
Abstract
The food and feeding ecology of albatrosses during the nonbreeding season is still poorly known, particularly with regard to the cephalopod component. This was studied in black-browed Thalassarche melanophris and grey-headed T. chrysostoma albatrosses by analysing boluses collected shortly after adults returned to colonies at Bird Island, South Georgia (54°S, 38°W), in 2009. Based on stable isotopic analyses of the lower beaks, we determined the habitat and trophic level (from δ13C and δ15N, respectively) of the most important cephalopods and assessed the relative importance of scavenging in terms of the albatrosses’ feeding regimes. Based on lower rostral lengths (LRLs), the main cephalopod species in the diets of both albatrosses was Kondakovia longimana, by frequency of occurrence (F > 90 %), number (N > 40 %) and mass (M > 80 %). The large estimated mass of many squid, including K. longimana, suggests that a high proportion (>80 % by mass) was scavenged, and that scavenging is much more important during the nonbreeding season than would be expected from breeding-season diets. The diversity of cephalopods consumed by nonbreeding birds in our study was similar to that recorded during previous breeding seasons, but included two new species [Moroteuthis sp. B (Imber) and ?Mastigoteuthis A (Clarke)]. Based on similarities in LRL, δ13C and δ15N, the squid consumed may have been from the same oceanic populations or region, with the exception of Taonius sp. B (Voss) and K. longimana, which, based on significant differences in δ15N values, suggest that they may have originated from different stocks, indicating differences in the albatrosses’ feeding regimes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Albatrosses are regarded as apex predators in subantarctic and Antarctic ecosystems, feeding on a wide diversity of prey, including cephalopods (Xavier and Cherel 2009). These molluscs play an important role in the ecology of the Southern Ocean, as key links in the food web between abundant mesopelagic fish and crustaceans, and higher predators such as albatrosses and marine mammals (Collins and Rodhouse 2006). Although free-living cephalopods in the Southern Ocean are elusive, which limits opportunities for ship-based studies, albatrosses can be used as biological sampling tools; the tracking and diet sampling of these marine birds improve our knowledge not only of their foraging behaviour but also of the distribution and ecology of their cephalopod prey (Xavier et al. 2006).
Albatrosses cover vast distances when foraging during the breeding and nonbreeding periods (Nel et al. 2001; Phillips et al. 2004; Xavier et al. 2004; Croxall et al. 2005). Black-browed (Thalassarche melanophris) and grey-headed (T. chrysostoma) albatrosses nest in dense colonies on subantarctic islands, including at South Georgia, which hold the largest grey-headed, and third largest black-browed albatross populations, respectively, in the world (Poncet et al. 2006). Tracking data from Bird Island indicate that during the chick-rearing period (January to June), both species forage mainly in Antarctic and subantarctic waters (Xavier et al. 2003b; Phillips et al. 2004). During the nonbreeding season, most black-browed albatrosses from South Georgia migrate to waters off southern Africa, and a small minority to the Patagonian Shelf or Australasia (Phillips et al. 2005). There is even greater individual variation in migration strategies of grey-headed albatrosses. Although most birds utilise oceanic waters, they may remain entirely in the southwest Atlantic or spend varying proportions of time in the Atlantic, Indian and Pacific oceans and can make one or two circumpolar migrations around the Antarctic continent (Croxall et al. 2005). Stable isotope analyses of feathers confirm that black-browed albatrosses from South Georgia moult in productive neritic waters of the Benguela Current during the nonbreeding period, but that grey-headed albatrosses moult in subantarctic waters, near the subtropical front (Phillips et al. 2009; Cherel et al. 2013).
Previous studies of black-browed and grey-headed albatrosses during the chick-rearing period at South Georgia highlighted the considerable annual variation in dietary components, although black-browed albatrosses typically fed on crustaceans, cephalopods and fish (36–40, 31 and 27–35 %, respectively, of the diet by mass), and grey-headed albatrosses on cephalopods (50–55 % by mass) and, to a lesser extent, lamprey Geotria australis (10 % by mass) and other prey (Prince 1980; Rodhouse and Prince 1993; Xavier et al. 2003a, 2013). Although many live prey are obtained by plunge diving (Cherel and Klages 1998), a number of the cephalopods (comprising as much as 13–14 % of the estimated total mass of all prey) were potentially obtained by scavenging (Xavier and Croxall 2007). Prior to the present study, the species composition of the diet of most albatrosses during the nonbreeding period was unknown due to the difficulties of sampling birds that spend their entire time at sea. Nonetheless, stable isotope analyses of feathers indicated that grey-headed and black-browed albatrosses fed, respectively, at low to mid, or at high trophic levels, within the Southern Ocean (Phillips et al. 2009).
The aims of the present study were to (1) investigate the cephalopod component of the diet of black-browed and grey-headed albatrosses at the end of the nonbreeding period, (2) estimate the size of individual squid to assess the relative importance of scavenging versus predation and (3) determine the habitat and trophic level of the most important cephalopod species using stable isotope analyses. Diet composition was based on analysis of boluses (pellets or casts) regurgitated voluntarily by adult albatrosses shortly after they returned to South Georgia to breed, at the end of the austral winter. Each bolus contains accumulated prey items (mainly cephalopod beaks), consumed in the latter part of the nonbreeding period, almost certainly over a period of several weeks given the long residency time of squid beaks in the stomach of seabirds recorded in previous studies (Furness et al. 1984). The primary advantages of analysing boluses include the ease of collection and minimal disturbance of birds, since handling is not required (Xavier et al. 2005). Stable isotope ratios were analysed in lower beaks found in these boluses to determine the habitat (δ13C) and relative trophic level (δ15N) of the squid, the former based on the negative latitudinal gradient in δ13C in the Southern Ocean (Cherel and Hobson 2005). Hence, δ13C values indicate water mass (subtropical vs. subantarctic or Antarctic), and higher vs. lower values for δ15N reflect the relative dependency on fish or squid compared with crustaceans (Cherel and Hobson 2007). Cephalopod beaks are hard structures which grow by accretion of proteins and chitin, and there is no turnover after synthesis. Consequently, they retain molecules built up from early development to time of death, and their isotopic signature integrates the feeding ecology of the animal over its whole life (Cherel and Hobson 2005).
Materials and methods
Sampling
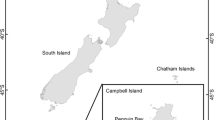
Boluses, regurgitated by adult black-browed and grey-headed albatrosses that had recently arrived at colonies at Bird Island, South Georgia (54°00′S 38°03′W), were collected from the ground during daily visits from September to December 2009 (Fig. 1; Table 1). All samples were either identified and measured at Bird Island or frozen at −20 °C and analysed at the British Antarctic Survey (BAS) headquarters (Cambridge, UK) or the Marine and Environmental Research Centre (MARE) of the University of Coimbra (Coimbra, Portugal). The components of the boluses (mostly indigestible items such as cephalopod beaks, cephalopod spermatophores, salps and penguin feathers; Xavier et al. 2003c) were identified to species level when possible. As seabirds retain squid beaks in the fore-gut for considerable periods (Furness et al. 1984), these beaks represent cephalopods consumed in the final weeks of the nonbreeding period. No fish or crustacean remains were recorded. Cephalopod beaks were separated into upper and lower, and the former were counted and discarded. The lower beaks were cleaned, counted and identified whenever possible to species level, and the lower rostral length (LRL) measured using vernier calipers to the nearest 0.1 mm (Xavier and Cherel 2009). Allometric equations were used to estimate dorsal mantle length (ML, mm) and the original wet body mass (M, g) from LRL using Xavier and Cherel (2009), Piatkowski et al. (2001), Clarke (1986), Lu and Williams (1994), Brown and Klages (1987), Rodhouse and Yeatman (1990), Rodhouse et al. (1990) and Cherel, unpublished data. The equations for Mastigoteuthis psychrophila were used for ?Mastigoteuthis A (Clarke) because there are no specific equations for the latter (Xavier and Cherel 2009) based on British Antarctic Survey, unpublished data.

Map of the Southern Ocean indicating the sampling site (Bird Island, South Georgia), and the 1,000 m isobath
Albatross diet composition was expressed in terms of the frequency of occurrence (F; number of samples with that cephalopod species/total number of samples), total number of lower beaks per cephalopod species (N), lower rostral lengths [LRL; mean, standard deviation (SD) and range], estimated mantle lengths [ML; total, mean, standard deviation (SD) and range] and estimated mass [M; total, mean, standard deviation (SD) and range]. The scavenging levels (cephalopods were put into 500 g categories) followed Croxall and Prince (1994).
Stable isotope analyses
Lower beaks were cleaned and preserved in 70 % ethanol, dried subsequently in an oven at 50 °C for 6–24 h to drive off the ethanol, reduced to a fine powder, and then part of the homogenised sample (0.30–0.55 mg) was encapsulated for stable isotope analysis (SIA). SIA was carried out only on cephalopod species represented by at least six lower beaks in samples from either species, with the exception of Taonius sp. B (Voss) of which there were four lower beaks in boluses from grey-headed albatrosses. Stable isotope ratios (δ15N and δ13C) were measured using a Continuous Flow Isotope Ratio Mass Spectrometer (CFIRMS) at MARE. The results are presented in δ notation as deviations from the standard references in parts per thousand (‰) according to the following equation: δ X = [(R sample/R standard) −1] × 1,000, where X represents 13C or 15N and Rsample the ratios 13C/12C or 15N/14N. Rstandard represents the international reference standard V-PDB (“Vienna”—PeeDee formation), and atmospheric N2 (AIR) is the standard for δ13C and δ15N, respectively.
The stable isotope ratios of cephalopod beaks were compared with those in feathers collected from black-browed and grey-headed albatrosses in January 2002, which represent diet during the preceding moulting (nonbreeding) period, i.e. austral winter 2001 (values reported in Phillips et al. 2009). To compare with results from previous studies, values for cephalopod lower beaks were converted into those expected for cephalopod muscle, and values for albatross feathers were converted into those expected for blood (Stowasser et al. 2012), by taking account of different isotopic discrimination factors for each tissue. SI ratios in cephalopod muscle (i.e. the bulk of tissue ingested by the albatross) were assumed to be 4.86 ‰ higher in δ15N and 0.75 ‰ lower in δ13C, than lower beaks (means for five species) (Hobson and Cherel 2006). Blood of black-browed and grey-headed albatrosses was assumed, respectively, to be 1.99 and 1.95 ‰ lower in δ13C, and 0.43 and 0.26 ‰ higher in δ15N than feathers (Cherel et al. 2014). Differences in tissue-to-diet discrimination factors are explained by tissue-specific biochemical composition, namely the composition of chitin and beak proteins (Schimmelmann and DeNiro 1988; Hobson and Cherel 2006), and amino acids, and lipid content (Wolf et al. 2009; Cherel et al. 2014).
Statistical analyses
LRL and stable isotope ratios of the cephalopod species recorded in the diet of both black-browed and grey-headed albatrosses were compared using T tests or Mann–Whitney U tests. Mean δ13C and δ15N of lower cephalopod beaks from black-browed and grey-headed albatrosses were compared using one-way ANOVA or Kruskal–Wallis tests. All statistical tests were carried out using Statistica version 10. Statistical significance was taken as P < 0.05.
Results
Black-browed albatross
A total of 17 cephalopod species were identified among the 115 lower beaks found in the 14 boluses collected from black-browed albatrosses at the end of the nonbreeding period (Table 1). The lower beaks belonged to adult and subadult squid (i.e. there were no beaks from juvenile squid). The most important by mass was Kondakovia longimana (F = 100 %, N = 40.9 %, M = 80.3 %; Tables 2, 3), followed by Moroteuthis knipovitchi and Alluroteuthis antarcticus. The longest rostrum was recorded in K. longimana and the shortest in Nototeuthis dimegacotyle. Estimated mantle lengths were longer in Taonius sp. B (Voss), followed by K. longimana and Galiteuthis glacialis, and shorter in Histioteuthis macrohista and Histioteuthis bonnellii corpuscula. The heaviest estimated squid caught by black-browed albatrosses was K. longimana (M = 4,027 g), and the lightest was Batoteuthis skolops (M = 23 g).
Assuming that squid heavier than 500 g were scavenged, 82.7 % by estimated mass of cephalopods in the diet of nonbreeding black-browed albatrosses were potentially scavenged (Table 4). This dropped to only 4.8 % by estimated mass that were scavenged using a cut-off value of 3,500 g (Table 4). The cephalopod species that included individuals with an estimated mass >500 g were K. longimana (one individual at >3,500 g), M. knipovitchi and A. antarcticus (Table 3).
There were significant differences in both δ13C and δ15N values among the five most common cephalopod species in the diet of black-browed albatrosses (Table 5; Fig. 2). δ13C values differed significantly only between M. knipovitchi and G. glacialis (Kruskal–Wallis test, H 42 = 12.1, P = 0.02), whereas δ15N values were lower in samples of K. longimana, G. glacialis and M. knipovitchi (values in these three taxa overlapped), and higher in Gonatus antarcticus and Taonius sp. B (Voss) (ANOVA, F (4,37) = 25.7, P < 0.01; Table 5; Fig. 2).

Adjusted stable carbon (δ13C) and nitrogen (δ15N) isotope ratios of lower beaks of cephalopod species found in the diet of a grey-headed and b black-browed albatrosses at the end of the nonbreeding period in 2009, and in feathers from eight males and eight females of the same albatross species grown in winter 2001 (Phillips et al. 2009). Conversion factors from Hobson and Cherel (2006) and Cherel et al. (2014). Bars represent SD
Grey-headed albatross
A total of 16 cephalopod species were identified among the 321 lower beaks found in the 32 boluses collected from grey-headed albatrosses at the end of the nonbreeding period (Table 1). The most important by mass was K. longimana (F = 90.6 %, N = 40.5 %, M = 90.5 %; Tables 2, 3), followed by G. antarcticus. The longest rostrum belonged to K. longimana, and the shortest rostrum belonged to Martialia hyadesi. The longest estimated mantle length was from Taonius sp. B (Voss), followed by K. longimana, and the shortest estimated mantle length was from H. macrohista, followed by H. b. corpuscula. The heaviest and the lightest estimated squid caught by grey-headed albatrosses were K. longimana (M = 3,632 and 10 g, respectively).
Assuming that squid heavier than 500 g were scavenged, a total of 92.1 % by estimated mass were potentially scavenged by grey-headed albatrosses during the later nonbreeding period (Table 4). This dropped to only 1.5 % by estimated mass that was scavenged using a cut-off value of 3,500 g (Table 4). The cephalopod species that included individuals with estimated mass >500 g were K. longimana (one individual at >3,500 g), G. antarcticus, A. antarcticus, Mesonychoteuthis hamiltoni and Todarodes sp. (Table 3).
There were significant differences in both mean δ13C and δ15N among the eight cephalopod species that were analysed (Table 5; Fig. 2). δ13C values were lower, and broadly overlapped, in B. skolops and K. longimana, and were higher in Histioteuthis atlantica, H. macrohista and M. hyadesi (Kruskal–Wallis test, H 69 = 53, P < 0.01). δ15N values were lower (and broadly overlapped), in K. longimana, G. glacialis and M. hyadesi, than in the other five species (ANOVA, F (4,37) = 25.7, P < 0.01; Table 5; Fig. 2).
Comparison of both albatrosses cephalopod component
Results of statistical tests (t tests or Mann–Whitney tests) comparing the lower rostral length, δ13C and δ15N of cephalopod species recorded in the diet of black-browed and grey-headed albatrosses are shown in Table 6. Comparisons of the adjusted isotopic values of cephalopod lower beaks and albatrosses feathers using Kruskal–Wallis tests are as follows. There was a significant difference in estimated δ13C values among feathers of black-browed albatrosses and lower beaks of all sampled cephalopod species (Kruskal–Wallis test, H 58 = 40.4, P < 0.01), and in estimated δ15N values among feathers and lower beaks of K. longimana and G.glacialis (Kruskal–Wallis test, H 58 = 41.0, P < 0.01). There was also a significant difference in estimated δ13C among feathers of grey-headed albatrosses and lower beaks of K. longimana and B. skolops (Kruskal–Wallis test, H 85 = 48.5, P < 0.01), and in estimated δ15N values among feathers and lower beaks of G. antarcticus, H. macrohista and H. atlantica (Kruskal–Wallis test, H 85 = 68.7, P < 0.01).
Discussion
Dietary items from boluses correspond to the prey taken over the last few weeks or months (Furness et al. 1984; Xavier et al. 2005). As they only include material that is difficult to digest, such as cephalopod beaks, boluses provide an incomplete picture of the overall diet. Moreover, most adult and some subadult beaks from boluses can be measured, because their wings are wholly darkened and resist digestion. In contrast, most subadult and juvenile beaks have no wings and thus cannot be measured. This was a particular problem for M. hyadesi in the diet of grey-headed albatrosses. Thus, there are two caveats associated with our approach: (1) considerable overestimation of mean size and mass for some species because only LRL from adult beaks was available (80 % of M. hyadesi’s subadult beaks were eroded; Table 2), and (2) a potentially huge underestimation of the importance by mass of M. hyadesi in the diet.
To improve the assessment of the contribution of different cephalopods to predator diets, both lower and upper beaks should be examined as the ratio frequently differs from parity (Xavier et al. 2011). However, as the difference was slight (<10 %) in our study (Table 1), there was no need to analyse upper beaks. Additionally, there are fewer descriptions of upper beak morphology which makes identification more difficult, and for many species, no allometric equations exist for estimating mass from measurements.
Cephalopods in albatross diets
There were no significant differences in LRL between the same cephalopod species recorded in the diet of the black-browed and grey-headed albatrosses, which could mean that adults of both species feed on the same squid populations at the end of their nonbreeding period (Table 6). The main cephalopod species represented in diets of both albatrosses was K. longimana, by frequency of occurrence, number and mass (Table 2). These percentages were higher than those reported for the breeding period (see Xavier et al. 2003a; British Antarctic Survey unpubl data). K. longimana is an important prey of numerous deep-diving predators including seals, fish (Patagonian toothfish Dissostichus eleginoides, Antarctic toothfish D. mawsoni and sleeper sharks Somniosus cf. microcephalus) and cetaceans (Collins and Rodhouse 2006). Thus, here we show that these adult and subadult larger squids are also available to nonbreeding black-browed and grey-headed albatrosses.
During the breeding period, both black-browed and grey-headed albatrosses feed mostly on live prey, including squid (only 25–64 % by mass potentially scavenged) (Croxall and Prince 1994; Xavier and Croxall 2007). In contrast, towards the end of the nonbreeding period, a much higher proportion of the squid were obtained by scavenging (>80 % by estimated mass; Table 4). A comparison of the sizes of squid caught by albatrosses from different populations indicates that nonbreeding black-browed and grey-headed albatrosses from South Georgia caught individuals of K. longimana that were of similar sizes to those consumed during the breeding season at Diego Ramirez, Prince Edward, Crozet and Kerguelen islands (Table 7). Similarly, nonbreeding grey-headed albatrosses from South Georgia caught individuals of M. hyadesi that were of similar estimated sizes to those consumed during the breeding season at Prince Edward and Crozet islands. This presumably reflects the very large foraging range during the nonbreeding period (Croxall et al. 2005; Phillips et al. 2005), and the availability and circumpolar distribution of this major cephalopod prey (Xavier and Cherel 2009). All M. hyadesi in the diet of nonbreeding grey-headed albatrosses were subadults (according to the lower beaks) and were <1 year old (average estimated size <200 mm)(González et al. 1997), whereas those individuals caught at Kerguelen and Campbell islands were older (average estimated size >200 mm). This supports the hypothesis that the spawning of M. hyadesi occurs somewhere in the north of the Scotia Sea, in subantarctic waters (e.g. Patagonian shelf slope), and that their eggs, paralarvae, juveniles and subadults are carried eastwards by the prevailing Antarctic Circumpolar Current System (Rodhouse 1997). In contrast, G. antarcticus in the diet of the nonbreeding grey-headed albatrosses were larger than those consumed during breeding at other locations, and included the biggest G. antarcticus specimen found so far, with a maximum mantle length of 424 mm (Table 3). This suggests that grey-headed albatrosses may feed on Gonatid squid that die after major spawning events during the winter (Xavier and Croxall 2007).
The relatively high δ15N in black-browed albatross tissues suggest that their diet during the nonbreeding season includes many of the same squid species, including G.antarcticus, Taonius sp. B (Voss) and M. knipovitchi, and potentially other types of prey also at relatively high trophic levels, including fish (Cherel et al. 2000a; Xavier et al. 2003a; Stowasser et al. 2012). It is not appropriate to compare δ13C in the squid beaks with that in feathers of black-browed albatrosses, as the latter moults in the Benguela Upwelling region (Phillips et al. 2009), whereas the cephalopods were obtained later in the nonbreeding season, when the birds would have been foraging in subantarctic waters, en route back to the colony (Phillips et al. 2005). In contrast, grey-headed albatrosses moult while in subantarctic waters (Connan et al. 2014), and values for δ13C in this species overlapped with those of many of their cephalopod prey in the samples analysed in this study (Fig. 2); hence, there is no evidence of a shift in their foraging habitat. Moreover, the relatively low δ15N in tissues of the grey-headed albatross confirms that their diet during the nonbreeding season includes predominantly low trophic-level prey, presumably including squid such as M. hyadesi, K. longimana and G. glacialis, and potentially crustaceans, jellyfish or salps (Connan et al. 2014).
Insights into cephalopods distribution
The diversity of cephalopods among the beaks recorded in this study (17 and 16 species, respectively, in samples from black-browed and grey-headed albatross) was similar to that recorded in combined samples of fresh and accumulated beaks collected in previous breeding seasons (6–16 and 10–19 cephalopod species in the diet of black-browed and grey-headed albatrosses, respectively) (see Xavier et al. 2003a; Xavier and Croxall 2007; unpubl data).
Nevertheless, Moroteuthis sp. B (Imber) was recorded for the first time in the diet of the nonbreeding black-browed and grey-headed albatrosses (Table 2). The distribution of this squid may therefore extend into the south Atlantic, but beyond the range of breeding birds from South Georgia, from its known distribution in the southern Indian Ocean, where it is a regular prey of Patagonian toothfish and king penguins Aptenodytes patagonicus (Cherel and Weimerskirch 1999; Cherel et al. 2004). ? Mastigoteuthis A (Clarke) was for the first time documented in the diet of the grey-headed albatross, but is known to occur in that of wandering albatrosses from South Georgia (Xavier et al. 2011). This species is possibly a synonym of the recently described chiroteuthid squid Asperoteuthis nesisi (Arkhipkin and Laptikhovsky 2008).
Of the other important squid species, only A. antarcticus in black-browed albatross diet and G. antarcticus in grey-headed albatross diet were caught in greater numbers by nonbreeding birds than during the breeding period, although not in terms of mass (Xavier et al. 2003a; British Antarctic Survey unpublished data). M. hyadesi was caught less often by grey-headed albatrosses than during previous breeding seasons, although not in terms of overall proportion, but it represented a smaller proportion of the total estimated mass as 80 % of the beaks were from subadult squid and too eroded to measure (see Xavier et al. 2003a; British Antarctic Survey unpubl data). The frequency of occurrence, overall proportion and mass of M. knipovitchi in the diet of black-browed albatrosses were similar to that recorded during their breeding period (see Xavier et al. 2003a; British Antarctic Survey unpubl data).
Carbon and nitrogen isotopic values of the lower beaks matched with previous studies (Cherel and Hobson 2005; Stowasser et al. 2012). Based on δ13C and δ15N values, squid consumed by both black-browed and grey-headed albatrosses may have been from the same oceanic populations and regions. Only δ15N values in Taonius sp. B (Voss) and K. longimana differed significantly in samples from the two albatross species (Table 6), suggesting different feeding regimes or origins from different stocks. However, those discrepancies could reflect individual preferences of albatrosses, which show high fidelity to foraging sites used during the nonbreeding season (Phillips et al. 2005). Individual specialisation is also apparent in stable isotope ratios in tissues sampled from birds at this colony, and reflects variation in relative species contribution to diets, and some degree of sexual segregation (Phillips et al. 2011).
The community of cephalopods living in the waters used by the albatrosses towards the end of the nonbreeding period in effect occupy only two trophic levels (Fig. 2), one trophic level less than that of the cephalopod community around the Kerguelen Islands (Cherel and Hobson 2005). The likely explanation is that there were no isotopic data from the colossal squid, the apex predator in Kerguelen waters, which features very rarely in the diet of grey-headed albatross at South Georgia (grey-headed albatross diet in Table 2; Xavier et al. 2003a). Overall, the South Georgia community showed higher δ15N and lower δ13C values than the cephalopod community of Kerguelen (>1.5 ‰ and 0.4 ‰, respectively, for M. knipovitchi, K. longimana, G. antarcticus and M. hyadesi) (Cherel and Hobson 2005; Cherel et al. 2011). The isotopic variation can be explained by differences in the base of the food web (δ15N) and by the location of South Georgia in Antarctic waters, whereas the Kerguelen Islands lie further north, in the Polar Frontal Zone (δ13C). There is a stepwise increase in δ13C by ~0.5–1.0 ‰ and in δ15N by ~3–5 ‰ with trophic level, and, in the Southern Ocean, δ13C differs among water masses, decreasing from neritic to oceanic, and benthic to pelagic habitats, and from north to south (Cherel and Hobson 2007; Phillips et al. 2009). Stable isotope ratios in the albatross tissues will integrate those of all types of prey consumed, not just the cephalopod component. As different environmental factors affect stable isotope signatures of particulate organic matter (POM; Stowasser et al. 2012), values in predator tissues will vary accordingly.
In conclusion, the main cephalopod species represented in diets of both nonbreeding black-browed and grey-headed albatrosses was K. longimana. The large estimated mass of many of the squid in this study suggest that a higher proportion were scavenged (>80 % by estimated mass) than during the breeding season. The diversity of cephalopods was similar to that recorded for these two predator species during previous breeding seasons. Nevertheless, there were two new species, and other differences in species contributions by mass and number. Based on LRL, δ13C and δ15N values, squid consumed by both black-browed and grey-headed albatrosses may have been from the same oceanic populations or region, except for Taonius sp. B (Voss) and K. longimana. The community of cephalopods in the waters used by the albatrosses towards the end of the nonbreeding period spanned only two trophic levels. Our results therefore provide new insights into albatross diet and ecology during this little known period of their life cycle. Future work should be focused on (1) comparison of boluses obtained from birds tracked using loggers to relate squid in the diet to individual nonbreeding distribution; and (2) concurrent deployment of bird-borne cameras and stomach temperature probes or other devices to determine time of ingestion, mass and identity of the squid prey.
References
Arata J, Xavier JC (2003) The diet of black-browed albatrosses at the Diego Ramirez Islands, Chile. Polar Biol 26:638–647. doi:10.1007/s00300-003-0530-z
Arata J, Robertson G, Valencia J, Xavier JC, Moreno CA (2004) Diet of grey-headed albatrosses at the Diego Ramírez Islands, Chile: ecological implications. Antarct Sci 16:263–275. doi:10.1017/S095410200400207X
Arkhipkin AI, Laptikhovsky VV (2008) Discovery of the fourth species of the enigmatic chiroteuthid squid Asperoteuthis (Cephalopoda: Oegopsida) and extension of the range of the genus to the South Atlantic. J Molluscan Stud 74:203–207. doi:10.1093/mollus/eyn007
Brown CR, Klages NT (1987) Seasonal and annual variation in diets of Macaroni (Eudyptes chrysolophus chrysolophus) and Southern rockhopper (E. chrysocome chrysocome) penguins at sub-Antarctic Marion Island. J Zool 212:7–28. doi:10.1111/j.1469-7998.1987.tb05111.x
Cherel Y, Hobson KA (2005) Stable isotopes, beaks and predators: a new tool to study the trophic ecology of cephalopods, including giant and colossal squids. Proc R Soc Lond B Biol Sci 272:1601–1607. doi:10.1098/rspb.2005.3115
Cherel Y, Hobson KA (2007) Geographical variation in carbon stable isotope signatures of marine predators: a tool to investigate their foraging areas in the Southern Ocean. Mar Ecol Prog Ser 329:281–287. doi:10.3354/meps329281
Cherel Y, Klages N (1998) A review of the food of albatrosses. In: Robertson G, Gales R (eds) Albatross Biology and Conservation. Surrey Beatty & Sons, Chipping Norton, pp 113–136
Cherel Y, Weimerskirch H (1999) Spawning cycle of onychoteuthid squids in the southern Indian Ocean: new information from seabird predators. Mar Ecol Prog Ser 188:93–104. doi:10.3354/meps188093
Cherel Y, Hobson KA, Weimerskirch H (2000a) Using stable-isotope analysis of feathers to distinguish moulting and breeding origins of seabirds. Oecologia 122:155–162. doi:10.1007/pl00008843
Cherel Y, Weimerskirch H, Trouve C (2000b) Food and feeding ecology of the neritic-slope forager black-browed albatross and its relationships with commercial fisheries in Kerguelen waters. Mar Ecol Prog Ser 207:183–199. doi:10.3354/meps207183
Cherel Y, Weimerskirch H, Trouve C (2002) Dietary evidence for spatial foraging segregation in sympatric albatrosses (Diomedea spp.) rearing chicks at Iles Nuageuses, Kerguelen. Mar Biol 141:1117–1129. doi:10.1007/s00227-002-0907-5
Cherel Y, Duhamel G, Gasco N (2004) Cephalopod fauna of subantarctic islands: new information from predators. Mar Ecol Prog Ser 266:143–156. doi:10.3354/meps266143
Cherel Y, Gasco N, Duhamel G (2011) Top predators and stable isotopes document the cephalopod fauna and its trophic relationships in Kerguelen waters. In: Duhamel G, Welsford D (eds) The Kerguelen Plateau: marine ecosystem and fisheries, Proceedings of the 1st international Science Symposium on the Kerguelen Plateau. Société Française d’Ichtyologie, Abbeville, pp 99–108
Cherel Y, Jaeger A, Alderman R, Jaquemet S, Richard P, Wanless RM, Phillips RA, Thompson DR (2013) A comprehensive isotopic investigation of habitat preferences in nonbreeding albatrosses from the Southern Ocean. Ecography 36:277–286. doi:10.1111/j.1600-0587.2012.07466.x
Cherel Y, Jaquemet S, Maglio A, Jaeger A (2014) Differences in δ13C and δ15N values between feathers and blood of seabird chicks: implications for non-invasive isotopic investigations. Mar Biol 161:229–237. doi:10.1007/s00227-013-2314-5
Clarke MR (1986) A handbook for the identification of cephalopod beaks. Clarendon Press, Oxford
Collins MA, Rodhouse PG (2006) Southern Ocean Cephalopods. In: Alan J. Southward CMY, Lee AF (eds) Adv Mar Biol. Academic Press, pp 191–265
Connan M, McQuaid C, Bonnevie B, Smale M, Cherel Y, Klages N (2014) Combined stomach content, lipid and stable isotope analyses reveal spatial and trophic partitioning among three sympatric albatrosses from the Southern Ocean. Mar Ecol Prog Ser 497:259–272. doi:10.3354/meps10606
Croxall JP, Prince PA (1994) Dead or alive, night or day: how do albatrosses catch squid? Antarct Sci 6:155–162. doi:10.1017/S0954102094000246
Croxall JP, Prince PA (1996) Cephalopods as Prey. I. Seabirds. Philos Trans R Soc Lond Ser B Biol Sci 351:1023–1043. doi:10.1098/rstb.1996.0091
Croxall JP, Silk JR, Phillips RA, Afanasyev V, Briggs DR (2005) Global circumnavigations: tracking year-round ranges of nonbreeding albatrosses. Science 307:249–250. doi:10.1126/science.1106042
Furness BL, Laugksch RC, Duffy DC (1984) Cephalopod beaks and studies of seabird diets. Auk 101:619–620
González AF, Trathan PN, Yau C, Rodhouse PG (1997) Interactions between oceanography, ecology and fishery biology of the ommastrephid squid Martialia hyadesi in the South Atlantic. Mar Ecol Prog Ser 152:205–215. doi:10.3354/meps152205
Hobson KA, Cherel Y (2006) Isotopic reconstruction of marine food webs using cephalopod beaks: new insight from captively raised Sepia officinalis. Can J Zool 84:766–770. doi:10.1139/z06-049
Lu CC, Williams R (1994) Contribution to the biology of squid in the Prydz Bay region, Antarctica. Antarct Sci 6:223–229. doi:10.1017/S0954102094000349
Nel DC, Lutjeharms JRE, Pakhomov EA, Ansorge IJ, Ryan PG, Klages NTW (2001) Exploitation of mesoscale oceanographic features by grey-headed albatross Thalassarche chrysostoma in the southern Indian Ocean. Mar Ecol Prog Ser 217:15–26. doi:10.3354/meps217015
Phillips RA, Silk JR, Phalan B, Catry P, Croxall JP (2004) Seasonal sexual segregation in two Thalassarche albatross species: competitive exclusion, reproductive role specialization or foraging niche divergence? Proc R Soc Lond B Biol Sci 271:1283–1291. doi:10.1098/rspb.2004.2718
Phillips RA, Silk JRD, Croxall JP, Afanasyev V, Bennett VJ (2005) Summer distribution and migration of nonbreeding albatrosses: individual consistencies and implications for conservation. Ecology 86:2386–2396. doi:10.1890/04-1885
Phillips R, Bearhop S, McGill RR, Dawson D (2009) Stable isotopes reveal individual variation in migration strategies and habitat preferences in a suite of seabirds during the nonbreeding period. Oecologia 160:795–806. doi:10.1007/s00442-009-1342-9
Phillips R, McGill RR, Dawson D, Bearhop S (2011) Sexual segregation in distribution, diet and trophic level of seabirds: insights from stable isotope analysis. Mar Biol 158:2199–2208. doi:10.1007/s00227-011-1725-4
Piatkowski U, Pütz K, Heinemann H (2001) Cephalopod prey of king penguins (Aptenodytes patagonicus) breeding at Volunteer Beach, Falkland Islands, during austral winter 1996. Fish Res 52:79–90. doi:10.1016/S0165-7836(01)00232-6
Poncet S, Robertson G, Phillips RA, Lawton K, Phalan B, Trathan PN, Croxall JP (2006) Status and distribution of wandering, black-browed and grey-headed albatrosses breeding at South Georgia. Polar Biol 29:772–781. doi:10.1007/s00300-006-0114-9
Prince PA (1980) The food and feeding ecology of grey-headed albatross Diomedea chrysostoma and black-browed albatross D. melanophris. Ibis 122:476–488. doi:10.1111/j.1474-919X.1980.tb00902.x
Richoux N, Jaquemet S, Bonnevie B, Cherel Y, McQuaid C (2010) Trophic ecology of grey-headed albatrosses from Marion Island, Southern Ocean: insights from stomach contents and diet tracers. Mar Biol 157:1755–1766. doi:10.1007/s00227-010-1448-y
Rodhouse PG (1997) Precautionary measures for a new fishery on Martialia hyadesi (Cephalopoda, Ommastrephidae) in the Scotia Sea: an ecological approach. CCAMLR Sci 4:125–139
Rodhouse PG, Prince PA (1993) Cephalopod prey of the black-browed albatross Diomedea melanophrys at South Georgia. Polar Biol 13:373–376. doi:10.1007/bf01681978
Rodhouse PG, Yeatman J (1990) Redescription of Martialia hyadesi Rochebrune and Mabille, 1889 (Mollusca: Cephalopoda) from the Southern Ocean. Bull Br Mus Nat Hist (Zool) 56:135–143
Rodhouse PG, Prince PA, Clarke MR, Murray AWA (1990) Cephalopod prey of the grey-headed albatross Diomedea chrysostoma. Mar Biol 104:353–362. doi:10.1007/bf01314337
Schimmelmann A, DeNiro MJ (1988) Stable isotope studies on Chitin. II. The 13C/12C and 15N/14N ratios in arthropod chitin. Contrib Mar Sci 29:113–130
Stowasser G, Atkinson A, McGill RAR, Phillips RA, Collins MA, Pond DW (2012) Food web dynamics in the Scotia Sea in summer: a stable isotope study. Deep Sea Res Part 2 Top Stud Oceanogr 59–60:208–221. doi:10.1016/j.dsr2.2011.08.004
Waugh SM, Weimerskirch H, Cherel Y, Shankar U, Prince PA, Sagar PM (1999) Exploitation of the marine environment by two sympatric albatrosses in the Pacific Southern Ocean. Mar Ecol Prog Ser 177:243–254. doi:10.3354/meps177243
Wolf N, Carleton SA, Martínez del Rio C (2009) Ten years of experimental animal isotopic ecology. Funct Ecol 23:17–26. doi:10.1111/j.1365-2435.2009.01529.x
Xavier JC, Cherel Y (2009) Cephalopod beak guide for the Southern Ocean. British Antarctic Survey, Cambridge
Xavier JC, Croxall JP (2007) Predator-prey interactions: why do larger albatrosses eat bigger squid? J Zool 271:408–417. doi:10.1111/j.1469-7998.2006.00224.x
Xavier JC, Croxall JP, Reid K (2003a) Interannual variation in the diets of two albatross species breeding at South Georgia: implications for breeding performance. Ibis 145:593–610. doi:10.1046/j.1474-919X.2003.00196.x
Xavier JC, Croxall JP, Trathan PN, Rodhouse PG (2003b) Interannual variation in the cephalopod component of the diet of the wandering albatross, Diomedea exulans breeding at Bird Island, South Georgia. Mar Biol 142:611–622. doi:10.1007/s00227-002-0962-y
Xavier JC, Croxall JP, Trathan PN, Wood AG (2003c) Feeding strategies and diets of breeding grey-headed and wandering albatrosses at South Georgia. Mar Biol 143:221–232. doi:10.1007/s00227-003-1049-0
Xavier JC, Trathan PN, Croxall JP, Wood AG, Podestá G, Rodhouse PG (2004) Foraging ecology and interactions with fisheries of wandering albatrosses (Diomedea exulans) breeding at South Georgia. Fish Oceanogr 13:324–344. doi:10.1111/j.1365-2419.2004.00298.x
Xavier JC, Croxall JP, Cresswell KA, Burger AE (2005) Boluses: an effective method for assessing the proportions of cephalopods in the diet of albatrosses. The Auk 122:1182–1190. doi:10.1642/0004-8038(2005)122[1182:BAEMFA]2.0.CO;2
Xavier JC, Tarling GA, Croxall JP (2006) Determining prey distribution patterns from stomach-contents of satellite-tracked high-predators of the Southern Ocean. Ecography 29:260–272. doi:10.1111/j.2006.0906-7590.04525.x
Xavier JC, Phillips RA, Cherel Y (2011) Cephalopods in marine predator diet assessments: why identifying upper and lower beaks is important. ICES J Mar Sci 68:1857–1864. doi:10.1093/icesjms/fsr103
Xavier JC, Louzao M, Thorpe SE, Ward P, Hill C, Roberts D, Croxall JP, Phillips RA (2013) Seasonal changes in the diet and feeding behaviour of a top predator indicate a flexible response to deteriorating oceanographic conditions. Mar Biol 160:1597–1606. doi:10.1007/s00227-013-2212-x
Acknowledgments
We are grateful to Derren Fox, Stacey Adlard and Ewan Edwards at Bird Island research station for helping with sample collection in winter 2009, and Janet Silk for creating Fig. 1.This research was supported by the Ministry of Science and Higher Education, Portugal (Fundação para a Ciência e a Tecnologia), the British Antarctic Survey and the Tinker Foundation, under the research programs CEPH, SCAR AnT-ERA, PROPOLAR and ICED. This study represents a contribution to the Ecosystems component of the British Antarctic Survey Polar Science for Planet Earth Programme, funded by The Natural Environment Research Council.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Alvito, P.M., Rosa, R., Phillips, R.A. et al. Cephalopods in the diet of nonbreeding black-browed and grey-headed albatrosses from South Georgia. Polar Biol 38, 631–641 (2015). https://doi.org/10.1007/s00300-014-1626-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-014-1626-3