
Abstract
Analytical modeling of predator–prey systems has shown that specialist natural enemies can slow, stop and even reverse pest invasions, assuming that the prey population displays a strong Allee effect in its growth. We aimed to formalize the conditions in which spatial biological control can be achieved by generalists, through an analytical approach based on reaction–diffusion equations. Using comparison principles, we obtain sufficient conditions for control and for invasion, based on scalar bistable partial differential equations. The ability of generalist predators to control prey populations with logistic growth lies in the bistable dynamics of the coupled system, rather than in the bistability of prey-only dynamics as observed for specialist predators attacking prey populations displaying Allee effects. As a consequence, prey control is predicted to be possible when space is considered in additional situations other than those identified without considering space. The reverse situation is also possible. None of these considerations apply to spatial predator–prey systems with specialist natural enemies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
1.1 Modeling the biological control of invasive pests
Biological invasions are a major contemporary problem (Pimentel 2011; Garnier et al. 2012; Mistro et al. 2012; Potapov and Rajakaryne 2013; Wang et al. 2013; Savage and Renton 2013) for which few solutions are available, all of which are very costly. The use of natural enemies for the biological control of invading insects is one of the most promising possibilities (Moffat et al. 2013; Li et al. 2014; Ye et al. 2014; Basnet and Mukhopadhyay 2014). As invasion is essentially a spatial process, the potential of natural enemies to stop or even reverse an invasion is of particular interest. The fundamental analytical work of Owen and Lewis (2001) showed that specialist predators could potentially slow, stop or reverse the spread of invasive pests. The reversal of pest spread by specialist predation requires a strong Allee effect for the pest-only dynamics, defined as a negative growth rate for the prey population at low density. In the presence of a weak Allee effect, the predator can stop, but not reverse the wave of invasion. These conclusions have been confirmed in several other theoretical studies (Cai et al. 2014; Boukal et al. 2007; Morozov and Petrovskii 2009).
Generalist predators can also control prey effectively (Erbach et al. 2014; Chakraborty 2015). Their use could be promoted through conservation biological control programs without the need for exogenous specialist natural enemies. Unfortunately, the role of generalist predators in the spatial control of their prey has been much less studied than that of specialist predators, due to the intrinsic difficulties of having to work with a system of equations rather than with a single scalar equation. However, two important studies have been carried out in this area: the analytical and comprehensive study of Du and Shi (2007), and the preliminary simulation study of Fagan et al. (2002). Both used the same model structure as we do here, with logistic growth for both prey and predator populations, and a type II functional response for predators. The convergence of these models was strengthened further by the in-depth analysis of Magal et al. (2008) in which space was not considered. It is difficult to use these models in a spatial context: the work of Du and Shi (2007) cannot deal with invasion and traveling waves, because it deals with a bounded space. The numerical simulations of Fagan et al. (2002) are restricted to a few parameter values. They are, however, valuable, because they suggest conditions in which a generalist predator might be able to stop, and even reverse the invasion wave of a pest population displaying logistic growth. Fagan et al. also reported the results of field studies indicating that predators with diffusion coefficients higher than those of their prey are poor control organisms. The authors provided an explanation for this finding founded on logical arguments, but without a firm mathematical foundation. This result has been confirmed by a few numerical simulations including space, as reported by Magal et al. (2008), revealing a strong dependence of system dynamics on the relative rates of diffusion of the prey and the predator. It is thus important to take space into account by adding diffusion terms for both predator and prey. This conclusion accounts for the interest of scientists in questions of this type (Lewis et al. 2013; Hastings 2000; De Roos et al. 1991, 1998).
There are therefore hopes that it might be possible to extend the conditions for the control of invasive prey organisms to (i) generalist predators and (ii) prey populations displaying growth patterns not dependent on the restrictive assumption of Allee effects. Such control approaches would have a major impact in the field, given the high degree of generalism obtained. The aim of this study was, therefore to formalize the conditions in which spatial biological control can be achieved by generalists, through an analytical approach based on traveling waves solutions of reaction–diffusion equations.
Traveling wave solution describes a constant profile U moving through space at a speed c. Such waves are often observed in nonlinear reaction–diffusion systems modeling various phenomena. They are particularly suitable for describing the propagation of invasive fronts. In systems modeling a single species, described by a scalar equation, this type of solution is very well understood (Fischer 1937; Kolmogorov et al. 1937; Volpert et al. 1994 for a complete theory). Two particular classes of equations can be distinguished: monostable equations (like the Fisher-KPP equations) and bistable equations (often modeling the Allee effect). In monostable equations, there is a minimal wave speed \(c^*\) such that, for any \(c\ge c^*\), a wave solution with speed c exists. In bistable equations traveling waves exist for a unique speed \(c^*\). The sign of this speed \(c^*\) distinguishes between invasion or extinction of prey, which is a key property for our purposes.
For interactions of several species (described by a multidimensional system), the situation is much more complex. However, for some type of interaction, cooperation for instance, the system possesses a strong structural property, namely monotonicity. Essentially, this monotony makes it possible to use the comparison principle, which is always possible for one-dimensional systems, and the theory is then complete (see Volpert et al. 1994). Unfortunately, our system, and prey–predator systems in general, do not have such a monotonous structure. This method is then unsuitable for monotonous systems and only a few results have been published. One of the key reasons for this is as follows: when we search for traveling wave solutions for a system with N equations, we obtain a system of N second order ordinary equations that can be reformulated as a system of 2N first order ordinary equations. In the scalar case (\(N=1\)), it is therefore possible to study trajectories in a plane, available using classical tools for two-dimensional dynamical systems. For several species (\(N>1\)), it is necessary to study trajectories in a 2N-dimensional space, which may be very difficult.
Hence, the first rigorous results demonstrating the existence of traveling waves in prey–predator systems were based on a generalization, to the fourth dimension, of the classical shooting method in the phase plane (Dunbar 1984a, b). This approach has since been generalised (Huang et al. 2003; Xu and Weng 2012). However, all these studies simply investigate the mere existence of traveling waves. They do not determine the direction of the wave or the global dynamics for general initial conditions. Other methods have recently been developed in similar models (Huang and Weng 2013; Ducrot and Langlais 2012), but they are subject to the same limitations. A last approach is to use the degree theory (see e.g. Giovangigli 1990; Volpert et al. 1994) to obtain the existence of traveling waves. These homotopy methods may occasionally give some information on the speed c. Unfortunately, this needs additional estimates which are very difficult to obtain here. We therefore required another method.
The analysis provided in Magal et al. (2008) gives conditions for preys’ control by predators, but this analysis was carried out largely without reference to space. Thus, we have extended the system of Magal et al. (2008) by adding spatial diffusion. We find that the conditions for control are very different from those for the system in which space is not considered. The conditions for prey extinction and invasion are discussed in terms of two essential parameters: the encouter rate E and the handling time h. Increasing E clearly increases predator pressure. Conversely, increasing h decreases predator pressure.
The paper is organized as follows. In Sect. 2 we present the mathematical model and the main result of this work: Theorem 2.1 describes invasion conditions for the ODE system and the Theorems 2.4 and 2.6 the invasion conditions for the PDE system. The mathematical results are completed by numerical simulations in Sect. 3. The results are discussed in Sect. 4. The final Sect. 5 is devoted to the mathematical proofs.
2 Model and main results
2.1 Mathematical model
We analyze a system of partial differential equations for a prey population with logistic growth, and a generalist predator population with logistic growth on alternative prey in the absence of the invading host. The functional response is of Holling type II. The prey–predator interactions are modeled by the following partial differential equation system:
with:
-
u(t, x) \(=\) prey density at time t and at point x.
-
v(t, x) \(=\) predator density at time t and at point x.
-
\(D_u \) \(=\) diffusion rate of prey
-
\( D_v \) \(=\) diffusion rate of predators
-
\( r_1 \) \(=\) growth rate of prey
-
\( r_2 \) \(=\) growth rate of predators
-
\(K_1 \) \(=\) carrying capacity of prey
-
\( K_2 \) \(=\) carrying capacity of predators in absence of focal prey
-
E \(=\) encounter rate
-
h \(=\) handling time
-
\(\gamma \) \(=\) conversion efficiency
-
\(u_0\ge 0\) the initial concentration of prey, \(D_u, D_v, r_1, r_2, K_1, K_2, E, h \text { and } \gamma \) are positive constant parameters.
We carried out the following adimensionalization:
-
\(t^{\prime }=r_1 t\); \(x^{\prime }=x\sqrt{\frac{r_1}{D_u}}\); \(u^{\prime }\left( x^{\prime },t^{\prime }\right) =u(t,x)/K_1\); \(v^{\prime }\left( x^{\prime },t^{\prime }\right) =v(t,x)/K_2\)
-
\(d^{\prime }=D_v/D_u\); \(r^{\prime }=r_2/r_1\); \(E^{\prime }= E K_2/r_1\); \(h^{\prime }=r_1 h K_1/K_2\) ; \(\gamma ^{\prime }=\gamma K_1/K_2\) ; \(\alpha =\frac{\gamma ^{\prime }}{r^{\prime }}\).
Removing the sign, to simplify the notation, the system reads
with the initial conditionsFootnote 1
2.2 Main results
We distinguish two ways in which a predator can control the prey, one taking space into account and the other not considering this factor (mathematical definitions are provided in Definition 2.2.1).
-
The spatially uniform extinction results exclusively from local demographic processes and is independent of space.
-
The extinction wave is due to both demographic and diffusive processes and may take various forms, from a traveling front to a pulse.
Conversly, invasion is defined as prey survival and we distinguish two ways in which the prey can invade.
-
The spatially uniform invasion, which is independent of space.
-
The non-uniform invasion, described by various spatial dynamics, from Turing phenomena to invasion waves.
Definition 2.2.1
Let \((u_0(x),v_0(x))\) be an initial condition verifying (3) and \((u(t,x),v(t,x))\) be the corresponding solution of (2).
-
Extinction of prey occurs if
$$\begin{aligned} \forall x\in \mathbb {R},\quad \lim _{t\rightarrow +\infty } u(t,x)=0. \end{aligned}$$ -
Prey extinction is uniform if it is uniform with respect to \(x\in \mathbb {R}\), that is, if there exists a map \(\phi (t)\) verifying
$$\begin{aligned} \forall x\in \mathbb {R},\quad \forall t>t_0,\quad 0\le u(t,x)\le \phi (t)\quad \text { and}\quad \lim _{t\rightarrow +\infty } \phi (t)=0. \end{aligned}$$ -
Prey extinction is non uniform if there is extinction but no uniform extinction.
-
Invasion of prey occurs if there is no extinction, that is if
$$\begin{aligned} \exists x\in \mathbb {R},\quad \limsup _{t\rightarrow +\infty } u(t,x)>0 \end{aligned}$$
2.2.1 Analysis of the associated ODE system
If space is not taken into account, system (2) may be rewritten as follows.
System (4) is well understood (Magal et al. 2008). Indeed, it is clear that there are always three trivial stationary states: (0, 0) and (1, 0), which are unstable and (0, 1), which is asymptotically stable if, and only if, \(E>1\). Moreover, there are no more than three non-trivial positive steady states. We are interested principally in the case \(E>1\). In this case, there are either no or two stationary positive steady states. If the two steady states exist, denoted \((\widehat{u},\widehat{v})\) and \((u^*,v^*)\) with \(\widehat{u}<u^*\) and \(\widehat{v}<v^*\), then \((\widehat{u},\widehat{v})\) is always unstable and \((u^*,v^*)\) is most often stable. In this case, there are two stable nonnegative solutions, (0, 1) and \((u^*,v^*)\) and the system is bistable.
We are interested principally in the conditions for prey extinction. If \(E<1\), then (0, 1) is unstable and no extinction occurs. We are therefore interested only in the case \(E>1\). Now, if \(E>1\), there are two possibilities. In the non bistable case, (0, 1) is globally stable and there is extinction. In the bistable case, provided that u is initially small enough, say \(u_0<\mu _1\) for some \(0<\mu _1<1\), then \(u(t)\rightarrow 0\) as \(t\rightarrow +\infty \). Conversely, if u is initially large enough, say \(0<\mu _2<u_0\), then \( u(t) \rightarrow u^*\) as \(t\rightarrow +\infty \). Thus, in this case, the outcome—extinction or invasion—depends on the initial conditions. The following result provides an explicit statement of the above in the parameter space (E; h) and is proven in Sect. 5.1.
Theorem 2.1
Let \(E>1\) and \(\alpha \ge 0\) be fixed.
-
(i)
Existence of positive solutions. There exists a unique \(h^*=h^*(E,\alpha )\) such that
-
If \(h< h^*\), then there is no positive stationary solution and there is extinction of prey for the ODE system.
-
If \(h>h^*(E,\alpha )\), then there exist two positive solutions for the ODE system \((\widehat{u},\widehat{v})\) and \((u^*,v^*)\) with \(\widehat{u}<u^*\).
-
-
(ii)
Stability of the solutions. Let \(h>h^*(E,\alpha )\). The solution \((\widehat{u},\widehat{v})\) is always unstable. Moreover, there exists a unique \(h^{**}(E,\alpha )>h^*(E,\alpha )\) such that
-
If \(h>h^{**}(E,\alpha )\) then \((u^*,v^*)\) is stable.
-
If \(h^*(E,\alpha )<h<h^{**}(E,\alpha )\), the stability of \((u^*,v^*)\) depends on r. It is unstable if r is small enough and stable otherwise.
-

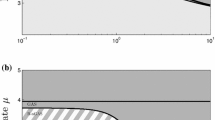
Description of the dynamics of the ODE system (4) in the \(E-h\) plane. If \(E<1\), the control solution (0, 1) is unstable. In this zone there exists at least one positive steady state and the prey never disappears entirely. If \(E>1\), then the control solution (0, 1) is always (locally) stable. Moreover, the \(E>1\) zone is the union of three subzones. Below the \(h^*\) curve, (0, 1) is the only non-negative steady state and is a global attractor: this is a monostable zone. Above the \(h^*\) curve, there are two additional positive steady states, one of which is always unstable while the second, denoted \((u^*,v^*)\), may be stable or unstable. Above the \(h^{**}\) curve, \((u^*,v^*)\) is always stable. In this subzone, the asymptotic behavior depends only on the initial conditions: this is a bistable zone. Between the \(h^*\) and the \(h^{**}\) curves, the stability of \((u^*,v^*)\) depends on other parameters: this is a conditional bistable zone. For illustrative purpose, the size of this last subzone has been considerably increased
Remark 1
Our calculations show that the gap between \(h^*\) and \(h^{**}\) is very small, so that, roughly speaking, \((u^*,v^*)\) is stable whenever it exists. However, if h belongs to the conditional stability zone, i.e. \(h\in (h^*,h^{**})\), and if r is very small, stability is lost and the system becomes excitable. This explains, in particular, the presence of pulses for small values of r when dealing with spatial interactions (see Sect. 3).
The map \((E,\alpha )\mapsto h^*(E,\alpha )\) has the following properties, as proved in Sect. 5.1.
Properties 2.2
Let \(\alpha \ge 0\) be fixed. The map \(E\mapsto h^*(E,\alpha )\) is increasing and one has the explicit limits:
Let \(E\ge 1\) be fixed. The map \(\alpha \mapsto h^*(E,\alpha )\) is increasing and one has the explicit limits:
Figure 1 illustrates the maps \(h^*\) and \(h^{**}\) and the possible outcomes for system (4).
2.2.2 Analysis of the PDE system
We wish to identify the parameter conditions required to obtain prey extinction in the PDE system (2). A simple stability analysis shows that, if \(E<1\), then invasion occurs in the PDE system. If \(E>1\), then the situation for the PDE system is more complex. In this situation, the spatial structure and diffusion processes result in additional conditions for extinction. The rationale is explained in detail below.
Let us assume that there is a positive stable stationary solution of (2) denoted by \((u^*,v^*)\) and that the initial condition u(x, 0) is close to \(u^*\) at some places x and close to 0 at other places. Since both \(u^*\) and 0 are stable, the demographic phenomena lead to an agregation near \(u^*\) and an agregation near 0. However, diffusion allows individuals to move around in space, so one of 0 or \(u^*\) may be the final global attractor. In other words, there may be a (stable) traveling wave joining \(u^*\) to 0. The direction of this wave, given by the sign of the speed of the wave, indicates whether extinction or invasion occurs. However, there are difficulties associated with this argument.
-
There can be no homogeneous stationary solution of (2), only stable heterogeneous positive stationary solutions. In other words, it is possible that \(h<h^*(E,\alpha )\) without control occurring.
-
Even in the case of bistability (\(h>h^*(E,\alpha )\)), the bistable system (2) is neither competitive nor cooperative. Little theoretical knowledge is available concerning the occurrence of traveling waves in such systems, with even less known about the stability and direction of the wave.
Using super and subsolutions, we show here how to obtain the conditions sufficient (but not necessary) for extinction and for invasion, based on well known scalar bistable PDEs. Roughly speaking, let (u(t, x), v(t, x)) be the solution of (2). If we find a positive constant \(\underline{v}\) such that, for anyFootnote 2 \((t,x)\in \mathbb {R}^+\times \mathbb {R}\), \(v(t,x)\ge \underline{v}\) then it comes
Let \(\overline{u}\) be the solution of
The comparison principle implies that \(\overline{u}(t,x)\ge u(t,x)\). Now, if \(\overline{u}(x,t) \rightarrow 0\) when \(t \rightarrow +\infty \) then \(u(t,x)\rightarrow 0\) when \(t \rightarrow +\infty \) and extinction of prey occurs (see Fig. 2a). Moreover, if \(\overline{u}(x,t)=\phi (t)\) does not depend on x, then there is aspatial control (see Fig. 2b).
Conversely, if we can identify a positive constant \(\overline{v}\) such that \(v(x,t)\le \overline{v}\), then it comes
Now, define \(\underline{u}(t,x)\) as the solution of
The comparison principle implies that \(\underline{u}(t,x) \le u(t,x)\). It follows that if for some \(x\in \mathbb {R}\), \({ \limsup _{t\rightarrow +\infty } \underline{u}(x,t) >0}\), then \({\limsup _{t\rightarrow +\infty } u(t,x)>0}\) and there is (non uniform) invasion (see Fig. 3).
These arguments give rise to the following theorems yielding sufficient conditions, in terms of the parameters E, \(\alpha \) and h, for extinction or invasion to occur. All theorems are proven in Sect. 5. We begin with a sufficient condition for uniform extinction.

a Sufficient condition for extinction. The solution u(t, x) is majored by a supersolution \(\overline{u}(t,x)\). If \({\lim _{t\rightarrow +\infty }\overline{u}(t,x)=0}\), then \({\lim _{t\rightarrow +\infty }u(t,x)=0}\) and there is extinction. b Sufficient condition for uniform extinction. The solution u(t, x) is majored by a supersolution \(\phi (t)\) which does not depend on x. If \({ \lim _{t\rightarrow +\infty } \phi (t)=0}\), then \({\lim _{t\rightarrow +\infty } u(t,x)=0}\) uniformly in x and there is uniform extinction

Sufficient condition for invasion. The solution u(t, x) is minored by a subsolution \(\underline{u}(t,x)\). If \({\limsup _{t\rightarrow +\infty } \underline{u}(t,x)>0}\), then \({\limsup _{t\rightarrow +\infty } u(t,x)>0}\) and there is invasion
Theorem 2.3
Let \(E>1\) and define
If \(h<h_1(E)\) then there is uniform extinction. In other words, for any initial condition verifying (3), there exists \(\phi (t)\ge 0\) such that any solution (u(t, x), v(t, x)) of (2) verifies
If \(h>h_1(E)\), then there can be invasion or extinction. The following theorem gives a sufficient condition for extinction to occur.
Theorem 2.4
Let \(E>1\) be fixed and let (u, v) be the solution of (2) and (3). Define \(\underline{v}=1\) and let \(\overline{u}\) be a solution of (6) together with \(\overline{u}(0,x)=u(0,x)\). There exists a unique \(h^-=h^-(E)>h_1(E)\), such that
-
If \(h<h^-(E)\), then \(\forall x\in \mathbb {R},\) \(\lim \nolimits _{t\rightarrow +\infty } \overline{u}(t,x)=0\)
-
If \(h>h^-(E)\), then \(\forall x\in \mathbb {R},\) \(\lim \nolimits _{t\rightarrow +\infty } \overline{u}(t,x)=\overline{\mu }\) where \(\overline{\mu }=\overline{\mu }(E,h)\) is a positive scalar.
As a consequence, if \(h<h^-(E)\) there is extinction of prey.
The map \(E\mapsto h^-(E)\) verifies the following properties proved in Sect. 5.3.
Properties 2.5
The map \(E\mapsto h^-(E)\) is increasing and admits the following explicit limits:
Our last result gives a sufficient condition for invasion to occur.
Theorem 2.6
Let \(E>1\) and \(\alpha \ge 0\) be fixed and let (u, v) be the solution of (2) and (3). Define \(\overline{v}=1+\alpha \frac{E}{1+Eh}\) and let \(\underline{u}\) be a solution of (6) together with \(\underline{u}(0,x)=u(0,x)\). There exists a unique \(h^+=h^+(E,\alpha )>h^*(E,\alpha )\) such that
-
If \(h<h^+(E,\alpha )\), then \(\forall x\in \mathbb {R},\) \(\lim \nolimits _{t\rightarrow +\infty } \underline{u}(t,x)=0\)
-
If \(h>h^+(E,\alpha )\), then \(\forall x\in \mathbb {R},\) \(\lim \nolimits _{t\rightarrow +\infty } \underline{u}(t,x)=\underline{\mu }\) where \(\underline{\mu }=\underline{\mu }(E,h)\) is a positive scalar.
As a consequence, if \(h>h^+(E,\alpha )\) there is invasion of prey.
Finally, the following result specifies the behavior of the map \(h^+\).
Properties 2.7
The maps \(E\mapsto h^+(E,\alpha )\) and \(\alpha \mapsto h^+(E,\alpha )\) are increasing. For any \(E>1\), \(h^+(E,0)= h^-(E)\) and \(\lim _{\alpha \rightarrow +\infty } h^+(E,\alpha )=+\infty \). Finally, \(\alpha \ge 0\) being fixed, one has the explicit limit
The results above are summarized in Fig. 4. In the domain \(\{(E,h),E>1, h^-(E)<h<h^+(E,\alpha )\}\), which we will refer to as the ‘transition zone’, it is not possible to draw any conclusions concerning whether prey invasion or extinction is likely to occur. Indeed, this zone can be separated into two subzones, according to the parameters values:
In Zone I, our numerical simulations show non-monotonous traveling waves. In zone II, simulations show various types of behavior, including pulse and even heterogeneous positive stationary solutions. This phenomena are discussed in the Sect. 3.

Description of the dynamic of the PDE system (2) in the \(E - H\) plan. If \(E<1\) there is always invasion. If \(E>1\) there is a uniform extinction for \(h<h_1(E)\), extinction for \(h_1(E)<h<h^-(E)\) and invasion for \(h^+(E,\alpha )<h\). The zone between \(h^-\) and \(h^+\) is called the transition zone. This transition zone is splitted into two subzones: Zone I and Zone II, separated by the \(h^*\) curve. In these two zones, both extinction or invasion of prey may occur due to various spatial phenomena
3 Numerical study of the transition zone
3.1 Influence of \(\alpha \)
The mathematical results above demonstrate the influence of the parameters E and h, and, indirectly, that of the conversion rate \(\alpha \), on the long-term behavior of the system. More precisely, when \(E>1\), prey extinction or invasion may occur, depending on the value of h. Indeed, we can define two values \(h^-=h^-(E)<h^+=h^+(E,\alpha )\) (see Theorems 2.4, 2.6). Extinction occurs if \(h<h^{-}\) and invasion occurs if \(h>h^+\). When \(h\in (h^-,h^+)\) we observe richer dynamics, which may depend on other factors. We refer to this zone as the transition zone. Note that, as \(h^-\) is not dependent on \(\alpha \) and \(h^+\) is an increasing function of \(\alpha \) (Proposition 2.7), the size of this transition zone increases with increasing \(\alpha \).

Computation of \(h^*(E,\alpha )\), \(h^-(E)\) and \(h^+(E,\alpha )\) for four values of \(\alpha \). The so-called transition zone lies between the red curve \(h^+\) and the blue curve \(h^-\). The transition zone increases with increasing \(\alpha \). This transition zone can be split into two subzones separated by \(h^*\) (black line) (color figure online)
A first clue to the possible dynamics in the transition zone is provided by an understanding of the dynamics of the ODE system (4) described in the Theorem 2.1. The dynamic of (4) is essentially dependentFootnote 3 on the position of h relative to \(h^*=h^*(E,\alpha )\). When \(h<h^*\), there is no positive stationary solution, whereas for \(h>h^*\) there are two positive stationary solutions, one of which, the larger of the two, is (nearly always) stable.
The position of \(h^*\) relative to \(h^-\) and \(h^+\) provides a first description of the transition zone. By virtue of Proposition 2.2, one gets the following. We always have \(h^*<h^+\) but the position of \(h^*\) relative to \(h^-\) is dependent on \(\alpha \). On the one hand, from the facts that \(h^*(1,\alpha )>h^-(1)\) for \(\alpha >0\) and \(h^*(E,0)= h_1(E)<h^{-}(E)\) for \(E>1\), we deduce that \(h^->h^*\) for large enough values of E and small enough values of \(\alpha \). On the other hand, \(h^*\) is an increasing function of \(\alpha \) tending to \(+\infty \). We obtain that \(h^*(E,\alpha )>h^-(E)\) for large values of \(\alpha \) and any \(E>1\).
Remark 2
When \(h^->h^*\), which may occur for sufficiently small values of \(\alpha \), we see that taking space into account automatically increases the potential of extinction of preys.
The transition zone can thus be separated into two subzones: one in which \(h<h^*\) (Zone II) and one in which \(h>h^*\) (Zone I). Figure 5 sums up this discussion. As we will see below, both extinction and invasion are possible in each of these zones, but the phenomena at work differ considerably, according to whether \(h<h^*\) or \(h>h^*\). These phenomena are studied in more detail below, using a numerical approach.
3.2 Extinction or invasion: influence of r and d
Our numerical analysis shows that both invasion and extinction are possible in the transition zone. When space is taken into account we see that \(h^*\) does not separate the zone of invasion from that of extinction. These two zones are, indeed, separated by a new critical value of the handling time denoted \(h_{crit}\in (h^-,h^+)\), which is dependent on E and \(\alpha \), of course, but also on the relative rates of growth (r) and diffusion (d) of the predator population:
As expected, when \(h>h_{crit}\), prey invasion is observed, whereas extinction is observed when \(h<h_{crit}\). Thus, higher values of \(h_{crit}\) are associated with more effective predation and thus with less effective invasion by the prey.
Figure 6 completes the theoretical scheme represented in Fig. 4, by presenting an example of the curve \((E,h_{crit})\) for a particular selection of values for the parameters \(\alpha \), r and d in the \(E-h\) plan.
Like \(h^*\) and \(h^+\), \(h_{crit}\) increases with both E and \(\alpha \). This naturally translates into the fact that, higher values of E increases the chance of meeting between predators and prey and that at higher \(\alpha \) values, the predator is able to make greater use of the prey and can therefore eliminate it.

Numerical computation of \(h_{crit}\) in the \(E-h\) space for the fixed values of \(\alpha =4\), \(d=1\) and \(r=1\). The transition zone \(h\in (h^-,h^+)\) is split into two subzones separated by \(h_{crit}\). Extinction occurs below \(h_{crit}\) and invasion occurs above

Numerical computation of \( h_{crit}\) with respect to d for four values of r with \(E=2\) and \(\alpha =4\). For a given value of the parameters, there is invasion of prey if \(h>h_{crit}\) and extinction if \(h<h_{crit}\). Therefore, the larger \(h_{crit}\) is, the greater is the potential for extinction of prey. We see that \(h_{crit}\) is increasing in r and (essentially) decreasing in d. Thus, an increase of d or a decrease of r decrease the impact of the predation on invasive prey
Given the multiple dependence of \(h_{crit}\) on different parameters, Fig. 6 can only represent a particular case, chosen for its simplicity. Figure 7 shows the relationships between \(h_{crit}\) and d for fixed values of E and \(\alpha \) and for various values of r. We see that \(h_{crit}\) increases with r. This translates into the fact that for small values of r, the predators growth rate is small and their effectiveness reduced. Conversly, \(h_{crit}\) (essentially) decreases with increasing d. This is due to the fact that for large d, predators spread into a zone in which prey are not present, resulting in a weakening predation.
Remark 3
For small values of r and intermediate values of d, predators may increase their effectiveness by increasing d. In that case, the predator growth rate being small, predators density remains high for a long time even if prey are absent. Now, if d is large enough but not large, predators may spread into a zone in which prey are not present and remain there at a high density and long enough to stop the prey to invade. This phenomenon may enable predators to form a barrier to prey’s movement, preventing thereby prey propagation. For too large values of d, the loss in predators effectiveness due to movement is too strong and the above phenomenon does not hold any more. This explains why, for small values of r, \(h_{crit}\) first increases and then decreases with increasing d.
3.3 Dynamics of the system in the transition zone
We will now describe the different dynamics occurring in the transition zone, summarized in Table 1. See Sect. 2.2 for a precise definition of the various quantities described here.
-
A] Invasion (\(h>h_{crit}\))
-
Turing instabilities: \(h<h^*\) and \(d\gg 1\) As d increases, predators spread out, moving into areas from which the prey is absent, leading to a decrease in the size of the predator population. This phenomenon leads to a decrease in predator density throughout the space occupied by the predator, allowing the prey to survive in certain zones. We thus obtain a periodic distribution in space and a constant distribution over time of the densities of the prey and predator. Mathematically, this phenomenon is described by a Turing bifurcation. Away from this bifurcation, other interesting regimes occur. For instance, we observe non-periodic stationary pattern.
-
Invasion traveling waves (ITW): \(h>h^*\) The invasion is described simply by an invasion traveling wave: a wave of propagation linking the two stable solutions \((u^*,v^*)\) and (0, 1) in the direction of the positive solution \((u^*,v^*)\). When r is large this traveling wave is monotone. By contrast, when \(r\ll 1\) it displays a rich dynamics. When \(r\ll 1\) and d is not too large, we observe that \(h_{crit}\) is approximately equal to \(h^*\) (see Fig. 7). This indicates that there is an invasion traveling wave if a positive solution exists. In this case, ahead of the front, \(v=1\) and, since \(r\ll 1\), the predator population increases very slowly. Besides, as \(h>h^-\), the prey invades the space when the predator is at concentration 1. This leads to front advancing. Behind the front, the predator has had sufficient time to increase the size of its population and, therefore, to decrease the size of the prey population. As \(h>h^*\), the population of the prey decreases towards the positive solution \(u^*\) and we observe a non-monotonous invasive traveling wave.
-
-
B] Extinction (\(h<h_{crit}\))
-
Pulse: \(h\in [h^-,h^*]\) and \(r\ll 1\) Since \(r\ll 1\), the front of the wave is similar to the ITW described above. However, as \(h<h^*\), here is no homogeneous positive solution \(u^*\). The prey population therefore decreases to zero behind the front, whereas it continues to advance in ahead of the front. We thus obtain a pulse.
-
Extinction traveling wave (ETW): \(h>h^*\) This corresponds to the simplest case described above. We observe a propagation wave linking the two stable solutions \((u^*,v^*)\) and (0, 1) in the direction of the control solution (0, 1).
-
Finally, Fig. 8 presents the map in the \(E-h\) plane for fixed values of \(\alpha \), d and r. It furthermore specifies the possible dynamics in each zone.

Different spatial dynamics in the transition zone. To ensure that all possible situations are represented, we choose \(\alpha =0.5\), \(r=0.01\) and \(d=10\). Note that traveling waves of extinction (ETW) and traveling waves of invasion (ITW) may be obtained outside the transition zone
4 Conclusion and discussion
4.1 Summary of the results
We have shown that invasion occurs if \(E<1\). If predators do not encounter their prey they cannot control them. If \(E>1\), then extinction or invasion can occur, depending on the parameters h, E, \(\alpha \) and r. Uniform extinction occurs for \(h<h_1(E)\) and extinction occurs for \(h_1(E)<h<h^-(E)\). Invasion occurs for \(h^+(E,\alpha )<h\). Thus, if h increases, we move from a zone of extinction without a consideration of space to a zone of extinction requiring a consideration of spatial aspects and then to a zone of invasion. For intermediate values of E, the zones of extinction increase with increasing E resulting in a higher potential of extinction, as \(h_1,h^*, h^-\) are increasing functions of E. When E is large, the zones of control do not depend on E any more because \(h_1,h^-, h^+\) have finite limits when \(E \rightarrow +\infty \). Thus, E can only play a role in prey control if it takes intermediate values and if h is not too large. In summary, for low values of E (\(E<1\)) or high values of E or h, the outcome of the interaction (extinction or invasion) is independent of E.
There is furthermore a transition zone splitted in two subzones, with various spatio-temporal phenomena and wherein both extinction and invasion can occur. The size of this transition zone greatly increases when the conversion rate \(\alpha \) increases. Depending on the relative positions of these two zones with regard to the zones of extinction and of invasion, four spatial dynamics were identified: extinction and invasion traveling waves, extinction pulse waves and heterogeneous stationary positive solutions of the Turing type.
4.2 Biological interpretation of the main results
We have shown that an increase in E increases the potential of extinction while an increase in h increases the potential of invasion. This translates the fact that a highly effective predator does have a high encounter rate and a small handling time. Furthermore, since \(h^+\) is an increasing function of \(\alpha \), an increase in \(\alpha \) decreases the potential of invasion and increases the size of the transition zone, which in turn increases the potential for the system to have complex dynamics. Finally, an increase of the diffusion rate d and a decrease of the amplitude of the predators growth rate r both increase the potential of invasion of prey. Thus, a generalist predator loses its effectiveness to exterminate invasive prey if it diffuses too fast or if it has a too slow dynamics.
The above results are stated in term of adimensionalized parameters (see Sect. 2.1). By choosing the appropriate spatio-temporal variables, we may define \(D_u=r_1=1\). Thus, the biological interpretations of d and r are accurate. Conversely, the definitions of the searching efficiency, \(E=\textit{EK}_2\), the handling time \(h= h \frac{K_1}{K_2}\) and the conversion rate \(\alpha =\frac{\gamma }{r}\) complicate the biological interpretation of these three parameters. Thus, in addition to the above discussion about the influence of E and h, we now discuss our results in terms of the other biological variables: \(K_1\), \(K_2\), \(\gamma \) and r. The parameter h being increasing in the carrying capacity \(K_1\) of prey, prey with high carrying capacity show a high risk of being invasive. Conversly, h and E are respectively decreasing and increasing in the carrying capacity of predators \(K_2\). Thus, predators with high carrying capacity have a high potential to control prey invasion. Otherwise \(\gamma <1\) means that predator growth is mostly due to alternative prey while \(\gamma >1\) implies that predator growth is due to consumption of the focal invasive prey. Finally, an increase in \(\gamma \) and a decrease in the amplitude of the predator growth rate r yield an increase of \(\alpha \). Therefore, predators with a preference for the invasive prey or predators with a slow dynamics might display a complex dynamics. In particular, the likelihood of the system to exhibit a pulse wave is then important.
4.3 The consideration of space often, but not always, increases the potential for control of pest invasion
The model analyzed here was studied without taking space into account, except for a numerical exploration in the discussion, in an article by Magal et al. (2008). As explained in the introduction, models of identical structure have been proposed independently by Fagan et al. (2002) and Chakraborty (2015). We will now discuss our results in the context of these previous studies. Adding a spatial component to predator–prey systems makes any prediction about the controllability of the system difficult, as it then depends on the values of several parameters. The comparison between situations with and without the consideration of space is epitomized by the distinction between \(h^*\), separating parameter regions of mono- and bi-stability in the ODE system, and \(h_{crit}\), separating parameter regions of invasion and extinction in the PDE system. We will now focus on the case of \(E>1\), as values of \(E<1\) do not promote control, predators encountering prey too infrequently.
If space is not taken into account, control occurs if \(h<h^*\), as 0 is a global attractor. This is still true in situations in which space is taken into account, if \(h<h_1\) where \(h_1\) is smaller than \(h^*\). When h is between \( h_1\) and \(h^*\), 0 is only a local attractor, so it is not possible to state that control is always attained. In this respect, adding consideration of space decreases the potential for control. Furthermore, when space is not taken into account, there is either extinction or invasion when \(h>h^*(E,\alpha )\), depending on initial conditions. Incorporating consideration of space changes the region where invasion occurs, for any values of the other parameters and for appropriate initial conditions, into \(h>h^+(E,\alpha )\), with \(h^+>h^*\). Thus, the consideration of space reduces the size of the zone wherein the invasion is certain and is detrimental to the invading prey. Finally, the relative levels of predator and prey diffusion also determine the potential for control. Our model shows that control is increased by predators being less mobile than prey. If predator mobility levels are too high, the predators become to thinly spread on the ground. For similar reasons, too high a level of prey mobility leaves the prey vulnerable to predators. This is entirely consistent with the experimental findings of Fagan et al. (2002). In conclusion, taking space into account can lead to an increase or a decrease in the controllability of invading prey by predators; the addition of space to the model has no generic implication for considerations of predator–prey dynamics (see also Lam and Ni 2012; Braverman et al. 2015).
4.4 How can generalist predators reverse invasion by pest?
The originality of this study lies in its consideration of a generalist predator in a spatial context. When studying generalist predators, it is common practice to assume that the functional response is of type III, due to switching between prey species (Erbach et al. 2014; van Leuven et al. 2007, 2013; Morozov and Petrovskii 2013). However, this approach is not mandatory, and other works (Basnet and Mukhopadhyay 2014; Krivan and Eisner 2006; Hoyle and Bowers 2007) have considered a type II functional response. Altering our model to include a type III functional response would be very costly in terms of understanding, because such responses lead to a loss of bistability. Its derivative would be null without prey, so some of our demonstration would fail and the analytical complexity would be greatly increased. However, traveling waves for specialist predator with type III functional response are known to exist (Li and Wu 2008) which indicates that our result may be extended to this case.
The complexity of analytical studies of spatial predator–prey interactions lies in the reaction terms being of alternative signs in the equations, making the study of the systems of equations essential (Dunbar 1984b; Huang et al. 2003; Huang and Weng 2013). Other interactions, such as competition of two species (all negative) and symbiosis (all positive), are simpler, as their studies are similar to the study of a single equation (Volpert et al. 1994; Alzahrani et al. 2012). This accounts for the slow scientific progress in this otherwise highly relevant topic. However, several major results have been obtained in recent decades, including those of the fundamental work of Owen and Lewis (2001). The finding of Owen and Lewis that predators can slow, stop, and even reverse invasion by their prey was based on the bistability of the prey-only dynamics of systems consisting of specialist predators attacking prey populations displaying Allee effects. By contrast, our work shows that the ability of generalist predators to control prey populations with logistic growth lies in the bistable dynamics of the coupled system. We also observe pseudo-Allee effects in our system, but their physics is quite different. An analysis of the ODE system identified parameter regions of monostable (extinction) and bistable (extinction or invasion) dynamics, but analysis of the associated PDE was able to distinguish different and additional regions of invasion and extinction. As a consequence, prey control was predicted to be possible when space was considered in additional situations other than those identified without considering space. The reverse situation was also possible. None of these considerations apply to spatial predator–prey systems with specialist natural enemies.
5 Proofs
5.1 Proof of Theorem (2.1)
Let \(E>1\) and \(\alpha \ge 0\) be fixed. For any \(h\ge 0\), the system (4) can be rewriten as
where \(\Theta _h(u)=\frac{E u}{1+Ehu}\), \(f_h(u)=\frac{1}{E}(1-u)(1+Ehu)\) and \(g_h(u)=1+\alpha \frac{Eu}{1+Ehu}\).
(i) Proof of the existence Define \(H(h,u)=f_h(u)-g_h(u)\). For a given \(h\ge 0\), a couple (u, v) is a positive stationary solution of (7) if and only if \(v=f_h(u)\) and
Now, fix \(u\in ]0,1[\). Since \(\partial _h H(h,u)=u(1-u)+\alpha \left( \frac{Eu}{1+Ehu}\right) ^2>0\), one sees that the map \(h\mapsto H(h,u)\) is increasing. From \(E>1\), we get \(H(0,u)<0\) and from \(u\in ]0,1[\) we get \(\lim \nolimits _{h\rightarrow +\infty } H(h,u)=+\infty .\) The map \(h\rightarrow H(h,u)\) being continuous, this implies that for any \(u\in ]0,1[\), there exists a unique \(h(u)>0\) such that \(\small {{\left\{ \begin{array}{ll} H(h,u)<0\quad \text {if}\,h<h(u),\\ H(h,u)=0\quad \text {if}\,h=h(u),\\ H(h,u)>0\quad \text {if}\,h>h(u).\\ \end{array}\right. }}\)

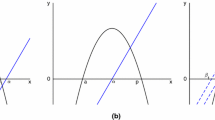
The four figures are computed for \(E=\alpha =2\), which gives \(h^*\approx 4.36\). The figure on the left represents the curve \(u\mapsto h(u)\) (in bold). Above the curve \(f_h(u)>g_h(u)\) while below the curve \(f_h(u)<g_h(u)\). For a given h, the ordinate u of a point of this curve verifies \(f_h(u)=g_h(u)\) and corresponds to the positive stationary solution \((u,f_h(u))\) of (7). The figures on the right represent the isoclines \(v=f_h(u)\) and \(v=g_h(u)\) for the three fixed values \(h=3\), \(h=4.36\) and \(h=5\). A positive stationary solution of the system (7) corresponds to an intersection of these two isoclines. The system (7) admits two positive solutions for \(h>h^*\), one (double) solution for the critical case \(h=h^*\) and zero positive solution for \(h<h^*\)
The smooth function \(u\mapsto h(u)\) may be computed explicitly by noting that for any \(h\ge 0\), the Eq. (8) is equivalent to the algebraic equation
wherein we have set
This yields the explicit formula
In particular
This implies that the minimum of h(u) is obtained for some \(u_{crit}\in ]0,1[\). We define
The definition of \(h^*\) shows that if \(h<h^*\), then (9) has zero solution. This also implies, together with the limits (11) and the continuity of \(u\mapsto h(u)\), that for any fixed \(h>h^*\) the Eq. (9) admits at least two solutionsFootnote 4 (see the Fig. 9). In addition to this, for any fixed \(h>0\), one has \(\deg (P_h)=3\) and \(P_h\) always admits a negative roots for \(P(0)=1-E<0\) and \(\lim \nolimits _{x\rightarrow -\infty }P_h(x)=+\infty \). This implies that (9) admits at most two solutions.Footnote 5 In conclusion, (9) has exactly two positive solutions if \(h>h^*\) and zero positive solution if \(h<h^*\). This ends the proof of (i).
(ii) Proof of the stability Let \(h>h^*\) be fixed. Let (u, v) be a positive stationary solution of (7). Since (u, v) verifies \(f_h(u)=g_h(u)=v\), the Jacobian matrix at (u, v) reads
hence
From the proof of (i), we know that the system (7) admits exactly two positive solutions denoted respectively as \((\widehat{u},\widehat{v})\) and \((u^*,v^*)\) with \(\widehat{u}<u_{crit}<u^*\) and such that
where \(u\mapsto h(u)\) is given by (10). In particular, \(\widehat{u}\) and \(u^*\) are the solution of
Differentiating the Eq. (14) with respect to u gives
Thus, using the known fact that \(\partial _h H(h(u),u)>0\), the identity (13) and the footnote 5, one gets \(\partial _u H(h,\widehat{u})<0\) and \(\partial _u H(h,u^*)>0\). Since \(\partial _u H(h,u)=-(g^{\prime }_h(u)-f_h^{\prime }(u))\), this shows that \(\det (J(\widehat{u},\widehat{v}))<0\) and the instability of \((\widehat{u},\widehat{v})\) follows.
By contrast, one has \(\det \big (J(u^*,v^*)\big )>0\) and it appears that the stability of \((u^*,v^*)\) is given by the sign of
In order to highlight the dependence on h, for any \(h>h^*\), we note \(u^*=u^*(h)\) and we also define \(\mu (h)=\frac{Eh-1}{2Eh}\). From \(f^{\prime }_h(u)=\frac{1}{2E^2 h}(\mu (h)-u)\) and (15), we infer the following:
-
If \(\mu (h)\le u^*(h)\) then \((u^*,v^*)\) is asymptotically stable.
-
If \(\mu (h)> u^*(h)\) then the stability of \((u^*,v^*)\) depends on r. More precisely, define \(r_{crit}=\frac{\Theta _h(u^*)f_h^{\prime }(u^*)}{v^*}>0.\) (It is easy to show that \(r_{crit}\le 1\)).
-
If \(r>r_{crit}\), then \((u^*,v^*)\) is asymptotically stable.
-
If \(r<r_{crit}\), then \((u^*,v^*)\) is unstable.
-
The sign of \(\mu (h)-u^*(h)\) with respect to the parameter h remains to be found.
On a first hand, the explicit expression of \(f_h^{\prime }\) shows that \(f^{\prime }_{h^*}\) is decreasing and that \(f^{\prime }_{h^*}(\mu (h^*))=0\). Moreover, the definition (12) of \(h^*\) yields \(u^*(h^*)=u_{crit}\) and \(f_{h^*}^{\prime }(u_{crit})=g_{h^*}^{\prime }(u_{crit})>0\). Hence, \(\mu (h^*)<u^*(h)\). It is also clear that \(\frac{1}{2}=\lim \nolimits _{h\rightarrow +\infty } \mu (h)<\lim \nolimits _{h\rightarrow +\infty } u^*(h)=1\). By continuity, we infer that the equation \(\mu (h)=u^*(h)\) has at least one solution in \((h^*,+\infty )\).
On another hand, if \(\mu (h)=u^*(h)\) then \(H(h,\mu (h))=0\), which may be rewritten as
A simple analysis shows that this equation has exactly one negative solution, one solution in \(\left( 0,\frac{1}{E}\right) \) and one solution in \(\left( \frac{1}{E},+\infty \right) \). Since \(h^*>\frac{1}{E}\) (see Footnote 4), this implies that \(\mu (h)=u^*(h)\) has exactly one solution in \((h^*,+\infty )\). We note this unique solution \(h^{**}=h^{**}(E,\alpha )\).
Finally, it is clear from the above arguments that \(\mu (h)<u^*(h)\) if \(h\in (h^*,h^{**})\) and that \(\mu (h)>u^*(h)\) if \(h>h^{**}\). This ends the proof of the theorem. \(\square \)
Proofs of Properties 2.2
Similarly to the proof of the theorem 2.1, and to highlight the role of the parameters E and \(\alpha \), let us define
From the proof of the theorem 2.1, we know that the quantity \(h^*=h^*(E,\alpha )\) and the corresponding \(u_{crit}=u_{crit}(E,\alpha )\) are characterized by the two equations
The implicit function theorem immediatly shows that the maps \((E,\alpha )\mapsto (h^*,u_{crit})\) belongs to \(C^1\big ((1,+\infty )\times [0,+\infty ); \mathbb {R}_+^2\big )\).
-
Proofs of the growth of \(E\mapsto h^*(E,\alpha )\) and of \(\alpha \mapsto h^*(E,\alpha )\)
Let \(\alpha \ge 0\) be fixed. Differentiate (16a) with respect to E and use (16b) yields
$$\begin{aligned} \partial _E H\left( E,\alpha ,h^*,u_{crit}\right) +\partial _E h^*(E,\alpha )\cdot \partial _h H\left( E,\alpha ,h^*,u_{crit}\right) =0. \end{aligned}$$We already know that \(\partial _h H<0\) and an explicit computation gives
$$\begin{aligned} \partial _E H\left( E,\alpha ,h^*,u_{crit}\right) =\frac{1}{E \left( 1+Eh^* u_{crit}\right) }>0. \end{aligned}$$It follows that \(\partial _E h^*(E,\alpha )>0\). Similar arguments show that \(\partial _\alpha h^*(E,\alpha )>0\).
-
Computation of \({\lim _{E\rightarrow 1}h^*(E,\alpha )}\)
Let \(\alpha \ge 0\) be fixed. Since \(h^*(\cdot ,\alpha )\) is increasing and positive on \((1,+\infty )\), there exists a nonnegative scalar \(h^*(\alpha )\) such that \(h^*(E,\alpha )\rightarrow h^*(\alpha )\) as \(E\rightarrow 1\). To compute this limit, denote \(P(E,\alpha ,h,u)=(1+Ehu) H(E,\alpha ,h,u)\). From (16) and the definition of \(h^*\), one see that \(h^*\) is the minimal value of h such that
$$\begin{aligned} \exists u\in [0,1],\quad P(E,\alpha ,h,u)=\partial _u P(E,\alpha ,h,u)=0. \end{aligned}$$(17)In other words, \(h^*\) is the minimal value of h such that \(P(E,\alpha ,h,\cdot )\) admits a multiple root in [0, 1].
By passing to the limit \(E\rightarrow 1\) in (17), we see that that \(h^*(\alpha )\) is the minimal value of h such that
$$\begin{aligned} \exists u\in [0,1],\quad P(1,h,\alpha ,u)=\partial _u P(1,h,\alpha ,u)=0. \end{aligned}$$(18)Explicit computations give
$$\begin{aligned} P(1,\alpha ,h,u)=u\left( h^2 u^2+h(2-h)u+\alpha +1-h\right) . \end{aligned}$$The multiplicity of the roots of \(P(1,\alpha ,h,\cdot )\) need now to be discussed.
\(\bullet \) If \(h<2\sqrt{\alpha }\) then 0 is the only root of \(P(1,\alpha ,h,\cdot )\) and the multiplicity is one.
\(\bullet \) If \(h\ge 2\sqrt{\alpha }\) then \(P(1,\alpha ,h,\cdot )\) has two other real roots and may be explicitly written as
$$\begin{aligned} P(1,\alpha ,h,u)=h^2 u(u-u_-)(u-u_+) \end{aligned}$$where
$$\begin{aligned} u_{\pm }=\frac{1}{2h}\left( h-2\pm \sqrt{(2-h)^2-4(\alpha +1-h)}\right) . \end{aligned}$$-
If \(h=2\sqrt{\alpha }\) then \(u_{-}=u_{+}=1-\frac{1}{\sqrt{\alpha }}\) and this multiple root belongs to [0, 1) if and only if \(\alpha \ge 1\).
-
If \(2\sqrt{\alpha }<h<\alpha +1\) then the three roots 0, \(u_-\) and \(u_+\) are distinct and there is no multiple root.
-
If \(h=\alpha +1\) then either \(0=u_-\) or \(0=u_+\), depending on the sign of \(\alpha -1\). In both cases 0 is a multiple root.
-
Finally, if \(h>\alpha +1\) then \(u_-<0<u_+\) and all the roots of \(P(1,\alpha ,h,\cdot )\) have multiplicity one.
The above discussion shows, using the characterization of \(h^*(\alpha )\), that
$$\begin{aligned} h^*(\alpha )=\left\{ \begin{array}{l} 1+\alpha \quad \text {if}\,\alpha \le 1\\ 2\sqrt{\alpha }\quad \text {if}\, \alpha > 1.\end{array}\right. \end{aligned}$$(19)Note that \(\alpha \mapsto h^*(\alpha )\) belongs to \(C^1([0,+\infty ),\mathbb {R}^+)\). \(\square \)
-
-
Computation of \({\lim _{E\rightarrow +\infty }h^*(E,\alpha )}\)
Since \(h^*(\cdot ,\alpha )\) is positive and increasing, one has
$$\begin{aligned} \lim _{E\rightarrow +\infty }h^*(E,\alpha ) = \frac{1}{\ell _\alpha } \end{aligned}$$for some nonnegative number \(\ell _\alpha \) (wherein we have set \(\frac{1}{0}=+\infty \)).
However, since \(u_{crit}(E,\alpha )\) is bounded, up to a subsequence again denoted by E, one has \(u_{crit}(E,\alpha )\rightarrow \mu \) for some positive number \(\mu \) (eventually depending on \(\alpha \)). By passing to the limit in (16a), we deduce \(\mu >0\); for otherwise one obtains \(0=H(E,\alpha ,h^*,u_{crit})\rightarrow +\infty \). It follows that taking \(E\rightarrow +\infty \) in (16b), one obtains \(\mu =1{/}2\). Thus, by passing to the limit in (16a), one gets
$$\begin{aligned} \frac{1}{\ell _\alpha ^2}=\frac{2}{\ell _\alpha }+2\alpha \end{aligned}$$and finally (\(\ell _\alpha \) being nonnegative)
$$\begin{aligned} \frac{1}{\ell _\alpha }=2+2\sqrt{1+\alpha }; \end{aligned}$$\(\square \)
-
Computation of \({h^*(E,0)}\) If \(\alpha =0\) one gets
$$\begin{aligned} P(E,h,0,u)=\frac{1}{E}(1+Ehu)\left( -Ehu^2+u(Eh-1)+1-E\right) . \end{aligned}$$(20)Standard computations show that \(P(E,h,0,\cdot )\) admits two nonnegative roots if and only if \(h>\frac{1}{E}(2E-1+\sqrt{E(E-1)}:=h_1(E)\) and one double nonnegative root if \(h=h_1(E)\). This shows that \(h^*(E,0)=h_1(E)\). \(\square \)
-
Computation of \({\lim _{\alpha \rightarrow +\infty }h^*(E,\alpha )}\) Since \(E\mapsto h^*(E,\alpha )\) is increasing one gets for any \(E\ge 1\), \(h^*(E,\alpha )\ge h^*(\alpha )\). The explicit expression (19) of \(h^*(\alpha )\) implies \( \lim \nolimits _{\alpha \rightarrow +\infty }h^*(E,\alpha )=+\infty .\) \(\square \)
5.2 Proof of the Theorem 2.3
Let \(E>1\) be fixed and (u(t, x), v(t, x)) be a solution of (2) with initial condition \((u_0(x),v_0(x))\) verifying (3).
Step 1 It is clear that \(u(t,x)\ge 0\) for any \(t>0\) which implies
Thus, using the comparison principle and the hypothesis that \(v(0,x)\ge 1\), we deduce \(v(t,x)\ge 1\) for any \(t\ge 0\) and \(x\in \mathbb {R}\). It follows that
By the comparison principle, we infer that any solution \(\overline{u}\) of
such that \(\overline{u}(0,x)\ge u_0(x)\) for any \(x\in \mathbb {R}\) satisfies
In particular, let \(\phi (t)\) be the solution of the ordinary differential equation
together with the initial condition \(\phi (0)=1\).
\(\phi \) is a homogeneous solution of (21) and from \(u(0,x)\le 1=\phi (0)\) we deduce
Step 2 Behavior of \(\phi (t)\) as \(t\rightarrow +\infty \).
It is clear that 0 is always a steady state of (22) and is asymptotically stable for \(E>1\).
Now, let \(\phi _0> 0\) be a positive steady state of (22). \(\phi _0\) is a root of the polynomial \(P(E,h,0,\cdot )\) which is studied in the proof of the property 2.2 [see (20)]. Hence, if \(h\ge h_1(E)\) then (22) has two positive steady states that we denote as \(u^-(E,h)\le u^+(E,h)\le 1\) explicitly given by
A linear analysis shows that for \(h>h_1(E)\), \(u^-(E,h)\) is unstable and \(u^+(E,h)\) is (asymptotically) stable.
Finally, classical arguments show that \(\phi (t)\rightarrow u^+(E,h)\) if \(h>h_1(E)\) and \(\phi (t)\rightarrow 0\) if \(h<h_1(E)\). This ends the proof of the theorem. \(\square \)
5.3 Proof of the Theorem 2.4
Let \(E>1\) be fixed and (u(t, x), v(t, x)) be a solution to (2) with initial condition \((u_0(x),v_0(x))\) verifying (3).
By the argument of the step 1 of the proof of 5.2, one already knows that:
where \(\overline{u}(t,x)\) is a solution of (21) with \(\overline{u}(0,x)=u_0(x)\). Moreover, from the step 2 of the proof of 5.2, one knows that if \(h>h_1(E)\) then the Eq. (21) is bistable since (21) admits two stable nonnegative steady states: \(u=0\) and \(\overline{\mu }=u^+(E,h)<1\). We prove here that, in that case, there exists a traveling wave connecting \(\overline{\mu }\) to 0 at a negative speed if and only if \(h<h^-(E)\) for some (implicit) number \(h^-(E)>h_1(E)\). This implies that for any \(x\in \mathbb {R}\), \(\overline{u}(t,x)\rightarrow 0\) if \(h<h^-(E)\) and \(\overline{u}(t,x)\rightarrow \overline{\mu }\) if \(h>h^{-}(E)\), which proves the theorem.

Graph of \(u\mapsto -W(E,h,u)\). The steady states 0, \(u^-\) and \(u^+\) correspond to critical points of the potential \(-W\). A stable steady state is a local minimum of W and an unstable steady state is a local maximum. One sees that 0 and \(u^+\) are two stable steady states. The sign of c characterizes which of them is the final global attractor: if \(c<0\), then 0 is the global attractor. If \(c>0\), then \(u^+\) is the global attractor
Let \(h>h_1(E)\) be fixed. It can be proven (see Fife 1979; Volpert et al. 1994 and the reference therein) that there exists a unique speed \(c=c(E,h)\) such that (21) admits a traveling solution of speed c which connects \(\overline{\mu }\) to 0. More precisely, there exists a profile \(U(\xi )\) verifying \(U(-\infty )=\overline{\mu }\), \(U(+\infty )=0\) and \(U'(\pm \infty )=0\), such that \(U(x-ct)=u(x,t)\) is a solution of (21). Moreover (see Fife 1979) this traveling wave describes the asymptotic behavior of all solutions provided the initial condition (3) are verified. The theorem is proven by showing that there exists \(h^{-}(E)\) such that \(c(E,h)<0\) if \(h<h^{-}(E)\) and \(c(E,h)>0\) if \(h>h^{-}(E)\). This result is a direct consequence of the two following lemma. The first lemma gives a characterization of the sign of c by an explicit function (see Fig. 10).
Lemma 5.1
Define
Then \(sign(c(E,h))=sign(W(E,h,\overline{\mu }))\) where \(\overline{\mu }:=u^+(E,h)\).
Proof
Denote \(c=c(E,h)\) and let \(\xi =x-ct\) and \(U(\xi )=u(x,t)\) with \(U(-\infty )=\overline{\mu }\), \(U(+\infty )=0\) and \(U'(\pm \infty )=0\). We have
Multiplying by \(U^{\prime }\) and integrating over \(\mathbb {R}\) one gets
and \(sign(c)=sign(W(E,h,u^+(E,h))\) follows. \(\square \)
The second lemma gives the sign of W with respect to h.
Lemma 5.2
For any \(E>1\), there exists (a unique) \(h^-(E)>h_1(E)\) such that
-
There is invasion of prey for (21) (\(c(E,h)>0\)) if \(h\in (h^-(E),+\infty )\). In that case, for any \(x\in \mathbb {R}\), \(\overline{u}(t,x)\rightarrow u^+(E,h)\) as \(t\rightarrow +\infty \).
-
There is extinction of prey for (21) (\(c(E,h)<0\)) if \(h\in [h_1(E),h^-(E))\). In that case, for any \(x\in \mathbb {R}\), \(\overline{u}(t,x)\rightarrow 0\) as \(t\rightarrow +\infty \).
In particular, we infer from the inequality (24), that if \(h\in [h_1(E),h^-(E))\), then for any \(x\in \mathbb {R}\), \(u(t,x)\rightarrow 0\) as \(t\rightarrow +\infty \). This ends the proof of the theorem. \(\square \)
It remains to prove the Lemma 5.2.
Proof of Lemma 5.2
Define \(\mathcal {W}(E,h)=W(E,h,u^+(E,h))\). From the Lemma 5.1, we know that \(sign(c)=sign(\mathcal {W}(E,h))\). We show here that there exists \(h^-(E)>h_1(E)\) such that \(sign(\mathcal {W}(E,h))=sign(h-h^-(E))\).
Step 1 Differentiate \(\mathcal {W}\) with respect to h gives
By the very definition of W and \(u^+\), one has \(\partial _u W(E,h,u^+(E,h))=0\), which yields
Explicit calculations show that
Differentiate this expression with respect to h and denoting \(z=Ehu^+(E,h)\) provides
A standard analysis shows that the map \(z\mapsto z-2\ln (1+z)+\frac{z}{1+z}\) takes positive values for \(z>0\). It follows that the map \(h\mapsto \mathcal {W}(E,h)\) is increasing.
Step 2 Recalling that \(u^+\) is the largest roots of (20), one verifies that \(u^+(E,h)\rightarrow 1\) as \(h\rightarrow +\infty \) and \(\mathcal {W}(E,+\infty )=1/2-1/3=1/6>0\).
Step 3This step consists in proving that for any \(E>1\), \(\mathcal {W}(E,h_1(E))=W(E,h_1(E),u^+(E,h_1(E))):=g(E)\) is negative.
From the explicit expression (23) of \(u^+(E,h)\) and the definition of \(h_1(E)\), we get
On a first hand, we have
Straightforward calculations give
wherein we have set
A standard analysis shows that \(2\frac{z}{z+2}<\ln (1+z)\) for any \(z>0\). Thus \(g^{\prime }(E)<0\) for any \(E>1\).
On another hand, we have \(h_1(1)=1\) and \(u^+(1,1)=0\), so that \(g(1)=0\). It follows that \(g(E)<0\) for any \(E>1\).
Conclusion For any \(E> 1\), the map \(h\mapsto \mathcal {W}(E,h)\) is increasing and verifies \(\mathcal {W}(E,h_1(E))<0\) and \(\lim \nolimits _{h\rightarrow +\infty }\mathcal {W}(E,h)=1/6\). By continuity, for any \(E>1\), there exists a unique \(h^-=h^-(E)\in (h_1(E),+\infty ) \) such that \(sign(W(E,h,u^+))=sign(h-h^-(E))\). This ends the proof of lemma 5.2. \(\square \)
Proofs of Properties 2.5
Let \(E>1\) be fixed.
-
Growth of \(h^-(\cdot )\)
Recall that \(h^-(E)\) is characterized by
$$\begin{aligned} W\left( E,h^-(E),u^+(E,h^-(E))\right) =0 \end{aligned}$$(26)where the expressions of W and \(u^+\) are respectively given in (25) and (23). By definition of W, one has
$$\begin{aligned} \partial _u W\left( E,h^-(E),u^+\left( E,h^-(E)\right) \right) =0. \end{aligned}$$Hence
$$\begin{aligned} \partial _E W\left( E,h^-(E),u^+(E,h^-(E))\right) +\frac{d h^-}{dE} (E)\cdot \partial _h W\left( E,h^-(E),u^+(E,h^-(E))\right) =0. \end{aligned}$$The explicit computation of \(\partial _E W\) and \(\partial _h W\) are done in proof 5.3 and one gets
$$\begin{aligned} \partial _E W\left( E,h^-(E),u^+(E,h^-(E))\right) <0\quad \text { and }\quad \partial _h W\left( E,h^-(E),u^+(E,h^-(E))\right) >0 \end{aligned}$$so that
$$\begin{aligned} \frac{d h^-}{dE} (E)=-\frac{\partial _E W\left( E,h^-(E),u^+\left( E,h^-(E)\right) \right) }{ \partial _h W\left( E,h^-(E),u^+\left( E,h^-(E)\right) \right) }>0 \end{aligned}$$as needed. \(\square \)
-
Limit of \(h^-(E)\) as \(E\rightarrow 1\)
Let \(E>1\). One knows that \(h^{-}(\cdot )\) is increasing on \((1,+\infty )\) and minored by \(h_1(E)>0\). Hence \(h^-(E)\) admits a limit \(\ell ^-\) as \(E\rightarrow 1\), and from \(h_1(1)=1\), one obtains
$$\begin{aligned} \ell ^-\ge 1. \end{aligned}$$The explicit expressions (23) of \(u^\pm \) yield
$$\begin{aligned} u^{-}\left( 1,\ell ^-\right) =0\text { and }u^+\left( 1,\ell ^-\right) =\frac{1-\ell ^-}{\ell ^-}. \end{aligned}$$Assume by contradiction that \(u^+(1,\ell ^-)>0\). Then, for any \(h\ge 1\) and \(0< u<u^+(h,1)\), we have
$$\begin{aligned} \partial _uW(1,h,u)=u(1-u)-\frac{1}{1+hu}>0\quad \text {and}\quad W(1,h,0)=0. \end{aligned}$$This implies \(W(1,\ell ^-,u^+(1,\ell ^-))<0\), which contradicts (26).
It follows that \(u^+(1,\ell ^-)=0\). Thus \(\ell ^-=1\), which reads \(\lim \limits _{E\rightarrow 1} h^-(E)=1.\) \(\square \)
-
Limit of \(h^-(E)\) as \(E\rightarrow +\infty \)
One knowns that \(h^-(E)\) is increasing so that there is some \(\ell \ge 0\) such that \({\lim _{E\rightarrow +\infty }h^-(E)= \frac{1}{\ell }}\) (wherein we have set \(\frac{1}{\infty }=0\)). By taking the limit \(E\rightarrow +\infty \) in (23), we obtain
$$\begin{aligned} \lim _{E\rightarrow +\infty } u^+\left( E,h^-(E)\right) =u_\ell :=\frac{1}{2}\left( 1+\sqrt{1-4\ell }\right) >0. \end{aligned}$$and then, by taking the limit in (26),
$$\begin{aligned} \frac{u_\ell ^2}{2}-\frac{u_\ell ^3}{3}-\frac{u_\ell }{\ell }=0. \end{aligned}$$Thus, \(\ell =\frac{3}{16}\) which reads, \(\lim \nolimits _{E\rightarrow +\infty } h^-(E)=\frac{16}{3}.\)
5.4 Invasion conditions, proof of Theorem 2.6
As in the proof 5.3, we start here by showing that \(0\le \underline{u}(t,x)\le u(t,x)\) for some function \(\underline{u}\) verifying a scalar reaction–diffusion equation (27) depending on E, h and \(\alpha \). Next, we show that there exists \(h^+(E,\alpha )\) such that \(\lim _{t\rightarrow +\infty }\underline{u}(t,x)=\underline{\mu }>0\) when \(h>h^+(E,\alpha )\). This step is done using the proof of Theorem 2.4 together with an appropriate change of variables.
Step 1 Let \(E>1\) be fixed. In general, using \(0\le u\le 1\), one has \(1\le v\le \overline{v}\) with
From the estimate \(v\le \overline{v},\) we get \(\underline{u}(t,x)\le u(t,x)\) where \(\underline{u}\) verifies
Denoting \(\widetilde{E}=E\overline{v}\) and \(\widetilde{h}=\frac{h}{\overline{v}}\), this equation reads simply
Note that, removing the \(\widetilde{}\), this equation is nothing but (21). As a consequence, the proof of the Theorem 2.4 implies the following result on (28).
Lemma 5.3
Let \(\widetilde{u}(t,x)\) be a solution of (28) verifying the initial condition (3). There exists \(h^-(\widetilde{E})>h_1(\widetilde{E})\) such that
-
if \(\widetilde{h}<h^-(\widetilde{E})\) then for any \(x\in \mathbb {R}\), \(\lim \nolimits _{t\rightarrow +\infty }\widetilde{u}(t,x)= 0\) (extinction),
-
if \(\widetilde{h}>h^-(\widetilde{E})\) then for any \(x\in \mathbb {R}\), \(\lim \nolimits _{t\rightarrow +\infty }\widetilde{u}(t,x)= \overline{\mu }(\widetilde{E},\widetilde{h})>0\) (invasion).
Step 2 Recalling that \(\widetilde{E}=E \overline{v}(E,h,\alpha )\) and \(\widetilde{h}=\frac{h}{ \overline{v}(E,h,\alpha )}\) do depend on h, we set \(\underline{\mu }=\underline{\mu }(E,h)=\overline{\mu }(\widetilde{E},\widetilde{h})\) and the previous lemma gives the following implicit condition on h for invasion to occur.
For any \(x\in \mathbb {R}\), \(\lim \limits _{t\rightarrow +\infty }u(t,x)= \underline{\mu }\), provided:
The following lemma gives an equivalent condition for this implicit condition to occur. This ends the proof of theorem 2.4.
Lemma 5.4
For any \(E>1\) and \(\alpha \ge 0\), there exists (a unique) \(h^+(E,\alpha )\ge h^-(E)\) such that:
Proof of Lemma 5.4
Let \(E>1\) and \(\alpha \ge 0\) be fixed and define the function
By the construction of \(h^-\) via the implicit function theorem, one knows that \(\mathcal {F}_{E,\alpha }(\cdot )\) is a \(C^1\) function. Moreover, since \(h^-\) and \(\overline{v}\) are bounded, we have \(\mathcal {F}_{E,\alpha }(+\infty )=+\infty \) and
Therefore, it suffices to show that \(\mathcal {F}_{E,\alpha }\) is increasing.
One has
From the expression of \(\overline{v}\), we infer \(\partial _h\overline{v}<0\) and from the proof of the properties 2.5, we know that \(\frac{dh^-}{dE}>0\). It follows that \(\mathcal {F}_{E,\alpha }^{\prime }(h)>0\). This ends the proof of the lemma. \(\square \)
Proofs of Properties 2.7
-
Proof of the growth of \(h^+(E,\cdot )\) and \(h^+(\cdot ,\alpha )\)
\(h^+(E,\alpha )\) is characterized by an equality in (29), that is,
$$\begin{aligned} \mathcal {F}_{E,\alpha }\left( h^+(E,\alpha )\right) =0 \end{aligned}$$(31)where \(\mathcal {F}_{E,\alpha }\) is defined in (30). Differentiating (31) with respect to E gives
$$\begin{aligned} \partial _E h^+(E,\alpha )\cdot \mathcal {F}^{\prime }_{E,\alpha }\left( h^+(E,\alpha ) \right) +\partial _E \mathcal {F}_{E,\alpha } \left( h^+(E,\alpha )\right) =0 \end{aligned}$$and then
$$\begin{aligned} \partial _E h^+(E,\alpha )\cdot \mathcal {F}^{\prime }_{E,\alpha }\left( h^+(E,\alpha )\right) = \left[ \left( E\overline{v}+h^-\right) \partial _E \overline{v}+\overline{v}^2\right] \cdot \frac{dh^-}{dE}\left( E \overline{v}\right) . \end{aligned}$$wherein we have set
$$\begin{aligned} h^-=h^-\left( E \overline{v}\right) ,\quad \overline{v}=\overline{v}\left( E,h^+(E,\alpha ),\alpha \right) \quad \text { and }\quad \partial _E\overline{v}=\partial _E\overline{v}\left( E,h^+(E,\alpha ),\alpha \right) . \end{aligned}$$One already knows that \(\mathcal {F}^{\prime }_{E,\alpha }(h^+(E,\alpha ) )>0\) and that \(\frac{dh^-}{dE}>0\). A direct computation shows that \(\partial _E\overline{v}>0\), which leads to \(\partial _E h^+(E,\alpha )>0\) as needed.
Similarly, differentiating (31) with respect to \(\alpha \) gives, with obvious notations,
$$\begin{aligned} \partial _\alpha h^+(E,\alpha )\cdot \mathcal {F}^{\prime }_{E,\alpha }(h^+(E,\alpha ) )=\partial _\alpha \overline{v}\cdot \left( h^-+E\frac{dh^-}{dE}\right) , \end{aligned}$$and since \(\partial _\alpha \overline{v}>0\), one obtains \(\partial _\alpha h^+(E,\alpha )>0\).
-
Limits of \(h^+(E,\alpha )\) as \(\alpha \rightarrow 0\) and \(\alpha \rightarrow +\infty \)
From \(\overline{v}(E,h,0)=1\) we deduce \(h^+(E,0)=h^-(E)\). Now, it is clear from the construction of \(h^+\), that \(h^+(E,\alpha )\ge h^*(E,\alpha )\). From the properties 2.2, we obtain \(h^+(E,\alpha ) \rightarrow +\infty \) as \(\alpha \rightarrow +\infty .\) \(\square \)
-
Limit of \(h^+(E,\alpha )\) as \(E\rightarrow +\infty \).
First, recall that \(h^+(E,\alpha )\) is characterized by (31), which reads
$$\begin{aligned} h^+(E,\alpha )=\left( 1+\alpha \frac{E}{1+Eh^+(E,\alpha )}\right) \cdot h^-\left( E+\alpha \frac{E^2}{1+Eh^+(E,\alpha )}\right) . \end{aligned}$$(32)Since \(h^+(\cdot ,\alpha )\) is increasing, there exists \(\ell _\alpha \ge 0\) such that \(\lim _{E\rightarrow +\infty } h^+(E,\alpha )=\frac{1}{\ell _\alpha }.\) Taking the limit \(E\rightarrow +\infty \) in (32) and using \(h^-(E)\rightarrow \frac{16}{3}\), one obtains \(\frac{1}{\ell _\alpha }=\frac{16}{3} \cdot (1+\alpha \ell _\alpha )\). The resolution of this equation ends the proof. \(\square \)
Notes
All our results remain true for various different initial conditions. The essential condition is that the solutions of the scalar systems we consider converge to traveling wave solutions. In particular, compact support may be allowed for \(u_0\). See Fife (1979) for a detailed discussion.
It suffices that this condition occurs for \(t>t_0\) for some \(t_0>0\).
The dynamics generally also depends on a quantity \(h^{**}\), defined in the theorem 2.1, slightly greater than \(h^*\) that is not taken into account here for the sake of simplicity.
Remark that \(h^*>\frac{1}{E}\), because \(h\le \frac{1}{E}\), implies that \(f_h'<0\) on (0, 1) and (8) has no solution.
These arguments also show that \(u\mapsto h(u)\) is decreasing on \((0,u^*)\) and increasing on \((u^*,1)\); for otherwise it is possible to choose \(h>0\) such that there is at least four different \(u\in (0,1)\) such that \(h=h(u)\), which is equivalent to \(P_h\) having at least four roots. See the Fig. 9.
References
Alzahrani EO, Davidson FA, Dodds N (2012) Reversing invasion in bistable systems. J Math Biol 65:1101–1124
Basnet K, Mukhopadhyay A (2014) Biocontrol potential of the lynx spider Oxyopes javanus (Araneae: Oxyopidae) against the tea mosquito bug, Helopeltis theivora (Heteroptera:Miridae). Int J Trop Insect Sci 34(4):232–238
Braverman E, Kamrujjaman M, Korobenko L (2015) Competitive spatially distributed population dynamics models: does diversity in diffusion strategies promote coexistence? Math Biosci 264:63–73
Boukal DS, Sabelis MW, Berec L (2007) How predator functional responses and Allee effects in prey affect the paradox of enrichment and population collapses. Theor Popul Biol 72:136–147
Cai Y, Banerjee M, Kang Y, Wang W (2014) Spatiotemporal complexity in a predator–prey model with weak allee effects. Math Biosci Eng 11:1247–1274
Chakraborty S (2015) The influence of generalist predators in spatially extended predator–prey systems. Ecol Complex 23:50–60
De Roos AM, Mccauley E, Wilson WG (1991) Mobility versus density-limited predator–prey dynamics on different spatial scales. Proc R Soc Lond B Biol Sci 246(1316):117–122
De Roos AM, Mccauley E, Wilson WG (1998) Pattern formation and the spatial scale of interaction between predators and their prey. Theor Popul Biol 53(2):108–130
Du Y, Shi J (2007) Allee effect and bistability in a spatially heterogeneous predator–prey model. Trans Am Math Soc 359(9):4557–4593
Ducrot A, Langlais M (2012) A singular reaction–diffusion system modelling prey–predator interactions: invasion and co-extinction waves. J Differ Equ 253:502–532
Dunbar SR (1984a) Traveling wave solutions of diffusive Lotka–Volterra equations. J Math Biol 17:11–32
Dunbar SR (1984b) Traveling wave solutions of diffusive Lotka–Volterra equations: a heteroclinic connection in R4. Trans Am Math Soc 286:557–594
Erbach A, Lutscher F, Seo G (2014) Bistability and limit cycles in generalist predator–prey dynamics. Ecol Complex 14:48–55
Fagan WF, Lewis MA, Neurbert MG, Pvd Driessche (2002) Invasion theory and biological control. Ecol Lett 5:148–157
Fife PC (1979) Long time behavior of solutions of bistable nonlinear diffusion equations. Arch Ration Mech Anal 70:31–46
Fischer RA (1937) The wave of advance of an advantageous gene. Ann Eugen 7:353–369
Garnier J, Roques L, Hamel F (2012) Success rate of a biological invasion in terms of the spatial distribution of the founding population. Bull Math Biol 74(2):453–473
Giovangigli V (1990) Nonadiabatic plane laminar flames and their singular limits. SIAM J Math Anal 21(5):1305–1325
Hastings A (2000) Parasitoid spread: lessons for and from invasion biology. In: Hochberg ME, Ives AR (eds) Parasitoids population biology. Princeton University Press, Princeton, pp 70–82
Hoyle A, Bowers RG (2007) When is evolutionary branching in predator–prey systems possible with an explicit carrying capacity? Math Biosci 210:1–16
Huang J, Lu G, Ruan S (2003) Existence of traveling wave solutions in a diffusive predator–prey model. J Math Biol 46:132–152
Huang Y, Weng P (2013) Traveling waves for a diffusive predator–prey system with a general functional response. Nonlinear Anal Real World Appl 14:940–959
Krivan V, Eisner J (2006) The effect of the Holling type II functional response on apparent competition. Theor Popul Biol 70:421–430
Kolmogorov AN, Petrowskii I, Piscounov N (1937) Etude de l’équation de la diffusion avec croissance de la quantité de matiére et son application à un problème biologique. Mosc Univ Math Bull 1:1–25
Lam KY, Ni WM (2012) Uniqueness and complete dynamics in heterogeneous competition–diffusion systems. SIAM J Appl Math 72:1695–1712
Li WT, Wu SL (2008) Traveling waves in a diffusive predator-prey model with Holling type-III functional response. Chaos Solitons Fractals 37:476–486
Lewis MA, Maini PK, Petrovskii SV (2013) Dispersal, individual movement and spatial ecology. Springer, Berlin
Li DS, Liao C, Zhang BX, Song ZW (2014) Biological control of insect pests in litchi orchards in China. Biol Control 68:23–36
Magal C, Cosner C, Ruan S, Casas J (2008) Control of invasive hosts by generalist parasitoids. Math Med Biol 25:1–20
Mistro DC, Rodrigues LAD, Petrovskii S (2012) Spatiotemporal complexity of biological invasion in a space- and time-discrete predator–prey system with the strong Allee effect. Ecol Complex 9:16–32
Moffat CE, Lalonde RG, Ensing DJ, De Clerck-Floate RA, Grosskopf-Lachat G, Pither J (2013) Frequency-dependent host species use by a candidate biological control insect within its native European range. Biol Control 67:498–508
Morozov A, Petrovskii S (2009) Excitable population dynamics, biological control failure, and spatiotemporal pattern formation in a model ecosystem. Bull Math Biol 71:863–887
Morozov A, Petrovskii S (2013) Feeding on multiple sources: towards a universal parameterization of the functional response of a generalist predator allowing for switching. PLoS One 8(9):e74586. doi:10.1371/journal.pone.0074586
Owen MR, Lewis MA (2001) How predation can slow, stop or reverse a prey invasion. Bull Math Biol 63:655–684
Pimentel D (2011) Biological invasions: economic and environmental costs of alien plant, animal, and microbe species, 2nd edn. CRC Press, New York
Potapov A, Rajakaruna H (2013) Allee threshold and stochasticity in biological invasions: colonization time at low propagule pressure. J Theor Biol 337:1–14
Savage D, Renton M (2013) Requirements, design and implementation of a general model of biological invasion. Ecol Model 272:394–409
van Leeuwen E, Jansen VAA, Bright PW (2007) How population dynamics shape the functional response in a one-predator–two-prey system. Ecology 88(6):1571–1581
van Leeuwen R, Brannsrom A, Jansen VAA, Dieckmann U, Rossberg AG (2013) A generalized functional response for predators that switch between multiple prey species. J Theor Biol 328:89–98
Volpert AI, Volpert VA, Volpert VA (1994) Traveling wave solutions of parabolic systems. American Mathematical Society, Providence
Wang W, Feng X, Chen X (2013) Biological invasion and coexistence in intraguild predation. J Appl Math 2013:12. doi:10.1155/2013/925141
Xu Z, Weng P (2012) Traveling waves in a diffusive predator–prey model with general functional response. Electron J Differ Equ 197:1–13
Ye GY, Xiao Q, Chen M, Chen X, Yuan Z, Stanley DW, Hu C (2014) Tea: biological control of insect and mite pests in China. Biol Control 68:73–91
Acknowledgements
The authors would like to thank the two anonymous reviewers for their valuable comments and suggestions to improve the quality of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Madec, S., Casas, J., Barles, G. et al. Bistability induced by generalist natural enemies can reverse pest invasions. J. Math. Biol. 75, 543–575 (2017). https://doi.org/10.1007/s00285-017-1093-x
Received:
Revised:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00285-017-1093-x
Keywords
- Reaction diffusion system
- Long time dynamics
- Traveling wave
- Invasion process
- Biological control
- Prey–predator interaction
- Generalist predator