
Abstract
One proposed benefit for the seemingly costly behaviour of food calling is the recruitment of social allies and mates by the signaller. In chimpanzees, food calls are only produced for approximately half of all feeding events. Therefore, we investigated the influence of social and ecological context on the probability of making a food call upon arriving to a food patch in a group of wild chimpanzees. First, we tested whether feeding events where food calls had been uttered did in fact attract more individuals to join the caller. Secondly, we examined the influence of two sources of audience effects: those who were physically present with the caller and those who were presumed nearby but out of sight, and thirdly the effect of various ecological factors. We found that when feeding on fruit species, events where food calls had been produced had a higher probability of group mates arriving, even whilst controlling for the effect of pant hoots. Furthermore, the probability of uttering a food call was motivated by social more than ecological context. Specifically, high-ranking males were more likely to make food calls when estrous females were nearby, while low-ranking males and females generally called more when more females were nearby, irrespective of their reproductive state. These effects were independent of the increase in food call probability when male callers were accompanied by more males. Our findings support the recruitment function of food calls and suggest that high-ranking males call to attract estrous females to food patches to obtain mating opportunities.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Vocalizations uttered by animals while foraging or feeding are known as food-associated calls and are found in a multitude of species across birds and mammals (Clay et al. 2012). Often, vocalizations considered food-specific are found to be produced in other contexts as well. For example, a food-associated call may generally signal excitement when encountering something pleasant in the environment and therefore can be produced in many contexts (Dittus 1984; Clay et al. 2012). Regardless, even these food-associated calls act as if they are referential to listeners, and many studies report that food-associated calls attract conspecifics and trigger appropriate feeding behaviour (birds: Marler et al. 1986; Elgar 1986; Heinrich and Marzluff 1991; bats: Wilkinson and Boughman 1998; dolphins: Janik 2000; primates: Goodall 1986; Chapman and Lefebvre 1990; Caine et al. 1995; Di Bitetti 2003; Slocombe and Zuberbühler 2005; Kalan et al. 2015). However, from a signaller’s perspective, it remains puzzling why one should produce food-associated calls, hereafter ‘food calls’, which attract other individuals to a food patch whereby the larger group size results in increased feeding competition (Chapman et al. 1995; Sterck et al. 1997). In fact, there are multiple reasons why attracting group mates to a food patch can confer advantages to the caller and offset the costs of feeding competition, namely protection from predators (Elgar 1986), resource or territory defence (Sterck et al. 1997; Wilkinson and Boughman 1998) or attracting mates and allies (Stokes and Williams 1972; Mitani and Nishida 1993). For other species, it has been shown that signallers produce food calls to primarily benefit their kin (Hauser and Marler 1993a). Additionally, food calls may be a cooperative form of signalling where benefits are derived to the caller via a reciprocal exchange of information about food sources (Trivers 1971). There is also some evidence to suggest that animals who fail to produce food calls that alert group members may suffer punishment. A study of rhesus macaques found that individuals who failed to call upon discovering food suffered increased rates of aggression from conspecifics (Hauser and Marler 1993b). When animals obtain a fitness benefit by calling to attract group mates, this is referred to as recruitment (Heinrich and Marzluff 1991). To date, only a few studies have directly tested for the attraction ability or recruitment potential of food calls (Elgar 1986; Chapman and Lefebvre 1990; Heinrich and Marzluff 1991; Wilkinson and Boughman 1998).
Although food calls are generally observed to attract others to a food patch (Clay et al. 2012) whether their primary function is to recruit individuals or if this happens as an accidental by-product via information parasitism by receivers still remains a controversial point (Heinrich and Marzluff 1991; Janik 2000). In other words, if signals are not given intentionally, receivers could still learn to associate food calls with the presence of food and therefore exploit this information and join callers. However, this becomes much more complex if signallers have some degree of control over call production. For example, in chimpanzees, there is surmounting evidence for audience effects in their communication whereby callers produce vocalizations selectively and direct them towards specific individuals (Mitani and Nishida 1993; Slocombe and Zuberbühler 2007; Townsend et al. 2008; Crockford et al. 2012; Schel et al. 2013). With regards to food calls, chimpanzees only utter them sometimes while feeding (on average 40–60 % of all feeding events; Slocombe et al. 2010; Kalan et al. 2015); therefore, there must be something else signallers are taking into account other than simply the presence of food. A recent playback study by Schel et al. (2013) showed that food call production was influenced by the presence of close social partners and dominant individuals. Additional chimpanzee research showed how food calls made during a feeding event could function to maintain proximity to social allies and to prolong feeding activity (Slocombe et al. 2010; Fedurek and Slocombe 2013). Together, these studies demonstrate that food call production by chimpanzees is not a hardwired response to food but instead appears to be socially motivated towards particular individuals. However, it remains to be tested whether food calls function to recruit other chimpanzees to the food patch as has been suggested repeatedly by many researchers studying captive and wild chimpanzees (Marler and Tenaza 1977; Wrangham 1977; Goodall 1986; Hauser and Wrangham 1987; Slocombe and Zuberbühler 2005, 2006). The only evidence for recruitment using food calls has been demonstrated in spider monkeys (Chapman and Lefebvre 1990), another primate species besides chimpanzees and bonobos to exhibit a fission-fusion social system. Spider monkeys producing food calls, or ‘whinny’ vocalizations, were observed to attract conspecifics to feeding trees (Chapman and Lefebvre 1990), and the call has been shown to be used to stay in contact with subgroups nearby (Ramos-Fernández 2005).
There is evidence from captivity and the wild that the acoustic variation present in chimpanzee food calls is modified with respect to the preference or value of the food item (Slocombe and Zuberbühler 2005, 2006) and patch size (Kalan et al. 2015). Early experiments also demonstrated that food call rate increased with the quantity of food presented (Hauser and Wrangham 1987; Hauser et al. 1993). Wrangham (1977) originally proposed that long-distance pant hoot vocalizations might be used by male chimpanzees to attract estrous females to feeding trees in order to gain mating opportunities. There is some evidence that these pant hoot vocalizations could indicate male status (Clark and Wrangham 1994), but pant hoots are mostly produced during travel, not feeding contexts (Fedurek et al. 2014), which suggests that their primary function is likely to regulate group coordination (Mitani and Nishida 1993). Therefore, to address Wrangham’s (1977) original hypothesis, it may be more appropriate to focus on chimpanzee food calls, also known as ‘rough grunts’, which grade from soft grunts to loud barks and are specifically produced by chimpanzees only in a foraging or feeding context (Goodall 1986; Slocombe and Zuberbühler 2006).
Chimpanzees have an extensive repertoire of gestures (Hobaiter and Byrne 2011) and facial expressions (Goodall 1986) to communicate during face-to-face interactions; therefore, it is essential to investigate whether vocalizations might be targeted at individuals that are not in the immediate presence of the caller, especially since chimpanzee food calls can be heard outside the distance of a cohesive party (Kalan et al. 2015). Thus, the aim of this study was to determine what motivates a chimpanzee to initially emit a food call or not when arriving to a food patch by investigating the surrounding social and ecological context. Firstly, we tested whether there is support for the ‘recruitment hypothesis’ by examining whether food calls do indeed attract other chimpanzees to a food patch. Secondly, if food calls are produced by signallers with the purpose to attract others, we expected chimpanzees to be selective about for whom they produce calls and thus to exhibit particular audience effects (Zuberbühler 2008). Inspired by the seminal studies on primate vocalizations by Mitani and Nishida (1993) and Chapman and Lefebvre (1990), we looked at two potential sources of audience effects: those individuals who were accompanying the signaller (i.e., physically present) and those individuals who were not in visual contact with the signaller but crucially could be believed to be nearby and within earshot when the signaller arrives at a food patch. Importantly, we tested for both types of audience effects simultaneously since the presence of some individuals has already been shown to be an important factor for food calls (Slocombe et al. 2010; Fedurek and Slocombe 2013) in order to elucidate which effects may be more important in driving call production. Additionally, due to our previous results where the acoustic structure of food calls varied according to patch size (Kalan et al. 2015), as well as the experimental studies mentioned above, we tested ecological variables for their potential influence on call production. Specifically, we predicted that food call production would depend on the time into the season since the motivation to inform and communicate with group mates may dissipate once all individuals are aware of trees bearing edible fruits. Hence, along with predictors of tree productivity and fruit availability, we included a proxy for time in fruiting season.
Materials and methods
Study subjects
Data were collected in Taï National Park, Côte d’Ivoire (5° 45 N, 7° 07 W; see Boesch and Boesch-Achermann 2000 for details of study site), from July 2011 through May 2012. All day focal follows were conducted for five male and four female chimpanzees of the habituated south group. This group comprised a total of 19 individuals with five dependent offspring at the end of the study period. All focal individuals were 15 years of age or older, except for two sub-adult males. Due to the small number of males in the community, sub-adult males were included who were the third and fourth ranked males of the group by the end of the study period (Table 1). Hence, they held important social roles within the group and were physically comparable in their development to the other males, of which none were over the age of 18. Dominance ranks of all individuals were determined by observing the directionality of pant-grunts, a greeting vocalization that is produced solely up the hierarchy which is a standard method for assessing dominance relationships of chimpanzees (Wittig and Boesch 2003). Amongst females, pant-grunts were rare, so we used additional observations on displacements at feeding sites and agonistic interactions to assign ranks (Wittig and Boesch 2003). It was not clear between two females who had the higher rank during the study period; thus to be conservative, we gave them the same rank. Males, unlike females, changed ranks during the study period (Table 1).
Data collection
Continuous behavioural and vocalization data were collected for a focal individual during dawn to dusk focal follows (average duration ± standard error 8.88 ± 2.9 h). A feeding event started when a focal individual began eating or collecting food items and lasted until the focal ceased to feed. If the focal was observed to resume feeding at the same patch, we recorded this as a continuation of the initial feeding event. During feeding events, it was noted whether pant hoots were produced by any other chimpanzee present. Every feeding event was given a unique ID, and the species eaten was noted. Only complete feeding events where we had continuous data from the arrival of the focal to the end of the feeding event were used in this analysis. Meat, insects, terrestrial herbaceous vegetation (THV) and other food items such as mushrooms were not included in this analysis since they are not easily confined to a particular location or patch. Thus, only feeding events for fruit, nuts or leaves were used since their availability was tree-dependent, and therefore, a single food patch could be easily identified.
To measure tree productivity, AK, or a field assistant, documented tree diameter at breast height (dbh; Chapman et al. 1992) during feeding events for any tree with a dbh > 20 cm. For fruits that were eaten on the ground by chimpanzees (seven species), AK observed what types of fruits chimpanzees rejected to determine which fruits on the ground would be readily eaten by chimpanzees (e.g., chimpanzees ignore rotting fruits). AK then counted the number of edible fruits in a 1-m strip from the base of the fruit tree and then multiplied this by the patch area under the tree. For all other species that were fed on while being in the tree, AK estimated a fruit count in one of two ways, but consistently in the same manner for any particular species, in order to standardize counts relative to one another, using the maximum and minimum count observed for a species. Either the number of fruits eaten by the focal individual within 1 min was counted, two separate times within the first 10 min of feeding, and then averaged to get an intake rate. For other species where it was possible to see the number of fruits in the tree crown using binoculars, AK estimated a minimum and maximum count for ripe fruits visible in the tree crown, and an average between the two values was taken. Moran’s I, a measurement of spatial aggregation or distribution, had been calculated for the fruit tree species in the south group territory in a previous study (Wittiger and Boesch 2013) and therefore were readily available. To obtain a proxy for time in fruiting season, we checked all feeding observations of the group for any given species and designated the first feeding event as day zero. From this reference point, the number of days were counted to determine at what time in the season subsequent feeding events had occurred for that species.
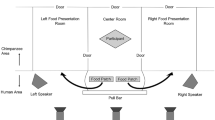
Throughout the day, party composition was monitored continuously, and the times of arrivals of other individuals to the food patch were noted. A party included any individual travelling independently (>5 years old) and within visual distance of the focal, and thus the observer. Visual detection distance is at most 25–50 m in the Taï forest. We also noted whether the focal’s party was joining others or if others had arrived to join the focal’s party. Additionally, we noted down whether the food patch was empty of chimpanzees upon our arrival or if other individuals were already present. If other individuals were present at the patch, they were from then on considered as part of the focal’s party since they were in visual contact with one other.
Since we used natural observations in this study and not experimental manipulation, we noted whether vocalizations were emitted by a focal chimpanzee within the first minute of arrival at a food patch. This is important for two reasons: (1) As a feeding event progresses, party composition changes and social interactions take place and other vocalizations are produced, thereby making it difficult to determine what may have driven call production, but calls uttered upon arrival are particularly likely to be triggered by the initial social and ecological environment; (2) chimpanzees are known to chorus or otherwise vocalize together back and forth in a way that strengthens already established bonds (Fedurek et al. 2013). With respect to food calls and pant hoots, this is more likely to occur after the first vocalizations have been elicited at a food patch or when other chimpanzees arrive (personal observation); thus, to control for confounds of these additional sources of arousal, we isolated the first minute upon arrival.
Definitions of audience members
Studies of chimpanzee vocalizations have consistently shown that behaviour is influenced by who may be nearby, or within auditory distance of the signaller, and not only by who may be in visual contact (Goodall 1986; Mitani and Nishida 1993; Slocombe and Zuberbühler 2007; Schel et al. 2013). To calculate which individuals may have been within auditory distance but not in visual sight, any individual observed with the focal within the 30-min period prior to arriving at the food patch but crucially, did not arrive with the focal, was considered to be possibly within a distance of a few hundred metres from the patch, therefore ‘nearby’ (Mitani and Nishida 1993). We recorded chimpanzee food grunts and barks on a Song Meter SM2+ device (Wildlife Acoustics) when chimpanzees were circa 250 m away (AK unpublished data) and therefore believe that food calls can travel up to a maximum of ca. 300 m in the Taï forest (Kalan et al. 2015). Interestingly, this happens to be the same distance that a spider monkey food call, or ‘whinny’, has been estimated to travel in a forest (Ramos-Fernández 2005). We chose a maximum interval of 30 min, because this was a sufficient amount of time for a receiver to transverse a putative 300 m distance while also reasonably assuming that a signaller may have a salient memory for having been with a specific individual within the last 30-min period and may believe that the individual is nearby (Mitani and Nishida 1993). The maximum 30-min delay was also chosen, because it was used in a recent experimental study on chimpanzee foraging cognition (Sayers and Menzel 2012). Since we are interested in vocal communication, it is essential that when individuals are not in visual contact, a vocal signal becomes the most effective means of communication. Thus, our definition of party composition is specifically concerned with visual contact, and our definition of nearby individuals specifically considers those individuals that the focal may reasonably believe are within earshot. Using this definition, nearby individuals ranged from being last seen with the focal between 3 min up to the full 30 min before arriving to the food source. Individuals that had been observed with the focal longer than 30 min ago were not considered to be nearby.
A female chimpanzee was considered to be in estrus when her sexual swelling was maximally tumescent, meaning the skin around the anogential region was fully swollen, tight and shiny which usually attracts males to copulate with her (Deschner et al. 2003). It has been shown that the majority of ovulations occur when these sexual swellings are maximally tumescent, thereby providing an indicator of the female’s conception probability to males (Deschner et al. 2003). In this study, we were not specifically concerned with conception, which would require hormone data, but rather the physical signal of the swelling that can be observed by conspecifics. Since we do not assume to know what may be more salient to a chimpanzee, whether it is the reproductive state of females or simply the sex of the individuals present, any estrous female was also scored as an adult female. Only three females were ever observed to be in estrus during this study period although never all three at the same time, one of which was nulliparous. Hence, although the number of adult females could range from 0 to 6, the number of estrous females observed ranged from 0 to a maximum of 2 and the number of males from 0 to a maximum of 5.
Statistical analysis
We ran three main analyses using generalized linear mixed models (GLMMs; (Baayen 2008) and two additional post hoc GLMMs. All GLMMs had a binomial error structure with a logit link function and were implemented using the function ‘glmer’ from the package ‘lme4’ (Bates et al. 2014) in R version 3.0.2 (R Core Team 2013). We accounted for potential temporal autocorrelation in the residuals (due to data points being near in time) of all models using the method detailed by Fürtbauer et al. (2011) and included it as a control predictor in our models (Mundry 2014). The term was dropped from the final model if the estimate was negative. For every GLMM, we included random slopes for all fixed effects within random intercepts, when applicable, to keep the probability of making a type I error at the desired level of 0.05 (Forstmeier and Schielzeth 2011; Barr et al. 2013). All continuous covariates were z-transformed to a mean of 0 and standard deviation of 1 before running the model (Schielzeth 2010). Model stability was evaluated by comparing model estimates where levels of the random effect(s) were dropped one at a time to ensure that they did not change model estimates significantly. We also checked for collinearity problems amongst predictors (Quinn and Keough 2002) by deriving variance inflation factors (VIFs) using the function ‘vif’ of the package ‘car’ (Fox et al. 2012) applied to standard linear models with fixed effects only. For each GLMM, a full versus null model comparison was conducted first to determine the significance of the test predictor variables as a whole (Forstmeier and Schielzeth 2011) using a likelihood ratio test (R function ‘anova’ (Dobson and Barnett 2008)). The null model comprised all model terms as the full model except for the fixed effects of interest. Only if the likelihood ratio test showed a significant difference (P < 0.05), we discuss the results of the model with respect to individual test predictors (Mundry 2014). The significance of individual predictors was also assessed using likelihood ratio tests. For all of the following models, stability assessment showed that estimates did not vary significantly and collinearity was not a problem, ensuring results were robust.
Recruitment model
The first model tested for the recruitment function of food calls, whereby conspecifics were expected to be attracted to feeding events where chimpanzees had uttered food calls during the feeding event. Specifically, we tested whether the arrival of other individuals (yes/no) to a feeding event was a function of whether food calls had been produced by the focal individual (yes/no), while also controlling for whether pant hoots had been produced during the feeding event (yes/no) which undoubtedly serve as a long-distance call and have been suggested to also attract group members to food sources when produced in a feeding context (Clark and Wrangham 1994). This model considered food calls and pant hoots produced at any time during the feeding event. For those events where arrivals of other chimpanzees had been observed, we specifically noted food calls and pant hoots that had been uttered before the first individual arrived. If multiple arrivals occurred during a feeding event, only the first arrival was considered. For events with no arrivals, food calls or pant hoots produced at any time were considered. It should be noted that for all feeding events observed, individuals who arrived always began to join in eating. Twenty-three events were removed because the whole group had arrived at the patch together (4/23 events food calls were still produced on arrival), leaving a total of 534 feeding events. We also removed any events where the first arrival occurred after 30 min of the focal arriving to the patch (7/153 events with arrivals) to limit the response window for an arrival to occur considering the maximum distance food calls can travel (see ‘Definitions of audience members’). The final sample size was 527 events for this analysis. The model controlled for food type (fruit, leaves or nuts) and party size, using a weighted mean according to duration and, importantly, included an offset term (McCullagh and Nelder 1989) to control for the time spent at the patch since the chance probability of new arrivals increases as event duration becomes longer. The random effect of food species (55 species) was accounted for, as well as the random slopes for food call and pant hoot production within food species (Table 1). We also ran the exact same model for fruit species only (N = 319 with 43 species) since previous research on food calls has shown that chimpanzees produce food calls more often for fruits than herbaceous foods (Fedurek and Slocombe 2013).
Audience effects model
The second analysis tested for audience effects on food call production, specifically, whether individuals that were nearby or present affected the probability of a chimpanzee to produce a food call upon arriving to a food patch. The response variable was whether the focal individual produced a food call upon arriving at a food patch within the first minute (yes/no). The critical test predictors included the two types of audience effects: presence of certain individuals accompanying the focal upon arrival and absent but nearby individuals. The following variables were therefore determined for the first minute of each feeding event, excluding the focal individual: the number of males present or nearby, the number of adult females present or nearby and the number of estrous females present or nearby. All six social audience effect test predictors were initially tested as three-way interactions with sex and rank of the caller since both sex and rank of a chimpanzee can affect their vocalization behaviour (Clark 1993). Again, food type and additional control predictors were time of day, also as a squared term, as a proxy for the variation in hunger due to daily activity patterns which may follow a quadratic rather than linear pattern, and whether the food patch was empty upon arrival or not. Every data point represented a single feeding event where we also had complete data on who was nearby (N = 497). The model included the random effects of focal ID (nine individuals) and food species (54 species) and included the random slopes for every interaction with a test predictor within focal ID and food species separately, as well as for all possible control variables (Table 1).
The assessment of model stability revealed all but one of the three-way interactions between the six test predictors and sex and rank to be somewhat unstable. Therefore, these interactions were dropped leaving only the respective two-way interactions (i.e., with sex and rank separately). At this point, model stability was achieved. Only once a full versus null model comparison was significant, we examined the significance of each predictor in its respective interaction (Forstmeier and Schielzeth 2011). If any particular interaction was non-significant, we dropped it and inspected the respective main effect in the final model to determine whether or not it had any influence. In this manner, we could draw robust inference regarding all social audience effects simultaneously.
Ecological model
The ecological variables tested for their potential influence on the probability of focal individuals uttering a food call upon arriving to a patch were dbh, Moran’s I, fruit count and days since the first feeding observation for a species. Thus, this model only included feeding events for fruit species where all ecological variables had been measured (N = 217). We put in two-way interactions between dbh, fruit count and Moran’s I since the effects of tree size and fruit count may be exacerbated or minimized for species that are clumped in their distribution. Sex, rank, arriving to an empty patch (yes/no), time, time2 and party size upon arrival were controlled for (we could include party size because it was no longer collinear with the other predictors; Table 1). Again, food species (seven species) and focal ID (nine individuals) were random effects in the model, and we included the random slope for every test and control predictor within the two random effects when applicable.
Post hoc analyses for recruitment of nearby chimpanzees
For those predictors where we found a significant effect of the number of nearby individuals in a certain class (e.g., estrous females) in the audience effects model, we ran post hoc analyses to determine whether events where food calls were produced upon arrival were significantly more likely to successfully recruit that specific class of nearby individuals. An event was considered successful in recruiting if at least one individual of that ‘targeted class’ was observed to arrive during the feeding event. Only the effects of sex and food calling on recruiting nearby individuals could be examined via post hoc analyses, because to test for the effect of rank and food calling on recruitment required an interaction between the two which was not possible to be fitted with the reduced datasets. To test for differences in recruitment among the two sexes, we could fit two separate GLMMs. One model for males (N = 35, 16 fruit species and 5 individuals) and another for females (N = 25, 12 species and 4 individuals), considering only those events where the potential targeted class of individuals was indeed nearby and for fruit species only, which allowed to reduce the complex random effects structure in the model. Only the test predictor, whether or not a food call had been produced by the focal upon arrival (yes/no) was included, while keeping the critical offset for event duration, and the random intercepts for species and focal ID as well as the random slopes for food calling within focal ID and food species. The critical control predictor of whether pant hoots had been produced during the event was included although additional control predictors could not be included because they caused model instability. The response variable was whether the targeted nearby individual(s) joined the feeding event (y/n).
Results
In total, we observed 557 complete feeding events where chimpanzees ate fruit (43 species), leaves (17 species) or nuts (3 species). Of those 557 events, food calls were produced in 41 %, with only 19 % of events eliciting food calls upon the first minute of arrival. In 89.9 % of the feeding events, there were no other chimpanzees already present at the food patch upon arrival. For comparison, pant hoots were produced even less often, having been uttered by focal individuals in 27 % of the 557 feeding events, with only 4 % of these events eliciting pant hoots by the focal within the first minute of arrival (Table 2).
The recruitment model showed that the arrival of other individuals to a feeding event was not influenced by food calling (estimate ± SE = 0.46 ± 0.35, Χ 2 = 1.61, df = 1, P = 0.21). Only pant hoots had a significant increase in the likelihood of chimpanzees to arrive (estimate ± SE = 0.72 ± 0.28, Χ 2 = 5.17, df = 1, P = 0.023). However, when we restricted our analysis to feeding events for fruit species only, arrivals of nearby individuals to the food patch were significantly more likely following food calls (estimate ± SE = 0.81 ± 0.31, Χ 2 = 5.16, df = 1, P = 0.023). Furthermore, this effect was similar to pant hoots which continued to have a significant positive effect on arrival probability (estimate ± SE = 0.85 ± 0.38, Χ 2 = 4.06, df = 1, P = 0.044).
Effects of audience and ecology on food call production
Of the 254 feeding events where a female chimpanzee was the focal, she uttered a food call upon arrival in 47 feeding events. The remaining 243 events where a male was the focal, he made a food call upon arrival in 51 events. The GLMM testing for audience effects showed that chimpanzee food call probability upon arriving to a patch was influenced by social factors (full-null model comparison Χ 2 = 44.45, df = 18, P = 0.0005; Table 3). There was a significant three-way interaction between sex and rank of the caller and the number of estrous females nearby, whereby high-ranking males had an increased probability of uttering a food call when there were more estrous females nearby (Table 3; Fig. 1). There was also a significant interaction between rank and the number of females nearby on food call probability (Table 3; Fig. 2), whereby both male and female chimpanzees of low rank uttered food calls more often when there were many females nearby irrespective of their reproductive state. With respect to the presence of particular individuals accompanying the focal chimpanzee, there was a significant influence for male chimpanzees to produce food calls more often when more males were in their party (Table 3; Appendix 1a), and males were less likely than females to produce a food call when adult females were accompanying them (Table 3; Appendix 1b). All other audience effects tested had non-significant effects (Table 4).

High-ranking male chimpanzees at Taï were more likely to utter a food call upon arrival as the number of estrous females nearby increased (N = 497). Low-ranking females generally called more often than high-ranking females, regardless of the number of estrous females nearby. The surface represents the fitted model; circles represent the averaged response per cell of the surface, and their volume corresponds to the relative number of observations in the respective cell. Filled in circles fall above the 3D model surface, depicted by solid vertical lines, while empty circles fall below it, depicted by dashed vertical lines

Both male (N = 243) and female (N = 254) Taï chimpanzees of low rank were more likely to produce food calls upon arriving to a food patch when many females were nearby. Plots are shown for the two sexes separately for clarity. The surface represents the fitted model; circles represent the averaged response per cell of the surface, and their volume corresponds to the relative number of observations in the respective cell. Filled in circles fall above the 3D model surface, depicted by solid vertical lines, while empty circles fall below it, depicted by dashed vertical lines
The only control variable having a significant influence on food call probability was food type (Χ 2 = 14.39, df = 2, P < 0.001), where food calls were most often produced for fruits, secondly for nuts and least often for leaves. However, the results of the GLMM testing for more precise ecological variables showed that there was no overall significance of ecological predictors on the probability of a chimpanzee producing a food call within the first minute of arrival to a food patch, both with the interactions included (full-null model comparison Χ 2 = 8.63, df = 7, P = 0.28) and when including main effects only (Χ 2 = 7.02, df = 4, P = 0.14).
Post hoc analyses of recruitment
There were 53 feeding events where a male chimpanzee was the focal, and at least one estrous female was nearby. A food call was produced on arrival for 19 of those events of which 15 were for fruit species. Of these 15 events, the estrous female was successfully recruited to the patch in 12 (80 %; Fig. 3). Post hoc analyses showed that recruitment success of male chimpanzees for an estrous female was significantly higher when they uttered a food call upon arriving to fruit trees (estimate ± SE = 3.38 ± 1.75, Χ 2 = 4.80, df = 1, P = 0.029). On the other hand, female chimpanzees did not differ in their recruitment success for an estrous female when they made food calls at fruit trees (estimate ± SE = 0.39 ± 1.57, Χ 2 = 0.059, df = 1, P = 0.81; Fig. 3). Of the 12 events where a food calling male chimpanzee was successful in recruiting an estrous female(s), that male was observed to subsequently copulate with that estrous female(s), either during or after the event had ended, for eight of the 12 events (67 %; Fig. 4). For the remaining four events, the male had been observed to copulate with that estrous female earlier in the day. Alternatively, of the 20 events where males had not uttered a food call upon arriving to a fruit tree, a nearby estrous female(s) was still attracted to the patch for five of these events (25 %; Fig. 3). The male focal was observed to copulate with that estrous female(s) during one of those feeding events. For another three of those events, the male had copulated with the estrous female earlier in the day, and for one of the events, we lost the focal after the feeding event so cannot be sure of what happened afterwards. Unfortunately, due to the reduced number of observations, we were not able to fit a stable model to test if this difference was statistically significant (Fig. 4).

The probability of a nearby estrous female(s) to be attracted to a food patch as a function of whether a food call had been produced by the focal chimpanzee, tested separately for males and females. In brackets is the total number of feeding events for fruit species where a food call had been made on arrival (black bar) or not (white bar)

The number of feeding events where an estrous female(s) had been successfully attracted, and the male focal chimpanzee was observed to copulate with her before, during or after the feeding event
Discussion
This study shows that the probability of a chimpanzee making a food call was influenced by social context more than ecological context. Having individuals both present with and nearby (i.e., within earshot), the caller affected whether chimpanzees made a food call upon arriving to a food patch. Overall, male chimpanzees were more likely to produce a food call when in the company of many males but less likely when accompanied by many females (Table 3). The effect of nearby individuals on food call probability was highly significant (Table 3). Both low-ranking males and females were more likely to utter a food call when there were many females nearby (Fig. 2). In particular, high-ranking males were more likely to call when more estrous females were nearby (Fig. 1). Crucially, when male chimpanzees were observed to utter food calls upon arrival, they were effective in recruiting these nearby estrous females to food patches (Fig. 3).
Similar to what was reported in other studies, fruit species elicited food calls more often than nuts or leaves (Slocombe et al. 2010). However, based on our previous findings (Kalan et al. 2015), we were surprised to not find an effect of other ecological variables on the probability of uttering a food call upon arrival. This could be an artefact of restricting our analysis to the first minute upon arrival, since ecological factors may become more salient as food is being ingested and its quality better assessed (e.g., number of fruits, patch size). In fact, the aforementioned study analyzed calls produced during a feeding event while controlling for call latency. On the other hand, our findings are congruent with the literature on animal communication whereby presence or absence of call production is often modulated by the social audience (Seyfarth and Cheney 2003; Zuberbühler 2008). Clearly, the mere presence of food is not sufficient to motivate call production since calls were produced on average for only 40 % of feeding events in this study (Table 2). Thus, particular social situations are more likely to elicit chimpanzee food calls as demonstrated in this study and others (Slocombe et al. 2010; Fedurek and Slocombe 2013; Schel et al. 2013).
Recruitment of nearby individuals
Our results provide support for chimpanzee food calls functioning as recruitment calls that attract nearby individuals to a food patch when feeding on fruit species, even when controlling for the effect of pant hoots. To our knowledge, these are the first data supporting the observation proposed by many researchers regarding the attractive properties of chimpanzee food calls (Marler and Tenaza 1977; Wrangham 1977; Goodall 1986; Hauser and Wrangham 1987; Slocombe and Zuberbühler 2005). Although, a previous study showed that food call structure was influenced by tree size for a particular fruit species and chimpanzees tended to be attracted to calls made specifically for large trees (Kalan et al. 2015). It would be worthwhile examining how much of this recruitment behaviour is dependent on a fission-fusion social system which is characteristic of both chimpanzees and spider monkeys (Chapman and Lefebvre 1990) and why some species use food calls to similarly attract conspecifics (Elgar 1986; Heinrich and Marzluff 1991; Caine et al. 1995; Wilkinson and Boughman 1998) while others repel conspecifics (Boinski and Campbell 1996). Species requiring greater defence at food patches from predators, such as house sparrows (Elgar 1986), or requiring defence of resources from other groups of the same species may benefit from recruiting others if this results in a higher probability of being able to successfully forage in a group than when alone (Heinrich and Marzluff 1991; Wilkinson and Boughman 1998). While in other species, such as chimpanzees and spider monkeys, food calls may ultimately serve a recruitment function as part of a strategy to manage a less predictable social environment, characteristic of fission-fusion societies (Chapman and Lefebvre 1990), rather than for defending food resources per se.
Although this is the first study to provide evidence for call production being adjusted with respect to the number of estrous females nearby, a seminal study by Mitani and Nishida (1993) did report observations of some males producing pant hoots when estrous females were nearby. However, Mitani and Nishida’s (1993) principal finding was that male chimpanzees called to attract nearby allies. We did not find support for this in our study, likely due to the low number of males present in this group and perhaps also due to the tumultuous social context during which these data were collected (including an alpha takeover and the disappearance of one adult male). Yet, the other audience effects supported in our data were similar to those reported in previous studies of chimpanzee food calls where males were more likely to utter a food call when in the presence of other males with whom they were strongly bonded (Slocombe et al. 2010; Fedurek and Slocombe 2013). Thus, to some extent, our results support Slocombe and colleagues’ (2010) bonding hypothesis for food calls where individuals call to coordinate feeding activity with others. Indeed, the suggestion that food calls may function to keep certain individuals together and to strengthen social bonds (Slocombe et al. 2010) does not exclude the putative motivation for food calling to recruit certain individuals to join the caller. In fact, we suggest that food calls may serve multiple functions and there may be fine scaled differences in food call structure that could help us to understand whether some food call subtypes (e.g., a soft grunt as opposed to a bark) function more to recruit others while other types are used to maintain contact with individuals already feeding with the caller (Fedurek and Slocombe 2013).
Reciprocal altruism in food calling
The apparent costly behaviour of food calls could be offset by a mechanism such as reciprocal altruism, namely an exchange of information about food resources (de Waal 1989) or trading information about food for mating opportunities (Roberts 1998). Our result suggests that male chimpanzees could be using food calls as a form of altruistic signalling which could potentially act as an indicator of male quality. In fact, Wrangham’s (1977) original hypothesis suggested that food calling may be a costly signal which could advertise an individual’s ability to suffer feeding competition to the benefit of potential mates and allies. This study provides support for this hypothesis, especially since males of higher rank were the most likely to utter food calls when an estrous female was nearby. The only other species where food calls have been reported to occur in a mating context is for gallinaceous birds, where males are observed to use food calls in courtship displays as a lure to attract females (Stokes and Williams 1972), and the production of food calls by chickens has been shown to be modulated by the presence of a hen (Evans and Marler 1994). Based on these studies and our present results, it is plausible that food calls in both chimpanzees and gallinaceous birds may have been principally produced in a foraging context but were secondarily adapted by males for their influence on females via sexual selection. However, whether food calling is an honest signal of a male’s fitness remains to be seen. It would be of particular interest to follow up the findings here using a playback study to see whether food calls produced by high-ranking males are better at attracting nearby estrous females compared to calls made by low rankers. It is crucial, however, that in those cases where males produced food calls when estrous females were nearby, they had a high success rate of attracting that individual to the food patch (80 %) and were even observed to successfully mate with her afterwards (67 %).
Low-ranking males increased food call probability when there were many females nearby, irrespective of their reproductive status. This is perhaps a peculiar finding since these males are not benefiting by attracting mating opportunities per se. Still, this could be an example of a long-term investment via reciprocal altruism whereby low-ranking males ‘invite’ females to share food in hopes of a delayed benefit for future mating opportunities. Similar behaviour has already been reported for the Taï chimpanzees, where long-term exchanges of meat for sex characterized some male-female dyads (Gomes et al. 2009).
Importantly, this study provides additional insights into calling behaviour of female chimpanzees, and here, we found similar effects as observed for males even though females vocalize less frequently overall (Clark 1993). Low-ranking females were more likely to utter a food call when more females were nearby (Fig. 3). There is evidence that female chimpanzees form strong social bonds with members of their own sex (Wakefield 2008; Lehmann and Boesch 2009) and female chimpanzees in Taï have been shown to exhibit a high degree of sociality and gregariousness (Riedel et al. 2011; Wittiger and Boesch 2013). Hence, females may benefit from food calling for other females via a process of reciprocal altruism as described above, which can ultimately serve to strengthen social relationships amongst females. Alternatively, the motivation for females to food call and attract other females may be driven by predator defence since Taï is characterized by a high density of leopards that attack and can kill chimpanzees (Boesch 1991).
Critically, our results emphasize the importance of animal vocalizations as a means of communicating with individuals that are not within visual contact. The use of vocalizations specifically for recruitment is particularly noteworthy for studies of animal cognition in determining whether animals are capable of understanding the minds of ‘targeted’ recipients (Crockford et al. 2012). As has been previously shown with spider monkey food calls (Chapman and Lefebvre 1990), the extent to which chimpanzees might use food calls to manipulate their social environment could provide significant insights into complex capabilities, particularly deception, and should thus be investigated further.
References
Baayen RH (2008) Analyzing linguistic data: a practical introduction to statistics. Cambridge University Press, Cambridge
Barr DJ, Levy R, Scheepers C, Tily HJ (2013) Random effects structure for confirmatory hypothesis testing: keep it maximal. J Mem Lang 68:255–278
Bates D, Maechler M, Matrix LT (2014) lme4: Linear mixed effects models using S4 classes. CRAN.R project, https://cran.r-project.org/web/packages/lme4/vignettes/lmer.pdf
Boesch C (1991) The effects of leopard predation on grouping patterns in forest chimpanzees. Behaviour 117:220–242
Boesch C, Boesch-Achermann H (2000) The chimpanzees of the Taï Forest: behavioural ecology and evolution. Oxford University Press, Oxford
Boinski S, Campbell AF (1996) The huh vocalization of white-faced capuchins: a spacing call disguised as a food call? Ethology 102:826–840
Caine NG, Addington RL, Windfelder TL (1995) Factors affecting the rates of food calls given by red-bellied tamarins. Anim Behav 50:53–60
Chapman CA, Lefebvre L (1990) Manipulating foraging group size: spider monkey food calls at fruiting trees. Anim Behav 39:891–896
Chapman CA, Chapman LJ, Wangham R et al (1992) Estimators of fruit abundance of tropical trees. Biotropica 24:527
Chapman CA, Chapman LJ, Wrangham RW (1995) Ecological constraints on group size: an analysis of spider monkey and chimpanzee subgroups. Behav Ecol Sociobiol 36:59–70
Clark AP (1993) Rank differences in the production of vocalizations by wild chimpanzees as a function of social context. Am J Primatol 31:159–179
Clark AP, Wrangham RW (1994) Chimpanzee arrival pant-hoots: do they signify food or status? Int J Primatol 15:185–205
Clay Z, Smith CL, Blumstein DT (2012) Food-associated vocalizations in mammals and birds: what do these calls really mean? Anim Behav 83:323–330
Crockford C, Wittig RM, Mundry R, Zuberbühler K (2012) Wild chimpanzees inform ignorant group members of danger. Curr Biol 22:142–146
de Waal FBM (1989) Food sharing and reciprocal obligations among chimpanzees. J Hum Evol 18:433–459
Deschner T, Heistermann M, Hodges K, Boesch C (2003) Timing and probability of ovulation in relation to sex skin swelling in wild West African chimpanzees, Pan troglodytes verus. Anim Behav 66:551–560
Di Bitetti MS (2003) Food-associated calls of tufted capuchin monkeys (Cebus apella nigritus) are functionally referential signals. Behaviour 140:565–592
Dittus WPJ (1984) Toque macaque food calls: semantic communication concerning food distribution in the environment. Anim Behav 32:470–477
Dobson AJ, Barnett AG (2008) An introduction to generalized linear models. Chapman & Hall/CRC, Boca Raton
Elgar MA (1986) The establishment of foraging flocks in house sparrows: risk of predation and daily temperature. Behav Ecol Sociobiol 19:433–438
Evans CS, Marler P (1994) Food calling and audience effects in male chickens, Gallus gallus: their relationships to food availability, courtship and social facilitation. Anim Behav 47:1159–1170
Fedurek P, Slocombe KE (2013) The social function of food-associated calls in male chimpanzees. Am J Primatol 75:726–739
Fedurek P, Machanda ZP, Schel AM, Slocombe KE (2013) Pant hoot chorusing and social bonds in male chimpanzees. Anim Behav 86:189–196
Fedurek P, Donnellan E, Slocombe KE (2014) Social and ecological correlates of long-distance pant hoot calls in male chimpanzees. Behav Ecol Sociobiol 68:1345–1355
Forstmeier W, Schielzeth H (2011) Cryptic multiple hypotheses testing in linear models: overestimated effect sizes and the winner’s curse. Behav Ecol Sociobiol 65:47–55
Fox J, Weisberg S, Bates D, Fox MJ (2012) Package “car”. https://cran.r-project.org/web/packages/car/car.pdf
Fürtbauer I, Mundry R, Heistermann M, Schülke O, Ostner J (2011) You mate, I mate:macaque females synchronize sex not cycles. PLoS ONE 6:e26144
Gomes CM, Mundry R, Boesch C (2009) Long-term reciprocation of grooming in wild West African chimpanzees. Proc R Soc Lond B 276:699–706
Goodall J (1986) The chimpanzees of gombe: patterns of behavior. Belknap Press of the Harvard University Press, Cambridge
Hauser MD, Marler P (1993a) Food-associated calls in rhesus macaques (Macaca mulatta): I. Socioecological factors. Behav Ecol 4:194–205
Hauser MD, Marler P (1993b) Food-associated calls in rhesus macaques (Macaca mulatta): II. Costs and benefits of call production and suppression. Behav Ecol 4:206–212
Hauser MD, Wrangham RW (1987) Manipulation of food calls in captive chimpanzees. Folia Primatol 48:207–210
Hauser MD, Teixidor P, Fields L, Flaherty R (1993) Food-elicited calls in chimpanzees: effects of food quantity and divisibility. Anim Behav 45:817–819
Heinrich B, Marzluff JM (1991) Do common ravens yell because they want to attract others? Behav Ecol Sociobiol 28:13–21
Hobaiter C, Byrne RW (2011) The gestural repertoire of the wild chimpanzee. Anim Cogn 14:745–767
Janik VM (2000) Food-related bray calls in wild bottlenose dolphins (Tursiops truncatus). Proc R Soc Lond B 267:923–927
Kalan AK, Mundry R, Boesch C (2015) Wild chimpanzees modify food call structure with respect to tree size for a particular fruit species. Anim Behav 101:1–9
Lehmann J, Boesch C (2009) Sociality of the dispersing sex: the nature of social bonds in West African female chimpanzees, Pan troglodytes. Anim Behav 77:377–387
Marler P, Tenaza RR (1977) Signaling behavior of apes with special reference to vocalization. In: Sebeok TA (ed) How animals communicate. Indiana University Press, Bloomington, pp 965–1033
Marler P, Dufty A, Pickert R (1986) Vocal communication in the domestic chicken: II. Is a sender sensitive to the presence and nature of a receiver? Anim Behav 34:194–198
McCullagh P, Nelder JA (1989) Generalized linear models, 2nd edn. Chapman and Hall, London
Mitani JC, Nishida T (1993) Contexts and social correlates of long-distance calling by male chimpanzees. Anim Behav 45:735–746
Mundry R (2014) Statistical issues and assumptions of phylogenetic generalized least squares. In: Garamszegi LZ (ed) Modern phylogenetic comparative methods and their application in evolutionary biology. Springer, Berlin, pp 131–153
Quinn GP, Keough MJ (2002) Experimental design and data analysis for biologists. Cambridge University Press, Cambridge
Ramos-Fernández G (2005) Vocal communication in a fission-fusion society: do spider monkeys stay in touch with close associates? Int J Primatol 26:1077–1092
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
Riedel J, Franz M, Boesch C (2011) How feeding competition determines female chimpanzee gregariousness and ranging in the Taï National Park, Côte d’Ivoire. Am J Primatol 73:305–313
Roberts G (1998) Competitive altruism: from reciprocity to the handicap principle. Proc R Soc Lond B 265:427–431
Sayers K, Menzel CR (2012) Memory and foraging theory: chimpanzee utilization of optimality heuristics in the rank-order recovery of hidden foods. Anim Behav 84:795–803
Schel AM, Machanda Z, Townsend SW et al (2013) Chimpanzee food calls are directed at specific individuals. Anim Behav 86:955–965
Schielzeth H (2010) Simple means to improve the interpretability of regression coefficients. Methods Ecol Evol 1:103–113
Seyfarth RM, Cheney DL (2003) Signalers and receivers in animal communication. Annu Rev Psychol 54:145–173
Slocombe KE, Zuberbühler K (2005) Functionally referential communication in a chimpanzee. Curr Biol 15:1779–1784
Slocombe KE, Zuberbühler K (2006) Food-associated calls in chimpanzees: responses to food types or food preferences? Anim Behav 72:989–999
Slocombe KE, Zuberbühler K (2007) Chimpanzees modify recruitment screams as a function of audience composition. Proc Natl Acad Sci U S A 104:17228–17233
Slocombe KE, Kaller T, Turman L, Townsend SW, Papworth S, Squibbs P, Zuberbühler K (2010) Production of food-associated calls in wild male chimpanzees is dependent on the composition of the audience. Behav Ecol Sociobiol 64:1959–1966
Sterck EHM, Watts DP, van Schaik CP (1997) The evolution of female social relationships in nonhuman primates. Behav Ecol Sociobiol 41:291–309
Stokes AW, Williams HW (1972) Courtship feeding calls in gallinaceous birds. Auk 89:177–180
Townsend SW, Deschner T, Zuberbühler K (2008) Female chimpanzees use copulation calls flexibly to prevent social competition. PLoS ONE 3:e2431
Trivers R (1971) The evolution of reciprocal altruism. Q Rev Biol 46:35–57
Wakefield ML (2008) Grouping patterns and competition among female Pan troglodytes schweinfurthii at Ngogo, Kibale National Park, Uganda. Int J Primatol 29:907–929
Wilkinson GS, Boughman JW (1998) Social calls coordinate foraging in greater spear-nosed bats. Anim Behav 55:337–350
Wittig RM, Boesch C (2003) Food competition and linear dominance hierarchy among female chimpanzees of the Taï National Park. Int J Primatol 24:847–867
Wittiger L, Boesch C (2013) Female gregariousness in Western Chimpanzees (Pan troglodytes verus) is influenced by resource aggregation and the number of females in estrus. Behav Ecol Sociobiol 67:1097–1111
Wrangham RW (1977) Feeding behaviour of chimpanzees in Gombe national park, Tanzania. In: Clutton-Brock TH (ed) Primate ecology: studies of feeding and ranging behavior in lemurs monkeys and apes. Academic Press, London, pp 503–538
Zuberbühler K (2008) Audience effects. Curr Biol 18:R189–R190
Acknowledgments
This study was funded by the Max Planck Society. We thank the Ministère de la Recherche Scientifique and the Ministère de l’Environnement et des Eaux et Forêts of Côte d’Ivoire and Office Ivorien des Parcs et Reserves for permission to conduct research in the country and Taï National Park. For logistical support, we thank the Centre Suisse de Recherche Scientifique and the Taï Chimpanzee Project, especially all field assistants and Roman Wittig. Thanks to Cathy Crockford for comments on earlier drafts as well as Roger Mundry and Colleen Stephens for statistical support. We also thank two anonymous reviewers for helpful comments that improved this manuscript.
Ethical standards
This research was conducted with permissions from the relevant Ivorian authorities in accordance with the national laws and animal care regulations of Côte d’Ivoire as well as Germany. The study complied with the ethical standards of the Max Planck Institute for Evolutionary Anthropology and the Primatology department’s ethical guidelines for non-invasive research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by D. P. Watts
Electronic supplementary material
Below is the link to the electronic supplementary material.
Appendix 1
(DOCX 180 kb)
Rights and permissions
About this article

Cite this article
Kalan, A.K., Boesch, C. Audience effects in chimpanzee food calls and their potential for recruiting others. Behav Ecol Sociobiol 69, 1701–1712 (2015). https://doi.org/10.1007/s00265-015-1982-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-015-1982-1