
Abstract
Managing biological invasions requires rapid, cost-effective assessments of introduced species’ occurrence, and a good understanding of the species’ vegetation associations. This is particularly true for species that are elusive or may spread rapidly. Finlayson’s squirrel (Callosciurus finlaysonii) is native to Thailand and southeastern Asia, and two introduced populations occur in peninsular Italy. One of the two introduced populations is rapidly expanding, but neither effective monitoring protocols nor reliable information on vegetation associations are available. To fill this gap, we conducted visual surveys and hair tube sampling in a periurban landscape of southern Italy to compare the effectiveness of these two methods in assessing presence of Finlayson’s squirrel. We also determined the species’ association with vegetation types at detection locations and nesting sites. Both visual and hair tube sampling effectively assessed the species’ presence, but hair tubes resulted in fewer false absences. Moreover, when we controlled for the costs of labor and equipment, hair tubes were 33.1% less expensive than visual sampling. Presence of squirrels and their nests was positively correlated with shrub species richness, indicating that the occurrence of forests with well-developed understory may inhibit the spread of the species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Countering the expansion of an introduced animal species, and minizing its undesirable effects, requires good knowledge of its distribution and the identification of the areas and vegetation types where expansion may occur (e.g., Ballari et al. 2016). Such information is fundamental to plan eradication or control actions and improve their effectiveness (Braysher 1993; Bertolino et al. 2005). Monitoring newly established populations of introduced species and assessing their vegetation associations is therefore necessary to better understand their spatial dynamics and inform effective management strategies.
Monitoring low-density, elusive, or nocturnal species is challenging (Mills et al. 2000; Thompson 2004; Kindberg et al. 2009; Srivathsa et al. 2014) because false absences may be common, i.e., the species often may not be detected despite being present (Gu and Swihart 2004; Mortelliti and Boitani 2007, 2008). False absences may limit the effectiveness of management actions, which might overlook sites where the species inaccurately is deemed to be absent (Gu and Swihart 2004). For mammals, indirect methods such as detection of footprints (Reynolds et al. 2004; Yarnell et al. 2014) or hair (Gurnell et al. 2004a; Harris et al. 2006) may complement or replace direct observation when the latter is ineffective, for instance when the species avoids areas in which humans are present and possibly is elusive. Moreover, monitoring or countering the spread of introduced species greatly depends on the availability of information on their habitat, including nesting or roosting sites (Palmer et al. 2013). For example, reproduction and wintering of introduced squirrels relies on the presence of nesting sites (dreys) (Setoguchi 1991; Okubo et al. 2005; Palmer et al. 2013).
Finlayson’s squirrel (Callosciurus finlaysonii) is an arboreal rodent native to Southeast Asia (Thorington et al. 2012). The species are ecologically adaptable, and, in its native and introduced ranges, may occur in several forest types. The species occurs in forests that are logged commercially, where it feeds opportunistically and seasonally on fruits, seeds, buds, and occasionally small animals (Lekagul and McNeely 1977; Bertolino et al. 2004). A few populations of this squirrel, introduced through the pet trade (Bertolino and Lurz 2013), have established outside its native range in the last 35-40 years. Two introduced populations are present in Singapore and Japan (Oshida et al. 2007). Two other introduced populations occur in Italy; a small population in the north (Mazzoglio et al. 2007) and a large population in the south resulted from the release, in the 1980s, of a few individuals in an urban park (Aloise and Bertolino 2005). The effects of this species in its introduced range include tree bark stripping, consumption of fruits and seeds in crops, and damage to electric cables (Bertolino et al. 2015; Mori et al. 2016). As in its native range, in its introduced range C. finlaysonii is associated with forests, where it builds nests from plants (Bertolino et al. 2004), but no detailed description of habitat is available. Although the population of C. finlaysonii in southern Italy has expanded along the Tyrrhenian coast, both naturally and through further human-assisted introductions (Aloise and Bertolino 2005), monitoring has been practically nonexistent (Bertolino et al. 2015).
Effective monitoring requires designing appropriate surveys. Given that C. finlaysonii is arboreal and associated with forests, in which direct visual observation may be difficult, indirect detection may be preferred over direct observation. To date, however, no study has assessed the effectiveness of alternative survey methods. Moreover, there is no comprehensive understanding of vegetation associations of C. finlaysonii, which would help focus survey efforts and inform future management, including population control or eradication.
We aimed to help fill these gaps. First, we determined whether C. finlaysonii were present by applying two detection methods, visual sampling (VS) and hair tube sampling (HTS), both of which are used to survey European red squirrels Sciurus vulgaris (Mortelliti and Boitani 2008). We compared the effectiveness and cost of these methods. Then, we assessed vegetation associations of C. finlaysonii at two levels: forest patches and nesting sites.
Methods
Patterns of Patch Occupancy and Comparison of Detection Methods
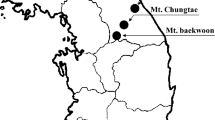
We used both VS and HTS to survey 22 forest patches (mean size ± SD: 4.3 ± 0.9 ha, range 0.6–18.1 ha) within a periurban landscape in the municipalities of Sapri (40.07°N, 15.64°E) and Palinuro (40.03°N, 15.28°E) on the Tyrrhenian coast of Campania (southern Italy; Fig. 1), where Finlayson’s squirrels were first recorded in 2004 (Aloise and Bertolino 2005). Forest patches varied in size, isolation, and species composition (Table 1), and were surrounded by an agricultural matrix of olive groves, cereal fields, and private vegetable gardens and by urban settlements.

Study region (Campania, shaded area) and sampling sites (inset; shaded areas in circles)
Hair tubes were commercially available PVC pipe segments (length 30 cm, diameter ~7 cm), at each end of which a small piece of plastic sheet covered with sticky tape was fixed to the roof (Gurnell et al. 2004a). We baited hair tubes with hazelnut cream and sunflower seeds and secured tubes horizontally on tree trunks and branches 1.5–2.0 m above ground. We established one transect per forest patch, each consisting of a linear sequence of hair tubes ca. 100 m apart. The number of hair tubes in each transect increased as patch size increased, and ranged from 2 to 12 (Mortelliti and Boitani 2008); for forest patches <1.5 ha, we positioned only one tube. We inspected hair tubes on 8 occasions separated by 10 days (80 days total). During each inspection, we replaced adhesive tape and stored trapped hairs for subsequent laboratory identification. Hairs were slide-mounted and identified with an Olympus BX 51 optic microscope equipped with 4–100× lenses. We examined hair morphology following Teerink (1991) and Venturini et al. (2008), and used a reference collection. We used this approach to distinguish C. finlaysonii hairs from those of other rodents that might visit hair tubes (Glis glis [edible dormouse], Muscardinus avellanarius [hazel dormouse], and Rattus rattus [black rat]). No other squirrel species occurred in the area.
For VS, one operator walked along a given transect, stopping 5 min near each tube and recording all squirrels sighted (Gurnell et al. 2004a). We therefore recorded squirrels on the basis of point counts, without accounting for their distance from the observer. VS also was conducted every 10 days for a total of 8 visits.
For each patch, we compiled the detection history (non-detection, 0; detection, 1) for HTS and VS separately. We then ran single-season occupancy models (Mortelliti and Boitani 2008), including a set of environmental covariates (Table 1) likely to be associated with patterns of occupancy in arboreal squirrels in a patchy landscape (Mortelliti and Boitani 2008). We expected the probability of presence in a given patch (Ψ: psi) to be associated with patch size, degree of isolation as measured by either the number of patches within 500 m of the patch edge or the distance between the focal patch and the closest neighboring patch, and vegetation structure. We selected the 500 m as the maximum distance that other squirrel species are known to move across a forest gap, representing a proxy for dispersal ability in a matrix of non-habitat (Bakker and Van Vuren 2004). We classified forest composition as mixed oaks, monospecific cork oak, or mixed (i.e., coniferous and broadleaved) forest. We measured shrub species richness as the number of shrub species recorded along each 10 m transect or, in two patches <1.5 ha large where only one hair tube was present, in a 10 m circular area around the tube. Because introduced C. finlaysonii often occur close to urban areas (Aloise and Bertolino 2005), we also included the minimum distance from the closest urban settlement as a covariate. We modeled detection probability (p) as a function of patch area, shrub species richness, and forest composition to account for potential biases in squirrel detection resulting from differences in the area within which transects were embedded or in station-dependent hair tube attractiveness or visibility.
We ran analyses with the R package Unmarked (Fiske and Chandler 2011). We evaluated sample size, i.e., the minimum number of visits necessary to assess whether the species was present (Reed 1996), as N = ln(αlevel)/ln(1−p), where α represents the probability of type I error (fixed at 0.05). We ranked models according to Akaike Information Criterion (AIC) values (Burnham and Anderson 2002). Because different models may provide similar results, we considered as valid all models with ΔAIC <2 from the model with the lowest AIC (Burnham and Anderson 2002). Among valid models, we assumed that the best supported were those with the lowest AIC values (MacKenzie et al. 2006). We calculated Akaike weights (wi) to assess the strength of evidence in support of each model (Srivathsa et al. 2014). We also used a stepwise process, implemented with the lme4 R package (Bates et al. 2015; R Core Team 2014), to generate a global model of Ψ and identify the covariates that were significantly associated with occupancy.
We also estimated the cost of implementing each survey method for the entire study period and per survey. Costs included materials (hair tubes, bait) and human effort (the time needed to assemble and place hair tubes and to commute between tubes and between sites, and time spent in the field and in the laboratory, at a standard rate of 10.00 €/h). We calculated total cost by multiplying cost per visit by the minimum number of visits needed to assess occupancy. We assessed cost per detection by dividing the total cost by the number of detections with the VS and HTS methods.
Selection of Nesting Areas
C. finlaysonii builds conspicuous nests of leaves and twigs on tree trunks, branches, or in the foliage (Setoguchi 1991; Okubo et al. 2005). We located squirrel nests by inspecting forest patches along all accessible trails separated by ca. 50 m, following the approach adopted by Palmer et al. (2013) for red-bellied squirrels Sciurus aureogaster introduced to Florida, and recorded their position with a Dakota 10—Garmin GPS receiver.
We measured nest site characteristics at three levels: nest, nesting tree, and plot. At the nest level, we measured height above ground and nest exposure (measured with a compass as the orientation of the nest relative to its support, which we classified as one of the eight main cardinal directions). The position and exposure of the nest may affect the timing and duration of solar radiation to the nest, influencing microclimate within the nest. We explored differences in nest aspect with a Rayleigh test, implemented in the CircStat R package (Agostinelli 2009).
For each tree in which a squirrel nest was built, we recorded tree species, tree height and diameter at breast height (DBH), whether vines were present on the trunk, percent canopy closure (assessed visually following Paletto and Tosi 2009), number of crown linkages (i.e., number of neighboring trees whose branches were ≤0.5 m from the occupied tree), location in the patch (core or edge of patch or clearing), and distance from the closest tree. We also generated 50 random coordinates within the study area with a random number grid on 1:10,000 maps. We used the tree closest to each random location as a control tree, at which we took the same measurements that we recorded for nesting trees. At the random location, we chose the closest tree with DBH >10 cm, the lowest value in the nesting trees data. To establish which trees are used for nesting and which features are associated with those trees, we compared the features of used trees with those of unused (random) trees. We used logistic regressions, including both categorical variables (tree species, whether the tree was live or dead, presence of vines, tree position within the forest patch) and normally distributed, continuous variables (tree size, canopy closure, and number of linkages).
At the plot level, we collected vegetation data within 10 m of nesting and random trees (Palmer et al. 2013). We recorded shrub species richness and abundance, tree species richness, and densities of live and dead trees (Pignatti 1982). We explored differences in vegetation between plots in which the species nested and random plots by running generalized linear models. We evaluated the direction and magnitude of effects by inspecting parameter estimates (β).
We ran all analyses with R 3.2.1 and set significance at p < 0.05. We present all results as mean ± SE with associated 95% confidence intervals (CI).
Results
Patterns of Patch Occupancy and Comparison of Detection Methods
We detected C. finlaysonii in 19 of 22 forest patches (86.4%). The proportion of occupied sites estimated by HTS was closer to the observed proportion than that estimated by VS (HTS: range: 83–87%, CI: 0.84–0.85; VS: 70–83%, CI: 0.72–0.80). VS and HTS resulted in 17 and 4% false absences, respectively. Detection probability (p) estimated with either method was similar (range 0.45–0.58), although it was significantly higher for HTS (CI: 0.56–0.58) than for VS (CI: 48.4–53.6; see Table 2). For both methods, the minimum number of visits per site necessary to establish presence was 5. No covariates were significantly associated with detection probability. In the global model, Ψ was positively correlated with shrub species richness and, to a lesser extent, negatively correlated with the distance to the closest patch; patch area and forest type were associated with occupancy as estimated by VS and HTS, respectively (Table 2).
The two methods had similar costs (Table 3), 2984€ and 2585€ for HTS and VS, respectively (Table 3), but after correcting for the number of surveys, HTS was 33.1% less expensive than VS (58.69€ vs. 87.67€ per detection). We provide a full list of the models we evaluated as Supplementary Material (Tables S1 and S2).
The models that minimized information loss (lowest AICc) and maximized explained proportion of variance (R2 values) (Table 4) indicated that occupancy was a function of shrub species richness when estimated with HTS, and a function of shrub species richness and distance from other forest patches when measured with VS.
Occupancy was positively associated with shrub species richness (0.24 and 0.47 for VS and HTS, respectively) and negatively associated with distance from other patches (−0.13 and −0.11; Table 2). VS suggested that the likelihood of squirrel occupancy significantly increased near human settlements (−0.67; Table 3). Forest type also was associated with occupancy by C. finlaysonii (0.688 and 0.083 for HTS and VS, respectively), with lowest values in conifer and cork oak stands, and highest values in mixed and mixed oak patches.
Selection of Nesting Areas
The 47 nests were located at heights of 7.7 ± 0.4 m above ground (range: 2.0–20.5 m), often (48.9%) between 5 and 6 m. Nests were located in all patches that VS and HTS indicated were occupied. Nests faced all directions, but most often southeast (Rayleigh’s Z = 123.81, p < 0.001, r = 0.32). Most (n = 33) nests were located in oaks (Quercus spp.). The remainder were located in strawberry trees (Arbutus unedo) (n = 7), olive trees (Olea europaea) (n = 3), Spanish broom (Spartium junceum) (n = 2), and wild plum trees (Prunus spp.) (n = 2). Nests occurred most often in downy oaks (Quercus pubescens) (β = 1.43, p < 0.05) and least often in cork oaks (Q. suber) (β = −2.46, p < 0.05).
The distance between random trees and closest nesting trees was 72–358 m. The size of nesting trees did not differ from that of random trees (height: β = 1.11, p = 0.11, CI: −1.59–5.0; DBH: β = 0.91, p = 0.061, CI: −1.64–1.96). Presence of vines on the trunk (β = −0.15, p = 0.54, CI: −1.11–3.12), tree position within the patch (β = 0.35, p = 0.91, CI: −2.12–3.21), canopy closure (β = −0.55, p = 0.78, CI: −9.9–15.4), and distance from the closest tree (β = 0.29, p = 0.13, CI: −1.21–3.39) did not vary between nest trees and random trees. Nesting trees had fewer crown linkages than random trees (nest trees: 3.6 ± 0.40; random trees: 5.3 ± 0.1; β = −3.19, p < 0.05, CI: −5.12 to −1.09). Plots around nesting trees had a lower abundance of shrubs (nesting plots: 20.70 ± 1.01; random plots: 35.44 ± 1.51; β = −2.78, p < 0.01, CI: −8.12 to −0.91) but greater species richness (nesting plots: 8.54 ± 2.38; random plots: 3.26 ± 1.20; β = 4.31, p < 0.01, CI: 1.12–6.33) than plots around random trees. Neither tree species richness (β = 1.91, p = 0.34; CI: −0.91–1.43) nor tree density (β = 0.36, p = 0.71, CI: −0.12–0.61) was significantly different between nesting and random plots.
Discussion
We found that HTS is more effective and costs less than VS to detect C. finlaysonii (Cagnacci et al. 2012). Our cost analysis did not take into account the fact that plastic hair tubes are robust enough to be reused for several surveys, further reducing costs. The proportion of sites in which we detected the species by HTS was ~4% lower than the estimated occupancy. Our results suggested that in forests, a minimum of five visits, 10 days apart, will provide reliable results. Although we conducted eight hair trapping surveys, we did not detect squirrels in additional sites after the fifth visit. The two methods differ markedly in sampling time. Whereas hair tubes typically are deployed over weeks or months, VS is conducted over short durations. VS may be affected by squirrels’ antipredatory responses, which include escaping or hiding from the observer (Haigh et al. 2017). When threatened, C. finlaysonii and congeners (Tamura and Yong 1993) emit loud alarm calls that increase detection probability (personal observation).
The positive association between occurrence and shrub species richness at both tree and plot levels might reflect the species’ opportunistic foraging on a range of plant parts, particularly seeds and fruits that are available on shrubs year round (Bertolino et al. 2004). The dependence of C. finlaysonii on large and well-connected forests, which is typical of tree squirrels (Celada et al. 1994; Rodríguez and Andrén 1999; Koprowski 2005), may explain the negative association between occupancy and distance to neighboring forest patches. The presence of C. finlaysonii, like that of other adaptable species often introduced to urban areas by the pet trade, is positively associated with proximity to human settlements (Jodice and Humphrey 1992; McCleery et al. 2007; Bonnington et al. 2014; Greene and McCleery 2017).
Although the diet of C. finlaysonii mainly consists of ripe fruits (Bertolino et al. 2004), in the study area these were available only for part of the year. Therefore, nesting on Q. pubescens may be a strategy to facilitate harvesting of acorns, providing squirrels with an additional food source, when other food items such as berries are unavailable. Southeast-facing dreys may facilitate exposure to the sun in the early morning while reducing exposure to cold wind from the north, thus providing a warm microclimate to nestlings. This may also explain the low heights of most nests. Most medium-sized tree squirrels (i.e., 100–500 g) build their nests at a similar height above ground (ca. 5 m) to shelter the nest from strong wind while precluding access to terrestrial predators or competitors that might steal the food stored in the nest (Rothwell 1979; Alvarenga and Talamoni 2005; Thorington et al. 2012). Given the tropical-subtropical origin of these species (Lekagul and McNeely 1977; Thorington et al. 2012), warm nesting sites also may be an adaptation to the cold spells in temperate regions (Terrien et al. 2011).
Our work highlights that hair tubes may be an effective means of surveying C. finlaysonii in dense vegetation or where the species may occur at low densities, e.g., where it might have colonized recently. We found that C. finlaysonii often occurs and builds nests in forest patches in which the species richness of the understory is relatively high. Such patches may be high priorities for monitoring and for management actions, such as control or eradication.
Introduced squirrels have had considerable effects in European ecosystems (Bertolino 2009; Bertolino and Lurz 2013). For example, native Sciurus vulgaris (Eurasian red squirrels) have been driven to extinction in much of their range within the United Kingdom by Sciurus carolinensis (Eastern gray squirrels) (Gurnell et al. 2004b) and, in Italy, their survival is jeopardized by both S. carolinensis (Bertolino et al. 2014b) and Callosciurus erythraeus (Pallas’ squirrels) (Mazzamuto et al. 2016). C. finlaysonii removes bark from trees, causing damage to forest and orchards (Mori et al. 2016; Bertolino et al. 2015), and may have negative effects on other species. The effects of introduced squirrels on other species and human activities may make active monitoring of their range useful for predicting and informing management of their expansion at larger extents (Di Febbraro et al. 2016; White et al. 2016). C. finlaysonii is believed to be expanding south, where it may compete with the newly described Sciurus meridionalis (Calabrian black squirrel) (Bertolino et al. 2014a), an endemic species occupying a small area (Wauters et al. 2017). We therefore urge that monitoring of the southern Italian population of C. finlaysonii be carried out to inform the adoption of effective control measures.
References
Agostinelli C (2009) CircStats: circular statistics, from “Topics in Circular Statistics” (2001). R package version 0.2-4. http://CRAN.R-project.org/package=CircStats
Aloise G, Bertolino S (2005) Free-ranging population of the Finlayson’s squirrel Callosciurus finlaysonii (Horsfield, 1824) (Rodentia, Sciuridae) in South Italy. Hystrix 16:70–74
Alvarenga CA, Talamoni SA (2005) Nests of the Brazilian squirrel Sciurus ingrami Thomas (Rodentia, Sciuridae). Rev Bras Zool 22:816–818
Bakker VJ, Van Vuren DH (2004) Gap‐crossing decisions by the red squirrel, a forest‐dependent small mammal. Conserv Biol 18:689–697
Ballari SA, Anderson CB, Valenzuela AEJ (2016) Understanding trends in biological invasions by introduced mammals in southern South America: a review of research and management. Mammal Rev 46:229–240
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Soft 67:1–48
Bertolino S (2009) Animal trade and non-indigenous species introduction: the world-wide spread of squirrels. Divers Distrib 15:701–708. https://doi.org/10.1111/j.1472-4642.2009.00574.x
Bertolino S, Lurz PW (2013) Callosciurus squirrels: worldwide introductions, ecological impacts and recommendations to prevent the establishment of new invasive populations. Mammal Rev 43:22–33. https://doi.org/10.1111/j.1365-2907.2011.00204.x
Bertolino S, Mazzoglio PJ, Vaiana M, Currado I (2004) Activity budget and foraging behavior of introduced Callosciurus finlaysonii (Rodentia, Sciuridae) in Italy. J Mammal 85:254–259. https://doi.org/10.1644/BPR-009
Bertolino S, Perrone A, Gola L (2005) Effectiveness of coypu control in small Italian wetland areas. Wildl Soc Bull 33:714–720. https://doi.org/10.2193/0091-7648. (2005)33[714:EOCCIS]2.0.CO;2
Bertolino S, Girardello M, Amori G (2014a) Identifying conservation priorities when data are scanty: a case study with small mammals in Italy. Mamm Biol 79:349–356. https://doi.org/10.1016/j.mambio.2014.06.006
Bertolino S, Colangelo P, Mori E, Capizzi D (2015) Good for management, not for conservation: an overview of research, conservation and management of Italian small mammals. Hystrix 26:25–35. https://doi.org/10.4404/hystrix-26.1-10263
Bertolino S, Cordero di Montezemolo N, Preatoni DG, Wauters LA, Martinoli A (2014b) A grey future for Europe: Sciurus carolinensis is replacing native red squirrels in Italy. Biol Invasions 16:53–62. https://doi.org/10.1007/s10530-013-0502-3
Bonnington C, Gaston KJ, Evans KL (2014) Squirrels in suburbia: influence of urbanisation on the occurrence and distribution of a common exotic mammal. Urban Ecosyst 17:533–546. https://doi.org/10.1007/s11252-013-0331-2
Braysher ML (1993) Managing vertebrate pests: principles and strategies. Australian Government Publishing Service, Canberra
Burnham KP, Anderson DR (2002) Model selection and inference—a practical information-theoretic approach, 2nd edn. Springer, New York
Cagnacci F, Cardini A, Ciucci P, Ferrari N, Mortelliti A, Preatoni DG, Russo D, Scandura M, Wauters LA, Amori G (2012) Less is more: a researcher’s survival guide in time of economic crisis. Hystrix 23:1–7. https://doi.org/10.4404/hystrix-23.2-8737
Celada C, Bogliani G, Gariboldi A, Maracci A (1994) Occupancy of isolated woodlots by the red squirrel Sciurus vulgaris L. in Italy. Biol Cons 69:177–183. https://doi.org/10.1016/0006-3207(94)90057-4
Di Febbraro M, Martinoli A, Russo D, Preatoni D, Bertolino S (2016) Modelling the effects of climate change on the risk of invasion by alien squirrels. Hystrix 27:1–8. https://doi.org/10.4404/hystrix-27.1-11776
Fiske I, Chandler R (2011) Unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. J Stat Soft 43:1–23
Greene DU, McCleery RA (2017) Multi-scale responses of fox squirrels to land-use changes in Florida: utilization mimics historic pine savannas. Ecol Manag 391:42–51. https://doi.org/10.1016/j.foreco.2017.02.001
Gu W, Swihart RK (2004) Absent or undetected? Effects of non-detection of species occurrence on wildlife–habitat models. Biol Cons 116:195–203. https://doi.org/10.1016/S0006-3207(03)00190-3
Gurnell J, Lurz PWW, Shirley MDF, Cartmel S, Garson PJ, Magris L, Steele J (2004a) Monitoring red squirrels Sciurus vulgaris and grey squirrels Sciurus carolinensis in Britain. Mammal Rev 34:51–74. https://doi.org/10.1046/j.0305-1838.2003.00028.x
Gurnell J, Wauters LA, Lurz PW, Tosi G (2004b) Alien species and interspecific competition: effects of introduced eastern grey squirrels on red squirrel population dynamics. J Anim Ecol 73:26–35. https://doi.org/10.1111/j.1365-2656.2004.00791.x
Haigh A, Butler F, O’Riordan R, Palme R (2017) Managed parks as a refuge for the threatened red squirrel (Sciurus vulgaris) in light of human disturbance. Biol Cons 211:29–36
Harris DB, Gregory SD, Macdonald DW (2006) Space invaders? A search for patterns underlying the coexistence of alien black rats and Galápagos rice rats. Oecologia 149:276–288. https://doi.org/10.1007/s00442-006-0447-7
Jodice PG, Humphrey SR (1992) Activity and diet of an urban population of Big Cypress fox squirrels. J Wildl Manag 54:685–692. https://doi.org/10.2307/3809461
Kindberg J, Ericsson G, Swenson JE (2009) Monitoring rare or elusive large mammals using effort-corrected voluntary observers. Biol Cons 142:159–165. https://doi.org/10.1016/j.biocon.2008.10.009
Koprowski JL (2005) The response of tree squirrels to fragmentation: a review and synthesis. Anim Cons 8:369–376. https://doi.org/10.1017/S1367943005002416
Lekagul B, McNeely JA (1977) Mammals of Thailand. Association for the Conservation of Wildlife, Bangkok
MacKenzie DI, Nichols JD, Royle JA, Pollock KH, Bailey LL, Hines JE (2006) Occupancy estimation and modeling. Inferring patterns and dynamics of species occurrence. Elsevier Academic Press Editions, Burlington, MA
Mazzamuto MV, Bisi F, Wauters LA, Preatoni DG, Martinoli A (2016) Interspecific competition between alien Pallas’s squirrels and Eurasian red squirrels reduces density of the native species. Biol Inv 19:723–735. https://doi.org/10.1007/s10530-016-1310-3
Mazzoglio PJ, Blengio L, Bertolino S (2007) Individual recognition in Callosciurus finlaysonii (Horsfield, 1824) (Rodentia, Sciuridae) introduced into Italy. Mammalia 71:122–125. https://doi.org/10.1515/MAMM.2007.018
McCleery RA, Lopez RR, Silvy NJ, Kahlick SN (2007) Habitat use of fox squirrels in an urban environment. J Wildl Manag 71:1149–1157. https://doi.org/10.2193/2006-282
Mills LS, Citta JJ, Lair KP, Schwartz MK, Tallmond DA (2000) Estimating animal abundance using noninvasive DNA sampling: promise and pitfalls. Ecol Appl 10:283–294. https://doi.org/10.1890/1051-0761(2000)010[0283:EAAUND]2.0.CO;2
Mori E, Mazzoglio PJ, Rima PC, Aloise G, Bertolino S (2016) Bark-stripping damage by Callosciurus finlaysonii introduced into Italy. Mammalia 80:507–514. https://doi.org/10.1515/mammalia-2015-0107
Mortelliti A, Boitani L (2007) Estimating species’ absence, colonization and local extinction in patchy landscapes: an application of occupancy models with rodents. J Zool 273:244–248. https://doi.org/10.1111/j.1469-7998.2007.00320.x
Mortelliti A, Boitani L (2008) Inferring red squirrel (Sciurus vulgaris) absence with hair tubes surveys: a sampling protocol. Eur J Wildl Res 54:353–356. https://doi.org/10.1007/s10344-007-0135-x
Okubo M, Tamura N, Katsuki T (2005) Nest site and materials of the alien squirrel, Callosciurus erythraeus. Kanagawa prefecture. J Jpn Wildl Res Soc 31:5–10
Oshida T, Torii H, Lin LK, Lee JK, Chen YJ, Endo H, Sasaki M (2007) A preliminary study on origin of Callosciurus squirrels introduced into Japan. Mamm Study 32:75–82. https://doi.org/10.3106/1348-6160(2007)32[75:APSOOO]2.0.CO;2
Paletto A, Tosi V (2009) Forest canopy cover and canopy closure: comparison of assessment techniques. Eur J For Res 128:265–272
Palmer GH, Koprowski JL, Pernas AJ (2013) Nest tree and site selection of an introduced population of red-bellied squirrels (Sciurus aureogaster). J Mammal 94:1274–1281. https://doi.org/10.1644/12-MAMM-A-308.1
Pignatti S (1982) Flora d’Italia. Edagricole, Bologna
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Reed JM (1996) Using statistical probability to increase confidence of inferring species extinction. Cons Biol 10:1283–1285
Reynolds JC, Short MJ, Leigh RJ (2004) Development of population control strategies for mink Mustela vison, using floating rafts as monitors and trap sites. Biol Cons 120:533–543. https://doi.org/10.1016/j.biocon.2004.03.026
Rodríguez A, Andrén H (1999) A comparison of Eurasian red squirrel distribution in different fragmented landscapes. J Appl Ecol 36:649–662. https://doi.org/10.1046/j.1365-2664.1999.00426.x
Rothwell R (1979) Nest sites of red squirrels (Tamiasciurus hudsonicus) in the Laramie Range of southeastern Wyoming. J Mammal 60:404–405. https://doi.org/10.2307/1379815
Setoguchi M (1991) Nest-site selection and nest-building behaviour of red-bellied tree squirrels on Tomogashima Island, Japan. J Mammal 72:163–170. https://doi.org/10.2307/1381991
Srivathsa A, Karanth KK, Jathanna D, Kumar NS, Karanth KU (2014) On a dhole trail: examining ecological and anthropogenic correlates of dhole habitat occupancy in the Western Ghats of India. PLoS ONE 9:e98803. https://doi.org/10.1371/journal.pone.0098803
Tamura N, Yong HS (1993) Vocalizations in response to predators in three species of Malaysian Callosciurus (Sciuridae). J Mammal 74:703–714
Teerink BJ (1991) Hair of West-European mammals. Cambridge University Press, Cambridge
Terrien J, Perret M, Aujard F (2011) Behavioral thermoregulation in mammals: a review. Front Biosci 16:1428–1444
Thompson W (2004) Sampling rare or elusive species: concepts, designs, and techniques for estimating population parameters. Island Press, Washington, DC
Thorington RW, Koprowski JL, Steele MA, Whatton JF (2012) Squirrels of the world. John Hopkins University Press, Baltimore, MD
Venturini M, Garrone A, Fasce E, Bertolino S, Balduzzi A (2008) L’uso di hair-tubes per il monitoraggio dello scoiattolo di Finlayson Callosciurus finlaysonii (Horsfield, 1923). Atti del 3° Workshop “Cantieri della Biodiversità”, 11–12 Settembre. Palazzo Squarcialupi, Siena, p 48
Wauters LA, Amori G, Aloise G, Gippoliti S, Agnelli P, Galimberti A, Casiraghi M, Preatoni D, Martinoli A (2017) New endemic mammal species for Europe: Sciurus meridionalis (Rodentia, Sciuridae). Hystrix 28:1–8. https://doi.org/10.4404/hystrix-28.1-12015
White A, Lurz PWW, Bryce J, Tonkin M, Ramoo K, Bamforth L, Jarrott A, Boots M (2016) Modelling disease spread in real landscapes: squirrelpox spread in Southern Scotland as a case study. Hystrix. https://doi.org/10.4404/hystrix-27.1-11657
Yarnell RW, Pacheco M, Williams B, Neumann JL, Rymer DJ, Baker PJ (2014) Using occupancy analysis to validate the use of footprint tunnels as a method for monitoring the hedgehog Erinaceus europaeus. Mammal Rev 44:234–238. https://doi.org/10.1111/mam.12026
Acknowledgements
We thank Alessia D’Auria for assisting with laboratory procedures. This project did not receive any specific funding. Thanks also go to the associate editor who handled our paper and two anonymous reviewers whose recommendations helped us to improve our manuscript greatly.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Ethical Approval
All procedures performed in studies involving wild species were in accordance with the ethical standards of the Institutional or National Research Committee and with approved ethical standards.
Electronic supplementary material
Rights and permissions
About this article

Cite this article
Ancillotto, L., Notomista, T., Mori, E. et al. Assessment of Detection Methods and Vegetation Associations for Introduced Finlayson’s Squirrels (Callosciurus finlaysonii) in Italy. Environmental Management 61, 875–883 (2018). https://doi.org/10.1007/s00267-018-1013-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-018-1013-x