
Abstract
Element cycling in aquatic systems is driven chiefly by planktonic processes, and the structure of the planktonic food web determines the efficiency of carbon transfer through trophic levels. However, few studies have comprehensively evaluated all planktonic food-web components in tropical regions. The aim of this study was to unravel the top-down controls (metazooplankton community structure), bottom-up controls (resource availability), and hydrologic (water residence time) and physical (temperature) variables that affect different components of the microbial food web (MFW) carbon stock in tropical reservoirs, through structural equation models (SEM). We conducted a field study in four deep Brazilian reservoirs (Balbina, Tucuruí, Três Marias, and Funil) with different trophic states (oligo-, meso-, and eutrophic). We found evidence of a high contribution of the MFW (up to 50% of total planktonic carbon), especially in the less-eutrophic reservoirs (Balbina and Tucuruí). Bottom-up and top-down effects assessed through SEM indicated negative interactions between soluble reactive phosphorus and phototrophic picoplankton (PPP), dissolved inorganic nitrogen, and heterotrophic nanoflagellates (HNF). Copepods positively affected ciliates, and cladocerans positively affected heterotrophic bacteria (HB) and PPP. Higher copepod/cladoceran ratios and an indirect positive effect of copepods on HB might strengthen HB-HNF coupling. We also found low values for the degree of uncoupling (D) and a low HNF/HB ratio compared with literature data (mostly from temperate regions). This study demonstrates the importance of evaluating the whole size spectrum (including microbial compartments) of the different planktonic compartments, in order to capture the complex carbon dynamics of tropical aquatic ecosystems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Inland waters play an important role in the global carbon cycle, being extremely active in the transport, mineralization, and storage of carbon [1]. Tropical freshwaters are particularly important for global biogeochemical cycles, as they are responsible for approximately 60% of the total carbon emissions from inland waters [2]. In addition to enhanced carbon emissions associated with high temperatures, low-latitude lakes and reservoirs usually stratify for longer periods, and hypolimnetic waters remain warm, creating optimal conditions for anoxia [3], and develop high concentrations of CO2 and CH4 that may later enter the atmosphere [4, 5]. A recent study reported that carbon emissions in tropical reservoirs were negatively correlated with age and latitude, and these emissions were particularly high in the tropical Amazon region [6].
The mechanisms that lead to these high emissions in tropical reservoirs are complex, as they involve multiple factors including stratification, primary production, community respiration, nutrient loading, food-web structure, and others. In large reservoirs, cycling of the main elements is driven chiefly by the planktonic food web, which can regenerate and transfer these elements to higher trophic levels [7, 8]. The food-web structure may influence and alter carbon transfer and emissions in freshwater systems [9–11]. Tropical aquatic systems have some particular food-web configurations, but comprehensive studies on all planktonic food-web components are extremely rare [12, 13].
Aquatic food webs in warm lakes are expected to have a smaller biomass of large-bodied zooplankton compared to temperate systems because high temperatures affect organism physiology either directly or indirectly by increasing fish predation on zooplankton [14–17]. In tropical regions, omnivorous fish are more frequent and high biomasses of small-sized and young-of-the-year fish prevail throughout the year, exerting higher predation pressure on large-bodied zooplankton, especially large cladocerans [16, 18]. Consequently, zooplankton in warm lakes is expected to be small-bodied, with a high proportion of protozooplankton (ciliates and nanoflagellates), which might impact bacterioplankton through high grazing pressure [12, 19]. Metazooplankton composition has strong direct and indirect (cascading) effects on the microbial food web (MFW) [20]. Large daphnids are keystone species in temperate systems, as they alter the MFW through their large impact on protozooplankton [21]. A characteristic of tropical aquatic food webs is the absence (or generally low abundance) of large daphnids [22].
Among the components of the microbial food web, protozooplankton (ciliates and heterotrophic nanoflagellates) is a key compartment, contributing to carbon and energy flux because these plankters can grow faster than phytoplankton [23, 24]. Heterotrophic nanoflagellates (HNF) play a particularly important role in aquatic microbial communities through predation on the smaller size fractions of planktonic communities (phototrophic picoplankton—PPP, and heterotrophic bacteria—HB), channeling picoplanktonic production to the higher trophic levels [23]. One way of measuring this impact is to study the degree of uncoupling (D) between HNF and their prey [25]. Previous studies have reported that rotifers and small cladocerans efficiently control HNF populations, and negatively affect the degree of coupling between HNF and the bacterial community [26–28]; while copepods can enhance the efficiency of the HB-HNF coupling mechanisms [20]. A recent study comparing tropical and temperate HNF-HB abundance showed lower numbers in tropical systems, and the lack of coupling was explained by predation on HNF by ciliates and small cladocerans [29].
In addition, despite the indications of a weak [30] or absent [29] relationship between HB abundance and chlorophyll-a in different tropical freshwater systems, a recent study in large African lakes reported a high degree of HB—primary production coupling, with a high percentage of dissolved primary production (extracellular organic carbon released by phytoplankton) [31]. Moreover, a recent meta-analysis revealed that bacterial remineralization rates per bacterial production unit in tropical freshwaters are at least twofold higher than in temperate regions [32]. For these reasons, it is important to elucidate the strength of the MFW in tropical aquatic systems, particularly its contribution to biomass stocks and fate. Since microbial metabolism has major impacts on carbon pathways and ultimately on gas exchange with the atmosphere [12, 32], microbes might comprise an important fraction of the planktonic carbon stock in these systems. Trophic status is also an important characteristic of the system that might influence planktonic trophic structure and can determine if MFW is a carbon sink or link [21, 33, 34]. In a trophic-state gradient, all planktonic components should increase their biomass in more-eutrophic systems [33], while the contribution of the MFW to total planktonic carbon should decrease [21].
The aim of this study was to elucidate the roles of top-down (i.e., metazooplankton community structure), bottom-up (resource availability), and abiotic variables as controls on different microbial food-web compartments in tropical reservoirs. We monitored key components of the planktonic microbial and classical food webs in four tropical reservoirs with different metazooplankton community structures, covering a range of trophic states (productivity gradient), and analyzed food-web interactions using structural equation models (SEM). We hypothesized that the microbial food-web components contribute considerably to the carbon pool in these tropical systems, especially in the less-productive ones, where heterotrophy is expected to predominate over autotrophy.
Material and Methods
Study Sites
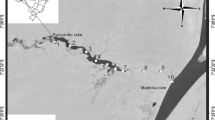
Sampling was conducted in four deep tropical hydroelectric reservoirs in Brazil (Fig. 1), with different morphologies, hydrodynamics, and trophic states (Table 1), in three different biomes: Amazonia (Balbina, Tucuruí), Cerrado (Três Marias), and Atlantic Forest (Funil). The Amazonian reservoirs are situated in different regions of northern Brazil: Balbina Reservoir is located in a mainly forested area of the central Amazon (79.0% [35]; Table 1) and is fed by a black-water river (Uatumã basin), while Tucuruí is located in the eastern Amazon, fed by a clear-water river (Tocantins/Araguaia basin). The drainage basin surrounding Tucuruí is extensively deforested, with vast agricultural areas (51.6% [35], Table 1). Três Marias Reservoir is located on the Central Brazilian Plateau (São Francisco basin) where the main vegetation is the Cerrado (Brazilian savanna) domain, which, however, has also been extensively altered for agriculture (70.5% [35], Table 1). Funil Reservoir in southeastern Brazil drains a densely populated area (Paraíba do Sul basin, 5.6% urban area [35], Table 1), receiving a high nutrient load [36]. This is a eutrophic reservoir with recurrent intense blooms of the cyanobacteria Dolichospermum circinale (Rabenhorst ex Bornet et Flahault) Wacklin, Hoffmann et Komárek, Cylindrospermopsis raciborskii (Woloszyńska) Seenayya et Subba Raju, and Microcystis aeruginosa (Kützing) Kützing [36, 37]. Balbina, Tucuruí, and Três Marias are large dendritic reservoirs, while Funil has a more-compact flooded area. The Amazonian reservoirs Balbina and Tucuruí are of nearly the same age of impoundment (∼30 years), while Três Marias and Funil were established ∼50 years ago, and the four reservoirs are monomictic with a hypoxic hypolimnion [36, 38–40].

Map and location of the Amazonian Balbina (a) and Tucuruí (b) reservoirs, and the non-Amazonian Três Marias (c) and Funil (d) reservoirs. Dots indicate locations of sampling stations. Dams are indicated by arrows
Sampling and Data Analysis
Physical, chemical, and biological variables of the four waterbodies were recorded at three sampling points of the reservoirs (including one point near the dam), during four periods of the year in 2011 and 2012, totaling 12 measurements per reservoir. The sampling period comprised the hydrological pulses in the Amazonian Balbina and Tucuruí reservoirs, and two rainy and two dry seasons in Três Marias and Funil reservoirs. Water temperature (WT) and pH were obtained with a YSI 6920 multiparameter probe (Yellow Springs Instruments, USA). The euphotic zone (Z eu) was measured as 2.7× Secchi disk extinction depth [50]. Integrated samples from the surface were taken using a 1.5-m-long tube sampler. Total phosphorus (TP), total nitrogen (TN), soluble reactive phosphorus (SRP), dissolved inorganic nitrogen (DIN), and dissolved organic carbon (DOC) were also measured from the integrated samples. TP and TN were estimated from unfiltered water samples, while DOC, SRP, and DIN (the sum of ammonium, nitrate, and nitrite) were filtered through glass-fiber filters (GF/C Whatman) and analyzed according to Wetzel and Likens [51]. The reservoir trophic states were assessed based on subsurface median concentrations of TP, according to Nürnberg [52]. Water residence time (RT, in days) was estimated considering the inflow discharge and the reservoir volume on daily basis, from data provided by the Brazilian National Electricity Regulatory Agency (ANEEL). The RT for each sampling period was calculated as the mean during the preceding month.
Water samples for picoplankton (PPP and HB), protozooplankton (HNF and ciliates), and phytoplankton for quantitative analysis were collected using the same tube sampler, while samples of metazooplankton (rotifers, cladocerans, calanoid copepods, cyclopoid copepods, copepodites, and nauplii) were collected by vertical hauls of a plankton net with 50-μm mesh, in the euphotic zone. For phytoplankton and ciliates, 100-mL samples were preserved with neutral Lugol’s and acid Lugol (2% final concentration) iodine solutions, respectively. For PPP and HNF, 100-mL samples were preserved with 10% glutaraldehyde (1% final concentration), and for HB, 10-mL samples were preserved in formalin at 2% final concentration. The metazooplankton 100-mL concentrates were preserved in 4% buffered formaldehyde.
Phytoplankton (Phyto) and ciliate (Cili) populations were concentrated by sedimentation and enumerated in random fields [53] for phytoplankton under an inverted microscope (Zeiss Oberkochen Axiovert 10, Carl Zeiss, Germany) at ×400 magnification [54], and all of a 10-mL chamber in phase-contrast illumination for ciliates. Phytoplankton taxonomic classes are according to Van den Hoeck et al. [55], except for cyanobacteria [56, 57]. Phytoplankters were identified to species as far as possible, according to appropriate literature, and ciliates were assigned to morphotypes. In general, at least 30 cells of each species or morphotype were measured directly under the microscope. Biovolume (mm3 L−1) for the two communities was estimated by multiplying the density of each species by the mean volume of its cells [58]. The carbon content (μg C L−1) for each plankton component was estimated. The carbon content of phytoplankton species was obtained from the biovolume through a conversion formula [59], and that of ciliates through a conversion factor of 140 fg μm−3 [60] after a correction factor of 1.4 [61].
For PPP and HB enumeration, 10 and 2 mL, respectively, were filtered through 0.2 μm black polycarbonate filters (Millipore) and stained with DAPI (4,6 diamidino-2-phenylindole) [62]. The filters were mounted on microscope slides with immersion oil, and the picoplankton abundance was estimated with the aid of an epifluorescence microscope (Zeiss Axio Observer A1, Carl Zeiss, Germany) at ×1000 magnification, by counting 200 cells, or 30 random fields in cases of low abundance [63, 64]. The blue excitation light filter set (EX BP 450-490, BS FT 510, EM LP 515) was used to quantify picocyanobacteria and picoeukaryotic cells, while the green excitation light filter set (H 546/12 BS FT 580 EM LP 590) was used only for picocyanobacteria. HB were visible under ultraviolet (UV) excitation (filter set: EX BP 365/12, BS FT 395 EM LP 397). PPP cells were measured directly under an epifluorescence microscope; the biovolume was calculated from the geometric formula for a prolate spheroid or sphere, and the carbon content was estimated using a conversion factor of 200 fg C μm−3 [65]. HB biomass was estimated from application of the major and minor axis measurements of cells in the formula proposed by Massana et al. [66], and biomass conversion according to Norland [67].
For HNF enumeration, 10 mL was filtered through 0.8 μm polycarbonate filters (Millipore) stained with DAPI, and the residue was mounted on microscope slides with immersion oil. The cells were counted with an epifluorescence microscope (Zeiss Axio Observer A1, Carl Zeiss, Germany) at ×1000 magnification using the UV filter set; the photosynthetic flagellates were distinguished with the blue and green filter sets. HNF cells were measured directly under the epifluorescence microscope. Biovolume was calculated from geometric formulas, using a conversion factor of 220 fg C μm−3 [68] for the carbon content.
The metazooplankton community (Metazoo) was enumerated using either a Sedgwick-Rafter chamber under a microscope (for rotifers, nauplii, and cladocerans) or in open chambers under a stereomicroscope (for copepodites and adult copepods). At least 250 individuals and 100 of the more abundant species were counted. Densities (individuals per liter) were estimated based on the volume of water that passed through the net. The organisms were classified in large groups: rotifers, cladocerans, calanoid and cyclopoid copepods, nauplii, and copepodites. Cladoceran and copepod biomasses were measured from the dry weight of 20 individuals after they were oven-dried for 24 h at 60 °C. Rotifer biomass was estimated from the median weight of each species as given in the literature [51], multiplied by its density in each sample [51], and the biomass of nauplii according to allometric formulas [69]. The carbon content (μg C L−1) of all metazooplanktonic components was estimated as 50% of dry weight [70]. In order to determine if the reservoirs had different metazooplanktonic community structures, and their influence on microbial food-web components (especially on HNF and HB) and/or phytoplankton, we calculated the ratio of copepods to cladocerans (COP/CLAD) for each reservoir.
Statistical Analysis
Variables were tested for differences among reservoirs using one-way ANOVA and a post hoc Tukey test (p < 0.05). We assumed marginally significant p values between 0.05 and 0.07. Data were Log(x + 1) transformed to achieve a normal distribution. We used SEM to detect direct and indirect effects of bottom-up and top-down factors for planktonic components and abiotic variables (Online Resource Table A1). Biotic components and abiotic variables in each sampling were used in the analysis. The goodness-of-fit of the model was tested using a Chi-square test and Bollen-Stine bootstrap, due to the small sample size (n = 48, [71]). A non-significant chi-square test indicates that there is no deviation between the observed covariance matrix and the proposed model in SEM, and the model can be accepted. We used the following indexes to decide if the model showed a good fit: (i) a Root Mean Square Error of Approximation (RMSEA) equal to or higher than 0.07 [72]; (ii) Standardized Root Mean Square Residual (SRMR) less than 0.05; (iii) Comparative Fit Index (CFI) equal to or higher than 0.96; and (iv) Tucker-Lewis Index (TLI) equal to or lower than 0.96 [73]. Akaike’s Information Criterion (AIC) was used to determine the most parsimonious model. The SEM graphs were constructed using CytoScape Software. The ANOVA and SEM analyses were performed with the “lavaan” package ([74], http://www.jstatsoft.org/v48/i02/) in R 3.0.3 (R Development Core Team).
We calculated the degree of uncoupling (D) between HB-HNF according to specific empirical models from the literature (below), predicted by a model for observed HB abundance (cell mL−1; equation 6 in [24]):
Results
Water Residence Time and Abiotic Variables
The median water residence times (RT) were significantly higher in Balbina (662 days) and Três Marias (410 days; F (3,44) = 47.31, p < 0.0001) than in Tucuruí (77 days) and Funil (34 days) reservoirs (Table 2). Water temperature (WT) was significantly higher (F (3,42) = 74.18, p < 0.0001) in the Amazonian reservoirs (Balbina, median = 30.8 °C; and Tucuruí, 30.2 °C), compared to Três Marias and Funil, and a slightly acidic median pH (6.3) was recorded in Balbina. DOC concentrations were also significantly higher in Funil (median = 5.33 mg L−1, F (3,41) = 10.23, p < 0.0001; Table 2).
Nutrient concentrations also differed among the reservoirs: TN concentrations were significantly higher in Funil (median = 107.50 μM; F (3,44) = 20.17, p < 0.0001; Table 2; Fig. 2a), and TP was lower in Balbina (median = 0.35 μM; F (3,44) = 15.77, p < 0.0001; Table 2; Fig. 2b). The gradient of trophic states, as expressed in TP concentrations, ranged from oligo-mesotrophic (Balbina) to mesotrophic (Tucuruí and Três Marias) and eutrophic (Funil; Table 2).

Gradients of a total nitrogen; b total phosphorus; c euphotic zone; and d phytoplankton carbon content in two Amazonian Balbina (BALB) and Tucuruí (TUC) and two non-Amazonian Três Marias (TM) and Funil (FUN) reservoirs. The variation of each variable within the reservoirs is expressed by box-and-whisker plots, where the line within boxes indicates the median value, the boxes encompass 25 and 75% and the whiskers 10 and 90%, while the dots encompass 95% of the data. Different letters inside the plots indicate statistically significant differences (p < 0.05)
As expected, Z eu was significantly lower (median = 3.9 m; F (3,42) = 7.31, p = 0.0005; Table 2; Fig. 2c) and DIN was significantly higher (median = 77.0 μM; F (3,44) = 26.59, p < 0.0001; Table 2) in the eutrophic reservoir (Funil). SRP did not differ significantly among the reservoirs (Table 2).
Carbon Content in Pelagic Food-Web Components
The Amazonian Balbina Reservoir showed significantly lower carbon concentrations of all components of the plankton pelagic food web (Fig. 3; for details, see Table A2 in Online Resource 2). The eutrophic Funil Reservoir showed a higher phytoplankton carbon stock (median = 374.72 μg C L−1; F (3,44) = 9.69, p < 0.0001; Fig. 2d; for details, see Table A2 in Online Resource 2) compared with Balbina, Turucuí, and Três Marias, which were not significantly different from each other. Total picoplankton biomass (the sum of HB and PPP) did not differ among the reservoirs, while HB biomass was significantly lower in Balbina compared to Funil (median = 21.9 μg C L−1; F (3,44) = 3.73, p = 0.02). PPP biomass did not differ among the reservoirs (Table A2 in Online Resource 2).

Carbon content in pelagic food-web components of a heterotrophic bacteria (HB) and autotrophic picoplankton (PPP); b heterotrophic nanoflagellates (HNF) and ciliates (Cili); c metazooplankton (Metazoo); and d copepod/cladoceran ratio (COP/CLAD) in Balbina (BALB), Tucuruí (TUC), Três Marias (TM), and Funil (FUN) reservoirs. Graphs A and B illustrate the variation of each variable by bars, where the lines indicate the mean and the whiskers indicate the standard deviation. Graphs C and D illustrate the variation of each variable within the reservoirs, expressed by box-and-whisker plots, where the line within boxes indicates the median value, and the boxes, whiskers, and dots encompass 75, 90, and 95% of the data, respectively. Different letters in plots c and d indicate significant differences (p < 0.05)
Analyzing protozooplankton components, significantly lower HNF (median 8.2 μg C L−1) and ciliate (median 1.9 μg C L−1) carbon stocks were recorded in Balbina Reservoir (F (3,44) = 18.99, p < 0.0001 and F (3,43) = 4.98, p = 0.005, respectively), while no differences were recorded among Tucuruí, Três Marias, and Funil reservoirs (Fig. 3b; Table A2 in Online Resource 2).
Concerning metazooplankton components, rotifers contributed less than 1% to the carbon stock and showed significant differences among the reservoirs (F (3,43) = 6.18, p = 0.001), with the lowest carbon concentration in Balbina (median = 0.02 μg C L−1; Fig. 3c; Table A2 in Online Resource 2) and the highest concentration in Três Marias (median = 1.9 μg C L−1; Fig. 3c; Table A2 in Online Resource 2). Três Marias also had the highest carbon stocks of cladocerans (median = 55.2 μg C L−1) and copepods (median = 386.8 μg C L−1; F (3,44) = 29.00, p < 0.0001 and F (3,44) = 33.9, p < 0.0001, respectively; Table A2 in Online Resource 2).
The carbon stock of the total protozooplankton community (the sum of HNF and Cili, F (3,43) = 17.06, p < 0.0001, see Table A2 in Online Resource 2) and the sum of the microbial food-web components (MFW=HB+PPP+HNF+Cili, F (3,44) = 12.34, p < 0.0001, see Table A2 in Online Resource 2) were significantly lower in Balbina, as was the metazooplankton (the sum of copepods, cladocerans, and rotifers; median = 11.38 μg C L−1; F (3,44) = 31.78, p < 0.0001). The highest metazooplankton carbon stocks were observed in Três Marias (median = 479.58 μg C L−1; Fig. 3c; Table A2 in Online Resource 2). The Amazonian reservoirs Balbina and Tucuruí showed lower COP/CLAD ratios compared to Três Marias and Funil (F (3,43) = 16.87, p < 0.0001; Fig. 3d; Table A2 in Online Resource 2).
The carbon contributions of phytoplankton, zooplankton, and MFW components varied among the waterbodies, but always with a higher contribution of phytoplankton (Fig. 4a). MFW components contributed approximately 30% of the total carbon stock of planktonic biota in the Amazonian reservoirs, a significantly higher proportion compared to Três Marias and Funil (∼10%; p < 0.0001, F (3,44) = 8.98; Fig. 4b; Table A2 in Online Resource 2). The MFW/Metazoo ratio was more variable among the reservoirs and was significantly higher in Balbina and lower in Três Marias (F (3,44) = 20.26, p < 0.0001; Fig. 4c; Table A2 in Online Resource 2). We observed an inverse relationship between the contribution of MFW and the total autotrophic carbon pool, as a proxy of productivity (Fig. 4d; Table A2 in Online Resource 2). The seasonal variation of MFW and the autotrophic contribution to total carbon in each reservoir was not very high (Fig. 4d), except for two periods in Balbina (rainy period, mean Phyto = 41.6 μg C L−1 and %MFW = 52%) and Funil (rainy period, median Phyto = 1,695 μg C L−1 and %MFW = 7.2%).

a Microbial food web (MFW), phytoplankton (PHYTO), and metazooplankton (Metazoo) carbon stocks; b contribution of microbial food web (MFW) to total pelagic carbon; c microbial food web (MFW)/metazooplankton (Metazoo) carbon ratio. The variation of each variable within the reservoirs is expressed by box-and-whisker plots, where the line within boxes indicates the median value, and the boxes, whiskers, and dots encompass 75, 90, and 95% of the data, respectively. Different letters in the plots indicate significant differences (p < 0.05). d Contribution of microbial food web (MFW) along a range of autotrophic carbon levels (as a proxy for trophic state) in Balbina (BALB), Tucuruí (TUC), Três Marias (TM), and Funil (FUN) reservoirs. Error bars indicate standard error. Note that the y-axis in panel c and x-axis in panel d are in log scale
Direct and Indirect Effects of Bottom-Up and Top-Down Interactions
The SEM analysis for bottom-up relationships for each plankton compartment, and also with RT and WT, indicated that there was no significant deviation between the observed covariance matrix and the predicted model (χ 2 = 11.22; df = 9; p = 0.63, AIC = 51.11), and good-fit indices (CFI = 0.99, TLI = 0.89, RMSEA = 0.07, SRMR = 0.032; Fig. 5).

Final structure equation model (SEM) with bottom-up variables, abiotic factors, and planktonic food-web components (χ 2 = 11.22; df = 9; p = 0.63, AIC = 51.11). Solid lines indicate significant effects (p < 0.05). Double lines indicate marginally significant (p < 0.7) effects. Numbers in arrows indicate standardized coefficient estimates. The direction of the arrows goes from the independent to dependent variables. SRP = soluble reactive phosphorus, DIN = dissolved inorganic nitrogen, WT = water temperature, RT = water residence time, PPP = picophytoplankton, HB = heterotrophic bacteria, Phyto = phytoplankton, Cili = ciliates, HNF = heterotrophic nanoflagellates, Rot = rotifers, Clad = cladocerans, Cop = copepods, e = standard errors
SRP showed a negative effect on PPP (p = 0.002) and DIN on HNF (p = 0.005). Phytoplankton (zooplanktonic and HB resource) showed a positive effect on the HB community (p = 0.01), but with no significant effect on zooplankton components (Fig. 5). HNF showed positive effects on rotifers (p = 0.03), cladocerans (p = 0.04), and copepods (p = 0.05), and a marginally significant effect on ciliates (p = 0.06; Fig. 5). RT showed a negative effect on HB (p = 0.03) and HNF (p = 0.03), and a positive effect on rotifers (p = 0.02) and copepods (p = 0.01). WT was significantly related only to HNF, with negative effects (p = 0.04). DOC, TN, TP, and Z eu showed no significant relationship to any biotic component.
With respect to the top-down relationships for each planktonic component, the SEM analysis also indicated no significant deviation between the observed covariance matrix and the predicted model (χ 2 = 3.74; df = 3; p = 0.36, AIC = 303.88), and good-fit indices (CFI = 0.99, TLI = 0.90, RMSEA = 0.07, SRMR = 0.03, Fig. 6). PPP was positively affected by cladocerans (p = 0.04) and negatively by HNF (p = 0.01), while HB was positively affected by cladocerans (p = 0.01). Phytoplankton was negatively affected by cladocerans (p = 0.02), and strongly and positively by copepods (p = 0.002; Fig. 6). HNF positively affected ciliates (p = 0.007), while ciliates were positively affected by copepods (p = 0.007). We also observed a positive indirect effect of copepods on HNF, through ciliates (standard coefficient = 0.51; p = 0.02), and also a positive indirect effect of cladocerans on HB, through ciliates (standard coefficient = 0.32; p = 0.03) and through HNF (standard coefficient = 0.36; p = 0.01). Cladocerans also positively affected PPP through ciliates (standard coefficient = 0.37; p = 0.02) and HNF (standard coefficient = 0.31; p = 0.05) No planktonic community showed a significant relationship to rotifers.

Final structure equation model (SEM) with top-down variables and planktonic food-web components (χ 2 = 3.74; df = 3; p = 0.36, AIC = 303.8). Solid lines indicate significant relationships (p < 0.05). Widths of arrows are proportional to the standardized coefficient estimates (also indicated by numbers). The direction of the arrows goes from the independent to dependent variables. HB = heterotrophic bacteria, PPP = picophytoplankton, Phyto = phytoplankton, Cili = ciliates, HNF = heterotrophic nanoflagellates, Clad = cladocerans, Cop = copepods, e = standard errors
The low degree of uncoupling (D) in all the reservoirs indicates a strong coupling between HB and HNF (Table A2 in Online Resource 2). Although the lowest median D value was recorded in Três Marias, the D values did not differ significantly among the systems.
Discussion
We hypothesized that the microbial food-web components contribute substantially to the carbon pool in these tropical systems, especially in the less-productive systems. The MFW in the four low-latitude reservoirs studied contributed highly to total planktonic carbon, especially in the less-eutrophic Amazonian reservoirs, where this contribution ranged from 20 to 50% of the total planktonic carbon.
Comparing our mean carbon pool and the relative contribution of the biotic components with other systems (Table 3), our four reservoirs had similar MFW and phytoplankton carbon stocks to other subtropical reservoirs and to shallow and deep lakes [7, 75, 76]. Furthermore, the phytoplankton carbon stocks obtained in this study were higher than those found in 55 German lakes with comparable trophic states [33], but within the same range as in other Brazilian reservoirs [37, 77]. It has been suggested that tropical systems might have a different aquatic food-web structure than temperate systems, with higher contributions from PPP and protozoans, lower contributions from daphnids, and higher phytoplankton/zooplankton ratios [12, 16]. However, a meta-analysis considering HB and HNF abundance data, including HNF/HB ratios, from temperate and tropical fresh waters showed that both compartments of MFW were less abundant in the tropical counterparts, although their ratios were similar [29]. The bacterial carbon observed in our study is comparable to mesotrophic systems of German lakes [33], but lower than in other temperate and subtropical systems (Table 3), including Brazilian ones [77]. Furthermore, Tucuruí, Três Marias, and Funil reservoirs contained fewer ciliates but a higher HNF carbon pool, compared to temperate and subtropical systems (Table 3), including subtropical Brazilian reservoirs [78, 79].
The relative contribution of MFW to the carbon pool decreased significantly with the increase in the trophic state, while phytoplankton was the main contributor to the biotic carbon stock in the reservoirs, as also evidenced by the lower MFW/Phyto ratio. This pattern has been observed in subtropical and temperate lakes and reservoirs [21, 33, 77] (Table 3), and also reveals a tendency toward a dominance of autotrophy over heterotrophy with increasing eutrophy [80–82]. The relationship between MFW contribution and phytoplankton may reflect the type of land use and the trophic state [7, 33, 83, 84]. The relative contribution of the MFW carbon stock was higher in the most oligotrophic system, which had the largest forested and the smallest urban areas, unlike the most eutrophic system, where the forested area was lower, with more agricultural and urban areas, concomitant with the higher phytoplankton biomass. This relationship was not as clear in the mesotrophic reservoirs, where the forested and agricultural areas were very similar, and with little difference in the extent of urban areas.
Abiotic Variables and Water Residence Time Regulation
In reservoirs and lakes, the water residence time (RT) is a key variable structuring planktonic communities and affecting their bacterial, phytoplankton, and zooplankton components [77, 85–87]. High RT causes higher phosphorus retention [88] and a loss of DOM reactivity by photodegradation [89], affecting bacterial processes [90] and also affecting HNF carbon sources. This could explain the negative effect of RT on the HB and HNF biomass, with the lowest carbon values recorded in Balbina Reservoir. High flushing rates and low residence times do not favor the development of crustacean zooplankton, which have longer life cycles [86, 91], and may have positive or negative effects on rotifers [86, 92]. Três Marias exhibited a high RT, which showed a positive effect on metazooplankton (copepods and rotifers). However, Balbina, with the highest RT, contained the lowest metazooplankton biomass. In this case, the nutrient limitation in this system might override the response of the zooplankton to the RT. As observed in other systems with different trophic statuses and RTs, such as Funil [77], eutrophication may exert a stronger positive effect on zooplankton biomass, despite the high flushing rate in this reservoir.
Temperature is also an important variable affecting most plankton metabolic processes [86, 93, 94], and our results indicate that temperature was negatively related to HNF biomass. Considering that plankton abundance and carbon stocks were lower in the Amazonian reservoirs, which also had higher mean temperatures and lower nutrient concentrations, it seems that the potential phosphorus limitation on HB and phytoplankton growth could be the main bottom-up limiting factor for these biotic components.
Alongside temperature, nutrients are a main mechanism limiting plankton metabolic processes. The four reservoirs had low SRP concentrations, and P was probably a main potential limiting factor for phytoplankton growth (<0.3 μM; [95]), whereas a similar limitation was not observed for nitrogen, since DIN concentrations were always above 7 μM [93]. This pattern of P limitation for phytoplankton and HB is commonly observed in freshwater aquatic systems [37, 96, 97], where HB seem to be more efficient in taking up P [98, 99]. Furthermore, among the picoplankton community, PPP and HB often also compete for the same resource, mainly P, but PPP may have an advantage over HB since the latter has comparatively less P storage capability [100]. Our results revealed a negative effect of SRP on PPP, in agreement with previous observations [7, 101, 102]. Although we found no significant differences in PPP carbon stock among the reservoirs, the markedly high PPP abundance throughout the year and across productivity gradients also agrees with predictions for tropical systems [12]. The contribution of PPP to total plankton carbon ranged from 6.8% in Funil to 15% in Balbina, showing the importance of PPP for the carbon cycle, mainly in the more oligotrophic systems. It also evidences a highly productive autotrophic carbon source that is available throughout the year to the proto- and microzooplankton (composed of HNF, ciliates, small rotifers, and early life stages of zooplankton) and is often neglected in limnological studies [103].
DIN is also an important source that is assimilated and converted to organic nitrogen by phytoplankton and bacteria, which can be reassimilated by other MFW autotrophic and heterotrophic components [104]. In our study, SEM analysis evidenced a DIN negative effect on HNF. The HNF carbon stock was lowest in Balbina Reservoir, which also had lower DIN concentrations. Moreover, the increase of DIN (as a proxy of eutrophication) also benefits potential predators of HNF, such as metazooplankton (rotifers, cladocerans, and copepods) and ciliates [33], which were positively affected by HNF in the SEM analysis.
Bottom-Up and Top-Down Regulation
The positive relationship between HB and phytoplankton is commonly reported in the literature [30, 96, 105, 106]. This relationship is explained by phytoplankton excretion of labile DOC, which is readily available for HB consumption (e.g., [107]). In addition to furnishing some evidence of a weak positive relationship between HB and phytoplankton in reservoirs in tropical systems [30], a recent study in large African lakes with different trophic states reported a high degree of HB–phytoplankton coupling, due to a higher percentage of extracellular organic carbon released by phytoplankton [31]. The positive relationship evidenced in the SEM analysis also supports this view of close HB–phytoplankton coupling in tropical systems.
PPP is an important food item and carbon source for pelagic ciliates and HNF, which can supply higher trophic levels [76, 108, 109]. Some studies have reported higher ciliate and HNF clearance rates on PPP (herbivory) than on HB (bactivory) [109, 110]. The authors of these studies argued that herbivory represents a more efficient means of energy transfer from primary producers to other parts of the food web and has important biogeochemical implications because of the autotrophic origin of the carbon versus recycled organic carbon. Considering that we found a significant fraction of planktonic carbon in protozooplankton, especially HNF, and also a negative relationship with PPP, it is probable that grazing rates on PPP were considerably high. Moreover, mixotrophy among phytoplankton also has been shown to be an important outcome explaining HB and autotrophic picoplankton losses in freshwater systems [111–113]. The carbon stock of potentially mixotrophic flagellates in the phytoplankton (mostly chrysophyceans, dinoflagellates, and cryptomonads) represented less than 20% of the total phytoplankton carbon stock in our study (6.3% in Funil, 8.9% in Balbina, 13.5% in Tucurui, and 20% in Três Marias) and did not differ significantly among the reservoirs. Ciliates are also important predators of picoplanktonic (autotrophic and heterotrophic) production and a link between matter and energy flux in the planktonic metabolism, and may exceed HNF predation [110, 114–116]. The main morphotype groups that contributed to total ciliate carbon in these reservoirs were Prostomatida, Oligotrichia, Hymenostomatida, Pleurostomatida, Peritrichia, and Gymnostomatida (data not shown), which are planktonic organisms [117–119]. Food items for each group are variable [120], but in general Oligotrichia, Peritrichia, and Hymenostomatida are mainly picoplanktivorous (autotrophic or heterotrophic), while Pleurostomatida, Gymnostomatida, and Protosmatida are omnivorous, herbivorous, or raptorial feeders [76, 116]. Nevertheless, no relationship was evidenced by the SEM analysis. Unfortunately, we could not estimate the species of ciliates and HNF, nor the real effect of bactivory or herbivory of mixotrophic flagellates on the picoplanktonic community, since for this, direct measurements of grazing rates would be necessary. However, we speculate that not only the HB community but also the PPP components were probably affected by the predation pressure of the protozoan community.
The HB carbon stock was significantly lower in Balbina than in Tucuruí, and did not differ between the other reservoirs. HB abundance generally varies less than other planktonic food-web components across environments [121], independently of the zooplankton community structure [122]. In most oligotrophic environments, HB abundance tends to be more top-down regulated by HNF, while bottom-up or controlled by metazoan predation in more eutrophic systems [121]. Since Balbina is the least eutrophic system studied, and showed a high relative contribution of MFWcomponents (%MFW) and cladocerans (low COP/CLAD ratio), the HB community might be experiencing a stronger top-down regulation in this reservoir than in the more eutrophic ones.
Zooplankton can also interact directly through predation with different trophic levels of MFW [12]. We did not observe a high contribution of rotifers to the consumer community in any of the four reservoirs. Although fish can control zooplankton and influence the size structure of the planktonic community [34, 123], few species of fishes occupy the upper layers of limnetic areas and deep layers of large reservoirs, and most of the ichthyofauna occurs in the littoral zone [124, 125]. Nevertheless, the main feeding guilds of fishes in these reservoirs are the piscivores (in all reservoirs and especially in Três Marias), detritivores, and omnivores (especially in Funil) [47–49]. A fish community composed mainly of piscivores will strongly impact other fish, and consequently the zooplankton and phytoplankton structure [123], including reduction of cladocerans and dominance of cyanobacteria [126].
With respect to the other metazooplankton groups, we observed two clear clusters of COP/CLAD ratios, with higher values in Três Marias and Funil reservoirs, which can establish different relationships between the classical and MFW. Copepods are selective feeders, preferring larger prey than the picoplankton spectrum [20], and may not be as effective in HNF predation [127]. On the other hand, cladocerans are effective predators on phytoplankton [93] and transport energy more efficiently through a direct link between the MFW and the classical food chain [128, 129]. Our analysis captured a positive direct effect of copepods on ciliates and phytoplankton biomass, and also an indirect positive effect on HNF. Likewise, cladocerans had positive direct and indirect effects on HB and PPP, but a negative effect on phytoplankton. Previous studies have reported positive relationships between phytoplankton and zooplankton in reservoirs [37, 130]. In a gradient of ultraoligotrophic to mesotrophic conditions, calanoid copepods, rotifers, copepodites, and nauplii became more effective than cladocerans in consuming protozoans, particularly ciliates [7, 131]. Moreover, laboratory experiments with Daphnia and copepods also found that copepods strongly impact ciliates [122]. All these studies concord with the results obtained in our models of the coupling between metazoans and protozoans.
Usually, the degree of HNF-HB uncoupling (D) is expected to be higher in daphnid-dominated than in copepod-dominated systems [26]. In this study, we did not observe significant differences in D values among the reservoirs, but our results showed rather low values compared to other studies [28, 121], especially in Três Marias where the highest copepod biomass was recorded. Conversely, HNF/HB carbon ratios equal to or higher than 1.0 were recorded in 50% of our samples, especially in reservoirs with higher COP/CLAD ratios. These values were more frequent than in similar reports from temperate lakes [26]. These observations suggest a strong HNF-HB coupling in tropical reservoirs. Despite this, we found no significant direct effect of HNF on HB, which may be influenced by the low variances of HB and HNF among the reservoirs. However, the positive relationship between copepods and ciliates and the significant positive indirect effect of copepods on HNF through ciliates provide empirical evidence for the higher relative importance of HNF in copepod-dominated food webs [122, 128, 132].
We also observed a positive relationship between HNF and ciliates, which can be explained by the overlap of controlling factors on the two communities, either by source supply or by predation [7, 33, 133]. Furthermore, the recycled nutrients provided by protozoans (mainly N, P, and DOC) probably stimulate bacterial growth [132]. This constant nutrient and carbon recycling, allied to the fact that bacterial respiration rates per bacterial production unit in tropical freshwaters are at least twofold higher in the tropics [32], may contribute to the CO2 supersaturation that is commonly observed in tropical reservoirs [4].
In summary, the high contribution of the MFW, especially HNF, to the carbon pool, and the strong HNF-HB coupling as measured by the low D values support the view that MFW components may participate actively in the metabolic process in tropical reservoirs. Besides, the MFW may be directly and indirectly influenced through different pathways by cladocerans and copepods. The biotic pelagic components of classical and MFW were more closely related to resources (inferred from the lowest AIC model test), configuring an important bottom-up mechanism. The food-web structure, especially top predators, can alter the efficiency of energy transfer through trophic levels [9, 12, 121]. In turn, the MFW exhibits many pathways and intermediate trophic levels, which may contribute to a higher energy loss through respiration [108]. High plankton respiration rates and inefficient food webs [32, 134] may help to account for the large greenhouse gas emissions in tropical reservoirs [4, 6]. Therefore, this study demonstrates the importance of evaluating the whole size spectrum (including the microbial compartments) of the different planktonic compartments, in order to better capture the complex carbon-flux dynamics of tropical aquatic ecosystems.
References
Cole JJ, Prairie YT, Caraco NF et al (2007) Plumbing the global carbon cycle: integrating inland waters into the terrestrial carbon budget. Ecosystems 10:171–184. doi:10.1007/s10021-006-9013-8
Aufdenkampe AK, Mayorga E, Raymond PA et al (2011) Riverine coupling of biogeochemical cycles between land, oceans, and atmosphere. Front Ecol Environ 9:53–60. doi:10.1890/100014
Lewis WM Jr (2010) Biogeochemistry of tropical lakes. Verh Internat Verein Limnol 30:1595–1603
Tranvik LJ, Downing JA, Cotner JB et al (2009) Lakes and reservoirs as regulators of carbon cycling and climate. Limnol Oceanogr 54:2298–2314. doi:10.4319/lo.2009.54.6_part_2.2298
Kosten S, Roland F, Da Motta Marques DML et al (2010) Climate-dependent CO2 emissions from lakes. Glob Biogeochem Cycles 24, GB2007. doi:10.1029/2009GB003618
Barros NO, Cole JJ, Tranvik LJ et al (2011) Carbon emission from hydroelectric reservoirs linked to reservoir age and latitude. Nat Geosci 4:593–596. doi:10.1038/ngeo1211
Burns CW, Galbraith LM (2007) Relating planktonic microbial food web structure in lentic freshwater ecosystems to water quality and land use. J Plankton Res 29:127–139. doi:10.1093/plankt/fbm001
Kilham P, Kilham SS (1990) Endless summer: internal loading processes dominate nutrient cycling in tropical lakes. Freshw Biol 23:379–390. doi:10.1111/j.1365-2427.1990.tb00280.x
Schindler DE, Carpenter SR, Cole JJ et al (1997) Influence of food web structure on carbon exchange between lakes and the atmosphere. Science 277:248–251. doi:10.1126/science.277.5323.248
Atwood TB, Hammill E, Greig HS et al (2013) Predator-induced reduction of freshwater carbon dioxide emissions. Nat Geosci 6:191–194. doi:10.1038/ngeo1734
Müller-Navarra DC (2008) Food web paradigms: the biochemical view on trophic interactions. Int Rev Hydrobiol 93:489–505. doi:10.1002/iroh.200711046
Sarmento H (2012) New paradigms in tropical limnology: the importance of the microbial food web. Hydrobiologia 686:1–14. doi:10.1007/s10750-012-1011-6
Sarmento H, Amado AM, Descy J-P (2013) Climate change in tropical fresh waters (comment on the paper “Plankton dynamics under different climatic conditions in space and time” by de Senerpont Domis et al., 2013). Freshw Biol 58:2208–2210. doi:10.1111/fwb.12140
Iglesias C, Mazzeo N, Meerhoff M et al (2011) High predation is of key importance for dominance of small-bodied zooplankton in warm shallow lakes: evidence from lakes, fish exclosures and surface sediments. Hydrobiologia 667:133–147. doi:10.1007/s10750-011-0645-0
Kruk C, Segura AM, Peeters ETHM et al (2012) Phytoplankton species predictability increase towards warmer regions. Limnol Oceanogr 57:1126–1135. doi:10.4319/lo.2012.57.4.1126
Fernando CH (1994) Zooplankton, fish and fisheries in tropical freshwaters. Hydrobiologia 272:105–123. doi:10.1007/BF00006516
Lewis WM Jr (1996) Tropical lakes: how latitude makes a difference. In: Schiemer F, Boland KT (eds) Perspect Trop Limnol. SPB Academic Publishing, Amsterdam, pp 43–64
Jeppesen E, Meerhoff M, Holmgren K et al (2010) Impacts of climate warming on lake fish community structure and potential effects on ecosystem function. Hydrobiologia 646:73–90. doi:10.1007/s10750-010-0171-5
Jeppesen E, Meerhoff M, Jacobsen BA et al (2007) Restoration of shallow lakes by nutrient control and biomanipulation—the successful strategy varies with lake size and climate. Hydrobiologia 581:269–285. doi:10.1007/s10750-006-0507-3
Sommer U, Sommer F (2006) Cladocerans versus copepods: the cause of contrasting top-down controls on freshwater and marine phytoplankton. Oecologia 147:183–194. doi:10.1007/s00442-005-0320-0
Porter KG, Paerl HW, Hodson R et al (1988) Microbial interactions in lake food webs. In: Carpenter SR (ed) Complex interact lake communities. Springer, New York, pp 209–227
Dumont HJ (1994) On the diversity of the Cladocera in the tropics. Hydrobiologia 272:27–38. doi:10.1007/BF00006510
Sherr EB, Sherr BF (2002) Significance of predation by protists in aquatic microbial food webs. Antonie Van Leeuwenhoek 81:293–308. doi:10.1023/A:1020591307260
Sherr EB, Sherr BF (1994) Bacterivory and herbivory: key roles of phagotrophic protists in pelagic food webs. Microb Ecol 28:223–235. doi:10.1007/BF00166812
Gasol JM (1994) A framework for the assessment of top-down vs bottom-up control of heterotrophic nanoflagellate abundance. Mar Ecol Prog Ser 113:291–300. doi:10.3354/meps113291
Gasol JM, Simons AM, Kalff J (1995) Patterns in the top-down versus bottom-up regulation of heterotrophic nanoflagellates in temperate lakes. J Plankton Res 17:1879–1903. doi:10.1093/plankt/17.10.1879
Tadonléké RD, Pinel-Alloul B, Bourbonnais N, Pick FR (2004) Factors affecting the bacteria-heterotrophic nanoflagellate relationship in oligo-mesotrophic lakes. J Plankton Res 26:681–695. doi:10.1093/plankt/fbh060
Fermani P, Diovisalvi N, Torremorell A et al (2013) The microbial food web structure of a hypertrophic warm-temperate shallow lake, as affected by contrasting zooplankton assemblages. Hydrobiologia 714:115–130. doi:10.1007/s10750-013-1528-3
Segóvia BT, Domingues CD, Meira BR et al (2016) Coupling between heterotrophic nanoflagellates and bacteria in fresh waters: does latitude make a difference? Front Microbiol 7:1–11. doi:10.3389/fmicb.2016.00114
Roland F, Lobão LM, Vidal LO et al (2010) Relationships between pelagic bacteria and phytoplankton abundances in contrasting tropical freshwaters. Aquat Microb Ecol 60:261–272. doi:10.3354/ame01429
Morana C, Sarmento H, Descy J-P et al (2014) Production of dissolved organic matter by phytoplankton and its uptake by heterotrophic prokaryotes in large tropical lakes. Limnol Oceanogr 59:1364–1375. doi:10.4319/lo.2014.59.4.1364
Amado AM, Meirelles-Pereira F, Vidal LDO et al (2013) Tropical freshwater ecosystems have lower bacterial growth efficiency than temperate ones. Front Microbiol 21(4):167. doi:10.3389/fmicb.2013.00167
Auer B, Elzer U, Arndt H (2004) Comparison of pelagic food webs in lakes along a trophic gradient and with seasonal aspects: influence of resource and predation. J Plankton Res 26:697–709. doi:10.1093/plankt/fbh058
Stockner JG, Porter KG (1988) Microbial food webs in freshwater planktonic ecosystems. In: Carpenter SR (ed) Complex interact. Lake communities. Springer-Verlag New York Inc, pp 70–83
Agência Nacional de Energia Elétrica A (2014) Projeto BALCAR: Emissões de Gases de Efeito Estufa em Reservatórios de Centrais Hidrelétricas. ANEEL - Agência Nacional de Energia Elétrica, Rio de Janeiro
Soares MCS, Marinho MM, Huszar VLM et al (2008) The effects of water retention time and watershed features on the limnology of two tropical reservoirs in Brazil. Lakes Reserv Res Manag 13:257–269. doi:10.1111/j.1440-1770.2008.00379.x
Rangel LM, da Silva LHS, Rosa PG et al (2012) Phytoplankton biomass is mainly controlled by hydrology and phosphorus concentrations in tropical hydroelectric reservoirs. Hydrobiologia 693:13–28. doi:10.1007/s10750-012-1083-3
Kemenes A, Forsberg BR, Melack JM (2011) CO2 emissions from a tropical hydroelectric reservoir (Balbina, Brazil). J Geophys Res Biogeosci 116, G03004. doi:10.1029/2010JG001465
Tundisi JG, Santos MA, Menezes CFS (2005) Tucurui reservoir: experience and lessons learned brief. (Lake Basin Manag Initiat 1–20.) Available at: www.worldlakes.org/uploads/26_Tucurui_Reservoir_27February2006.pdf
Esteves FDA, Amorim JC, Cardoso EL, Barbosa FAR (1985) Caracterização limnológica preliminar da represa de Três Marias (MG) com base em alguns parâmetros ambientais básicos. Ciênc Cult 37:608–617
La Rovere EL, Mendes FE (2000) Tucuruí Hydropower Complex Tucuruí Hydropower Complex, Brazil, A WCD case study prepared as an input to the World Commission on Dams, Cape Town
Soares MCS, Marinho MM, Azevedo SMFO et al (2012) Eutrophication and retention time affecting spatial heterogeneity in a tropical reservoir. Limnologica 42:197–203. doi:10.1016/j.limno.2011.11.002
da Silva CC, Ferreira EJG, de Deus CP (2008) Diet of Bryconops alburnoides and B. caudomaculatus (Osteichthyes: Characiformes) in the region affected by Balbina Hydroelectric Dam (Amazon drainage, Brazil). Neotropical Ichthyol 6:237–242. doi:10.1590/S1679-62252008000200011
Guérin F, Abril G, Richard S et al (2006) Methane and carbon dioxide emissions from tropical reservoirs: significance of downstream rivers. Geophys Res Lett 33, L21407. doi:10.1029/2006GL027929
Rosas FCW, de Mattos GE, Cabral MMM (2007) The use of hydroelectric lakes by giant otters Pteronura brasiliensis: Balbina lake in central Amazonia, Brazil. Oryx 41:520–524. doi:10.1017/S0030605307005121
ANEEL (Agência Nacional de Energia Elétrica) (2002) Atlas de energia elétrica do Brasil. ANEEL, Brasília
Gomes LC, Fernandes R, Luiz EA (2004) Development of reservoir fisheries management in Brazil based on imported paradigms. Acta Sci Biol Sci 26:309–315
Becker B, Galhardo BDOS, Macedo DR et al (2016) Influence of limnological zones on the spatial distribution of fish assemblages in three Brazilian reservoirs. J Limnol 75:156–168. doi:10.4081/jlimnol.2015.1266
Santos ABI, Terra BDF, Araújo FG (2010) Influence of the river flow on the structure of fish assemblage along the longitudinal gradient from river to reservoir. Zoologia 27:732–740. doi:10.1590/S1984-46702010000500010
Cole GA (1994) Textbook of limnology. Waveland Press Inc, Long Grove, Illinois
Wetzel RG, Likens GE (1991) Limnological analyses. Springer, New York
Nürnberg GK (1996) Trophic state of clear and colored, soft- and hardwater lakes with special consideration of nutrients, anoxia, phytoplankton and fish. Lake Reservoir Manage 12:432–447. doi:10.1080/07438149609354283
Uhelingher V (1964) Summary for policymakers. In: Intergovernmental Panel on Climate Change (ed) Climate change 2013—physical science basis. Cambridge University Press, Cambridge, pp 1–30
Utermöhl H (1958) Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Mitt Int Ver Theor Angew Limnol 9:1–38
Van den Hoeck C, Mann DG, Jahns HM (1997) Algae. An introduction to phycology. Cambridge University Press, Cambridge
Komárek J, Anagnostidis K (1999) Cyanoprokaryota I. Teil Chroococcales. I. In: Ettl H, Gärtner G, Heynig H, Mollenhauer D (eds) Süsswasserflora von Mitteleuropa. Gustav Fischer, Stuttgart, p 548
Komárek J, Anagnostidis K (2005) Cyanoprocaryota. 2. Oscillatoriales. In: Ettl H, Gärtner G, Heynig H, Mollenhauer D (eds) Süsswasserflora von Mittleuropa. Gustav Fisher, Stuttgart, p 759
Hillebrand H, Dürselen C-D, Kirschtel D et al (1999) Biovolume calculation for pelagic and benthic microalgae. J Phycol 35:403–424. doi:10.1046/j.1529-8817.1999.3520403.x
Rocha O, Duncan A (1985) The relationship between cell carbon and cell volume in freshwater algal species used in zooplanktonic studies. J Plankton Res 7:279–294
Putt M, Stoecker DK (1989) An experimentally determined carbon: volume ratio for marine “oligotrichous” ciliates from estuarine and coastal waters. Limnol Oceanogr 34:1097–1103. doi:10.4319/lo.1989.34.6.1097
Muller H, Geller W (1993) Maximum growth rates of aquatic ciliated protozoa: the dependence on body size and temperature reconsidered. Arch Hydrobiol 126:315–327
Porter KG, Feig YS (1980) The use of DAPI for identifying and counting aquatic microflora. Limnol Oceanogr 25:943–948
Maclsaac EA, Stockner GS (1993) Enumeration of phototrophic picoplankton by autoflorescence microscopy. In: Kemp PF, Sherr BF, Sherr EB, Cole JJ (eds) The handbook of methods in aquatic microbial ecology. Lewis Publishers, Boca Raton, pp 187–197
Kirchman DL (1993) Statistical analysis of direct counts of microbial abundance. In: Kemp PF, Sherr BF, Sherr EB, Cole JJ (eds) The handbook of methods in aquatic microbial ecology. Lewis Publishers, Boca Raton, pp 117–120
Weisse T (1993) Dynamics of autotrophic picoplankton in marine and freshwater ecosystems. In: Jones JG (ed) Advances in microbial ecology. Plenum Press, New York, pp 327–370
Massana R, Gasol JM, Bjørnsen PK (1997) Measurement of bacterial size via image analysis of epifluorescence preparations: description of an inexpensive system and solutions to some of the most common problems. Sci Mar 61:397–407. doi:10.1017/CBO9781107415324.004
Norland S (1993) The relationship between biomass and volume of bacteria. In: Kemp PF, Sherr BF, Sherr EB, Cole JJ (eds) The handbook of methods in aquatic microbial ecology. Lewis Publishers, Boca Raton, pp 303–307
Borsheim KY, Bratbak G (1987) Cell volume to cell carbon conversion factors for a bacterivorous Monas sp. enriched from seawater. Mar Ecol Prog Ser 36:171–175. doi:10.3354/meps036171
Culver DA, Boucherle MM, Bean DJ, Fletcher JW (1985) Biomass of freshwater crustacean zooplankton from length–weight regressions. Can J Fish Aquat Sci 42:1380–1390. doi:10.1139/f85-173
Latja R, Salomen K (1978) Carbon analysis for determination of individual biomasses of planktonic animals. Verh Internat Verein Limnol 20:2556–2560
Bollen KA, Stine RA (1992) Bootstrapping goodness-of-fit measures in structural equation models. Sociol Methods Res 21:205–229. doi:10.1177/0049124192021002004
Steiger JH (2007) Understanding the limitations of global fit assessment in structural equation modeling. Personal Individ Differ 42:893–898. doi:10.1016/j.paid.2006.09.017
Hu L, Bentler PM (1999) Cutoff criteria for fit indexes in covariance structure analysis: conventional criteria versus new alternatives. Struct Equ Model Multidiscip J 6:1–55. doi:10.1080/10705519909540118
Rosseel Y (2014) Structural Equation Modeling with lavaan. 1–128
Havens KE, Beaver JR, East TL (2007) Plankton biomass partitioning in a eutrophic subtropical lake: comparison with results from temperate lake ecosystems. J Plankton Res 29:1087–1097. doi:10.1093/plankt/fbm083
Šimek K, Macek M, Pern J et al (1996) Can freshwater planktonic ciliates survive on a diet of picoplankton? J Plankton Res 18:597–613. doi:10.1093/plankt/18.4.597
Silva LHS, Huszar VLM, Marinho MM et al (2014) Drivers of phytoplankton, bacterioplankton, and zooplankton carbon biomass in tropical hydroelectric reservoirs. Limnologica 48:1–10. doi:10.1016/j.limno.2014.04.004
Pereira DG, Velho LFM, Pagioro TA, Lansac-Tôha FA (2005) Abundância de nanoflagelados heterotróficos no plâncton de reservatórios com distintos graus de trofia. Acta Sci Biol Sci 27:43–50
Velho LFM, Pereira DG, Pagioro TA et al (2005) Abundance, biomass and size structure of planktonic ciliates in reservoirs with distinct trophic states. Acta Limnol Bras 17:361–371
Gaedke U, Kamjunke N (2006) Structural and functional properties of low- and high-diversity planktonic food webs. J Plankton Res 28:707–718. doi:10.1093/plankt/fbl003
del Giorgio PA, Gasol JM (1995) Biomass distribution in freshwater plankton communities. Am Nat 146:135–152
Cotner JB, Biddanda BA (2002) Small players, large role: microbial influence on biogeochemical processes in pelagic aquatic ecosystems. Ecosystems 5:105–121. doi:10.1007/s10021-001-0059-3
Katsiapi M, Mazaris AD, Charalampous E, Moustaka-Gouni M (2012) Watershed land use types as drivers of freshwater phytoplankton structure. Hydrobiologia 698:121–131. doi:10.1007/s10750-012-1095-z
Galbraith LM, Burns CW (2010) Drivers of ciliate and phytoplankton community structure across a range of water bodies in southern New Zealand. J Plankton Res 32:327–339. doi:10.1093/plankt/fbp134
Lindström ES, Agterveld MPK, Zwart G (2005) Distribution of typical freshwater bacterial groups is associated with pH, temperature, and lake water retention time. Appl Environ Microbiol 71:8201–8206. doi:10.1128/AEM.71.12.8201
Obertegger U, Flaim G, Braioni MG et al (2007) Water residence time as a driving force of zooplankton structure and succession. Aquat Sci 69:575–583. doi:10.1007/s00027-007-0924-z
Beaver JR, Jensen DE, Casamatta DA et al (2013) Response of phytoplankton and zooplankton communities in six reservoirs of the middle Missouri River (USA) to drought conditions and a major flood event. Hydrobiologia 705:173–189. doi:10.1007/s10750-012-1397-1
Straškraba M (1999) Retention time as a key variable of reservoir limnology. In: Tundisi TG, Straškraba M (eds) Theoretical reservervoir ecology and its applications. Brazilian Academy and Backhuys Publishers, São Carlos, pp 358–410
Mari X, Rochelle-Newall E, Torréton J-P et al (2007) Water residence time: a regulatory factor of the DOM to POM transfer efficiency. Limnol Oceanogr 52:808–819. doi:10.4319/lo.2007.52.2.0808
Amaral JHF, Suhett AL, De Melo S, Farjalla VF (2013) Seasonal variation and interaction of photodegradation and microbial metabolism of DOC in black water Amazonian ecosystems. Aquat Microb Ecol 70:157–168. doi:10.3354/ame01651
Pace ML, Findlay SEG, Lints D (1992) Zooplankton in advective environments: the Hudson River community and a comparative analysis. Can J Fish Aquat Sci 49:1060–1069. doi:10.1139/f92-117
Basu BK, Pick FR (1996) Factors regulating phytoplankton and zooplankton biomass in temperate rivers. Limnol Oceanogr 41:1572–1577
Reynolds CS (2006) Ecology of phytoplankton. Cambridge University Press, New York. doi:10.1017/CBO9781107415324.004
Sarmento H, Montoya JM, Vazquez-Dominguez E et al (2010) Warming effects on marine microbial food web processes: how far can we go when it comes to predictions? Philos Trans R Soc Lond B Biol Sci 365:2137–2149. doi:10.1098/rstb.2010.0045
Sasai Y (1989) Lake restoration by reduction of nutrient loading: expectations, experiences, extrapolations. Academia Verlag Richarz, St. Augustin
Artigas J, Soley S, Pérez-Baliero MC et al (2012) Phosphorus use by planktonic communities in a large regulated Mediterranean river. Sci Total Environ 426:180–187. doi:10.1016/j.scitotenv.2012.03.032
Farjalla VF, Esteves FDA, Bozelli RL, Roland F (2002) Nutrient limitation of bacterial production in clear water Amazonian ecosystems. Hydrobiologia 489:197–205. doi:10.1023/A:1023288922394
Bratbak G, Thingstad TF (1985) Phytoplankton-bacteria interactions: an apparant paradox? Analysis of a model system with both competition and commensalism. Mar Ecol Prog Ser 25:23–30. doi:10.3354/meps025023
Mindl B, Sonntag B, Pernthaler J et al (2005) Effects of phosphorus loading on interactions of algae and bacteria: reinvestigation of the ‘phytoplankton-bacteria paradox’ in a continuous cultivation system. Aquat Microb Ecol 38:203–213. doi:10.3354/ame038203
Brett MT, Lubnow FS, Villar-Argaiz M et al (1999) Nutrient control of bacterioplankton and phytoplankton dynamics. Aquat Ecol 33:135–145. doi:10.1023/A:1009998915078
Callieri C, Stockner JG (2002) Freshwater autotrophic picoplankton: a review. J Limnol 61:1–14. doi:10.4081/jlimnol.2002.1
Takamura N, Nojiri Y (1994) Picophytoplankton biomass in relation to lake trophic state and the TN:TP ratio of lake water in Japan. J Phycol 30:439–444
Sarmento H, Unrein F, Isumbisho M et al (2008) Abundance and distribution of picoplankton in tropical, oligotrophic Lake Kivu, eastern Africa. Freshw Biol 53:756–771. doi:10.1111/j.1365-2427.2007.01939.x
McCarthy MJ, Lavrentyev PJ, Yang L et al (2007) Nitrogen dynamics and microbial food web structure during a summer cyanobacterial bloom in a subtropical, shallow, well-mixed, eutrophic lake (Lake Taihu, China). Hydrobiologia 581:195–207. doi:10.1007/s10750-006-0496-2
Baines SB, Pace ML (1991) The production of dissolved organic matter by phytoplankton and its importance to bacteria: patterns across marine and freshwater systems. Limnol Oceanogr 36:1078–1090
Jankowski K, Schindler DE, Horner-Devine MC (2014) Resource availability and spatial heterogeneity control bacterial community response to nutrient enrichment in lakes. PLoS ONE 9:e86991. doi:10.1371/journal.pone.0086991
Sarmento H, Gasol JM (2012) Use of phytoplankton-derived dissolved organic carbon by different types of bacterioplankton. Environ Microbiol 14:2348–2360. doi:10.1111/j.1462-2920.2012.02787.x
Pirlot S, Unrein F, Descy J, Servais P (2007) Fate of heterotrophic bacteria in Lake Tanganyika (East Africa). Microb Ecol 62:354–364. doi:10.1111/j.1574-6941.2007.00396.x
Tarbe A-L, Unrein F, Stenuite S et al (2011) Protist Herbivory: a key pathway in the pelagic food web of lake Tanganyika. Microb Ecol 62:314–323. doi:10.1007/s00248-011-9817-8
Šimek K, Hartman P, Nedoma J et al (1997) Community structure, picoplankton grazing and Zooplankton control of heterotrophic nanoflagellates in a eutrophic reservoir during the summer phytoplankton maximum. Aquat Microb Ecol 12:49–63. doi:10.3354/ame012049
Jansson M, Blomqvist P, Jonsson A, Bergstrøm A-K (1996) Nutrient limitation of bacterioplankton, autotrophic and mixotrophic phytoplankton, and heterotrophic nanoflagellates in Lake Örträsket. Limnol Oceanogr 41:1552–1559. doi:10.4319/lo.1996.41.7.1552
Gerea M, Saad JEF, Izaguirre I et al (2015) Presence, abundance and bacterivory of the mixotrophic algae Pseudopedinella (Dictyochophyceae) in freshwater environments. Aquat Microb Ecol 76:219–232. doi:10.3354/ame01780
Saad JEF, Unrein F, Tribelli PM et al (2016) Influence of lake trophic conditions on the dominant mixotrophic algal assemblages. J Plankton Res 38:818–829. doi:10.1093/plankt/fbw029
Azam F, Fenchel T, Field JG et al (1983) The ecological role of water-column microbes in the sea. Mar Ecol Prog Ser 10:257–263. doi:10.3354/meps010257
Callieri C, Heinimaa S (1997) Microbial loop in the large subalpine lakes. Mem Ist Ital Idrobiol 56:143–156
Šimek K, Jürgens K, Nedoma J et al (2000) Ecological role and bacterial grazing of Halteria spp.: small freshwater oligotrichs as dominant pelagic ciliate bacterivores. Aquat Microb Ecol 22:43–56. doi:10.3354/ame022043
Lansac-Tôha FA, Velho LFM, Perenha MCZ et al (2004) Abundance of planktonic ciliates in a cascading reservoirs of the Paranapanema River, Brazil. Acta Sci Biol Sci 26:407–413
Pfister G, Auer B, Arndt H (2002) Pelagic ciliates (Protozoa, Ciliophora) of different brackish and freshwater lakes—a community analysis at the species level. Limnol Ecol Manag Inland Waters 32:147–168. doi:10.1016/S0075-9511(02)80005-6
Beaver JR, Crisman TL (1989) The role of ciliated protozoa in pelagic freshwater ecosystems. Microb Ecol 17:111–136. doi:10.1007/BF02011847
Foissner W, Berger H (1996) A user-friendly guide to the ciliates (Protozoa, Ciliophora) commonly used by hydrobiologists as bioindicators in rivers, lakes, and waste waters, with notes on their ecology. Freshw Biol 35:375–482
Gasol JM, Pedrós-Alió C, Vaqué D (2002) Regulation of bacterial assemblages in oligotrophic plankton systems: results from experimental and empirical approaches. Antonie Van Leeuwenhoek 81:435–452. doi:10.1023/A:1020578418898
Zöllner E, Santer B, Boersma M et al (2003) Cascading predation effects of Daphnia and copepods on microbial food web components. Freshw Biol 48:2174–2193. doi:10.1046/j.1365-2426.2003.01158.x
Carpenter SR, Kitchell JF, Hodgson JR (1985) Cascading trophic interactions and lake productivity. Bioscience 35:634–639
Araújo-Lima CARM, Agostinho AA, Fabré NN (1995) Trophic aspects of fish communities in Brazilian rivers and reservoirs. In: Tundisi JG, de Bicudo CEM, Matsumura-Tundisi T (eds) Limnology in Brazil. ABC/SBL, Rio de Janeiro, pp 105–136
Agostinho AA, Pelicice FM, Gomes LC (2008) Dams and the fish fauna of the Neotropical region: impacts and management related to diversity and fisheries. Braz J Biol 68:1119–1132. doi:10.1590/S1519-69842008000500019
Pinto-Coelho RM, Bezerra-Neto JF, De Miranda FP et al (2008) The inverted trophic cascade in tropical plankton communities: impacts of exotic fish in the Middle Rio Doce lake district, Minas Gerais, Brazil. Braz J Biol 68:1025–1037. doi:10.1590/S1519-69842008000500010
Jürgens K, Wickham SA, Rothhaupt KO, Santer B (1996) Feeding rates of macro- and microzooplankton on heterotrophic nanoflagellates. Limnol Oceanogr 41:1833–1839
Jürgens K, Jeppesen E (2000) The impact of metazooplankton on the structure of the microbial food web in a shallow, hypertrophic lake. J Plankton Res 22:1047–1070. doi:10.1093/plankt/22.6.1047
Burns CW, Schallenberg M (1996) Relative impacts of copepods, cladocerans and nutrients on the microbial food web of a mesotrophic lake. J Plankton Res 18:683–714. doi:10.1093/plankt/20.8.1501
Bonecker CC, Nagae MY, Bletller MCM et al (2007) Zooplankton biomass in tropical reservoirs in southern Brazil. Hydrobiologia 579:115–123. doi:10.1007/s10750-006-0391-x
Burns CW, Schallenberg M (2001) Calanoid copepods versus cladocerans: consumer effects on protozoa in lakes of different trophic status. Limnol Oceanogr 46:1558–1565. doi:10.4319/lo.2001.46.6.1558
Jürgens K, Matz C (2002) Predation as a shaping force for the phenotypic and genotypic composition of planktonic bacteria. Antonie Van Leeuwenhoek 81:413–434. doi:10.1023/A:1020505204959
Auer B, Arndt H (2001) Taxonomic composition and biomass of heterotrophic flagellates in relation to lake trophy and season. Freshw Biol 46:959–972. doi:10.1046/j.1365-2427.2001.00730.x
Brown JH, Gillooly JF, Allen AP et al (2004) Toward a metabolic theory of ecology. Ecology 85:1771–1789. doi:10.1890/03-9000
Acknowledgments
We are grateful to Gladson Rezende Marques for laboratory analyses, and to Carlos Henrique Eckhardt Duque-Estrada and Ramôn Pereira de Souza for fieldwork assistance. We are also grateful to Inácio Domingos (Universidade Federal do Rio de Janeiro) and Roberto Dias (Universidade Federal de Juiz de Fora) for assistance in determining morphotypes in the ciliate community, and to Janet Reid for English review. This study was funded by the Centrais Elétricas do Norte do Brasil S/A (ELETRONORTE). We also thank the Electrical Energy Research Center (Cepel), Eletrobrás Amazonas Energia, Eletrobrás Eletronorte/Tucuruí staff, Waimiri-Atroari Program, and Uatumã Biological Reserve (REBio-Uatumã) for logistical support in Balbina and Tucuruí reservoirs. C.D.D. is grateful to the Brazilian National Council for Scientific and Technological Development (CNPq) for a PhD fellowship. H.S.’s work was supported by CNPq and the Foundation for Research Support of the State of São Paulo (FAPESP, process 2014/14139-3).
Author information
Authors and Affiliations
Corresponding author
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 20 kb)
Rights and permissions
About this article

Cite this article
Domingues, C.D., da Silva, L.H.S., Rangel, L.M. et al. Microbial Food-Web Drivers in Tropical Reservoirs. Microb Ecol 73, 505–520 (2017). https://doi.org/10.1007/s00248-016-0899-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-016-0899-1