
Abstract
Viviparity has evolved from oviparity approximately 142 times among vertebrates. Different theories have been proposed to explain the evolution of each of its traits in the different taxa. None, however, is applicable to all the viviparous vertebrates, since the derived ecological advantages such as controlling incubating temperature or protecting eggs against predation differ amongst clades. Most theories have been developed under a co-adaptive perspective, whereas less attention has been paid to conflict. We developed a broad panorama of the gradual evolution, from oviparity to advanced forms of viviparity, that includes the different environmental and co-adaptive selective pressures that have been suggested to be at the root of the different instances of viviparity and of the diverse maternal–foetal adaptations for nutrient transfer seen amongst vertebrates. Furthermore, we highlight the importance of conflict as a crucial driver of the evolution of many of those traits, including the evolution of epigenetic control of maternal resources. We suggest that the different types of matrotrophic viviparity, and probably also some reversals to oviparity, have been the result of an antagonistic coevolution between mothers, fathers and offspring, and their genomes. We additionally suggest that the appearance of a trait that allowed or favoured the evolution of internal development and matrotrophy generates a new selective environment that promotes further adaptations or counteradaptations, leading to the observed diversity of forms of embryonic development, nourishment, and transfer of maternal nutrients, and ultimately to the diversity of extant viviparous taxa.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Even in its simplest forms, reproduction is costly, since either self-dividing or producing and releasing gametes use resources (Stearns 1989; Blacher et al. 2017; Tarwater and Arcese 2017). These costs are referred to as reproductive investment because they are incurred in order to gain fitness. Investment in individual offspring can be made in one step, as when females produce yolk-provisioned eggs, or may be deferred through a gestation period via embryo-maternal (or paternal) interactions, and parental investment frequently continues after hatching or birth. However, although producing one descendant may convey the same benefit to both parents, the net fitness gain by each parent may differ if their respective parental contributions are not equal (Gross and Sargent 1985). For instance, withholding parental investment can be advantageous if the partner can provide enough resources for the offspring to reach independence, because the same fitness will be accrued with less investment. This is a type of sexual conflict, which more generally occurs when the evolutionary interest of males and females differ, or when their optima cannot simultaneously be realised (Trivers 1972). Sexual conflict can arise in relation to courtship, current or future mating decisions (Parker 1974; Smuts and Smuts 1993; Arnqvist and Rowe 2002; Magurran and Seghers 1994), as well as over parental investment (Trivers 1972), and it may lead to the evolution of traits (e. g. behaviours or protein production, which are determined by gene expression) that are beneficial to one sex, but that impose a cost to the other (Parker 1974). Such antagonistic coevolution resembles an evolutionary arms race, where adaptations to bring the interaction closer to the optimum value for one sex are met by counter-adaptations towards the optimum value for the other sex.
For a parent, it is typically beneficial to make its partner to invest more than itself in their common offspring (McNamara et al. 2003; Osorno and Székely 2004), because it enables a reduction in the amount of care it provides. Thus, sexually antagonistic coevolution is expected to give rise to attributes that induce partners to increase their reproductive investment, and to traits to resist such inducement (Chapman et al. 2003). Although demonstrations of antagonistic coevolution often deal with phenotypic traits such as those used to induce mating (Arnqvist and Rowe 2002; Buckling and Rainey 2002; Macías Garcia and Ramirez 2005), it has also been reported in relation to traits that influence provisioning to developing offspring, such as the augmented maternal investment by birds exposed to attractive male traits (Burley 1981; Gil et al. 1999). Sexual conflict can occur at the simpler but very transcendental level of the expression of genes in charge of regulating nutrient transfer during offspring development (Moore and Haig 1991) or it might also be related to signalling pathways that control resource allocation (Zwoinska et al. 2014).
We find that although the potential role of parent–offspring and sexual conflict in driving the evolution of viviparity has been recognised by some authors (Crespi and Semeniuk 2004; Blackburn 2015a; Geist et al. 2019), in general, it has been undervalued. Conflict is in fact absent from most reviews regarding the evolution of viviparity and its traits in several viviparous taxa (Wake 1992; Murphy and Thompson 2011), and although some authors recognise the impact of epigenetic regulation on the evolution of viviparity, this is frequently associated with changes in the environment (Albergotti and Guillette 2011). Thus, we argue that conflict can, in fact, explain more attributes of the different modes of viviparity than has previously been contemplated Additionally, we propose that once one trait related to viviparity and matrotrophy evolves, it sets up a new selective environment that promotes the evolution of other traits, leading to a diversity of forms of embryonic development and nutrition and thus contributes to the current diversity of viviparous vertebrates.
The evolution of viviparity
In vertebrates, oviparity is the ancestral reproductive pattern, in which propagules enclosed within an egg envelope are fertilised—outside or inside the female body—and develop and hatch commonly in the external environment. Oviparous embryos are usually nourished exclusively from the vitellum (egg yolk). Viviparity, by contrast, is a mode of reproduction characterised by internal fertilisation and egg retention, in which embryos fully develop within the female reproductive tract and are released to the external environment as free-living animals. The condition where embryos develop with minimal interaction to the maternal tissues, beyond some gas exchange, and hence embryonic nutrition depends on the yolk, is known as lecithotrophic viviparity, whereas the condition where embryonic nutrition is provided by the mother once the egg yolk is depleted is known as matrotrophic viviparity (Wourms et al. 1988).
Animal viviparity has evolved independently over 160 times, including 142 instances of convergent evolution amongst vertebrates (Blackburn 1999, 2015b). Given the diversity of conditions currently experienced by viviparous taxa, it is not clear which selective force, or forces, promoted viviparity in the first place. This mode of reproduction confers a variety of demonstrated or suspected fitness benefits, several of which have been proposed as the primary forces driving its evolution (Blackburn 1999), yet each tends to be relevant only in some of the viviparous taxa, and thus, we lack a unifying theoretical framework for the evolution of animal viviparity and matrotrophy (embryonic nutrition via maternal resources other than yolk; Table 1, see Supplementary Information). Here, we attempt to fill this gap in relation to vertebrate viviparity.
It is unlikely that all the viviparous clades followed precisely the same path in the evolution of viviparity and/or matrotrophy. Still, we argue that the underlying evolutionary conflicts, which are common to- but played differently in the various viviparous clades, performed a key role in the evolution of traits related to viviparity and/or matrotrophy. We also argue that the diversity of reproductive patterns and their particular adaptations are the result of different combinations of conflict-driven selective pressures and the ecological context in which they take place.
Both gradualist (Blackburn 1992; Whittington et al. 2022) and saltationist models (Blackburn 1995) have been proposed to explain the evolution of viviparity and placentation. Yet neither gradual nor rapid transition from oviparity to matrotrophic viviparity would have been possible without the evolution of 1) the initial acquisition of internal fertilisation and egg retention, which seem to be a pre-requisite for the evolution of viviparity sensu stricto (Blackburn 1999), followed by 2) internal embryonic development within typically the female reproductive tract, where embryos are nourished only with nutrients contained in the vitellum (yolk) of the ovum (Blackburn 2000). Subsequently, 3) a lengthening of developing time (extended uterine gestation) matched by an increasing supplementation of nutrients released by the mother into her reproductive tract (incipient matrotrophic viviparity), and culminating, in some cases, in what we know as 4) matrotrophic viviparity (Blackburn 2000), where instead of yolk, nutrients are gradually provided by the mother in the form of oviductal secretions or through placental organs (see Supplementary Information). Thus, from an egg-laying ancestor whose egg may already have had genetic and physiological attributes to enable a primitive form of matrotrophy (Wourms 1981), females would have become able to give birth to fully developed, independent, and particularly, as increasingly effective adaptations for embryonic nutrition evolved, bigger offspring than their oviparous counterparts (Sibly et al. 2018). The above route, however, has not been followed by all viviparous taxa (Blackburn 1992), and species are often in intermediate states, as in the case of caecilians that evolved dermatotrophy, a type of oviparity where oviparous new-borns ingest some modified and nutritious maternal skin (San Mauro et al. 2014; Kupfer et al. 2016). Indeed, lecithotrophy and matrotrophy are not absolute conditions, but rather are extremes of a continuum of female investment (Wourms et al. 1988; Stewart and Thompson 1996; Blackburn 1998; Riesch et al. 2014). Several species regarded as lecithotrophic, also present some form of matrotrophy (e. g. histotrophy, which is a type of embryonic nutrition based on maternal nutrients that are absorbed by the embryo through specialised structures, such as the skin or gill epithelium) at the end of the embryonic development, after hatching and before birth, such as the stingrays (Hamlett et al. 2005) or have a very simple placenta, as in some reptiles (Stewart 1992). This highlights the need for a theoretical framework that may accommodate also more divergent paths towards matrotrophic viviparity.
Conflict and the evolution of viviparity
Conflict during the evolution of viviparity and matrotrophy: proposed pathway
Theories proposed to explain the evolution of viviparity, matrotrophy, and placentation are diverse and often contradictory, and it is unlikely that a single explanation holds for the whole of the complex, multi-stage transition from oviparity to matrotrophic viviparity among vertebrates. More probably, it must have been shaped by a variety of selective pressures that interacted simultaneously or sequentially, and even such interactions or sequences of events may have differed in the distinct lineages that became matrotrophic, thus promoting lineage diversification. Typically, reviews and theoretical papers have focussed on only one, or a few, of the evolutionary transitions that must have occurred between oviparity and matrotrophic viviparity. Here we present a possible scenario whereby natural selection and genetic conflicts of interests may have driven, through a diversity of particular pathways, the evolution of the several instances of vertebrate matrotrophy from an oviparous ancestral stage (Fig. 1).

Possible pathway leading to the evolution of matrotrophic viviparity and placentae from an ancestral oviparous condition. The evolution of matrotrophic viviparity and placentotrophy from oviparity was probably the result of several selective pressures acting together. Although the appearance of live-bearing reproduction had mutual benefits for mothers and their offspring, different types of conflict also must have favoured the evolution of internal fertilisation, the staggered delivery and intake of maternal resources, and the diversity of forms of matrotrophy and placentae
Internal fertilisation
Following the idea that females are generally the limiting sex, as their reproductive output is normally set by the number of eggs they can produce, whereas that of the males is determined by how many female eggs they have the capacity to fertilise (Trivers 1985), there is a premium for males to gain access to fertile females before other males do. This leads to protandry, by which males emerge/arrive at the breeding ground before females (Wiklund and Fagerström 1977). This form of male–male competition that favours males to be prepared to mate before their rivals would, in externally fertilising species, promote a heightened readiness to ejaculate just as, or shortly before, spawning occurs (Fig. 1). In the extreme, any male feature that can allow males to deliver sperm directly into the female reproductive tract before egg laying would be favoured by intrasexual selection, even in the absence of any female adaptation for egg retention. Accordingly, it has been proposed that internal fertilisation initially evolved because: a) it can reduce the intensity of sperm competition (Parker 1970), or b) even in the absence of sperm competition, it can reduce the risk of sperm being lost, and hence increase the probability that eggs are fertilised (Parker 1984). If the original driving force was reducing the intensity of sperm competition, then internal fertilisation might have evolved through intra-sexual conflict, as male competition would determine paternity (in the absence of cryptic female choice). The ubiquity of external fertilisation in several taxa inhabiting all types of aquatic environments, such as many fish and amphibian species, suggests that sperm loss is not a very strong selective force against external fertilisation. Concurrently, the frequent evolution of male coercive means to ensure a successful copulation, such as the presence of claspers (Shibukawa et al. 2012), and a wide variety of male adaptations to deal with sperm competition in both oviparous and viviparous clades (Stockley et al. 1997) support our view that male–male conflict lies at the root of the evolution of internal fertilisation.
Hypotheses about the origin of internal insemination cannot be tested in amniotes, as they lack variation in the occurrence of this trait. It is even likely that, rather than being an adaptation to life outside water, internal insemination was a prerequisite to the independence from an aquatic medium for reproduction. Therefore, the following predictions pertain only anamniote vertebrates. If the attempts of males to monopolise females and fertilise their eggs favoured the evolution of internal fertilisation, then we expect that internal insemination has evolved more often in clades where multiple paternity amongst externally fertilising species is common. We also expect female adaptations to counter male monopolisation, such as sperm storage and superfetation, to be more common in clades where males have evolved more coercive means of securing internal insemination.
Alternatively, it has been suggested that external fertilisation predicts the occurrence of male parental care (Benun Sutton and Wilson 2019) and that the transition from external to internal fertilisation is linked to the transition from paternal to maternal care among fishes and amphibians (Beck 1998; Mank et al. 2005; Kahn et al. 2013), two groups of viviparous vertebrates where paternal care is common.
Again, lack of variation in insemination mode, and the fact that in the overwhelming majority of mammals (Balshine 2012) and reptiles parental care is provided by females makes this prediction impossible to test in amniotes although we note that in the absence of conflict, we would expect bi-parental and uniparental care to be equally likely to occur. Yet the proposal that the sex that releases the gametes first is the one that can desert parental care (Dawkins and Carlise 1976), can be tested in anamniotes, where we expect that any adaptation that allow males to release their gametes inside the female body and to fertilise her eggs, such as adaptations for sperm transfer and motility (Costa et al. 2016; Yokoe et al. 2016) would be a favoured outcome of intrasexual (male–male) conflict over egg fertilisation. This suggest that conflict related to parental care could also have acted as a selective force in the evolution of male strategies that initially favoured internal fertilisation as a means to monopolise eggs, and subsequently allowed males to decrease their parental investment in the offspring.
As a third possibility, connected to the above argument, it has been suggested that external fertilisation evolved from internal fertilisation among the vertebrate ancestors (Long et al. 2014). This is also compatible with our proposal that sexual conflict underlies the evolution of fertilisation mode, since even if internal fertilisation evolved initially among gnathostomes, external fertilisation could have subsequently evolved as a female adaptation to desert parental care in aquatic environments (where sperm loss is not a major concern).
Egg retention
Egg retention is considered a result of adaptations to counteract the effect of harsh environmental factors and pre-hatching predation risks (natural selection pressures; Andrews and Rose 1994). According to this view, increased offspring survival would have followed from intrauterine development, hence in a more secure and stable environment that also accelerated their development or that allowed females to enhance offspring fitness by manipulating some of their phenotypic traits (Shine 2014). This may sometimes have been the case, but we note that harsh environments are just as likely to promote the production of resistant eggs that can survive hard environmental conditions, even if their parents themselves cannot. This, for instance, is the case of annual fishes (Cyprinodontiformes; Murphy et al. 1999), a diverse group of tropical minnows that inhabit ephemeral water bodies, which they re-populate each wet season from draught-resistant eggs (Rodao et al. 2015). Hence, while sometimes they may promote egg retention, harsh environments may in other cases lead to the production and laying of even more independent eggs. Thus, as with internal fertilisation, egg retention may have evolved in response to different selective pressures, some of which may be related to conflict (Fig. 1). (Motz and Callard 1988; Guillette et al. 1991a; Callard et al. 1992).
The presence and conserved function of some hormones that promote or delay oviposition is ubiquitous among vertebrates (e. g. luteinising hormone (LH), arginine vasotocin (AVT); Pickford 1952; Bercu et al. 1980; Guillette and Jones 1982; Guillette et al. 1991b; Wang et al. 2008). Thus, it is likely that the production of hormones that induce egg retention—or the lack of hormones that induce oviposition—evolved first as a female mechanism to control oviposition depending on a variety of circumstances, such suboptimal places for oviposition, or to avoid predators or infanticide (Schneider 1999; Matsushima and Kawata 2005; Montserrat et al. 2007; Thiem 2020). Such mechanism was open to co-option by males or offspring to induce a longer egg retention, either i) via chemical suppression of female hormones that induce oviposition, such as AVT; ii) via embryonic production of hormones that induce egg retention, such as LH or hormones similar to progesterone and oestrogen (Motz and Callard 1988; Guillette et al. 1991a; Callard et al. 1992); or iii) if males or embryos could somehow promote embryonic development before eggs are laid, such as the embryonated eggs of caecilians by influencing their growth rate (see belowMotz and Callard 1988; Guillette et al. 1991a; Callard et al. 1992).
Testing this conflict hypotheses, however, is currently complicated due to the lack of information regarding the identity and regulation of hormones than induce oviposition, except for AVT, and egg retention in non-mammalian vertebrates. Nonetheless, if egg retention evolved as a female strategy to increase the embryos’ survival or as a consequence of conflict, this became a key precondition for the emergence of an antagonistic coevolution between the mother and the offspring and/or the father regarding the allocation of maternal resources. Once embryos were retained inside the female body, the eggshell became permeable and post-fertilisation mother-embryo communication was promoted, mothers became susceptible to embryonic physiological manipulation or to the embryos gaining control over nutrient consumption rate and amount (see section Mother offspring conflict and the evolution of matrotrophic viviparity). Under this scenario, we then expect to find among oviparous species with different degrees of egg retention, correspondingly variable offspring adaptations that allow them to i) hatch inside the female body if certain development stage is reached and ii) increase their nutrient consumption (through large or efficient embryonic components of the egg membranes that favour a more effective nutrient acquisition or through teeth or structures to eat maternal tissues or siblings). We would also expect to find an ancestral version of those embryonic traits exclusively in oviparous clades with some degree of egg retention, but not in taxa with no egg retention, and a modified version (improved or vestigial) of such adaptations in viviparous and closely related species, such as the case of the deciduous dentition of matrotrophic caecilians (Wake 1977b), and the specialised dentition of the oviparous taxa (Kupfer et al. 2006b).
At present, several examples among vertebrates of eggs that are laid with embryos in an advanced developmental have been documented. This is the case of the embryonated eggs of some caecilians (Kupfer et al. 2006a) and of sporadic accounts of developing fish embryos being laid by otherwise oviparous species (Hayakawa and Munehara 2001, 2003), something for which fish eggs may be regarded as pre-adapted (Wourms 1981). Among reptiles, there are frequent cases of eggs laid with embryos in stages 29–31 (Blackburn 1995). However, there is evidence that shows that a successful egg retention also depends on female’s anatomy and embryonic factors specific to each species (Andrews 1997). If egg retention is a consequence of internal fertilisation and a pre-requisite for viviparity, but its evolution is not equally successful in every taxon, then we expect the evolution of a more frequent and successful egg retention (which does not have a negative impact in embryo’s developmental time compared to developmental status when born, or survival) in species with: 1) egg or embryonic primitive structures similar to placental analogues or equivalents, and 2) a relatively thin and at least partially permeable eggshell, and 3) small clutch sizes.
Staggered embryo provisioning
In internally fertilising egg laying species insemination is often followed by a short-term egg retention during which the already yolked egg receives additional nutrients and, in some taxa, a hard, protective shell is added (Kupfer et al. 2006b). Long-term egg retention is incompatible with the presence of a hard shell that would impede gas exchange and could injure the female if accidentally broken. Mothers of strict lecithotrophic species face two constraints: 1) their provisioning of resources cannot be modified over time if conditions improve, and 2) females are encumbered with the fully provisioned eggs through the gestation period, which compromises movement performance (Ghalambor et al. 2004). Those constraints place a premium on what we call here staggered embryo provisioning, i.e. the production of eggs with little yolk (with a subsequent supplementation of nutrients via oviductal secretions or the consumption of other siblings/eggs after hatching inside the female body), or with no yolk and a gradual supply through gestation.
The diversity of staggered embryo provisioning patterns in vertebrates is vast. Among therian mammals, which are highly matrotrophic, embryonic nutrition is possible thanks to the yolk sac and the chorioallantoic placentae (Smith 2015). Viviparous squamates, just like mammals, also evolved a placenta that results from the apposition of the chorioallantois and a specialised derivative of the yolk sac, to the lining of the uterus of the mother (Stewart and Blackburn 1988). Although most viviparous squamates are relatively lecithotrophic, and mothers only transfer small amounts of nutrients to the developing embryos through the placenta, there are six clades with considerable embryo provisioning via placentotrophy (transfer of maternal nutrients via the placenta; see Supplementary Information), which in a few cases can be also supplemented with additional resources (reviewed in Blackburn 2015b).
Amphibians have evolved staggered embryo provisioning several times, but unlike mammals and reptiles, this has been accompanied by a diversity of embryonic and maternal adaptations for oophagy (ingestion of fertilised or unfertilised ova), embryophagy (ingestion of siblings), histophagy (ingestion of maternal secretions), matrophagy (ingestion of maternal tissues) or a combination according to each species (Table 1, see Supplementary Information; Wake 1977b; Guex and Chen 1986; Dopazo and Alberch 1994; Buckley et al. 2007; Buckley 2012). Such adaptations are sometimes shared by several species but seem to be the result of multiple origins (reviewed in Blackburn 2015b). Substantial matrotrophy has also evolved in the form of histotrophy, histophagy (ingestion of maternal secretion via specialised structures), placentotrophy, oophagy, embryophagy, or a combination among teleosts (Turner 1936; Wourms et al. 1988; Hollenberg and Wourms 1994, 1995; Meisner and Burns 1997), and Chondrichthyans (Springer 1948; Gilmore et al. 1983; Wourms et al. 1988; Hamlett and Hysell 1998). Relatively, lecithotropic chondrichthyans may also present some degree of staggered embryo provisioning, in the form of oophagy and histotrophy (Wourms 1977; Wourms et al. 1988; Compagno 2001). Although different taxa seem to share the same patterns of embryonic provisioning, it is important to note that the same matrotrophy patterns can have notoriously different adaptations across taxa (reviewed in Blackburn 2015b).
In contrast to strict lecithotrophy, exclusive or predominant matrotrophic viviparity allows females to carry the full biomass of the clutch for only a fraction of the gestation period, hence reducing the energetic cost of mobility and the period of greater vulnerability to predation (Hagmayer et al. 2020), and to adjust the rate at which she delivers resources to the embryos in response to changes in ecological conditions (Pollux and Reznick 2011). Full maternal control of this process may not be adaptive to the embryos (Einum and Fleming 2000), or to all the embryos in a clutch, and this may lead to a departure from the honest signalling of embryonic needs (Godfray 1995; Haig 1996) and to the evolution of means that allow the embryos to control the flow of resources from the mother, such as the embryonic component(s) of the placentae (i. e. to changes in the pay-off matrix of mothers and embryos). Thus, although staggered provisioning might be beneficial for both the mother and the embryo, specific forms of matrotrophic viviparity may not be evolutionarily stable, as they can be open to invasion by manipulative strategies from either the embryos or the father(s) of the clutch (see section Mother offspring conflict and the evolution of matrotrophic viviparity).
Placentae can be considered, in general, as the most advanced form of matrotrophy because of their capacity for nutrient transfer and gas exchange (Ostrovsky et al. 2016). If placental diversity is due to divergent adaptations to a variety of ecological conditions calling for special requirements in the transport of nutrients (as it has been suggested in adaptive theories for the evolution of placenta: see above), then we would expect the same type of adaptations and placentae in closely related species that live in similar environments. However, the environment experienced by, say, zebras and gazelles is essentially the same, whereas their placentae are widely different (Roberts et al. 2016). Considerations such as these led some authors to emphasise the importance of intense selective pressures, such as conflict, as important factors that shaped changes in the anatomy and function of the mammalian placenta during mammalian cladogenesis (Wildman et al. 2006; Roberts et al. 2016). It also led Crespi and Semeniuk (2004; see also Klisch and Mess 2007) to propose that antagonistic parent–offspring coevolution might explain the differentiation of placental types within and across taxa (Uribe and García Alarcón 2005; Mess and Carter 2007). However, this type of conflict may only influence the evolution of viviparity and its traits once internal embryonic development and matrotrophy emerged. Parent–offspring conflict has been implicated in the differentiation of placental types (a form of matrotrophy); however, there are also other forms of matrotrophy whose evolution may also have been linked to conflict. If this is true, then we expect to see across phylogenies, evidence of antagonistic coevolution between mothers and embryos (or fathers) in the form of i) gradual increases of invasiveness of embryonic component of the placentae, ii) a continuum in the degree of nutrient ingestion in the form of ova, siblings or maternal tissues within clades or families. Also, since the outcome of conflict over embryo provisioning is likely to be context-dependent, we may also expect iii) occasional polymorphisms in the degree of matrotrophy (matrotrophy index, MI = dry mass of the offspring at birth divided by the dry mass of the egg at fertilisation) within species, which can be coupled with polymorphisms in the mode of nutrient transfer. We also expect to see maternal adaptations to regulate the embryonic consumption of resources, such as reversals to oviparity (or via gene expression) in clades which also include highly developed placental systems (see section Genetic conflict and the evolution of genomic imprinting).
Mother offspring conflict and the evolution of matrotrophic viviparity
We have argued ways in which sexual conflict could have influenced or been the main driver of the evolution of most of the salient traits associated with viviparity. In this last section, we will develop the proposal that mother–offspring conflict has played a crucial role in the evolution of matrotrophy. Crespi and Semeniuk (2004) advanced a very well supported proposal showing that mother–offspring conflict is present in many forms of vertebrate viviparity, and that it is the main leading force behind the evolutionary diversification of mammalian placentae. Here, we extend that proposal and suggest that the offspring drive to extract as much nutrients as possible from the mother lies at the root of the mother–offspring conflict, the main selective force that led to the evolution and diversification of the extant forms of vertebrate matrotrophy and their particularities (Fig. 1).
Pre-adaptations for the evolution of matrotrophy in the different vertebrate groups
It is likely that certain pre-existing adaptations, such as a semi-permeable eggshell and/or egg membranes that allowed some small nutrients to enter the egg, or the appearance of new characteristics such as new embryonic adaptations to ingest dissolved nutrients, maternal tissues, or other eggs or siblings, facilitated the evolution of the diverse matrotrophic systems.
In the case of fish, for example, Morrison et al. (2017) found evidence suggesting that their eggs are preadapted for the evolution of matrotrophy (small molecules from the surrounding environment can traverse the egg membranes using mechanisms of active transport) and proposed that this is likely the main reason why matrotrophy has evolved considerably more times among fishes (Blackburn 2015b) than in reptiles and mammals (Blackburn 2015b).
Among mammals and reptiles, however, matrotrophy evolved in the form of placentotrophy, in which the chorioallantoic membrane and other pre-existing tissues of the amniote egg have been recruited for the formation of the placenta to enhance the maternal–embryonic communication and nutrient transfer (Griffith and Wagner 2017; Kent 2018). Additionally, this type of placenta co-opted the endocrine function of the egg’s chorioallantoic membrane and shows expression of genes that are important for resource uptake during pregnancy (Griffith et al. 2017).
Finally, amphibian embryos evolved adaptations to feed on maternal nutrients, tissues, siblings, or other eggs, such as the modification of gills or skin (ectotrophoblast), foetal dentition, precocial development of the jaws, teeth, jaw musculature, and of the digestive tract (Wake 2015). In fish, similar as well as different traits have emerged to facilitate histophagy, histotrophy, placentotrophy, oophagy, and embryophagy (Blackburn 2015b).
Although all these new traits and specialisations were influential in the evolution of the different forms of matrotrophy, the ability of the embryos to hatch inside the female body, probably associated with the need to seek more nutrients (Wake and Hanken 1982; Buckley et al. 2007), was probably the key feature that allowed and favoured increased and diverse mother–embryo and embryo–embryo interactions, which subsequently could have favoured the other mentioned adaptations. Given that the evolutionary interests of the several actors are seldom completely coincident during reproduction (Trivers 1974; Arnqvist and Rowe 2005; Royle et al. 2012), this would have led to different levels of conflict among the members of the family (mother and embryos, or among siblings), and thus probably been responsible for the diversification within the different types of matrotrophy.
Internal hatching and the evolution of mother–offspring conflict
The release of embryos from any type of eggshell or hard, impermeable membrane inside the female body, and the permanence there of embryos for at least part of their development is a trait shared by all viviparous matrotrophic taxa. The removal of barriers between embryonic and maternal tissues allowed embryos to have access to new-, or to increase their access to sources of food. Once offspring are able to reach the maternal tissues, they can get into closer contact with her physiology, or other eggs or developing siblings, potentially gaining some control over their own nutrient intake to a level that may be sub-optimal for the mother.
In species that evolved matrophagy, for example, embryos consume maternal tissues, which is a form of maternal cannibalism and is likely to be costly for the mother, since embryos can ingest more tissue than what may be ideal for her to transfer to them. The interests of mother and offspring can also clash under oophagy and embryophagy, since the developing embryos ingest unfertilised eggs—potential siblings—and siblings in advanced stages of embryonic development (Gilmore et al. 2005). Among these species, although the embryo or embryos that are born develop faster and/or are better fed and with a greater chance of survival, eating potential siblings in the form of ova or developing siblings in which the mother already invested, may negatively impact her fitness.
Placentae can be another tool used by offspring to increase their nutrient intake. Reptilian and mammalian placentae can be very invasive (Blackburn and Flemming 2009; Kent 2018) and produce hormones able to manipulate and increase the maternal nutrient supply (Haig 1996). Even in the cases of non-invasive placentae, such as those of horses, embryos can influence the placental supply of nutrients by producing hormones, such as insulin like growth factor 2, that increases maternal resource allocation (Allen et al. 2002).
Although there is little evidence of hormonal manipulation in histotrophic and histophagic matrotrophy, as a rule, hormones are implicated in maternal–embryonic communication across taxa (Bowman et al. 2021). Thus, even where mothers seem to be in total control of nutrient supply, embryos may have the physiological tools to send deceiving signals of nutritional state or can develop other traits, such as organs or tissues that favour a continuous and a more effective intake of maternal secretions (Hamlett 1999; Blackburn 2015a).
The different means available to embryos for increasing the acquisition of maternal resources are not always mutually exclusive, and there are species where more than one form of matrotrophy co-occur. For example, among goodeid fish, embryos of G. multiradiatus and A. splendens, apart from receiving nutrients constantly through the trophotaenial placenta, also may ingest other eggs or viable siblings (Greven and Grossherr 1992). As mentioned above, embryos of S. salamandra start to ingest sibling eggs, and in some cases also less developed sibling embryos once they have consumed their own yolk (Buckley et al. 2007).
While the various forms of matrotrophy and their diverse features may constitute maternal strategies to provide additional resources to the developing embryos, the evidence mentioned above supports the idea that, at least in several cases, embryos have made use of those features to increase their nutrient uptake to levels that are probably sub-optimal for the mother. This suggests that the conflict that results from the offspring measures to increase their nutrient ingestion could have prompted the evolution of the different embryonic and maternal adaptations and counter-adaptations to gain control over the amount and pace of embryonic nutrient intake (see section Further evidence supporting the role of conflict during the evolution of viviparity and matrotrophy among vertebrates), and that such antagonistic coevolution would be responsible for the diversity of forms of matrotrophic viviparity seen in vertebrates. If so, we expect to see among highly matrotrophic groups with considerable embryonic control over nutrient ingestion rate or maternal allocation (i.e. placentotrophy, oophagy, embryophagy, and matrophagy) the evolution of maternal counter-adaptations such as reversals to oviparity or differential gene expression in tissues in contact with the embryos. More specifically, we expect to find more reversals to oviparity in clades with the aforementioned matrotrophic patterns (especially with highly invasive placentae or when more than one pattern has evolved in one species), and a clear correlation between the MI and i) the number of reversals to oviparity within clades, or ii) the number of differentially expressed genes that play a role in nutrient demand and transfer between the maternal and the embryonic component of the placentas, or between parental alleles in the offspring (see section Genetic conflict and the evolution of genomic imprinting).
Other theories for the evolution of matrotrophy and placentation that involve conflict
Both mutually adaptive and conflict hypotheses have been proposed to explain the evolution of matrotrophy and the origin and evolution of placentae. The driving forces behind the transition from lecithotrophy to matrotrophy are thought to be ecological, such as the locomotor costs imposed by a prolonged period bearing yolked eggs (Blackburn 1999; Ghalambor et al. 2004; Pollux et al. 2009), or the possibility of modifying temporarily the allocation of resources to embryos depending on resource availability (Blackburn 1999; Marsh-Matthews and Deaton 2006; Pollux and Reznick 2011), without compromising offspring survival. Alternatively, as proposed by Zeh and Zeh (2000), we also suspect that the transition may have been driven by postzygotic genetic conflicts, either among mother and offspring, between partners, or among siblings. The proposal is that once embryos were able to hatch inside the female body, genetic weapons, such as embryonic or paternal genes capable of manipulating the maternal physiology, could have promoted the evolution of protracted nutrient transfer. Thus, rather than being consequence of diverse ecological factors, the differences in maternal provisioning among viviparous animals would have been an outcome of the way intra-family genetic conflict over the allocation of maternal resources evolves in different taxa -and may have promoted lineage divergence (Helmstetter et al. 2016; Zeh and Zeh 2000). Indeed, while the basic set of hormones involved in the regulation of reproductive processes is highly conserved among vertebrates, their sources, functions, and targets differ among taxa; therefore, Crespi and Semeniuk (2004) proposed that such variation may reflect a long evolutionary history of maternal–foetal antagonistic coevolution.
The increased and prolonged maternal provisioning during embryonic development takes place in several animal groups through the placenta or placenta-like structures (Blackburn et al. 1985). This probably evolved from pre-existing tissues that acquired new functional attributes, modified their developmental programs, and evolved novel cell types (Griffith and Wagner 2017), allowing a close association between mother and offspring tissues, and an efficient exchange of nutrients, gases and excretions (Mossman 1991; Wooding and Burton 2008). Mammalian placentation has been described and studied in great detail, yet it evolved first in fish (Wourms and Lombardi 1992), and several other groups (reptiles, amphibians, invertebrates and plants) have also independently evolved placenta-like structures (Kaye et al. 1972; Haig and Westoby 1991; Blackburn 1999; Reznick et al. 2002) that are responsible for increasing of embryonic dry weight from zygote to birth of up to 38,700% (Goodeid fish; Lombardi and Wourms 1979).
Although the primary function of placentae is shared among taxa (Faber et al. 1992), it is one of the most morphologically and physiologically diverse vertebrate organs (Mossman 1991). This organ constitutes a key functional link in the transition from lecithotrophy to matrotrophy, providing the physiological scenario in which conflict can be expressed. In fact, some authors beyond Crespi and Semeniuk (2004) have proposed and found evidence that suggests that the divergent interests between mother and offspring, followed by a rapid antagonistic coevolution, were the main causes of the diversifying evolution of the mammalian placenta (Klisch and Mess 2007), and that the proteins that that mediate in this organ the mother-embryo interactions may often be targets of evolutionary conflict (Chuong et al. 2010). Furthermore, there is evidence that most imprinted genes—those that are expressed in a parent-of-origin manner—are expressed in the placenta (Kaneko-Ishino et al. 2003), and some of them are essential for placental development and growth (Baker et al. 1993; Renfree et al. 2013). Among these, the genes that increase embryonic growth are usually paternally expressed, whereas those that tend to restrict growth are maternally expressed (Renfree et al. 2013); thus, additional types of conflict, such as conflict between males and females, the parental alleles in the offspring or between half-siblings in the womb, are likely playing a role in the evolution of the placenta and its physiology (Burt and Trivers 1998; Parker 2006; Moore 2012).
Genetic conflict and the evolution of genomic imprinting
Genomic imprinting is a widespread phenomenon, in which certain genes are expressed in a parent-of-origin manner, usually as a result of DNA methylation or histone modifications (Tilghman 1999).
Trivers (1974) was the first to mention the possibility of a conflict between parents and offspring related to differing genetic interests, which could drive offspring to employ physiological weapons to manipulate maternal investment. During pregnancy, different sources of genetic conflict may arise: i) between genes expressed in the mother and in the foetus/placenta, or ii) between maternally derived and paternally derived genes within the embryonic genome (Haig 1996). Whenever there are mother–offspring interactions, four sources of genes can be recognisable: a) genes expressed in the mother, b) genes expressed in the offspring, c) maternally inherited genes expressed in the offspring, and d) paternally inherited genes expressed in the offspring (Crespi and Semeniuk 2004).
The evolution of the gene expression type (a) and (b) may be shaped by a process of antagonistic coevolution between mother and embryo. Under this scenario, a genetic conflict may arise between maternal and foetal genes, where expression of the latter will be selected to increase the transport of maternal nutrients, and maternal genes will be selected to be expressed so that nutrient transport takes place at a level that is optimal for the mother (Haig 1993). Indeed, in mammals, hormones produced by the embryonic placenta may be interpreted as foetal attempts to manipulate maternal metabolism for the offspring’s benefit (Haig 1996). Gene expression type (c) and (d), on the contrary, may reflect a conflict between parental alleles expressed in the offspring, where paternally derived alleles will be selected to favour a more efficient nutrient acquisition and maternally derived alleles will be selected to favour an even distribution of maternal resources among broods (Haig and Westoby 1989). This kind of conflict has been interpreted as the force that drove the evolution of genomic imprinting.
The Kinship Theory of genomic imprinting mainly proposes the parent-of-origin gene expression evolved as a consequence of a conflict between the interest of the paternally inherited alleles (padumnal alleles or patrigenes) and maternally inherited alleles (madumnal alleles or matrigenes) over maternal investment during offspring development (Moore and Haig 1991; Haig 2000). According to the Kinship theory, because the conflict is associated with maternal allocation of resources, it is predicted that the genes that increase nutrient demand from the mother will be paternally expressed and the genes that restrict embryo growth will be maternally expressed (Haig 1996, 2000; Renfree et al. 2013). This is more likely to evolve when: 1) offspring are sired by more than one father, either among litters or in the same litter, 2) provisioning of offspring during development is largely performed by the mothers, and 3) there are genes expressed on the offspring that can manipulate the amount of resources that the mother provides (Wilkins and Haig 2003).
A classic example of genomic imprinting that is consistent with kinship theory is the expression pattern of the insulin-like Growth Factor 2 (igf2) and its receptor igf2r (Haig 2004). IGF2 is a protein that, among other functions, promotes growth and cellular differentiation during development (Cohick and Clemmons 1993). It also regulates the placental supply of nutrients, and the genetic demand for nutrients by the foetus (Constância et al. 2002; Fowden et al. 2006). On the other hand, igf2r encodes a membrane protein (cation independent mannose-6-phosphate receptor or IGF2R) that captures and transports the excess the of mannose-6-phosphate and IGF2 to the lysosomes for posterior degradation (Kornfeld and Mellman 1989), and thus it is essential for regulating normal foetal growth, circulating level of IGF2, and heart development (DeChiara et al. 1991; Lau et al. 1994). In therian mammals, the paternal allele of igf2 is expressed and the maternal allele is silent (DeChiara et al. 1990) while igf2r is maternally active and paternally silent (Barlow et al. 1991). This accords to the proposal by Moore and Haig (Moore and Haig 1991).
A new perspective of conflict and genomic imprinting
Haig (2000) proposed that the conflict between parental alleles drove the evolution of genomic imprinting. More generally, this form of control of gene expression may have evolved in the context of sexual conflict. For example, in the case of IGF2, a protein that controls the demands for nutrients by the embryo, and the nutrients delivery by the placenta, the cost of an increase in maternal investment (imposed by a paternally induced excess of the embryonic protein) is only faced by females. This makes the interest of both sexes diverge and clash, especially if fathers do not sire subsequent litters that the female may produce, leading to an evolutionary arms race between sexes that takes place at a genomic level and reflects allele conflict, as Haig (2000) suggested. However, it is difficult to establish whether the alleles are in conflict directly with each other, or if their evolution is directed by the conflict between sexes. In other words, the conflict that we see in viviparous polyandrous systems between parental alleles is part of the conflict between males and females.
It has been suggested that among vertebrates, genomic imprinting is a characteristic exclusive of viviparous mammals that appeared before the marsupials and eutherian split, and that evolved differentially in both groups, resulting in a larger number of imprinted genes in eutherians compared to marsupials (Renfree et al. 2013; Fig. 2). Thus, the evolution of genomic imprinting as a result of conflict between parental alleles, or between the sexes, over maternal allocation of resources has been extensively studied in mammals (Tycko and Morison 2002; Haig 2004). However, viviparity and maternal provisioning of resources during pregnancy (the main enhancers of this type of conflict) have been also documented in many vertebrate taxa, including fish. Although signs of genomic imprinting have not been found in reptiles, birds and marsupials so far (e.g. O’Neill et al. 2000; Griffith et al. 2016; Schwartz and Bronikowski 2016), the evolutionary foundation of genomic imprinting has been demonstrated in oviparous fish (Xie et al. 2009); reprogramming of DNA methylation similar to that of mammals has been demonstrated in zebra fish (MacKay et al. 2007; Daneshfozouna et al. 2015), and there is evidence that insulin-like growth factor 2 is under positive selection, which coincides with the evolution of placentation in fishes (O’Neill et al. 2007). Moreover, spontaneous abortion rate in crosses between populations of poecilid fish with a different level of polyandry has been related to the postzygotic reproductive isolation as a product of parent–offspring conflict (Schrader and Travis 2008), and there is already evidence suggesting that igf2 is imprinted in a family of highly matrotrophic viviparous fish (Saldivar Lemus et al. 2017).

Scheme of appearance of genomic imprinting. Genomic imprinting has been documented in therian mammals, and it is known to be absent (N) in monotremes or birds. There is evidence that suggest that it is also present in at least one species of the fish family Goodeidae (Saldivar Lemus et al. 2017). The possibility of the evolution of a parent-of-origin gene expression in reptiles, amphibians and fish has been poorly investigated (O’Neill et al. 2000; Griffith et al. 2016; Schwartz and Bronikowski 2016) and deserves further attention
The above suggests that the same type of conflict and antagonistic coevolution that has been documented in mammals may be occurring in other organisms with similarly strong conflict over maternal allocation of resources and with similar mating systems. Therefore, although some attempts to find evidence of genomic imprinting in vertebrate taxa other than mammals have been conducted, we suggest that more research is needed in viviparous vertebrates, especially in fish, in relation to genomic imprinting on genes in charge of embryo-maternal communication and nutrient transfer. In line with the genomic imprinting conflict theory, we expect to see in other groups that the paternally expressed alleles promote growth while maternal alleles restrain it, and that such balance as may be observed today would have arisen by coevolution, being facilitated by clustering of genes with antagonistic effects (Reik et al. 2003). While igf2 seems to be the most studied and best described example of genomic imprinting and conflict in mammals, there are many other genes that enhance or restrict growth during development in mammals (Table 2). Thus, we expect that further research will reveal even more genes involved, both in mammals and in other viviparous vertebrates. Additionally, although 1) genomic imprinting has not evolved in the same way even among organisms of the same group (Kalscheuer et al. 1993; Pearsall et al. 1996; Okamura et al. 2000), and 2) genomic imprinting of important genes, such as igf2, does not occur in all the matrotrophic groups, this does not necessarily mean that imprinting as a result of sexual conflict over maternal allocation of resources has not evolved in some other growth-related genes. And we note that almost none of those genes have so far been tested in other viviparous and more matrotrophic organisms than mammals.
Further evidence supporting the role of conflict during the evolution of viviparity and matrotrophy among vertebrates
An extended mode of amphibian reproduction involves the occurrence of a larval stage, which in several cases can undergo metamorphosis before hatching. Larvae may ingest unfertilised eggs provided by the mother (Buckley 2012), while in oviparous caecilians, females may exhibit extended egg retention, and thus lay embryonated eggs (Kupfer et al. 2006a). These allow the mother to control a staggered delivery of resources to the developing young, and there is little opportunity for the latter (or the father) to control the female investment in their own benefit. The embryos of viviparous salamanders and caecilians sometimes have specialised teeth that scrap the maternal tissues (Buckley 2012) and may have evolved because they promote the interests of the father (i. e. being the result of sexual conflict), unless its expression was a function of the embryos’ nutritional state, a condition that would suggest mother–offspring conflict. This form of embryotrophy (known as matrophagy, Table 1, see Supplementary Information) may preclude the evolution of an embryonic placenta, and in some salamander species is preceded by an oophagous phase. Oophagy (Table 1, see Supplementary Information) may allow the female to determine the maximum investment she is willing to make in a given reproductive effort, while the embryos regulate the tempo at which the resources are consumed. Embryophagy (Table 1, Supplementary Information), on the other hand, limits the control that can be exerted by the female on the rate at which resources are passed from the mother to the offspring, since embryos can ingest their viable siblings to which females have allocated nutrients during gestation. Although this form of amphibian embryotrophy is exclusive of the fire salamander Salamandra salamandra (Linnaeus, 1758) (Buckley et al. 2007; Buckley 2012) and little is known about its mating system, we predict a stronger selective pressure for the evolution of embryophagy in polygamous mating systems with multiple paternity. This is because the payoff of half-sibling ingestion is greater than that of ingesting full sibs, and because the cost of embryophagy is unequally shared between mother and fathers. We further predict that once embryophagy evolves, females have very few alternatives to prevent it, short of reverting to oviparity (as in Chondrichthyes; see Supplementary Information, Fig. 8.
Both histophagy (Table 1, Supplementary Information), occurring in some salamandrids (Blackburn 2015b) and bufonids (Xavier 1973; Wake 1980), and histotrophy (Table 1, Supplementary Information) found among the hemiphractid anurans (Savage 2002; Roberts et al. 2016) should allow females to control the nutrient delivery; yet in some caecilians and salamandrids, embryos can induce maternal secretions by abrading the uterine lining with their prenatal teeth (Wake 1977a; Guex and Chen 1986). If the extent of the secretion-inducing damage was against the interests of the mother, we would expect to see mechanisms like maternally induced epigenetic inhibition of transcription factors associated with teeth formation (e. g. AmeloD; Chiba et al 2019) in embryos, and such maternal resistance should be a derived, and the expression of the embryonic teeth an ancestral condition within salamandrid clades.
Most viviparous squamates are relatively lecithotrophic and transfer small quantities of nutrients through the placenta via histotrophy (Blackburn 1994; Stewart and Thompson 2000; Thompson and Speake 2006); nevertheless, substantial matrotrophy has evolved in six clades of scincid lizards (Blackburn 1992; Stewart and Blackburn 2014). All squamate placentae have maternal (uterine epithelium) and embryonic components (chorioallantois or specialised derivatives of the yolk sac), and in the genera Pseudemoia, Mabuya, and Eumecia, they have the placentome (an absorptive embryonic structure with maternal secretory elements; (Thompson et al. 2004; Adams et al. 2005; Blackburn 2000).
Social monogamy is uncommon among reptiles (Harrison 2013) and multiple paternity, which can involve as many as 50% of all litters, has been documented in all lizard and snake species investigated so far (Uller and Olsson 2008). Thus, if maternal–foetal communication in squamate reptiles is regulated chemically, and offspring signal their necessities via hormones synthesised in/released by the placentome, then both the embryo and (or) the father may influence maternal resource allocation.
Fish are characterised by a continuous progression of viviparous species, from cases where mothers do little more than merely protecting the ova inside them, to true viviparous species, where the nutritional, respiratory, and excretory demands of the embryos are satisfied by the mother (Amoroso 1960; Wourms and Lombardi 1992) through structures such as placentae or ovarian nipples (Turner 1940; Lombardi and Wourms 1979; Blackburn 2015b), or via oophagy, embryophagy, histotrophy, and histophagy (see Supplementary Information). Substantial matrotrophy has evolved in at least nine of the 12 clades of viviparous teleosts and in four of the eight clades of viviparous elasmobranchs (Blackburn 2015b). Embryos can absorb or ingest nutrients deposited in the ovarian lumen or the ovarian follicle across permeable surfaces (Wourms 1981; Kunz 2004). Although oophagy is widely distributed among viviparous fishes (Blackburn 2015b), and the mother can control the maximum quantity of nutrients she provides, allocation mediated by placentae, histotrophy and histophagy, may still be influenced by the embryos or the father (see section on genomic imprinting above).
The bewildering diversity of adaptations surrounding viviparity in vertebrates may obscure any underlying evolutionary pattern. Still, as mentioned earlier, we propose that 1) certain sequential transitions are more likely to have occurred than others (e.g. lecithotrophic viviparity—matrotrophic viviparity; unchecked embryonic control of nutrients transfer—reversion to oviparity) and 2) that conflict of interests between the participants (mother, father, embryos) would have promoted diversity, as the processes and outcomes would vary from one instance to other (see Figs. 3 and 4). We used two families of teleost fish that include oviparous and viviparous species, Goodeidae and Zenarchopteridae and traced the features related to mode of reproduction and control over maternal nutrient transfer. Oviparity was the ancestral state in both families, and it apparently was followed by lecithotrophic viviparity, sometimes complemented by some mode of matrotrophic viviparity (Zenarchopteridae), or by matrotrophic viviparity in the form of placental viviparity (Goodeidae). Although placental viviparity is not present among zenarchopterids, some species that belong to the genera Dermogenys have evolved some structures that allow a maternal–embryonic connection (Meisner and Burns 1997) and that could be considered as very primitive placentae. The embryonic nourishment of the different lecithotrophic and some matrotrophic species of this family is characterised mainly by histotrophy/histophagy, where the mothers can control the amount of resources they transmit to their embryos, or by oophagy and embryophagy, which allow embryos to gain control over the amount of resources they ingest (Meisner and Burns 1997; Reznick et al. 2007; Fig. 3). The evolution of an incipient placental viviparity among Dermogenys species and the reversion to oviparity by Hemirhamphodon tengah (Tan and Lim 2013) may reflect the conflict among mother and siblings for controlling the maternal allocation of resources within this family. There is evidence of a similar conflict being at play in the Goodeidae (Fig. 4). For instance, the placenta is almost completely absent in a non-basal species: the striped goodeid Ataeniobius toweri (Meek 1904) which may have allowed females to regain control over the nutrient allocation via some type of histotrophy or histophagy. The opposite case, where embryos ingest their siblings, evolved in species such as the butterfly splitfin Ameca splendens (Miller and Fitzsimons 1971) (Greven and Grossherr 1992) and the Amarillo fish Girardinichthys multiradiatus (Meek 1904) (YSL and CMG pers. obs.). This suggests that conflict for controlling maternal allocation of resources is continuously shaping the evolution of patterns of matrotrophy among vertebrates. Both sets of predictions (sequence of events and conflict-linked diversity) require comparative analyses. We are, however, still ignorant of the ways in which internal gestation works in most of the taxa where it occurs, and appropriate phylogenies at the family or sub-family level are often lacking. Nonetheless, there are studies that suggest that viviparity has led to lineage diversification in Cyprinodontiformes (Helmstetter et al. 2016) and squamates (Recknagel et al. 2021), although not in all taxa (Zúñiga-Vega et al. 2016). Even though lineage diversification in reptiles has been linked to the evolution of viviparity resulting from environmental conditions and of each species’ genetic background (Recknagel et al. 2021), sexual- and mother–offspring conflict can also be tested as the main driver for the evolution of this reproductive pattern that is producing such diversification of lineages among vertebrates. We conducted a preliminary exercise of tracing several features of viviparity on the phylogenies of vertebrates (see Supplementary Information) to try to infer whether forms of viviparity and matrotrophy that resulted from an antagonistic coevolution between mothers or offspring/fathers for the control of the maternal allocation of resources, could have led to more complex forms of maternal–foetal interactions, and potentially, to the resolution of conflict. While some works have advocated a role of viviparity-linked conflict on evolutionary diversification (e. g. Helmstetter et al. 2016; Recknagel et al. 2021), the relatively crude descriptions of the modes of viviparity in several taxa, and the resolution at which information can be traced in the phylogenies preclude at this point the evaluation of those hypothesis for many families or orders.

Phylogeny of Zenarchopteridae and the attributes of viviparity and control over maternal allocation of resources. This is not an ancestral state reconstruction, but rather a graphic representation of the traits across the family. The external group (family Belonidae) and genus Zenarchopterus are oviparous. After the split of Zenarchopterus, it seems that the ancestor of the other genera developed lecithotrophy. Nomorhamphus and Dermogenys are lecithotrophic, however, some species have evolved also matrotrophic viviparity independently (Reznick et al. 2007) in the form of maternal and embryonic control. In the specific case of the genus Dermogenys, structures that could be considered as a primitive placenta (embryonic-maternal control) evolved in some species. Hemirhamphodon is a lecithotrophic genus except for one species that is oviparous. Information regarding embryonic in Hemirhamphodon is scarce; however, they appear to be lecithotrophic (Reznick et al. 2007). Crossed-out icons indicate the loss of the character

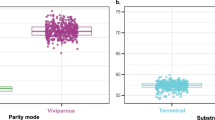
a Phylogeny of Goodeidae and the attributes of viviparity and control over maternal allocation of resources. All viviparous goodeids evolved viviparity from an oviparous ancestor. Although trophotaenial placenta (E-M control) is characteristic of the subfamily Goodeinae, which encompass the viviparous species, Ataeniobius toweri is characterised by the lack of a placenta and the evolution of some kind of histotrophy/histophagy (M control). Additionally, embryophagy (E embryonic control) has been reported in two species. b Photographs of males and females of selected goodeid species. Crossed-out icons indicate the loss of the character
Predictions and future directions
Our exploration of the modes of reproduction and the diverse types of embryonic nutrition among vertebrates allows some preliminary conclusions and some predictions:
a) We suggest that the evolution of internal fertilisation, which is a pre-requisite for the evolution of viviparity, was led by intra-sexual conflict, where the first copulating males can fertilise more eggs or where males that inseminate females can desert parental care. This is consistent with the fact that whenever claspers or other coercive means of internal fertilisation evolved, these are invariably present in males, and not in females. In the case of matrotrophic species lacking precopulatory female mate choice, b) we predict either the evolution of a strong cryptic female choice to select the sperm of the preferred male, or the evolution of a type of matrotrophic viviparity where females can control the amount of nutrients that are transferred to the developing embryos (i. e. histotrophy, histophagy, dermatrotrophy or some forms of oophagy where females provide unfertilised eggs specifically as a source of nutrients). In these species with maternal control of the nutrient transfer, however, we also expect c) counter adaptations in the offspring to gain control over maternal allocation, as in the case of placentotrophy—partial physiological control—or via embryophagy and oophagy of potential siblings. Further, d) we expect the evolution of these forms of matrotrophy more commonly occurring after the appearance of histotrophy, histophagy, or dermatrotrophy and not the other way around. Once evolved, some forms of embryonic control of maternal resources cannot be checked by the mother except by reverting to oviparity, which e) we predict would be more common in lineages where oophagy, embryophagy, or very invasive placentotrophy first evolved.
Several imprinted genes related to growth have been described to come to play during embryonic development in mammals (Table 2). We predict f) the evolution of genomic imprinting, or allele-specific DNA methylation patterns (seen as a primitive state of genomic imprinting), in genes related to growth in highly matrotrophic species with strong sexual conflict related to mating (e. g. polygamous species with highly dimorphic and/or courting males). In the specific case of placental species, as a result of the mother–offspring and sexual conflict, we expect g) more parent-of-origin expressed genes in placenta (as a whole) than in the developing embryo, with opposite expression patterns between the maternal and embryonic components of the placenta, or at least opposite DNA methylation levels in those genes in both components of the placenta. Under this scenario, h) the evolutionary rate of such genes should be asynchronous between components. Matrotrophic viviparity is very diverse among vertebrates, and we have argued that this is in part the evolutionary consequence of the different types of conflict among all the interacting parts. Most research has been devoted to the mother–offspring conflict and the evolution of placental matrotrophy in mammals. However, viviparity has evolved also in fish, amphibians and reptiles, and the diversity of the matrotrophic forms is greater and also likely the result of the conflicting interests of mothers, fathers, and offspring. To test these predictions, more research should be conducted on sequence evolution and DNA methylation patterns of key genes, Bayesian analyses to infer the order in which adaptations related to viviparity evolved, and phylogenetic analyses to infer the most likely sequence of events that led to the diverse forms of viviparity and matrotrophy that we see today.
Conclusions
-
(1)
Reproduction is costly, and selection has rewarded those organisms that can manipulate their partner to invest more than their fair share in raising their common offspring. The evolution of matrotrophic viviparity, and especially placentation, allowed offspring and fathers (via the expression of their alleles in the offspring) to influence the maternal allocation of resources.
-
(2)
Among reptiles, since there is conflicting evidence, it is impossible to determine if the evolution from oviparity to matrotrophic viviparity, including all the intermediate steps, and the subsequent appearance of a great diversity of forms of matrotrophy and placentae among vertebrates, has been gradual and shaped by several forces of natural and sexual selection or not. In other taxa, however, a higher diversity and complexity of matrotrophy forms can lead to different conclusions.
-
(3)
Although it is likely that egg retention and incipient matrotrophy would have been favoured at least partly because of the adaptive benefits that result from increasing offspring developmental stage at birth and their survival, while decreasing the mother predation risk and the possibility of losing the brood, we suggest that conflict also played a central role on its evolution. Additionally, we propose that the evolution of the variety of embryonic nourishment types in matrotrophic viviparous species and the enormous variety of mammalian placentae (and maybe of species among viviparous vertebrates) was probably due to the substantial conflict of interests between mother and offspring, and between mother and father—expressed in the genome of their offspring—regarding the optimal maternal allocation of resources during offspring development. This role of conflict seems to be constant and essential in the evolution of the different patterns of matrotrophy among vertebrates.
-
(4)
Patterns of lecithotrophy and matrotrophy are very diverse among vertebrates and so are their correlated traits. Although there are taxa that evolved one or the other, in most cases, species exhibit a type of viviparity that lies somewhere along that continuum. In many cases, once viviparity evolved, an evolutionary arms-race for the control of maternal resources seems to have ensued.
-
(5)
Since viviparity has evolved in most of the vertebrate clades and the patterns of viviparity and mating systems vary enormously across taxa, we suggest that the evolution of parent-of-origin methylation patterns related to maternal control of nutrient allocation has evolved in other viviparous taxa apart from mammals and that it should be investigated in amphibians, and reptiles, but especially in fish.
-
(6)
We propose that sexual conflict was a main driver during the evolution of most of the traits related to viviparity seen only as internal gestation, but mother–offspring conflict played a key role in the evolution of the different forms of matrotrophy.
Data availability
Data sharing is not applicable to this article as no datasets were generated or analysed during the current study.
References
Adams SM, Biazik JM, Thompson MB, Murphy CR (2005) Cyto-epitheliochorial placenta of the viviparous lizard Pseudemoia entrecasteauxii: a new placental morphotype. J Morphol 264:264–276
Albergotti LC, Guillette LJ Jr (2011) Viviparity in reptiles: evolution and reproductive endocrinology. In: Norris DO, Lopez KH (eds) Hormones and reproduction of vertebrates. Elsevier, Amsterdam, pp 247–275
Allen WR, Wilsher S, Turnbull C, Stewart F, Ousey J, Rossdale PD, Fowden AL (2002) Influence of maternal size on placental, fetal and postnatal growth in the horse. I Development in Utero Reproduction 123:445–453
Amoroso EC (1960) Viviparity in fishes. Symp Zool S 1:153–181
Andrews RM (1997) Evolution of viviparity: variation between two sceloporine lizards in the ability to extend egg retention. J Zool 243:579–595
Andrews RM, Rose BR (1994) Evolution of viviparity: constraints on egg retention. Physiol Zool 67:1006–1024
Andrews SC, Wood MD, Tunster SJ, Barton SC, Surani MA, John RM (2007) Cdkn1c (p57Kip2) is the major regulator of embryonic growth within its imprinted domain on mouse distal chromosome 7. BMC Dev Biol 7:53. https://doi.org/10.1186/1471-213X-7-53
Arnqvist G, Rowe L (2002) Antagonistic coevolution between the sexes in a group of insects. Nature 415:787–789. https://doi.org/10.1038/415787a
Arnqvist G, Rowe L (2005) Sexual conflict. Princeton University Press, Princeton, NJ
Baker J, Liu J-P, Robertson EJ, Efstratiadis A (1993) Role of insulin-like growth factors in embryonic and postnatal growth. Cell 75:73–82. https://doi.org/10.1016/S0092-8674(05)80085-6
Balshine S (2012) Patterns of parental care in vertebrates. In: Royle J, Smiseth PT, Kölliker M (eds) The evolution of parental care. Oxford University Press, Oxford, pp 62–80
Barlow DP, Stöger R, Herrmann BG, Saito K, Schweifer N (1991) The mouse insulin-like growth factor type-2 receptor is imprinted and closely linked to the Tme locus. Nature 349:84–87. https://doi.org/10.1038/349084a0
Beck CW (1998) Mode of fertilization and parental care in anurans. Dev Biol 55:439–449
Benun Sutton F, Wilson AB (2019) Where are all the moms? External fertilization predicts the rise of male parental care in bony fishes. Evolution 73(12):2451–2460
Bercu BB, Hyashi A, Poth M, Alexandrova M, Solof MS, Donahoe PK (1980) Luteinizing hormone-releasing hormone-induced delay of parturition. Endocrinology 107:504–508. https://doi.org/10.1210/endo-107-2-504
Blacher P, Huggins TJ, Bourke AFG (2017) Evolution of ageing, costs of reproduction and the fecundity – longevity trade-off in eusocial insects. Proc R Soc B 284:20170380. https://doi.org/10.1098/rspb.2017.0380
Blackburn DG (1992) Convergent evolution of viviparity, matrotrophy, and specializations for fetal nutrition in reptiles and other vertebrates. Am Zool 32:313–321
Blackburn DG (1994) Standardized criteria for the recognition of embryonic nutritional patterns in squamate reptiles. Copeia 1994:925–935
Blackburn DG (1995) Saltationist and Punctuated Equilibrium Models for the evolution of viviparity and placentation. J Theor Biol 174:199–216
Blackburn DG (1998) Structure, function, and evolution of the oviducts of squamate reptiles, with special reference to viviparity and placentation. J Exp Zool 282:560–617. https://doi.org/10.1002/(Sici)1097-010x(199811/12)282:4/5%3c560::Aid-Jez10%3e3.3.Co;2-A
Blackburn DG (1999) Viviparity and oviparity: Evolution and reproductive strategies. In: Knobil TE, Neill JD (eds) Encyclopedia of Reproduction. Academic Press, New York, pp 994–1003
Blackburn DG (2000) Classification of the reproductive patterns of amniotes. Herpetol Monogr 14:371–377
Blackburn DG (2015) Viviparous placentotrophy in reptiles and the parent-offspring conflict. J Exp Zool B 324:532–548. https://doi.org/10.1002/jez.b.22624
Blackburn DG (2015) Evolution of vertebrate viviparity and specializations for fetal nutrition: A quantitative and qualitative analysis. J Morphol 276:961–990. https://doi.org/10.1002/jmor.20272
Blackburn DG, Flemming AF (2009) Morphology, development, and evolution of fetal membranes and placentation in squamate reptiles. J Exp Zool B 312:579–589. https://doi.org/10.1002/jez.b.21234
Blackburn DG, Evans HE, Vitt LJ (1985) The evolution of fetal nutritional adaptations. Fortschr Zool 30:437–439
Bowman CE, Arany Z, Wolfgang MJ (2021) Regulation of maternal–fetal metabolic communication. Cell Mol Life Sci 78:1455–1486
Buckley D, Alcobendas M, García-París M, Wake MH (2007) Heterochrony, cannibalism, and the evolution of viviparity in Salamandra salamandra. Evol Dev 9:105–115. https://doi.org/10.1111/j.1525-142X.2006.00141.x
Buckley D (2012) Evolution of viviparity in salamanders (Amphibia, Caudata). In: eLS , pp 1–13. https://doi.org/10.1002/9780470015902.a0022851
Buckling A, Rainey PB (2002) Antagonistic coevolution between a bacterium and a bacteriophage. Proc R Soc Lond B 269:931–936. https://doi.org/10.1098/rspb.2001.1945
Burley N (1981) Sex ratio manipulation and selection for attractiveness. Science 211:721–722
Burt A, Trivers R (1998) Genetic conflicts in genomic imprinting. Proc R Soc Lond B 265:2393–2397
Callard IP, Fileti LA, Perez LE, Sorbera LA, Giannoukos G, Klosterman LL, Tsang P, McCracken JA (1992) Role of the corpus luteum and progesterone in the evolution of vertebrate viviparity. Integr Comp Biol 32:264–275. https://doi.org/10.1093/icb/32.2.264
Chapman T, Arnqvist G, Bangham J, Rowe L (2003) Sexual conflict. Trends Ecol Evol 18:41–48
Charalambous M, Smith FM, Bennett WR, Crew TE, Mackenzie F, Ward A (2003) Disruption of the imprinted Grb10 gene leads to disproportionate overgrowth by an Igf2-independent mechanism. P Natl Acad Sci USA 100:8292–8297. https://doi.org/10.1073/pnas.1532175100
Chiba Y, He B, Yoshizaki K et al (2019) The transcription factor AmeloD stimulates epithelial cell motility essential for tooth morphology. J Biol Chem 294:3406–3418. https://doi.org/10.1074/jbc.RA118.005298
Chuong EB, Tong W, Hoekstra HE (2010) Maternal-fetal conflict: rapidly evolving proteins in the rodent placenta. Mol Biol Evol 27:1221–1225. https://doi.org/10.1093/molbev/msq034
Cohick WS, Clemmons DR (1993) The insulin-like growth factors. Annu Rev Physiol 55:131–153
Compagno LVJ (2001) Sharks of the world. An annotated and illustrated catalogue of shark species known to date, vol. 2. Bullhead, mackerel and carpet sharks (Heterodontiformes, Lamniformes and Orectolobiformes). FAO species catalogue for fishery purposes no 1, vol. 2, Rom
Constância M, Hemberger M, Hughes J et al (2002) Placental-specific IGF-II is a major modulator of placental and fetal growth. Nature 417:945–948. https://doi.org/10.1038/nature00819
Costa WJEM, Amorim PF, Mattos JLO (2016) Molecular phylogeny and evolution of internal fertilization in South American seasonal cynopoeciline killifishes. Mol Phylogenet Evol 95:94–99. https://doi.org/10.1016/j.ympev.2015.11.011
Crespi B, Semeniuk C (2004) Parent-offspring conflict in the evolution of vertebrate reproductive mode. Am Nat 163:635–653. https://doi.org/10.1086/382734
Daneshfozouna H, Panjvinib F, Ghorbanic F, Farahmandd H (2015) A review of epigenetic imprints in aquatic animals. Fish Aquac J 6:1–3. https://doi.org/10.4172/2150-3508.1000119
Dawkins R, Carlise TR (1976) Parental investment, mate desertion and fallacy. Nature 262:131–133
DeChiara TM, Efstratiadis A, Robertson EJ (1990) A growth-deficiency phenotype in heterozygous mice carrying an insulin-like growth factor II gene disrupted by targeting. Nature 345:78–80
DeChiara TM, Robertson EJ, Efstratiadis A (1991) Parental imprinting of the mouse insulin-like growth factor II gene. Cell 64:849–859. https://doi.org/10.1016/0092-8674(91)90513-X
Dopazo H, Alberch P (1994) Preliminary results on optional viviparity and intrauterine siblicide in Salamandra salamandra populations from Northern Spain. Mertensiella 4:125–137
Duvillié B, Cordonnier N, Deltour L, Dandoy-Dron F, Itier JM, Monthioux E, Jami J, Joshi RL, Bucchuni D (1997) Phenotypic alterations in insulin-deficient mutant mice. P Natl Acad Sci USA 94:5137–5140. https://doi.org/10.1073/pnas.94.10.5137
Duvillié B, Bucchini D, Tang T, Jami J, Pàldi A (1998) Imprinting at the mouse Ins2 locus: evidence for cis- and trans-allelic interactions. Genomics 47:52–57. https://doi.org/10.1006/geno.1997.5070
Einum S, Fleming IA (2000) Highly fecund mothers sacrifice offspring survival to maximize fitness. Nature 405:565–567. https://doi.org/10.1038/35014600
Faber JJ, Thornburg KL, Binder ND (1992) Physiology of placental transfer in mammals. Am Zool 32:343–354
Fowden AL, Sibley C, Reik W, Constancia M (2006) Imprinted genes, placental development and fetal growth. Horm Res 65:50–58. https://doi.org/10.1159/000091506
Frank D, Fortino W, Clark L, Musalo R, Wang W, Saxena A, Li C-M, Reik W, Ludwig T, Tycko B (2002) Placental overgrowth in mice lacking the imprinted gene Ipl. P Natl Acad Sci USA 99:7490–7495. https://doi.org/10.1073/pnas.122039999
Gabory A, Ripoche MA, Le Digarcher A et al (2009) H19 acts as a trans regulator of the imprinted gene network controlling growth in mice. Development 136:3413–3421. https://doi.org/10.1242/dev.036061
Geist KS, Strassmann JE, Queller DC (2019) Family quarrels in seeds and rapid adaptive evolution in Arabidopsis. P Natl Acad Sci USA 116:9463–9468. https://doi.org/10.1073/pnas.1817733116
Ghalambor CK, Reznick DN, Walker JA (2004) Constraints on adaptive evolution: the functional trade-off between reproduction and fast-start swimming performance in the Trinidadian guppy (Poecilia reticulata). Am Nat 164:38–50. https://doi.org/10.1086/421412
Giddings SJ, King CD, Harman KW, Flood JF, Carnaghi LR (1994) Allele specific inactivation of insulin 1 and 2, in the mouse yolk sac, indicates imprinting. Nat Genet 6:310–313. https://doi.org/10.1038/ng0394-310
Gil D, Graves J, Hazon N, Wells A (1999) Male attractiveness and differential testosterone investment in zebra finch eggs. Science 286:126–128. https://doi.org/10.1126/science.286.5437.126
Gilmore RG, Dodrill JW, Linley PA (1983) Reproduction and embryonic development of the sand tiger shark Odontaspis taurus (Refinesque). Fish Bull 81:201–226
Gilmore RG, Putz O, Dodrill JW (2005) Oophagy, intrauterine cannibalism and reproductive strategy in lamnoid sharks. In: Hamlett WC (ed) Reproductive Biology and Phylogeny of Chondrichthyes: Sharks, batoids and chimaeras. Science Publishers, Enfield, NH, pp 435–462
Godfray HCT (1995) Signaling of need between parents and young: parent-offspring conflict and sibling rivalry. Am Nat 146:1–24
Greven H, Grossherr M (1992) Adelphophagy and oophagy in Ameca splendens Miller and Fitzsimons, 1971 (Goodeidae, Teleostei). Z Fischkd 12:193–197
Griffith OW, Wagner GP (2017) The placenta as a model for understanding the origin and evolution of vertebrate organs. Nat Ecol Evol 1:0072. https://doi.org/10.1038/s41559-017-0072
Griffith OW, Brandley MC, Belov K, Thompson MB (2016) Allelic expression of mammalian imprinted genes in a matrotrophic lizard, Pseudemoia entrecasteauxii. Dev Genes Evol 226:79–85. https://doi.org/10.1007/s00427-016-0531-x
Griffith OW, Brandley MC, Whittington CM, Belov K, Thompson MB (2017) Comparative genomics of hormonal signaling in the chorioallantoic membrane of oviparous and viviparous amniotes. Gen Comp Endocrinol 244:19–29. https://doi.org/10.1016/j.ygcen.2016.04.017
Gross MT, Sargent RC (1985) The evolution of male and female parental care in fishes. Am Zool 25:807–822
Guex GD, Chen PS (1986) Epitheliophagy: Intrauterine cell nourishment in the viviparous alpine salamander, Salamandra atra (Laur.). Experientia 42:1205–1218. https://doi.org/10.1007/BF01946392
Guillette LJ, Jones RE (1982) Further observations on arginine vasotocin-induced oviposition and parturition in lizards. J Herpetol 16:140–144
Guillette J, Louis J, DeMarco V, Palmer BD (1991) Exogenous progesterone or indomethacin delays parturition in the viviparous lizard Sceloporus jarrovi. Gen Comp Endocrinol 81:105–112
Guillette LJ, Propper CR, Cree A, Doresl RM (1991) Endocrinology of oviposition in the tuatara (Sphenodon punctatus)—II. Plasma arginine vasotocin concentrations during natural nesting. Comp Biochem Physiol A 100:819–822. https://doi.org/10.1016/0300-9629(91)90298-Q
Hagmayer A, Furness AI, Reznick DN, Dekker ML, Pollux BJA (2020) Predation risk shapes the degree of placentation in natural populations of live-bearing fish. Ecol Lett 23:831–840. https://doi.org/10.1111/ele.13487
Haig D (1993) Genetic conflicts in human pregnancy. Q Rev Biol 68:495–532
Haig D (1996) Placental hormones, genomic imprinting, and maternal-fetal communication. J Evol Biol 9:357–380. https://doi.org/10.1046/j.1420-9101.1996.9030357.x
Haig D (2000) The kinship theory of genomic imprinting. Annu Rev Ecol Syst 31:9–32
Haig D (2004) Genomic imprinting and kinship: how good is the evidence? Annu Rev Genet 38:553–585. https://doi.org/10.1146/annurev.genet.37.110801.142741
Haig D, Westoby M (1989) Parent-specific gene expression and the triploid endosperm. Am Nat 134:147–155. https://doi.org/10.1086/284971
Haig D, Westoby M (1991) Genomic imprinting in endosperm - its effect on seed development in crosses between species, and between different ploidies of the same species, and its implications for the evolution of apomixis. Phil Trans R Soc B 333:1–13. https://doi.org/10.1098/rstb.1991.0057
Hamlett WC (1999) Placenta and placental analogues in elasmobranchs. In: Knobil TE, Neill JD (eds) Encyclopedia of Reproduction. Academic Press, New York, pp 197–206
Hamlett WC, Hysell MK (1998) Uterine specializations in elasmobranchs. J Exp Zool 282:438–459
Hamlett WC, Kormanik G, Storrie M, Stevens B, Walker TI (2005) Chondrichthyan parity, lecithotrophy and matrotrophy. In: Hamlett WC (ed) Reproductive Biology and Phylogeny of Chondrichthyes: Sharks, Batoids and Chimaeras. Science Publishers, Enfield, NH, pp 395–434
Harrison A (2013) Size-assortative pairing and social monogamy in a neotropical lizard, Anolis limifrons (Squamata: Polychrotidae). Breviora 534:1–9. https://doi.org/10.3099/534.1
Hayakawa Y, Munehara H (2001) Facultatively internal fertilization and anomalous embryonic development of a non-copulatory sculpin Hemilepidotus gilberti Jordan and Starks (Scorpaeniformes: Cottidae). J Exp Mar Biol Ecol 256:51–58. https://doi.org/10.1016/S0022-0981(00)00303-8
Hayakawa Y, Munehara H (2003) Comparison of ovarian functions for keeping embryos by measurement of dissolved oxygen concentrations in ovaries of copulatory and non-copulatory oviparous fishes and viviparous fishes. J Exp Mar Biol Ecol 295:245–255. https://doi.org/10.1016/S0022-0981(03)00297-1
Helmstetter AJ, Papadopulos AST, Igea J, Van Dooren TJM, Leroi AM, Savolainen V (2016) Viviparity stimulates diversification in an order of fish. Nat Commun 7:11271. https://doi.org/10.1038/ncomms11271
Hollenberg F, Wourms JP (1994) Ultrastructure and protein uptake of the embryonic trophotaeniae of four species of goodeid fishes (Teleostei: Atheriniformes). J Morphol 219:105–129
Hollenberg F, Wourms JP (1995) Embryonic growth and maternal nutrient sources in goodeid fishes (Teleostei: Cyprinodontiformes). J Exp Zool A 271:379–394
Kahn AT, Schwanz LE, Kokko H (2013) Paternity protection can provide a kick-start for the evolution of male-only parental care. Evolution 67:2207–2217. https://doi.org/10.1111/evo.12103
Kalscheuer VM, Mariman EC, Schepens MT, Rehder H, Ropers H-H (1993) The insulin-like growth factor type-2 receptor gene is imprinted in the mouse but not in humans. Nat Genet 5:74–78
Kaneko-Ishino T, Kohda T, Ishino F (2003) The regulation and biological significance of genomic imprinting in mammals. J Biochem 133:699–711. https://doi.org/10.1093/jb/mvg090
Kaye MD, Jones WR, Anderson DT (1972) Immunology and placentation in viviparous invertebrates. J Reprod Fertil 1972:335–336. https://doi.org/10.1530/jrf.0.0310335
Kent GC (2018) Animal reproductive system. Encyclopedia Britannica, https://www.britannica.com/science/animal-reproductive-system
Klisch K, Mess A (2007) Evolutionary differentiation of cetartiodactyl placentae in the light of the viviparity-driven conflict hypothesis. Placenta 28:353–360. https://doi.org/10.1016/j.placenta.2006.03.014
Kornfeld S, Mellman I (1989) The biogenesis of lysosomes. Annu Rev Cell Dev Biol 5:483–525
Kunz Y (2004) Developmental biology of teleost fishes. Springer, Dordrecht
Kupfer A, Kramer A, Himstedt W, Greven H (2006) Copulation and egg retention in an oviparous Caecilian (Amphibia: Gymnophiona). Zool Anz 244:223–228. https://doi.org/10.1016/j.jcz.2005.12.001
Kupfer A, Müller H, Antoniazzi MM, Jared C, Greven H, Nussbaum RA, Wilkinson M (2006) Parental investment by skin feeding in a caecilian amphibian. Nature 440:926–929. https://doi.org/10.1038/nature04403
Kupfer A, Maxwell E, Reinhard S, Kuehnel S (2016) The evolution of parental investment in caecilian amphibians: a comparative approach. Biol J Linn Soc 119:4–14. https://doi.org/10.1111/bij.12805
Lau MMH, Stewart CEH, Liu Z et al (1994) Loss of the imprinted IGF2/cation-independent mannose 6-phosphate receptor results in fetal overgrowth and perinatal lethality. Genes Dev 8:2953–2963. https://doi.org/10.1101/gad.8.24.2953
Lefebvre L, Viville S, Barton SC, Ishino F, Keverne EB, Surani MA (1998) Abnormal maternal behaviour and growth retardation associated with loss of the imprinted gene Mest. Nat Genet 20:163–169. https://doi.org/10.1038/2464
Li Y, Behringer RR (1998) Esx1 is an X-chromosome-imprinted regulator of placental development and fetal growth. Nat Genet 20:309–311. https://doi.org/10.1038/3129
Li L, Keverne EB, Aparicio SA, Ishino F, Barton SC, Surani MA (1999) Regulation of maternal behavior and offspring growth by paternally expressed Peg3. Science (new York NY) 284:330–333. https://doi.org/10.1126/science.284.5412.330
Lombardi J, Wourms JP (1979) Structure, function, and evolution of trophotaeniae, placental analogues of viviparous fishes. Am Zool 19:976
Long JA, Mark-Kurik E, Johanson Z et al (2014) Copulation in antiarch placoderms and the origin of gnathostome internal fertilization. Nature 517:196–199. https://doi.org/10.1038/nature13825
Ludwig T, Eggenschwiler J, Fisher P, D’Ercole AJ, Davenport ML, Efstratiadis A (1996) Mouse mutants lacking the type 2 IGF receptor (IGF2R) are rescued from perinatal lethality in igf2 and igf1r null backgrounds. Dev Biol 535:517–535
Macías Garcia C, Ramirez E (2005) Evidence that sensory traps can evolve into honest signals. Nature 434:501–505. https://doi.org/10.1038/nature03351.1
MacKay AB, Mhanni AA, McGowan RA, Krone PH (2007) Immunological detection of changes in genomic DNA methylation during early zebrafish development. Genome 50:778–785. https://doi.org/10.1139/G07-055
Magurran AE, Seghers BH (1994) A cost of sexual harassment in the guppy, Poecilia reticulata. Proc R Soc Lond B 258:89–92
Mank JE, Promislow DEL, Avise JC (2005) Phylogenetic perspectives in the evolution of parental care in ray-finned fishes. Evolution 59:1570–1578. https://doi.org/10.1554/04-734.1.s1
Marsh-Matthews E, Deaton R (2006) Resources and offspring provisioning: A test of the Trexler-DeAngelis model for matrotrophy evolution. Ecology 87:3014–3020. https://doi.org/10.1890/0012-9658(2006)87[3014:RAOPAT]2.0.CO;2
Matoba S, Nakamuta S, Miura K, Hirose M, Shiura H, Kohda T, Nakamuta N, Ohuda A (2019) Paternal knockout of Slc38a4/SNAT4 causes placental hypoplasia associated with intrauterine growth restriction in mice. P Natl Acad Sci USA 116:21047–21053. https://doi.org/10.1073/pnas.1907884116
Matsushima N, Kawata M (2005) The choice of oviposition site and the effects of density and oviposition timing on survivorship in Rana japonica. Ecol Res 20:81–86. https://doi.org/10.1007/s11284-004-0010-0
McNamara JM, Houston AI, Barta Z, Osorno JL (2003) Should young ever be better off with one parent than with two? Behav Ecol 14:301–310. https://doi.org/10.1093/beheco/14.3.301
Meek SE (1904) The fresh-water fishes of Mexico north of the Isthmus of Tehuantepec. Field Columbian Mus Publ 93. Zool Ser 5:i–lxiii:1–252
Meisner AD, Burns JR (1997) Viviparity in the halfbeak genera Dermogenys and Nomorhamphus (Teleostei: Hemiramphidae). J Morphol 234:295–317. https://doi.org/10.1002/(SICI)1097-4687(199712)234:3%3c295::AID-JMOR7%3e3.0.CO;2-8
Mess A, Carter AM (2007) Evolution of the placenta during the early radiation of placental mammals. Comp Biochem Physiol A 148:769–779. https://doi.org/10.1016/j.cbpa.2007.01.029
Miller RR, Fitzsimons JM (1971) Ameca splendens, a new genus and species of goodeid fish from western Mexico, with remarks on the classification of the Goodeidae. Copeia 8:1–3
Montserrat M, Bas C, Magalhães S, Sabelis MW, De Roos AM, Janssen A (2007) Predators induce egg retention in prey. Oecologia 150:699–705. https://doi.org/10.1007/s00442-006-0527-8
Moore T (2012) Review: Parent-offspring conflict and the control of placental function. Placenta 33:S33–S36. https://doi.org/10.1016/j.placenta.2011.11.016
Moore T, Haig D (1991) Genomic imprinting in mammalian development: a parental tug-of-war. Trends Genet 7:45–49. https://doi.org/10.1016/0168-9525(91)90230-N
Morrison KR, Ngo V, Cardullo RA, Reznick DN (2017) How fish eggs are preadapted for the evolution of matrotrophy. Proc R Soc B 284:20171342. https://doi.org/10.1098/rspb.2017.1342
Mossman HW (1991) Comparative morphogenesis of the fetal membranes accesso and uterine structures. Placenta 12:1–5
Motz VA, Callard IP (1988) Seasonal variations in oviductal activity, extensibility, and tensile strength in Chrysemys picta. Gen Comp Endocrinol 72:453–460. https://doi.org/10.1016/0016-6480(88)90168-2
Murphy BF, Thompson MB (2011) A review of the evolution of viviparity in squamate reptiles: The past, present and future role of molecular biology and genomics. J Comp Physiol B 181:575–594. https://doi.org/10.1007/s00360-011-0584-0
Murphy WJ, Thomerson JE, Collier GE (1999) Phylogeny of the Neotropical killifish family Rivulidae (Cyprinodontiformes, Aplocheiloidei) inferred from mitochondrial DNA sequences. Mol Phylogenet Evol 13:289–301. https://doi.org/10.1006/mpev.1999.0656
O’Neill MJ, Ingram RS, Vrana PB, Tilghman SM (2000) Allelic expression of IGF2 in marsupials and birds. Dev Genes Evol 210:18–20. https://doi.org/10.1007/PL00008182
O’Neill MJ, Lawton BR, Mateos M, Carone DM, Ferreri GC, Hrbek T, Meredith RW, Reznick DN, O’Neill RJ (2007) Ancient and continuing Darwinian selection on insulin-like growth factor II in placental fishes. P Natl Acad Sci USA 104:12404–12409. https://doi.org/10.1073/pnas.0705048104
Okamura K, Hagiwara-Takeuchi Y, Li T, Vu TH, Hirai M, Hattori M, Sakaki Y, Hoffman AR, Ito T (2000) Comparative genome analysis of the mouse imprinted gene impact and its nonimprinted human homolog IMPACT: toward the structural basis for species-specific imprinting. Genome Res 10:1878–1889. https://doi.org/10.1101/gr.139200
Osorno JL, Székely T (2004) Sexual conflict and parental care in magnificent frigatebirds: Full compensation by deserted females. Dev Biol 68:337–342. https://doi.org/10.1016/j.anbehav.2003.06.027
Ostrovsky AN, Lidgard S, Gordon DP, Schwaha T, Genikhovich G, Ereskovsky AV (2016) Matrotrophy and placentation in invertebrates: a new paradigm. Biol Rev 91:673–711. https://doi.org/10.1111/brv.12189
Parker GA (1970) Sperm competition and its evolutionary consequences in the insects. Biol Rev 45:525–567. https://doi.org/10.1111/j.1469-185X.1970.tb01176.x
Parker GA (1974) Courtship persistence and female-guarding as male time investment strategies. Behaviour 48:157–184
Parker GA (1984) Sperm competition and the evolution of animal mating strategies. In: Smith RL (ed) Sperm Competition and the Evolution of Animal Mating Systems. Academic Press, London, pp 1–60
Parker GA (2006) Sexual conflict over mating and fertilization: an overview. Phil Trans R Soc B 361:235–259. https://doi.org/10.1098/rstb.2005.1785
Pearsall RS, Shibata H, Brozowska A, Yoshino K, Okuda K, DeJong PJ, Plass C, Chapman VM, Hayashizaki Y, Held WA (1996) Absence of imprinting in U2AFBPL, a human homologue of the imprinted mouse gene U2afbp-rs. Biochem Biophys Res Commun 222:171–7. S0006–291X(96)90716–0 [pii] https://doi.org/10.1006/bbrc.1996.0716
Pickford GE (1952) Induction of a spawning reflex in hypophysectomized killifish. Nature 170:807–808
Pollux BJA, Reznick DN (2011) Matrotrophy limits a female’s ability to adaptively adjust offspring size and fecundity in fluctuating environments. Funct Ecol 25:747–756. https://doi.org/10.1111/j.1365-2435.2011.01831.x
Pollux BJA, Pires MN, Banet AI, Reznick DN (2009) Evolution of placentas in the fish family Poeciliidae: an empirical study of macroevolution. Annu Rev Ecol Evol S 40:271–289. https://doi.org/10.1146/annurev.ecolsys.110308.120209
Recknagel H, Kamenos NA, Elmer KR (2021) Evolutionary origins of viviparity consistent with palaeoclimate and lineage diversification. J Evol Biol 34:1167–1176. https://doi.org/10.1111/jeb.13886
Reik W, Constância M, Fowden A, Anderson N, Dean W, Ferguson-Smith A, Tycko B, Sibley C (2003) Regulation of supply and demand for maternal nutrients in mammals by imprinted genes. J Physiol 547:35–44. https://doi.org/10.1113/jphysiol.2002.033274
Renfree MB, Suzuki S, Kaneko-Ishino T (2013) The origin and evolution of genomic imprinting and viviparity in mammals. Phil Trans R Soc B 368:20120151. https://doi.org/10.1098/rstb.2012.0151
Reznick DN, Mateos M, Springer MS (2002) independent origins and rapid fish genus Poeciliopsis. Science 298:1018–1020. https://doi.org/10.1126/science.1076018
Reznick DN, Meredith R, Collette BB (2007) Independent evolution of complex life history adaptations in two families of fishes, live-bearing halfbeaks (Zenarchopteridae, Beloniformes) and Poeciliidae (Cyprinodontiformes). Evolution 61:2570–2583. https://doi.org/10.1111/j.1558-5646.2007.00207.x
Riesch R, Plath M, Schlupp I, Tobler M, Brian Langerhans R (2014) Colonisation of toxic environments drives predictable life-history evolution in livebearing fishes (Poeciliidae). Ecol Lett 17:65–71. https://doi.org/10.1111/ele.12209
Roberts RM, Green JA, Schulz LC (2016) The evolution of the placenta. Reproduction 152:R179–R189. https://doi.org/10.1530/REP-16-0325
Rodao M, Montagne J, Clivio GA, Papa NG, Larrosa GC (2015) Sperm and egg envelope ultrastructure and some considerations on its evolutionary meaning. In: Berois N, García G, de Sá RO (eds) Annual Fishes: Life History Strategy, Diversity, and Evolution. CRC Press, Boca Raton, pp 47–62
Royle NJ, Smiseth PT, Kölliker M (eds) (2012) The evolution of parental care. Oxford University Press, Oxford
Saldivar Lemus Y, Vielle-Calzada J-P, Ritchie MG, Macías García C (2017) Asymmetric paternal effect on offspring size linked to parent-of-origin expression of an insulin-like growth factor. Ecol Evol 7:4465–4474. https://doi.org/10.1002/ece3.3025
San Mauro D, Gower DJ, Müller H, Loader SP, Zardoya R, Nussbaum RA, Wilkinson M (2014) Life-history evolution and mitogenomic phylogeny of caecilian amphibians. Mol Phylogenet Evol 73:177–189. https://doi.org/10.1016/j.ympev.2014.01.009
Savage J (2002) The amphibians and reptiles of Costa Rica: a herpetofauna between two continents, between two seas. University of Chicago Press, Chicago
Schneider JM (1999) Delayed oviposition: a female strategy to counter infanticide by males? Behav Ecol 10:567–571
Schrader M, Travis J (2008) Testing the viviparity-driven-conflict hypothesis: parent-offspring conflict and the evolution of reproductive isolation in a poeciliid fish. Am Nat 172:806–817. https://doi.org/10.1086/592999
Schwartz TS, Bronikowski AM (2016) Evolution and function of the insulin and insulin-like signaling network in ectothermic reptiles: some answers and more questions. Integr Comp Biol 56:171–184
Shibukawa K, Tran DD, Tran LX (2012) Phallostethus cuulong, a new species of priapiumfish (Actinopterygii: Atheriniformes: Phallostethidae) from the Vietnamese Mekong. Zootaxa 51:45–51
Shine R (2014) Evolution of an evolutionary hypothesis: a history of changing ideas about the adaptive significance of viviparity in reptiles. J Herpetol 48:147–161. https://doi.org/10.1670/13-075
Sibly RM, Kodric-Brown A, Luna SM, Brown JH (2018) The shark-tuna dichotomy: Why tuna lay tiny eggs but sharks produce large offspring. R Soc Open Sci 5:180453. https://doi.org/10.1098/rsos.180453
Smith KK (2015) Placental evolution in therian mammals. In: Dial KP, Shubin N, Brainerd EL (eds) Great transformations in vertebrate evolution. University of Chicago Press, Chicago, pp 205–225
Smuts BB, Smuts RW (1993) Male aggression and sexual coercion of females in nonhuman primates and other mammals: evidence and theoretical implications. Adv Stud Behav 22:1–63. https://doi.org/10.1016/S0065-3454(08)60404-0
Springer S (1948) Oviphagous embryos of the sand shark, Carcharias taurus. Copeia 1948:153–157. https://doi.org/10.2307/1438449
Stearns SC (1989) Trade-offs in life-history evolution. Funct Ecol 3:259–268
Stewart JR (1992) Placental structure and nutritional provision to embryos in predominantly lecithotrophic viviparous reptiles. Integr Comp Biol 32:303–312. https://doi.org/10.1093/icb/32.2.303
Stewart JR, Blackburn DG (1988) Reptilian placentation: structural diversity and terminology. Copeia 1988:839–852
Stewart JR, Blackburn DG (2014) Viviparity and placentation in lizards. In: Rheubert JL, Siegel DS, Trauth SE (eds) Reproductive biology and phylogeny of lizards and tuatara. CRC Press, Boca Raton, FL, pp 448–563
Stewart JR, Thompson MB (1996) Evolution of reptilian placentation: development of extraembryonic membranes of the Australian scincid lizards, Bassiana duperreyi (oviparous) and Pseudemoia entrecasteauxii (viviparous). J Morphol 227:349–370. https://doi.org/10.1002/(sici)1097-4687(199603)227:3%3c349::aid-jmor6%3e3.0.co;2-0
Stewart JR, Thompson MB (2000) Evolution of placentation among squamate reptiles: recent research and future directions. Comp Biochem Physiol A 127:411–431. https://doi.org/10.1016/S1095-6433(00)00273-7
Stockley P, Gage MJT, Parker GA, Møller AP (1997) Sperm competition in fishes: the evolution of testis size and ejaculate characteristics. Am Nat 149:933–954
Tan HH, Lim KKP (2013) Three new species of freshwater halfbeaks (Teleostei: Zenarchopteridae: Hemirahamphodon) from Borneo. Raffles Bull Zool 61:735–747
Tarwater CE, Arcese P (2017) Young females pay higher costs of reproduction in a short-lived bird. Behav Ecol Sociobiol 71:84. https://doi.org/10.1007/s00265-017-2309-1
Thiem L (2020) Hold on for one more day: oviductal egg retention as a mechanism for flexible reproductive phenology in eastern musk turtles (Sternotherus odoratus). MSc thesis, Austin Peay State University
Thompson MB, Speake BK (2006) A review of the evolution of viviparity in lizards: structure, function and physiology of the placenta. J Comp Physiol B 176:179–189. https://doi.org/10.1007/s00360-005-0048-5
Thompson MB, Adams SM, Herbert JF, Biazik JM, Murphy CR (2004) Placental function in lizards. Int Congr Ser 1275:218–225. https://doi.org/10.1016/j.ics.2004.08.055
Tilghman SM (1999) The sins of the fathers and mothers: Genomic imprinting in mammalian development. Cell 96:185–193. https://doi.org/10.1016/S0092-8674(00)80559-0
Trivers RL (1972) Parental investment and sexual selection. In: Campbell B (ed) Selection and the Descent of Man 1871–1971. Aldine de Gruyter, New York, pp 134–179
Trivers RL (1974) Parent-Offspring Conflict Am Zool 14:249–264. https://doi.org/10.1093/icb/14.1.249
Trivers RL (1985) Social evolution. Cummings Publishing Co., Menlo Park, CA
Turner CL (1936) The absorptive processes in the embryos of Parabrotula dentiens, a viviparous, deep-sea Brotulid fish. J Morphol 59:313–325. https://doi.org/10.1002/jmor.1050590205
Turner CL (1940) Pseudoamnion, pseudochorion, and follicular pseudoplacenta in poeciliid fishes. J Morphol 67:59–89
Tycko B, Morison IM (2002) Physiological functions of imprinted genes. J Cell Physiol 192:245–258. https://doi.org/10.1002/jcp.10129
Uller T, Olsson M (2008) Multiple paternity in reptiles: patterns and processes. Mol Ecol 17:2566–2580. https://doi.org/10.1111/j.1365-294X.2008.03772.x
Uribe MC, García Alarcón A (2005) The ovary of viviparous teleosts. Morphological differences between the ovaries of Goodea atripinnis and Ilydon whitei (Goodeidae). In: Uribe MC, Grier HJ (eds) Viviparous fishes. New Life Publication, Homestead, FL, pp 217–235
Wake MH (1977) The reproductive biology of caecilians: an evolutionary perspective. In: Taylor DH, Guttman SI (eds) The reproductive biology of amphibians. Plenum, New York, pp 73–101
Wake MH (1977) Fetal maintenance and its evolutionary significance in the Amphibia: Gymnophiona. J Herpetol 11:379–386
Wake MH (1980) The reproductive biology of Nectophrynoides malcolmi (Amphibia: Bufonidae), with comments on the evolution of reproductive modes in the genus Nectophrynoides. Copeia 1980:193–209
Wake MH (1992) Evolutionary scenarios, homology and convergence of structural specializations for vertebrate viviparity. Integr Comp Biol 32:256–263. https://doi.org/10.1093/icb/32.2.256
Wake MH (2015) Fetal adaptations for viviparity in amphibians. J Morphol 276:941–960. https://doi.org/10.1002/jmor.20271
Wake MH, Hanken J (1982) Development of the skull of Dermophis mexicanus (amphibia: Gymnophiona), with comments on skull kinesis and amphibian relationships. J Morphol 173:203–223. https://doi.org/10.1002/jmor.1051730208
Wang S, Yuen SSF, Randall DJ, Hung CY, Tsui TKN, Poon WL, Lai JCC, Zhang Y, Lin H (2008) Hypoxia inhibits fish spawning via LH-dependent final oocyte maturation. Comp Biochem Physiol C 148:363–369. https://doi.org/10.1016/j.cbpc.2008.03.014
Whittington CM, van Dyke JU, Liang SQT, Edwards SV, Shine R, Thompson MB, Grueber CE (2022) Understanding the evolution of viviparity using intraspecific variation in reproductive mode and transitional forms of pregnancy. Biol Rev (published online, https://doi.org/10.1111/brv.12836)
Wiklund C, Fagerström T (1977) Why do males emerge before females? Oecologia 31:153–158. https://doi.org/10.1007/BF00346917
Wildman DE, Chen C, Grossman LI, Goodman M, Romero R (2006) Evolution of the mammalian placenta revealed by phylogenetic analysis. P Natl Acad Sci USA 103:3203–3208
Wilkins JF, Haig D (2003) What good is genomic imprinting: the function of parent-specific gene expression. Nat Rev Genet 4:359–368. https://doi.org/10.1038/nrg1062
Wooding P, Burton G (2008) Comparative placentation: structures, functions and evolution. Springer, Berlin
Wourms JP (1977) Reproduction and development in chondrichthyan fishes. Am Zool 17:379–410
Wourms JP (1981) Viviparity: The maternal-fetal relationship in fishes. Integr Comp Biol 21:473–515. https://doi.org/10.1093/icb/21.2.473
Wourms JP, Lombardi J (1992) Reflections on the evolution of piscine viviparity. Integr Comp Biol 32:276–293. https://doi.org/10.1093/icb/32.2.276
Wourms JP, Grove B, Lombardi J (1988) The maternal-embryonic relationship in viviparous fishes. In: Hoar W, Randall D (eds) Fish Physiology, vol 11B. Academic Press, New York, pp 1–134
Xavier F (1973) Le cycle des voies génitales femelles de Nectophrynoides occidentalis Angel, Amphibien Anoure vivipare. Z Zellforsch 140:509–534. https://doi.org/10.1007/BF00306677
Xie B, Zhang L, Zheng K, Luo C (2009) The evolutionary foundation of genomic imprinting in lower vertebrates. Chin Sci Bull 54:1354–1360. https://doi.org/10.1007/s11434-009-0149-8
Yokoe M, Takayama-Watanabe E, Saito Y, Kutsuzawa M, Fujita K, Ochi H, Nakauchi Y, Watanabe A (2016) A novel cysteine knot protein for enhancing sperm motility that might facilitate the evolution of internal fertilization in amphibians. PLoS ONE 11:e0160445. https://doi.org/10.1371/journal.pone.0160445
Yu S, Yu D, Lee E, Eckhaus M, Lee R, Corria Z, Accili D, Westphal H, Weisntein LS (1998) Variable and tissue-specific hormone resistance in heterotrimeric Gs protein α-subunit (Gsα) knockout mice is due to tissue-specific imprinting of the Gsα gene. Genetics 95:8715–8720. https://doi.org/10.1073/pnas.95.15.8715
Yu S, Gavrilova O, Chen H, Lee R, Liu J, Pacak K, Parlow AF, Quon M, Reitman ML, Weinstein LS (2000) Paternal versus maternal transmission of a stimulatory G-protein α subunit knockout produces opposite effects on energy metabolism. J Clin Invest 105:615–623. https://doi.org/10.1172/JCI8437
Yu S, Castle A, Chen M, Lee R, Takeda K, Weinstein LS (2001) Increased insulin sensitivity in Gsα knockout mice. J Biol Chem 276:19994–19998. https://doi.org/10.1074/jbc.M010313200
Zeh DW, Zeh JA (2000) Reproductive mode and speciation: the viviparity-driven conflict hypothesis. BioEssays 22:938–946. https://doi.org/10.1002/1521-1878(200010)22:10%3c938::AID-BIES9%3e3.0.CO;2-9
Zúñiga-Vega JJ, Fuentes-G JA, Ossip-Drahos AG, Martins EP (2016) Repeated evolution of viviparity in phrynosomatid lizards constrained interspecific diversification in some life-history traits. Biol Lett 12:20160653. https://doi.org/10.1098/rsbl.2016.0653
Zwoinska M, Lind MI, Maklakov AA (2014) Sexual conflict: Male control of female longevity. Curr Biol 24:R196–R198. https://doi.org/10.1016/j.cub.2014.01.044
Acknowledgements
We thank Michael G. Ritchie for providing the parenthetical notation of the phylogeny of goodeids and for his comments on an earlier version of the manuscript, and Marcela Méndez-Janovitz and Nadia Neri-Vera for their guide during the construction of phylogenetic trees. YSL is grateful to the Doctorado en Ciencias Biomédicas (UNAM) and to CONACyT for providing a PhD scholarship (No. 46568). This manuscript was much improved thanks to the comments of two referees.
Funding
CONACyT provided a PhD scholarship (No. 46568) for YS-L.
Author information
Authors and Affiliations
Contributions
Authors contributed equally to this work. YS-L conceived the review, conducted the literature search, and generated a first draft that was subsequently reworked jointly by both YS-L and CMG.
Corresponding author
Ethics declarations
Ethical approval
This is not an empirical study, but a review, and no animals or humans were used to gather any data.
Consent for publication
All authors declare their consent for publication of this review.
Conflicts of interest
The authors declare that there is no conflict of interest.
Additional information
Communicated by F. Trillmich.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Saldívar-Lemus, Y., Macías Garcia, C. Conflict and the evolution of viviparity in vertebrates. Behav Ecol Sociobiol 76, 68 (2022). https://doi.org/10.1007/s00265-022-03171-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-022-03171-z