
Abstract
Purpose
To investigate the occurrence of apoptosis and autophagy on human gastric cancer cells after treatment by Tanshinone I, as well as the relationship between them.
Methods
BGC823 and SGC7901 cells were treated with Tanshinone I; the cell proliferation was measured using CCK-8 and clone formation assay; and the expression of apoptosis- and autophagy-associated proteins was detected by Western blot. Autophagic vacuoles in cells were observed with LC3 dyeing using confocal fluorescent microscopy, and apoptotic cells were detected via flow cytometry. Bcl-2 was overexpressed in gastric cells treated with Tanshinone I or not, and autophagy relative protein was investigated; the interaction between Beclin-1 and Bcl-2 was detected by immunoprecipitation. Cell apoptosis was detected when autophagy was inhibited by ATG7-siRNA. Tumor growth was assessed by subcutaneous inoculation of cells into BALB/c nude mice.
Results
Tanshinone I inhibited the proliferation of BGC823 and SGC7901 cells, and induced cell apoptosis by inhibiting anti-apoptosis protein Bcl-2. Tanshinone I also increased the conversion of LC3I to LC3II and triggered autophagosome formation, without changing the expression of Beclin-1. However, the Beclin-1 VPS34 complexes were increased after Tanshinone I treatment via inhibiting Bcl-2 expression. Moreover, disturbing autophagy by knockdown of ATG7 expression contributed to Tanshinone I-induced apoptosis. In vivo assay showed that combination with autophagy inhibitor chloroquine in nude mice bearing BGC823 xenograft significantly augmented the antitumor effect of Tanshinone I.
Conclusions
Tanshinone I induced apoptosis and pro-survival autophagy via inhibiting Bcl-2 expression on gastric cancer, and the combination of chloroquine and Tanshinone I could inhibit tumor growth more efficiently than monotherapy, which might be considered as an effective strategy for the treatment for gastric cancer.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Gastric cancer is the third leading cause of cancer-related death in the world. At present, the conventional treatment approach of gastric cancer is gastrectomy combined with chemotherapy or radiotherapy [1]. Due to the limited efficacy of existing drugs, the advanced gastric cancer still has a poor prognosis. Therefore, it is important to develop novel effective antitumor agents with known mechanism of action, including traditional Chinese herbal medicines.
Danshen (Salvia miltiorrhiza Bunge) is a traditional Chinese herb that has been successfully used for treating coronary heart diseases in clinics [2, 3]. More than 40 diterpene compounds (including the relatively abundant Tanshinones, Tanshinone I, Tanshinone IIA, crypto Tanshinone, and dihydro Tanshinone) have been isolated from Danshen [4]. These Tanshinones have shown cytotoxic effects on cell lines derived from human carcinomas of the colon, ovary, lung, mouth, and breast [5–11]. Recent findings indicate Tanshinone I can induce cyclin D1 proteasomal degradation and apoptosis in colon cancer [12, 13]. However, the antitumor activity of Tanshinone I on human cancer cell lines is not effective enough. Therefore, understanding the mechanisms of Tanshinone I action was critical for improving its anticancer effects.
Autophagy is a catabolic degradation process whereby cellular proteins and organelles are digested in lysosomes and recycling maintains cellular biosynthesis [14]. Accumulating evidence has shown natural products that have antitumor effects can induce autophagy in different cancer cell types. Resveratrol present in grapes can induce autophagy in ovarian cancer cell lines [15]. Triterpenoid saponins isolated from soybeans induce autophagy in colon cancer cells with decreasing Akt expression and increasing ERK expression [16]. In addition, EB1089, which is a vitamin D analog, induces autophagy in MCF-7 cells, and overexpression of Beclin-1 can sensitize these cells to EB 1089 [17]. However, drug-induced autophagy in tumor cells is a cell survival mechanism that suppresses apoptosis and eventually leads to treatment resistance [18, 19]. Moreover, no data upon the effect of Tanshinone I on apoptosis and autophagy have been reported until now. To understand the cross talk between autophagy and apoptosis, we investigated the effect of Tanshinone I on apoptosis and autophagy of gastric BGC823 and SGC7901 cells.
Materials and methods
Reagents
Tanshinone I, 5-diphenyl-2-H-tetrazolium bromide (MTT), chloroquine, propidium iodide (PI), and 3-methyladenine (3-MA) were purchased from Sigma, USA.
Streptomycin, penicillin, and Lipofectamine 2000 were purchased from Life Technology, USA.
Cell culture
The human BGC823 and SGC7901 gastric cancer cell lines were purchased from the American Type Culture Collection (ATCC). Cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM, Life Technology, USA) supplemented with 10 % fetal bovine serum (FBS, Life Technology, USA), streptomycin (100 U/mL), and penicillin (100 U/mL) at 37 °C in a humidified incubator of 5 % CO2.
Cytotoxicity assay
The cytotoxicity of Tanshinone I was assayed by the CCK-8 kit (Dojindo Laboratories, Japan). Cells were seeded at a density of 5 × 103/well in 100 μl of medium onto 96-well microplates (Costar Corning, USA). After 24-h incubation, Tanshinone I was added at various concentrations 2.5, 5, 10, 15, 20, 40 μg/ml, and five biological repeats were included for each concentration. After treatment with Tanshinone I for 48 h, 10 μl of CCK-8 was added to each well and cultivated for another 2 h. Absorbance at 450 nm was measured with a microplate reader (Bio-Rad, USA), using wells without cells as blanks. All experiments were performed in triplicate. The effect of Tanshinone I on the proliferation of cells was expressed as the inhibitory ratios (%), using the following formula: Cell inhibitory ratios (%) = [A450 (control) − A450 (Tanshinone I)]/[A450 (control) − A450 (blank)] × 100.
Clone formation assay
The colony is defined at least 50 cells. For clone formation assay, approximately 150 cells were plated in 24-well plates and treated with 2.5 μg/ml Tanshinone I for 2 weeks. Colonies were counted under microscopy after crystal violet staining. Clone formation rate = clone number/plated number × 100 %.
Deficiency in ATG7
To produce cells deficient in ATG7, siRNA targeting ATG7 was designed and applied to GBC823 and SGC7901 cells cultured in 75-cm2 flasks at 5 × 105 cells/flask. Briefly, three different siRNAs targeting ATG7 and negative-mismatched siRNA were purchased from RiboBio (CHINA). The optimal transfection efficiency was determined from our preliminary studies testing the range from 5 to 50 nM, and we selected the condition and the siRNA sequence (from three different sequences) for ATG7 protein that can cause about 60–70 % silencing effect with minimal cytotoxicity. GBC823 and SGC7901 cells were transfected with 50 nM of the selected annealed siRNAs targeting ATG7 or negative-mismatched siRNA in antibiotic-free media. Lipofectamine 2000 (Life Technology, USA) was used as the transfection reagent following the manufacturer’s instruction. After 48-h transfection, cells were trypsinized and collected for further analysis as described in the experimental procedure.
Flow cytometric analysis of apoptosis
Annexin V-FITC Apoptosis Detection Kit (Merck Millipore, USA) was used to evaluate the percentage of apoptosis cells. Cells were treated with Tanshinone I at different concentrations, or other while pretreated with 3-MA for 1 h before the addition of Tanshinone I. 5 × 105 treated cells were centrifuged at 1000×g for 5 min at room temperature followed by washing with cold PBS. Cells were re-suspended in cold 1 × binding buffer and added with 1.25 μl Annexin V-FITC for 15 min of incubation at room temperature in the dark. After 10 µl of propidium iodide (PI) was added, the suspension was analyzed by flow cytometry (FACScan).
Fluorescence microscopy for LC3 punctate
BGC823 and SGC7901 were seeded on poly-lysine-coated coverslips overnight and then treated with 2.5 μg/mL Tanshinone I for 24 h. Cells were fixed, dyed with LC3-FITC antibody and then mounted using SlowFade with 4,6-diamidino-2-phenylindole (DAPI; Life Technology, USA), and examined under a × 100 lens on an confocal fluorescence microscope.
Western blotting
Treated cells were collected and washed with PBS twice. The pellets were suspended in mammalian protein extraction reagent (Thermo fisher, USA) for at least 30 min on ice. The lysis buffer also contained 1 mM protease inhibitor cocktail, Na3VO4, NaF. Cell lysates were clarified by centrifugation at 12,000 × g for 10 min at 4 °C, and the supernatants were mixed with 5 × loading buffer and boiled for 10 min. The protein contents in the supernatant were measured by using a BCA protein assay kit II (Bio-Rad). Proteins were separated by SDS-PAGE electrophoresis and electrotransferred onto a PVDF membrane. The membrane was blocked in 5 % nonfat skim milk and probed with primary antibodies of PARP, Beclin-1, Bcl-2, VPS34, Bax, cleaved caspase 3, and LC3 (Cell signaling technology, USA) followed by incubation with horseradish peroxidase-conjugated secondary antibodies. Protein expression was detected by using enhanced chemiluminescence system (Amersham Pharmacia).
Tumor xenograft studies
All mouse experiments were performed in accordance with approved protocols from Shanghai Medical Experimental Animal Care Commission. GBC823 cells (5 × 105) in 100 µl of DMEM medium were injected subcutaneously in the right lateral flank of six-week-old nude mice. When the tumor volumes reached an average of approximately 100 mm3, we treated the mice with vehicle control, Tanshinone I at a concentration of 20 mg/kg q.d. or chloroquine at a concentration of 50 mg/kg q.d or the combination for 2 weeks p.o. Tumor volumes were measured every 3 days in two dimensions with vernier calipers. The tumor volumes were calculated using the following formula: length × width2 × 0.5. Two weeks after the final treatment, the mice were killed, and the tumors were surgically excised and weighed. Tumor tissues from the in vivo experiments were collected for Western blot analysis.
TUNEL assay
The TUNEL assay was performed according to the kit instructions (Qia39, Merck, USA). The tumor tissue sections were deparaffinized, rehydrated, and incubated with proteinase K (20 μg/mL) for 20 min at 37 °C. After several washes with TBS, the specimen was covered in 1 × equilibration buffer for 30 min and then incubated with a mixture of 57.0 µl of Fluorescein-FragELTM TdT Labeling Reaction Mix and 3 µl of TdT Enzyme for 1.5 h at 37 °C in the dark. Then, the slides were rinsed in TBS three times. A glass coverslip was mounted using Fluorescein-FragELTM Mounting Media, and slides were visualized under a fluorescence microscope (OLYMPUS IX71, Japan). TUNEL-positive cells were counted at × 400 magnification. The apoptotic index was calculated as a ratio of (apoptotic cell number)/(total cell number) in each field.
Statistical analysis
All data are presented as the mean ± SD. Statistical significance was determined by paired or unpaired Student’s t test in cases of standardized expression data. One-way ANOVA was performed for multiple group comparisons, and comparisons between two groups were conducted using the least significant difference method. P < 0.05 was considered significant.
Results
Growth inhibition by Tanshinone I in human gastric cancer cells
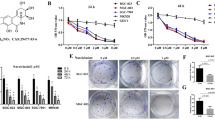
To evaluate the inhibitory effect of Tanshinone I on gastric cancer cells, BGC823 and SGC7901 cells were treated with gradient Tanshinone I for 48 h. As shown in Fig. 1a, b, the relative viability of cells subsequent to Tanshinone I treatment was decreased in a dose-dependent manner. In addition, colony formation assay suggested that the number of developed colonies generated from Tanshinone I-treated BGC823 and SGC7901 cell was less than controls (Fig. 1c, d). It was concluded that Tanshinone I could inhibit the BGC823 and SGC7901 cells growth in a gradient-dependent manner.

Tanshinone inhibited BGC823 and SGC7901 cell proliferation. a, b Cells viability of BGC823 (a) and SGC7901 (b) after Tanshinone I treatment at 2.5, 5, 10, or 20 μg/ml by CCK-8 assay. c, d Colony formation of BGC823 and SGC7901 cells after Tanshinone I treatment at 2.5 μg/ml. The data are expressed as the mean value ± SD. Statistical significance is indicated versus control (*P < 0.05; **P < 0.01; ***P < 0.001)
Apoptosis was induced by Tanshinone I in human gastric cancer cells
To examine whether Tanshinone I-induced cytotoxicity was due to the induction of apoptotic cell death, we investigated the level of apoptosis exerted by Tanshinone I in gastric cancer cells. Apoptosis in BGC823 and SGC7901 cells treated with Tanshinone I was analyzed by flow cytometry with Annexin V and PI staining. As shown in Fig. 2a, the apoptotic index was significantly increased in gastric cancer cells treated with Tanshinone I. We then examined the apoptosis-associated protein expression levels in BGC823 and SGC7901 cells. As shown in Fig. 2b, Tanshinone I treatment increased the expression of Bax and cleaved caspase 3 and PARP, while inhibited the expression of Bcl-2, a negative regulatory factor for autophagy. Taken together, Tanshinone I could induce apoptosis in BGC823 and SGC7901 cells.

Tanshinone I induced apoptosis in BGC823 and SGC7901 cells. a FACS assessing apoptosis induced in BGC823 and SGC7901 cells after Tanshinone I treatment at 5 and 10 μg/ml. The data are expressed as the mean value ± SD. Statistical significance is indicated versus control (*P < 0.05; **P < 0.01; ***P < 0.001). b Western blot assessing the change in apoptosis-related protein including caspase 3, PARP, Bax, and Bcl-2
Autophagy was induced by Tanshinone I in human gastric cancer cells
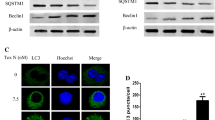
Due to natural products which can also induce autophagy in some cancer cells, we next explore whether Tanshinone I could induce autophagy. Autophagy in BGC823 and SGC7901 cells was detected via testing LC3 puncta by immunofluorescence staining with LC3 antibody and the conversion of LC3 I to LC3 II by Western blot, which are both classic index for autophagy detection [20]. Tanshinone I treatment resulted in significant autophagy induction, as evidenced by an increase in LC3 puncta (autophagosomes) (Fig. 3a). Consistently, Western blot data showed that Tanshinone I was able to cause an increased conversion of LC3-I into LC3-II in BGC823 and SGC7901 cells (Fig. 3c). However, the expression of Beclin-1, a vital autophagy-induced protein, was not changed. Taken these together, Tanshinone I can induce both apoptosis and autophagy in human gastric cancer cells.

Tanshinone I induced autophagy in BGC823 and SGC7901 cells. a Immunofluorescence evaluated the effect of Tanshinone I on LC3 punctates in BGC823 and SGC7901 cells. b LC3B and other autophagy-related protein were analyzed by Western blot
Tanshinone I inhibited Beclin-1 and Bcl-2 complex and induced autophagy in BGC823 and SGC7901 Cells
Bcl-2 is a negative regulatory factor for autophagy, which binds to Beclin-1 and disrupts the interaction between Beclin-1 and VPS34, leading to Beclin-1 homodimer formation and inhibiting autophagosome formation [21, 22]. To investigate whether Bcl-2 was involved in Tanshinone I-induced autophagy, Bcl-2 was overexpressed in BGC823 and SGC7901 cells to disturb the interaction between Beclin-1 and VPS34. As shown in Fig. 4a, Bcl-2 overexpression significantly inhibited Tanshinone I-induced autophagy. Immunoprecipitation results indicated that overexpression of Bcl-2 increased Beclin-1/Bcl-2 complex and decreased VPS34/Beclin-1 complex, which could inhibit autophagosome formation. Our data indicated that Tanshinone I induces autophagy though downregulating of Bcl-2/Beclin-1 combination and contributing to VPS34/Beclin-1 complex in BGC823 and SGC7901 cells.

Tanshinone I inhibited Beclin-1/Bcl-2 complex and induced autophagy in BGC823 and SGC7901 cells. a Overexpression of Bcl-2 inhibited Tanshinone I-induced autophagy by Western blot assay. b IP assessing Beclin-1 and Bcl-2 complex, and Beclin-1 and VPS34 complex after Bcl-2 overexpression or not
Combining Tanshinone I with autophagy inhibition enhanced Tanshinone I-induced cytotoxicity in vitro
To investigate the role of autophagy in the anticancer effects of Tanshinone I in vitro, ATG7-siRNA and VPS34 inhibitor 3-MA were used to inhibit autophagy. BGC823 and SGC7901 cells were treated with Tanshinone I at 5 µg/ml in combination with ATG7-siRNA at 50 nM or 3-MA at 3 mM for 24 h, and then, cell viability was analyzed. As shown in Fig. 5a, b, cell growth inhibition was enhanced when Tanshinone I in combination with ATG7-siRNA or 3-MA. We also observed that ATG7-siRNA or 3-MA treatment downregulated Tanshinone I-induced LC3II conversion and unregulated the apoptosis-associated proteins cleaved caspase 3 (Fig. 5c, d). Furthermore, FACS result suggested that the combination treatment induced apoptosis more significantly than monotherapy (Fig.5e). Taken together, combining Tanshinone I with autophagy inhibitor enhanced cytotoxicity in vitro, and autophagy might be a pro-survival process against Tanshinone I-induced cancer cell growth inhibition.

Combination of Tanshinone I with autophagy inhibitor enhanced Tanshinone I-induced cytotoxicity in vitro. a Cell viability of with combining ATG7-siRNA with Tanshinone I at 5 μg/ml. b Cell viability of with combining a class III PI3 K (hVPS34) inhibitor 3-MA at 3 mM with Tanshinone I at 5 μg/ml. c Western blot assessing the change in autophagy- and apoptosis-related protein after ATG7-siRNA and Tanshinone I treatment. d Western blot assessing the change in autophagy- and apoptosis-related protein after 3-MA and Tanshinone I treatment. e FACS evaluated the effect of Tanshinone I, siRNA, or the combination on apoptosis. The data are expressed as the mean value ± SD. Statistical significance is indicated versus control (*P < 0.05; **P < 0.01; ***P < 0.001)
Combining Tanshinone I with autophagy inhibitor enhanced Tanshinone I-induced antitumor effect in vivo
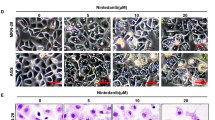
To examine the effect of autophagy inhibition on the in vivo anticancer efficacy of Tanshinone I, the nude mice bearing BGC823 tumor xenografts were treated by vehicle control, Tanshinone I at a concentration of 20 mg/kg q.d. or chloroquine at a concentration of 50 mg/kg q.d or the combination for 2 weeks p.o. The inhibitory ratios of Tanshinone I, chloroquine, and the combination on day 21 after first administration were 18.9, 57.5, and 70 %, respectively (Fig. 6a). Tumor weight was also measured at the end of the study (Fig. 6b), and the result was consistent with that of tumor volume. TUNEL staining demonstrated a significant increase in the number of apoptotic cells in the combination treatment group compared with the monotherapy groups (P < 0.01, Fig. 6c). Together, these data showed that the combination therapy of Tanshinone I and chloroquine had an addictive effect on reducing tumor proliferation and inducing tumor cell apoptosis than monotherapy in vivo.

Combination of Tanshinone I with autophagy inhibitor enhanced Tanshinone I-induced antitumor effect in vivo. a Growth curve of BGC823 xenografts upon treatment of vehicle or Tanshinone I at a concentration of 20 mg/kg q.d. or chloroquine at a concentration of 50 mg/kg q.d or the combination for 2 weeks p.o. Data are the mean tumor volumes ± SE. b Tumor volume of BGC823 xenografts upon treatment with control (vehicle), Tanshinone I, chloroquine, and the combination. c TUNEL showed the change in apoptosis in BGC823 xenografts upon treatment with control (vehicle), Tanshinone I, chloroquine, and the combination. The data are expressed as the mean value ± SD. d Western blot showed the changes in autophagy and apoptosis signaling events in BGC823 xenograft upon treatment with control (vehicle), Tanshinone I, chloroquine, and the combination. Statistical significance is indicated versus control (*P < 0.05; **P < 0.01; ***P < 0.001)
Discussion
Gastric cancer is the third most deadly cancer worldwide [1]. Due to its high metastatic recurrence, the prognosis of patients with advanced gastric cancer is poor [23]. At present, drugs are limited in clinical. Therefore, it is important to develop new effective therapy in gastric cancers. In this study, we found for the first time that Tanshinone I has potential anticancer activities in the human gastric cancer cells in vitro and in vivo, and exerts its anticancer effect by inducing apoptotic cell death.
Bcl-2, an anti-apoptosis and pro-survival protein, is overexpressed in multiple tumor types and contributes to drug resistance. Recent studies indicate that it can also inhibit autophagy via binding to Beclin-1, which was an autophagy-inducing protein [24]. Small-molecule antagonists of Bcl-2, ABT-737/263, can competitively disturb the Bcl-2/Beclin-1 complex to induce autophagy [24]. Our results suggest that the molecular mechanism underlining Tanshinone I induces autophagy and is likely to involve the downregulation of Bcl-2. We also found that overexpression of Bcl-2 could recover Tanshinone I-induced decreasing in Bcl-2/Beclin-1 complex. In this regard, our data demonstrate that Tanshinone I can induce autophagy in gastric cancer cells.
Recent studies indicate that anticancer drugs can trigger apoptosis and/or autophagy in cancer cells [25]. However, whether autophagy in response to anticancer drugs is pro-death or pro-survival is contentious. Tamoxifen and other antiestrogen agents can induce autophagy in breast cancer MCF-7 cells [26, 27]. This autophagy is mediated by a cell-permeable short-chain analog of a second messenger C2 ceramide and increased expression of Beclin-1, an autophagy-associated protein. Addition of 3-methyladenine (3-MA), an inhibitor of autophagosomes formation, prevents the cell death, suggesting that autophagy functions as a cell death program in this setting. It is interesting that bafilomycin A1, another autophagy inhibitor, radiosensitizes cancer cells by inducing apoptosis, suggesting that autophagy induced by radiation may be a protective mechanism [28]. In this regard, autophagy can be either pro-death or pro-survival depending upon the cellular context. In an effort to determine whether autophagy serves a pro-survival or pro-death role in response to Tanshinone I, we evaluated two genetic approaches to inhibit autophagy. Since autophagosome formation is regulated by a class III phosphoinositide-3-kinase (PI3 K) complex [24], we utilized the class III PI3 K inhibitor 3-methyladenine (3-MA) [29] and knocked down autophagy gene ATG7 by specific siRNAs. Data suggested that both 3-MA and ATG7-siRNA can significantly enhance apoptosis in combination with Tanshinone I. Both in vitro and in vivo assay indicate the consistent conclusion that Tanshinone I in combination with chloroquine, another autophagy inhibitor, suggested addictive efficacy in BGC823 tumor xenografts model.
In summary, this study demonstrates that Tanshinone I can induce apoptosis and pro-survival autophagy in vitro and in vivo via inhibiting Bcl-2 expression and increase the autophagy-inducing complex Beclin-1/VPS34 on gastric cancer. Autophagy induced by Tanshinone I serves a pro-survival role, and the combination of autophagy inhibitor chloroquine and Tanshinone I could increase cell apoptosis, which might be considered as an potential therapy for the treatment for gastric cancer.
References
Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D, Bray F (2015) Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer 136(5):E359–E386. doi:10.1002/ijc.29210
Fish JM, Welchons DR, Kim YS, Lee SH, Ho WK, Antzelevitch C (2006) Dimethyl lithospermate B, an extract of Danshen, suppresses arrhythmogenesis associated with the Brugada syndrome. Circulation 113(11):1393–1400. doi:10.1161/CIRCULATIONAHA.105.601690
Chang PN, Mao JC, Huang SH, Ning L, Wang ZJ, On T, Duan W, Zhu YZ (2006) Analysis of cardioprotective effects using purified Salvia miltiorrhiza extract on isolated rat hearts. J Pharmacol Sci 101(3):245–249
Zhou L, Zuo Z, Chow MS (2005) Danshen: an overview of its chemistry, pharmacology, pharmacokinetics, and clinical use. J Clin Pharmacol 45(12):1345–1359. doi:10.1177/0091270005282630
Chiu SC, Huang SY, Chang SF, Chen SP, Chen CC, Lin TH, Liu HH, Tsai TH, Lee SS, Pang CY, Hsieh TF (2014) Potential therapeutic roles of tanshinone IIA in human bladder cancer cells. Int J Mol Sci 15(9):15622–15637. doi:10.3390/ijms150915622
Hu T, Wang L, Zhang L, Lu L, Shen J, Chan RL, Li M, Wu WK, To KK, Cho CH (2015) Sensitivity of apoptosis-resistant colon cancer cells to tanshinones is mediated by autophagic cell death and p53-independent cytotoxicity. Phytomedicine 22(5):536–544. doi:10.1016/j.phymed.2015.03.010
Hou J, He J, Jin X, Hu T, Zhang Y (2013) Study on optimisation of extraction process of tanshinone IIA and its mechanism of induction of gastric cancer SGC7901 cell apoptosis. Afr J Tradit Complement Altern Med 10(6):456–458
Lin JY, Ke YM, Lai JS, Ho TF (2015) Tanshinone IIA enhances the effects of TRAIL by downregulating survivin in human ovarian carcinoma cells. Phytomedicine 22(10):929–938. doi:10.1016/j.phymed.2015.06.012
Gao H, Sun W, Zhao W, Hao W, Leung CH, Lu J, Chen X (2015) Total Tanshinones-induced apoptosis and autophagy via reactive oxygen species in lung cancer 95D cells. Am J Chin Med 43(6):1265–1279. doi:10.1142/S0192415X1550072X
Xu M, Cao FL, Li NY, Liu YQ, Li YP, Lv CL (2013) Tanshinone IIA reverses the malignant phenotype of SGC7901 gastric cancer cells. Asian Pac J Cancer Prev APJCP 14(1):173–177
Tseng PY, Lu WC, Hsieh MJ, Chien SY, Chen MK (2014) Tanshinone IIA induces apoptosis in human oral cancer KB cells through a mitochondria-dependent pathway. BioMed Res Int 2014:540516. doi:10.1155/2014/540516
Kim MK, Park GH, Eo HJ, Song HM, Lee JW, Kwon MJ, Koo JS, Jeong JB (2015) Tanshinone I induces cyclin D1 proteasomal degradation in an ERK1/2 dependent way in human colorectal cancer cells. Fitoterapia 101:162–168. doi:10.1016/j.fitote.2015.01.010
Su CC, Chen GW, Lin JG (2008) Growth inhibition and apoptosis induction by tanshinone I in human colon cancer Colo 205 cells. Int J Mol Med 22(5):613–618
Degenhardt K, Mathew R, Beaudoin B, Bray K, Anderson D, Chen G, Mukherjee C, Shi Y, Gelinas C, Fan Y, Nelson DA, Jin S, White E (2006) Autophagy promotes tumor cell survival and restricts necrosis, inflammation, and tumorigenesis. Cancer Cell 10(1):51–64. doi:10.1016/j.ccr.2006.06.001
Opipari AW Jr, Tan L, Boitano AE, Sorenson DR, Aurora A, Liu JR (2004) Resveratrol-induced autophagocytosis in ovarian cancer cells. Cancer Res 64(2):696–703
Ellington AA, Berhow MA, Singletary KW (2006) Inhibition of Akt signaling and enhanced ERK1/2 activity are involved in induction of macroautophagy by triterpenoid B-group soyasaponins in colon cancer cells. Carcinogenesis 27(2):298–306. doi:10.1093/carcin/bgi214
Hoyer-Hansen M, Bastholm L, Mathiasen IS, Elling F, Jaattela M (2005) Vitamin D analog EB1089 triggers dramatic lysosomal changes and Beclin 1-mediated autophagic cell death. Cell Death Differ 12(10):1297–1309. doi:10.1038/sj.cdd.4401651
Lu Z, Luo RZ, Lu Y, Zhang X, Yu Q, Khare S, Kondo S, Kondo Y, Yu Y, Mills GB, Liao WS, Bast RC Jr (2008) The tumor suppressor gene ARHI regulates autophagy and tumor dormancy in human ovarian cancer cells. J Clin Investig 118(12):3917–3929. doi:10.1172/JCI35512
Lum JJ, Bauer DE, Kong M, Harris MH, Li C, Lindsten T, Thompson CB (2005) Growth factor regulation of autophagy and cell survival in the absence of apoptosis. Cell 120(2):237–248. doi:10.1016/j.cell.2004.11.046
Tanida I, Ueno T, Kominami E (2008) LC3 and autophagy. Methods Mol Biol 445:77–88. doi:10.1007/978-1-59745-157-4_4
Pattingre S, Tassa A, Qu X, Garuti R, Liang XH, Mizushima N, Packer M, Schneider MD, Levine B (2005) Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell 122(6):927–939. doi:10.1016/j.cell.2005.07.002
Funderburk SF, Wang QJ, Yue Z (2010) The Beclin 1-VPS34 complex—at the crossroads of autophagy and beyond. Trends Cell Biol 20(6):355–362. doi:10.1016/j.tcb.2010.03.002
Cunningham D, Chua YJ (2007) East meets west in the treatment of gastric cancer. New Engl J Med 357(18):1863–1865. doi:10.1056/NEJMe078182
Levine B, Sinha S, Kroemer G (2008) Bcl-2 family members: dual regulators of apoptosis and autophagy. Autophagy 4(5):600–606
Amaravadi RK, Thompson CB (2007) The roles of therapy-induced autophagy and necrosis in cancer treatment. Clin Cancer Res 13(24):7271–7279. doi:10.1158/1078-0432.CCR-07-1595
Bursch W, Ellinger A, Kienzl H, Torok L, Pandey S, Sikorska M, Walker R, Hermann RS (1996) Active cell death induced by the anti-estrogens tamoxifen and ICI 164 384 in human mammary carcinoma cells (MCF-7) in culture: the role of autophagy. Carcinogenesis 17(8):1595–1607
Scarlatti F, Bauvy C, Ventruti A, Sala G, Cluzeaud F, Vandewalle A, Ghidoni R, Codogno P (2004) Ceramide-mediated macroautophagy involves inhibition of protein kinase B and up-regulation of beclin 1. J Biol Chem 279(18):18384–18391. doi:10.1074/jbc.M313561200
Daido S, Yamamoto A, Fujiwara K, Sawaya R, Kondo S, Kondo Y (2005) Inhibition of the DNA-dependent protein kinase catalytic subunit radiosensitizes malignant glioma cells by inducing autophagy. Cancer Res 65(10):4368–4375. doi:10.1158/0008-5472.CAN-04-4202
Seglen PO, Gordon PB (1982) 3-Methyladenine: specific inhibitor of autophagic/lysosomal protein degradation in isolated rat hepatocytes. Proc Natl Acad Sci USA 79(6):1889–1892
Acknowledgments
This work was supported by Shanghai Natural Science Foundation of China (13ZR1434300). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors have declared that no competing interests exist.
Rights and permissions
About this article

Cite this article
Jing, X., Xu, Y., Cheng, W. et al. Tanshinone I induces apoptosis and pro-survival autophagy in gastric cancers. Cancer Chemother Pharmacol 77, 1171–1181 (2016). https://doi.org/10.1007/s00280-016-3034-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00280-016-3034-6