002) Diet of Atlantic cod in the southern Gulf of St Lawrence as an index of ecosystem change, 1959–2000. J Fish Biol 60:902–992
Hidalgo F, Silliman B, Bazterrica M, Bertness M (2007) Predation on the Rocky Shores of Patagonia, Argentina. Estuar Coast 30:886–894
Hulton N, Purves P, McCulloch R, Sugden D, Bentley M (2002) The Last Glacial Maximum and deglaciation in southern South America. Quat Sci Rev 21:233–241
Hüne M, Ojeda J (2012) Estructura del ensamble de peces costeros de los canales y fiordos de la zona central de la Patagonia chilena (48°–52°S). Rev Biol Mar Oceanogr 47:451–460
Hüne M, Rivera G (2010) Contribución de poliquetos (Annelida: Polychaeta) en la dieta de tres especies de nototénidos (Perciformes: Notothenioidei) en la región de Magallanes. An Inst Patagonia 38:39–46
Hüne M, Vega R (2015) Spatial variation in the diet of Patagonotothen tessellata (Pisces, Nototheniidae) from the fjords and channels of southern Chilean Patagonia. Polar Biol 38:1613–1622. doi:10.1007/s00300-015-1726-8
Hüne M, González-Wevar C, Poulin E, Mansilla A, Fernández DA, Barrera-Oro E (2015) Low level of genetic divergence between Harpagifer fish species (Perciformes: Notothenioidei) suggests a Quaternary colonization of Patagonia from the Antarctic Peninsula. Polar Biol 38:607–617. doi:10.1007/s00300-014-1623-6
Hurtubia J (1973) Trophic diversity measurement in sympatric predatory species. Ecology 54:885–890
Hyslop EJ (1980) Stomach contents analysis—a review of methods and their application. J Fish Biol 17:411–429
Klingenberg CP, Ekau W (1996) A combined morphometric and phylogenetic analysis of an ecomorphological trend: pelagization in Antarctic fishes (Perciformes: Nototheniidae). Biol J Linn Soc 59:143–177
La Mesa M, Eastman JT, Vacchi M (2004) The role of notothenoid fish in the food web of the Ross Sea shelf waters: a review. Polar Biol 27:321–338
Leopold M, van Damme C, van der Veer H (1998) Diet of cormorans and the impact of cormorant predation on juvenile flatfish in the Dutch Wadden Sea. J Sea Res 40:93–107
Lloris D, Rucabado J (1991) Ictiofauna del Canal Beagle (Tierra del Fuego), aspectos ecológicos y análisis biogeográfico. Instituto Español de Oceanografía, Madrid
Magurran A (2004) Measuring biological diversity. Blackwell, Oxford
Mayr M, Berger A (1992) Territoriality and microhabitat selection in two intertidal New Zealand fish. J Fish Biol 40:243–256
Menge BA, Branch GM (2001) Rocky intertidal communities. In: Bertness M, Gaines S, Hay M (eds) Marine community ecology. Sinauer Associates, Sunderland, pp 221–251
Menge BA, Sutherland JP (1987) Community regulation: Variation in disturbance, competition, and predation in relation to environmental stress and recruitment. Am Nat 130:730–757
Moreno C, Jara F (1984) Ecological studies on fish fauna associated with Macrocystis pyrifera belts in the south Fueguian Islands, Chile. Mar Ecol Prog Ser 15:99–107
Morton JK, Platell ME, Gladstone W (2008) Differences in feeding ecology among three co-occurring species of wrasse (Teleostei: Labridae) on rocky reefs of temperate Australia. Mar Biol 154:577–592. doi:10.1007/s00227-008-0951-x
Murillo C, Oyarzún C (2002) Variación ontogenética en la dieta de Patagonotothen longipes (Steindachner 1876) (Perciformes: Nototheniidae) en el fiordo de Castro, Chiloe. Gayana 66:219–224. doi:10.4067/S0717-65382002000200018
Nakamura Y, Horimonchi M, Nakai T, Sano M (2003) Food habits of fishes in a seagrass bed on a fringing reef at Iriomote Island, Southern Japan. Ichthyol Res 50:15–22
Ojeda J, Rosenfeld S, Marambio J, Rozzi R, Mansilla A (2014) Patrones estacionales y espaciales de la diversidad de moluscos intermareales de bahía Róbalo, canal Beagle, Reserva de la Biosfera Cabo de Hornos, Chile. Rev Biol Mar Oceanogr 49:493–509
Oksanen J, Blanchet F, Kindt R, Legendre P, O’Hara R, Simpson G, Solymos P, Stevens M, Wagner H (2010) Vegan: Community ecology package, Version 2.0.6 http://CRAN.R-project.org/package=vegan
Pequeño G, Lamilla J, Lloris D, Rucabado J (1995) Comparación entre las ictiofaunas intermareales de los extremos austral y boreal de los canales patagónicos. Rev Biol Mar 30:155–177
Pérez A, Calvo J, Tresguerres M, Luquet C (2003) Aglomerularism in Harpagifer bispinis: a subantarctic notothenioid fish living at reduced salinity. Polar Biol 23:800–805
Pianka ER (1974) Niche overlap and diffuse competition. P Natl Acad Sci USA 71:2141–2145
Porter C, Santana A (2003) Rapid 20th century retreat of Ventisquero Marinelli in the Cordillera 486 Darwin Icefield. An Inst Patagonia 31:17–26
Prince E, Goodyear P (2006) Hypoxia-based habitat compression of tropical pelagic fishes. Fish Oceanogr 15:451–464
R Core Team (2013) R: a language and environment for statistical computing, Vienna. http://www.r-project.org
Rae GA, Calvo J (1995) Fecundity and reproductive habits in Patagonothen tessellata (Richardson, 1845) from the Beagle Channel, Argentina. Antarct Sci 7:235–240
Rae GA, Calvo J (1996) Histological analysis of gonadal development in Patagonotothen tessellata (Richardson 1845) (Nototheniidae: Pisces) from the Beagle Channel, Argentina. J Appl Ichthyol 12:31–38
Reyes P, Hüne M (2012) Peces del Sur de Chile. Ocho Libros Ediciones, Santiago de Chile
Rignot E, Rivera A, Casassa G (2003) Contribution of the Patagonia Icefields of South America to Sea Level Rise. Science 302:434–437
Rincón P, Lobón-Cerviá J (1993) Microhabitat use by stream-resident Brown trout: bioenergetic consequences. Trans Am Fish Soc 122:575–587
Ríos C, Mutschke E (1999) Community structure of intertidal Boulder-cobble fields in the Strait of Magellan Chile. Sci Mar 63(Suppl 1):193–201
Ross S (1986) Resource partitioning in fish assemblages: a review of field studies. Copeia 1986:352–388
Salas-Berrios F, Valdés-Aguilera J, Landaeta MF, Bustos CA, Pérez-Vargas A, Balbontín F (2013) Feeding habits and diet overlap of marine fish larvae from the peri-Antarctic Magellan region. Polar Biol 36:1401–1414. doi:10.1007/s00300-013-1359-8
Santana A, Porter C, Butorovic N, Olave C (2006) Primeros antecedentes climatológicos de Estaciones Automáticas (AWS) en el canal Beagle, Magallanes, Chile. An Inst Pat Ser Cienc Nat 34:5–20
SHOA (2008) Tabla de marea de la costa de Chile. Servicio Hidrográfico y Oceanográfico de la Armada de Chile, Valparaíso. http://www.shoa.cl
Sievers H, Silva N (2006) Masas de agua y circulación en los canales y fiordos australes. In: Silva N, Palma S (eds) Avances en el conocimiento de las aguas interiores chilenas, Puerto Montt a Cabo de Hornos. CONA, Valparaíso, pp 53–58
Sudgen DE, Hulton N, Purves RS (2002) Modelling the inception of the Patagonian icesheet. Quatern Int 95–96:55–64
Zamora E, Santana A (1979) Características climáticas de la costa occidental de la Patagonia, entre las latitudes 46°40′ y 56°30′(S). An Inst Pat 10:109–154
Zander CD, Berg J (1984) Feeding ecology of littoral gobiid and blennioid fishes of the Banyuls area (Mediterranean Sea) II. Prey selection and size preference. Vie Milieu 34:149–157
Acknowledgments
The authors would like to express their gratitude to Gladys Rivera, José Sánchez and Pedro Contreras for the onsite and logistic support. At the same time, this research was supported by the project Fondecyt 1151336 to MH.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Hüne, M., Vega, R. Feeding habits in two sympatric species of Notothenioidei, Patagonotothen cornucola and Harpagifer bispinis, in the Chilean Patagonian channels and fjords. Polar Biol 39, 2253–2262 (2016). https://doi.org/10.1007/s00300-016-1892-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-016-1892-3