
Abstract
Mercury (Hg), particularly as methylmercury (MeHg), is a nonessential, persistent, and bioaccumulative toxic element with high biomagnification capacity and is considered a threat to marine environments. We evaluated total Hg concentrations in liver, kidney, and brain in 62 individuals of 9 bird species linked to marine ecosystems from western Mediterranean admitted in a Wildlife Rehabilitation Center (WRC) (Alicante, Spain, 2005–2020). Age- and sex-related differences in Hg levels, as well as the cause of admission to the WRC, were also evaluated in certain species. The species studied were: northern gannet (Morus bassanus), European shag (Phalacrocorax aristotelis), great cormorant (Phalacrocorax carbo), osprey (Pandion haliaetus), Balearic shearwater (Puffinus mauretanicus), yellow-legged gull (Larus michahellis), razorbill (Alca torda), common tern (Sterna hirundo), and black-headed gull (Chroicocephalus ridibundus). Concentrations in feathers of 27 individuals, and concentrations in internal tissues in 7 other individuals of 7 different species were also reported but not statistically evaluated due to the limited number of samples. Results suggest that individuals were chronically exposed to Hg through diet. The differences in Hg concentrations among species may be explained by their diet habits. Mercury concentrations strongly correlated between tissues (r = 0.78–0.94, p < 0.001, n = 61–62). Some individuals of certain species (i.e., European shag, northern gannet, and great cormorant) showed Hg concentrations close to or above those described in the literature as causing reproductive alterations in other avian species. Consequently, certain individuals inhabiting western Mediterranean could be at risk of suffering long-term, Hg-related effects. Some of the species evaluated are listed within different categories of threat according to the International Union for Conservation of Nature (IUCN) and are endangered at a national level, so this study will provide valuable information for assessors and authorities in charge of the management of the environment and pollution.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Marine ecosystems are threatened by pollutants such as mercury (Hg), especially in its organic form as methylmercury (MeHg), a persistent, bioaccumulative, and toxic, nonessential element that is distributed worldwide (Cherel et al. 2018; Kenney et al. 2018). Natural processes and anthropogenic activities participate in the continuous release of Hg into the environment (Kenney et al. 2018; Ruus et al. 2015), which enters marine ecosystems mostly through wet and dry atmospheric deposition processes and runoff from industrial emissions (Carravieri et al. 2018; Ishii et al. 2017; Zamani-Ahmadmahmoodi et al. 2010). In the marine environment, the inorganic Hg is methylated and converted into MeHg, the most toxic and bioavailable form (Cherel et al. 2018; Kenney et al. 2018; Ruus et al. 2015). Methylmercury is assimilated by phytoplankton and zooplankton, becoming part of the food chain, where it bioaccumulates and biomagnifies as the trophic level increases. Consequently, top predators, such as seabirds achieve higher concentrations of this contaminant in their organs and tissues (Carravieri et al. 2018; Misztal-Szkudlinska et al. 2018).
Methylmercury is a neurotoxic and endocrine disruptor element that also alters behaviour, reproductive success, nestlings’ growth and development, metabolism, and immune responses (Carravieri et al. 2018; Fort et al. 2015; García-Fernández 2014), affecting principally reproduction in seabirds (Carravieri et al. 2018). Against this toxicity, organisms have protective mechanisms, such as the synthesis and binding to metallothioneins (MT), demethylation and formation of nontoxic complexes with selenium, or MeHg elimination through moult (Espín et al. 2012, 2016). These processes seem to be particularly effective in seabirds, explaining the tolerance of these predators to higher Hg concentrations compared with other bird species (García-Fernández 2014).
It is essential to conduct Hg biomonitoring studies in wildlife inhabiting marine ecosystems and, for this purpose, seabirds and other piscivorous birds (e.g., osprey, Pandion haliaetus) are considered good bioindicators of Hg-polluted marine environments because they are long-lived species, they bioaccumulate MeHg in their organism, and they are in a high trophic position in the food web (Carravieri et al. 2018; Espín et al. 2012; García-Fernández 2014; García-Fernández et al. 2020; Kojadinovic et al. 2007; Moura et al. 2018a; Ribeiro et al. 2009). In this sense, collecting tissues from birds that have died in massive mortality events or from dead specimens stored at Research or Wildlife Rehabilitation Centers may provide interesting data to examine Hg concentrations and the relationships between internal tissues in a broad range of species (Espín et al. 2012; Fort et al. 2015; Mallory et al. 2018).
The Mediterranean is a semiclosed sea with restricted water exchange and surrounded by industrialized countries, which entails a greater risk of Hg contamination (Espín et al. 2012; Pereira et al. 2019). However, data are scarce about the concentrations of this metal in certain bird species of the western Mediterranean. The purpose of this study was to evaluate the exposure to Hg in different seabird and aquatic bird species linked to marine ecosystems in eastern Spain. The specific objectives are: (1) to provide data on total Hg concentrations in liver, kidney, and brain of different seabird and aquatic bird species as well as in feathers of some individuals, and (2) to assess differences in total Hg concentrations among nine species, as well as between sexes, age groups and causes of admission in the Wildlife Rehabilitation Center (WRC) for four species where a sufficient number of samples was available. Based on the available literature, we hypothesize that larger species, as well as male and adult individuals, will present higher Hg concentrations. In addition, we expect to find higher Hg concentrations in internal tissues of those specimens suffering non-traumatic pathologies (i.e., individuals with symptoms of undernutrition due to infectious or parasitic diseases).
Materials and Methods
Species and Study Area
In this study, Hg exposure was evaluated in 62 individuals of 9 species of birds linked to marine ecosystems: 13 European shags (Phalacrocorax aristotelis), 13 yellow-legged gulls (Larus michahellis), 12 northern gannets (Morus bassanus), 8 great cormorants (Phalacrocorax carbo), 5 razorbills (Alca torda), 3 common terns (Sterna hirundo), 3 Balearic shearwaters (Puffinus mauretanicus), 3 osprey (Pandion haliaetus), and 2 black-headed gulls (Chroicocephalus ridibundus). Table 1 reports the main characteristics of these 9 species, including their habitat, diet, body weight, and conservation status. Mercury concentrations in 7 individuals of 7 other different species are also reported: Atlantic puffin (Fratercula arctica), ruddy turnstone (Arenaria interpres), Audouin’s gull (Ichthyaetus audouinii), Mediterranean gull (Ichthyaetus melanocephalus), Scopoli’s shearwater (Calonectris diomedea), little tern (Sternula albifrons), and grey heron (Ardea cinerea). Data for those 7 species where only one individual was available are presented for information purposes but are not included in the statistics nor discussed due to limitations in number of samples. All of these animals were found dead or injured along the Occidental Mediterranean coastline, at different locations in the province of Alicante, and were admitted in the WRC of Santa Faz (Alicante, eastern Spain; Fig. 1) between 2005 and 2020. The causes of admission in the WRC were trauma, drowning, fish-hook ingestion, electrocution, entanglement in fishing line and fishing net, and undernutrition as a result of other pathologies (e.g., infectious diseases).

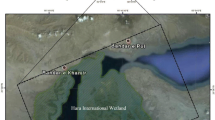
Map of the sampling area (coastline of Alicante, Spain). Three individuals were found in different locations in the province of Valencia (i.e., Montaverner, Oliva, and Valencia; not shown in map)
Sampling
Necropsies of the 69 individuals were performed in the WRC. A total of 206 samples of liver (n = 69), kidney (n = 68, no kidney sample was retained in a cormorant individual), and brain (n = 69) were collected in Eppendorf tubes, transported under cold conditions to the Toxicology laboratory at the University of Murcia, and stored frozen at −20 °C until analysis. Sterile Eppendorf tubes were used so that there was no possibility of contamination from the containers. Back feathers were only collected in 27 individuals and were kept in sterile sealed plastic bags at room temperature. In most cases, the age (n = 63 individuals), sex (n = 61), and body mass (n = 55) of the individuals were recorded. Age was determined through plumage patterns and morphological criteria and sex by direct visualization of the gonads during the necropsy.
Mercury Analysis
Total Hg (hereafter Hg) was analysed using a Milestone DMA-80 direct Hg analyser based on atomic absorption spectrophotometry, with a detection limit of 0.005 ng. Each sample (0.05 g wet weight for internal tissues and 0.005 g dry weight for feathers) was loaded in a quartz boat. The precision and accuracy of the method were previously evaluated using certified reference material (CRM; TORT-2, lobster hepatopancreas, National Research Council Canada), and blanks were also run in each sample set. A recovery percentage of 108.9 ± 4.1% (mean ± standard deviation, SD) and a coefficient of variation for repeatability of 3.7% were obtained. Feathers were washed using distilled water, Milli-Q water, and acetone before analytical determination to remove external contamination from the surface.
The percentage of humidity of the internal tissues was calculated in an Infrared Moisture Analyser MA35 (Sartorius) in order to express the results of total Hg in both wet weight (ww) and dry weight (dw) and compare them with other published studies.
Statistical Analysis
The results obtained were analysed using the IBM SPSS v.24 statistical package. A descriptive statistical analysis was performed by obtaining the mean ± SD and median (min–max) Hg concentrations. Species with only one individual available (n = 7 species; Table 2) and results from feathers (27 samples from 11 species; Table 2) were excluded to perform statistical tests and discuss results due to limitations in number of samples. The normality of the variables was tested using a Kolmogorov–Smirnov test and Hg concentrations in liver, kidney, and brain were log-transformed, obtaining a normal distribution after the transformation. ANOVA followed by Tukey’s tests for multiple comparison were performed to test significant differences in Hg concentrations between tissues and species (n = 9 species; Table 2). The relationships between the Hg concentrations in liver, kidney, and brain and their correlation with body mass were tested using Pearson’s correlation coefficient. For those species where male, female, juvenile, and adult individuals were available, as well as different causes of admission to the WRC (n = 4 species, i.e., Morus bassanus, Phalacrocorax aristotelis, Phalacrocorax carbo, and Larus michahellis), ANOVA was used to test differences in Hg concentrations according to sex, age and cause of admission. The causes of admission were classified into two groups, based on the probability to be related to loss of body mass: (1) traumatic type entry, which included trauma, drowning, hook ingestion, fishing line entanglement, and fishing net entanglement, and (2) nontraumatic type entry, which included individuals with symptoms of undernutrition as a result of other pathologies (e.g., parasitic or infectious diseases). For all analyses, the level of significance was set at p ≤ 0.05.
Results
Hg concentrations in liver, kidney, brain, and feathers for the different study species are shown in Table 2, and Hg concentrations reported in internal tissues of the same species in some publications are provided in Table 3 for comparison purposes. Mercury concentrations differed significantly between the nine species for the three internal tissue types (ANOVA test for liver: F = 10.09, kidney: F = 9.5 and brain: F = 7.8, p < 0.001; Table 2; Fig. 2). Tukey’s test results comparing Hg concentrations among species within each sample type show that, in general, northern gannet, European shag, and great cormorant—the greater species—were the ones that presented the highest Hg concentrations (mean Hg in liver: 7.16, 14.56, and 5.65 µg/g ww, respectively; Table 2), coinciding with our initial hypothesis. The osprey was the next species with the highest Hg concentration (mean Hg in liver: 1.98 µg/g ww; Table 2) but did not show significant differences with the rest of the species except for yellow-legged gull, razorbill, common tern, and black-headed gull in brain (Table 2). Mercury levels also differed among tissues (ANOVA test: F = 46.7, p < 0.001). Tukey’s test showed no differences between liver and kidney (p = 0.386), whereas the concentrations in these tissues were significantly higher than those found in the brain (p < 0.001) for the nine species studied (Table 2). For these nine species, the mean ratio of Hgliver:Hgkidney was 1.03 (0.63–1.61, n = 62; coefficient of variation, CV, of 26%), reflecting that liver and kidney values were similar, whereas the ratio of Hgliver:Hgbrain was 8.28 (2.25–14.86, n = 62, CV 70%) similar to the ratio of Hgkidney:Hgbrain (8.01, 1.87–16.45, n = 62, CV 87%), showing the higher Hg levels in the liver and kidney compared with the brain.

Log mercury concentrations (± SE; µg/g, wet weight) in liver, kidney, and brain of the species studied. MB Morus bassanus; PA Phalacrocorax aristotelis; PC Phalacrocorax carbo; PH Pandion haliaetus; PM Puffinus mauretanicus; LM Larus michahellis; AT Alca torda; SH Sterna hirundo; CR Chroicocephalus ridibundus
Pearson’s correlation coefficients showed that Hg concentrations in tissues were positively correlated with the body mass of the individuals (r Hg liver–Body mass = 0.450, r Hg kidney–Body mass = 0.537, r Hg brain–Body mass = 0.565, p < 0.005, n = 48). In addition, strong significant positive correlations were observed for Hg concentrations between tissues (rliver–kidney = 0.937, rliver–brain = 0.787, rkidney–brain = 0.784, p < 0.001, n = 61–62; Fig. 3).

Correlations between (log) mercury concentrations (µg/g, wet weight) in tissues of 9 wild bird species (rliver–kidney = 0.937, rliver–brain = 0.787, rkidney–brain = 0.784, p < 0.001, n = 61–62). AT Alca torda; CR Chroicocephalus ridibundus; LM Larus michahellis; MB Morus bassanus; PA Phalacrocorax aristotelis; PC Phalacrocorax carbo; PH Pandion haliaetus, PM Puffinus mauretanicus; SH Sterna hirundo
Differences in Hg concentrations according to sex, age, and cause of admission were evaluated in four species (i.e., northern gannet, European shag, great cormorant, and yellow-legged gulls). Adult European shags showed significantly higher Hg concentrations in liver and kidney than juvenile birds (F = 35.3 and 68.9, respectively, p < 0.001), as expected, whereas the opposite trend was found in the three tissue types in yellow-legged gulls (F = 28.5, 35.0, and 16.6 in liver, kidney, and brain, respectively, p < 0.003; Fig. 4). Sex-related differences were only observed in yellow-legged gulls. Females had lower Hg concentrations in tissues than males (F = 8.0, p = 0.018 in liver; F = 8.8, p = 0.014 in kidney; F = 5.8, p = 0.037 in brain; Fig. 4), which is in line with the literature data. Finally, significant differences in Hg concentrations according to the cause of admission to WRC were only found in liver for northern gannets (F = 6.3, p = 0.033) and European shags (F = 6.7, p = 0.029), birds suffering nontraumatic pathologies showing higher hepatic Hg concentrations than birds admitted due to traumatic causes (Fig. 5), which was expected according to our hypothesis.

Mean (and 95% CI) mercury concentrations (µg/g, w.w.) in tissues of Larus michahellis by age and gender. *Significant differences between ages and sexes were found in the three tissue types (p < 0.05)

Mean (and 95% CI) (log) mercury concentration (µg/g, w.w.) in liver of Morus bassanus and Phalacrocorax aristotelis according to cause of admission to the WRC (traumatic or nontraumatic). Significant differences according to the cause of admission were found in both species (p < 0.05)
Discussion
Tissue Hg Concentrations and Interspecific Differences
The pattern of Hg distribution in tissues of nine species linked to marine ecosystems was similar to other studies: liver ≥ kidney > brain (Table 3). Chronic exposure to Hg entails a balance in concentrations between compartments in the body, which explains the distribution pattern observed and the strong correlations found between Hg concentrations in liver, kidney, and brain (Fig. 3). The distribution of Hg in different organs depends on the form of Hg to which the individual is exposed, and the ratio of Hg in kidney and liver may be used to distinguish a chronic exposure to MeHg or inorganic Hg (Scheuhammer 1987). Thus, a kidney:liver ratio markedly greater than 1 reflects an exposure to inorganic Hg, whereas a ratio close to 1 (and < 2) is characteristic of MeHg exposure. In this study, the kidney:liver ratio was within the range 0.62–1.58 (mean ratio: 0.88) depending on the species, probably reflecting that the individuals evaluated were mainly exposed to MeHg. This is consistent with the fact that almost 100% of the total Hg detected in muscle of different fish species was in the form of MeHg (Scheuhammer 1987).
Several factors may explain variations in Hg concentrations between species (Table 2), some of them interspecific, such as detoxification capacity, size, diet, or migratory habits, and others intraspecific, such as age, sex, or body condition (Moura et al. 2018a; Ramos et al. 2013). One of the main factors that determine the interspecific differences in the pollutant load in the organism is the diet, being the main route of Hg exposure in marine vertebrates (Carravieri et al. 2018; Kojadinovic et al. 2007; Moura et al. 2018b; Ribeiro et al. 2009). Although the study area is an essential factor to consider due to the potential differences in Hg contamination, it has not been discussed in this study, because all individuals were found dead or injured along the Occidental Mediterranean coastline. Also, the exact origin of the migratory individuals before their arrival to the coast of Alicante is unknown. Despite this, it should be considered that the origin could partly explain the differences in Hg concentrations found in certain species. This may be critical in some cases, and an approach to relate Hg concentrations in an abiotic matrix with those in bird tissues is recommended for future studies (e.g., the Biota Sediment Accumulation Factor, Calle et al. 2015).
In general, the species studied are mainly piscivorous, which means that they are exposed to higher Hg levels than species with different diet habits, since fish accumulate high levels of this metal, especially as MeHg (Kojadinovic et al. 2007). Depending on the species of fish they ingest, they will be exposed to a different Hg amount. Demersal and benthic fish have higher Hg concentrations than pelagic fish because they occupy higher trophic levels and are closer to the bottom sediments (Arcos et al. 2002; Vizuete et al. 2018). The study species that presented a larger size (northern gannet, European shag, and great cormorant) showed the highest Hg concentrations (Table 2), which was supported by a positive correlation between Hg concentrations in tissues and body mass. This could be due to the consumption of larger prey, which can contain higher Hg levels than smaller prey of the same species (Zamani-Ahmadmahmoodi et al. 2014). Although the northern gannet feeds on pelagic fish, it ingests larger prey than the cormorants by feeding farther from the coast. Also, it selectively looks for places where it can take advantage of trawl fishery discards (BirdLife International 2018; Hamer et al. 2000; Kubetzki et al. 2009), which may lead to greater Hg exposure, because birds can consume species that they cannot access in a natural way, such as demersal or benthic fish. One of its main prey is the Atlantic mackerel (Scomber scombrus), a large size fish (215–455 mm) that feeds on plankton but mostly on smaller fish as its size increases, being more exposed to Hg than other fish species (Hamer et al. 2000; Olaso et al. 2005). High Hg levels in cormorants (i.e., European shag and great cormorant) can also be explained by the diving capacity of both species, which allows them to feed on benthic fish (Arcos et al. 2002; BirdLife International 2018, Misztal-Szkudlinska et al. 2018). The osprey feeds exclusively on fish, and the Balearic shearwater takes advantage of commercial fishery discards and ingests pelagic fish but feeds on smaller prey, so less Hg exposure can be expected (BirdLife International 2018; Louzao et al. 2012). Both species showed slightly (but not significant for most tissues) lower Hg concentrations that northern gannets, European shags, and great cormorants. Although the diet of razorbills and common terns are mainly based on fish, they presented lower Hg concentrations than the northern gannet and European shag, probably because these species ingest smaller and pelagic prey (BirdLife International 2018; Szostek and Becker 2015). Some fish included in the diet of these species are sardines (Sardina pilchardus) for razorbills and also anchovies (Engraulis encrasicolus) in the case of common terns, being small and pelagic fish species that can be found in the Mediterranean Sea (Costalago et al. 2015; Espín et al. 2012; Szostek and Becker 2015). Sardines present a size < 250 mm and anchovies from 10 to 130 mm. They mainly feed on phytoplankton and zooplankton, respectively, so they occupy a low trophic level (Borme et al. 2009; Costalago et al. 2015; Tudela and Palomera 1997). In addition, the common tern ingests mostly juvenile fish, so they are expected to accumulate a smaller amount of Hg (Szostek and Becker 2015). The yellow-legged and black-headed gulls showed lower Hg levels than European shags and cormorants, probably because they are opportunistic species also ingesting terrestrial and freshwater food, which have less Hg load than prey of marine origin (BirdLife International 2018; Ramos et al. 2013; Vizuete et al. 2018). In future studies, it would be interesting to analyse the stable isotope Nitrogen 15 (15δN) to determine the trophic level of each study species so that a comparison of Hg concentrations versus the trophic position can be made.
In general, Hg concentrations found in liver, kidney, and brain were similar to or lower than those observed in the same species from other countries, particularly for razorbill, osprey, black-headed gull, or Balearic shearwater (Table 3). However, for certain species (mainly northern gannet, European shag, and great cormorant) concentrations found in this study were higher than levels reported in the literature (Table 3).
Mercury concentrations in internal tissues are a key indicator of bioaccumulation. Measuring both liver and kidney simultaneously can provide information on the nature of exposure (i.e., chronic exposure to MeHg or inorganic Hg). Threshold concentrations (mainly in liver and kidney) associated with adverse effects in birds have been suggested for interpretation (Espín et al. 2016). However, due to ethical and legal reasons, sampling is generally possible where carcasses are found in the field or injured animals are euthanasied for welfare reasons. In addition, metabolism, demethylation and health condition (starvation versus healthy individuals) can influence the balance (e.g., remobilization of Hg) and alter tissue Hg concentrations. On the other hand, feathers are considered a good matrix for Hg determination since they can be obtained as moulted feathers, from carcasses or be plucked without permanently damaging the bird, being a minimally invasive matrix. Moreover, MeHg is uniformly deposited in feathers and they are a more stable matrix. However, this deposition only occurs during feather growth, reflecting Hg concentration in blood during this period, while internal tissues maintain a continuous exchange with blood, so they provide updated information even though Hg levels are affected by changes in diet and/or fat mobilization. In addition, feathers can be contaminated on the surface (although Hg external contamination is typically small), and moulting periods and patterns are different among species (Espín et al. 2016), which may pose some difficulties when comparing results. Although Hg concentrations were also analysed in feathers from some species in this study, a proper statistical analysis could not be done due to limitations in the number of samples. Mercury concentrations in feathers of most species were, in general, similar to those reported in other studies (Arcos et al. 2002; Cotín et al. 2012; Mazloomi et al. 2008; Misztal-Szkudlinska et al. 2012; Monteiro et al. 1999; Moreno et al. 2013; Otero et al. 2018; Paiva et al. 2008; Rumbold et al. 2001; Sanpera et al. 2008; Szumiło-Pilarska et al. 2016, 2017; Zolfaghari et al. 2009), whereas they were lower in the case of osprey (Cahill et al. 1998; DesGranges et al. 1998; Lounsbury-Billie et al. 2008) and higher for northern gannet, black-headed gull, Audouin’s gull and Scopoli’s shearwater (Arcos et al. 2002; Goutner et al. 2000, 2013; Mendes et al. 2008; Monteiro et al. 1995, 1999; Nardiello et al. 2019).
Sex, Age, and Cause of Admission to WRC
In this study, the influence of sex, age, and cause of admission on Hg exposure was evaluated in four species (i.e., northern gannet, European shag, great cormorant, and yellow-legged gulls). Sex-related differences in tissue Hg concentrations were only found in yellow-legged gulls, females showing lower Hg levels compared to males (Fig. 4). Different studies (Ishii et al. 2017; Vizuete et al. 2018) have demonstrated that, in adult individuals, females have lower Hg levels than males justified by their excretion capacity through egg laying. Regarding age differences, adult European shags showed higher Hg concentrations in liver and kidney than juvenile birds. Several authors agree that adult individuals have higher Hg concentrations than juveniles of the same species because of the greater accumulation of Hg in their body during their life (Moura et al. 2018b; Ribeiro et al. 2009; Saeki et al. 2000; Vizuete et al. 2018). However, the opposite trend was found in yellow-legged gulls in this study (Fig. 4), which might be due to their opportunistic diet habits (Table 1) and a different diet source between juvenile and adult birds. However, further studies with higher number of samples would be needed to better evaluate these sex and age-related differences.
Northern gannets and European shags suffering non-traumatic pathologies (i.e., specimens with symptoms of undernutrition as a result of pathologies such as infectious or parasitic diseases) showed higher hepatic Hg concentrations than birds admitted to the WRC due to traumatic causes (Fig. 5). In this regard, Sanpera et al. (2008) have observed that dehydrated individuals, with poor body condition and a state of weakness had higher Hg concentrations in their tissues as a result of a general redistribution of metals in the organs. Further studies with a larger number of samples within each cause of admission type would be necessary in order to evaluate deeply the effect of the cause of admission on Hg concentrations in the study species.
Risk Assessment
In the majority of cases, the individuals studied showed Hg concentrations below the critical levels related to reproductive disturbances in black ducks (Anas rubripes) (i.e., reduced egg production, hatchability, and survival of ducklings; liver: 23 µg/g, ww; kidney: 16 µg/g, ww; brain: 3.79 µg/g, ww; Finley and Stendell 1978) or marked behavioural changes in pigeons (i.e., declined rate of pecking, changes in posture and coordination; brain: 12–16 µg/g, Evans et al. 1982). However, two individuals of European shag showed tissue concentrations exceeding those critical levels in liver and kidney (liver: 27.94 and 110.57 µg/g, ww; kidney: 28.40 and 134.17 µg/g, ww; brain: 0.57 and 1.92 µg/g, ww). A northern gannet (liver: 19.33 µg/g, ww; kidney: 10.13 µg/g, ww; brain: 2.97 µg/g, ww) and a great cormorant (kidney: 16.06 µg/g, ww; liver: 10.58 µg/g, ww; brain: 0.53 µg/g, ww) had concentrations close to that levels. In addition, all the species studied showed mean hepatic Hg levels similar to or higher than those associated with altered behaviour and decreased reproductive success in laboratory-reared ducklings (liver: 1–2 µg/g, ww; reviewed by Zillioux et al. 1993). It is clear that these comparisons should be interpreted with caution due to the interspecific differences in tolerance to contaminants. In addition, total Hg is not the best indicator of toxic effects, and more importance should be given to the more toxic form, MeHg concentrations (Wolfe et al. 1998). However, these results suggest that Hg concentrations in the marine ecosystems of the western Mediterranean could constitute a risk situation for certain seabird individuals, especially for endangered species (at national level), such as the European shag, with only 49–55 breeding pairs in the Valencian Community in 2018 (D. G. Medi Natural i Avaluació Ambiental 2018), or the northern gannet under Special Protection in Spain (Table 1).
Conclusions
The results of this study suggest that individuals of nine bird species linked to marine ecosystems found dead in the western Mediterranean coasts were chronically exposed to MeHg. Mercury concentrations differed among species, which can be explained by their different dietary habits. In general, Hg concentrations found are similar to or higher than those reported in other studies worldwide. Some individuals of certain species (i.e., European shag, northern gannet, and great cormorant) showed Hg concentrations close to or higher than those described in the literature as causing reproductive alterations in other avian species. These comparisons should be made with caution due to the possible difference in sensitivity between species. However, our results suggest that certain individuals inhabiting marine ecosystems in the western Mediterranean could be at risk of suffering long-term, Hg-related effects on physiology, reproduction, and behaviour. Some of the species evaluated are listed within different categories of threat according to the International Union for Conservation of Nature (IUCN) (including Near Threatened and Critically Endangered species) and are endangered at a national level, so this study will provide valuable information for risk assessors and authorities in charge of the management of the environment and pollution. Further studies with a greater number of specimens of each species are necessary to better evaluate the effect of sex, age, and cause of admission to WRCs on Hg concentrations in the study species. The cause of admission to the WRC is essential, because it helps to relate the Hg concentrations found with the history and symptoms of the individuals. Therefore, this factor should be described and evaluated in future research.
Availability of Data and Materials
Available upon request.
References
Aazami J, KianiMehr N (2017) Survey of heavy metals in internal tissues of great cormorant collected from southern wetlands of Caspian Sea, Iran. Environ Monitor Assess 190(1):52
Aazami J, Esmaili-Sari A, Bahramifar N, Ghasempouri M, Savabieasfahani M (2011) Mercury in liver, kidney, feather and muscle of seabirds from major wetlands of the Caspian Sea, Iran. Bull Environ Contam Toxicol 86(6):657
Arcos JM, Ruiz X, Bearhop S, Furness RW (2002) Mercury levels in seabirds and their fish prey at the Ebro Delta (NW Mediterranean): the role of trawler discards as a source of contamination. Mar Ecol Progr Ser 232:281–290
BirdLife International (2018) The IUCN red list of threatened species. https://www.iucnredlist.org/. Consulted 14 May 2019
BOE (Boletín Oficial del Estado) (2011) Real Decreto 139/2011, de 4 de febrero, para el desarrollo del Listado de Especies Silvestres en Régimen de Protección Especial y del Catálogo Español de Especies Amenazadas. Ministerio de Medio Ambiente, y Medio Rural y Marino, 46:20912–20951
Borme D, Tirelli V, Brandt SB, Umani SF, Arneri E (2009) Diet of Engraulis encrasicolus in the northern Adriatic Sea (Mediterranean): ontogenetic changes and feeding selectivity. Mar Ecol Progr Ser 392:193–209
Braune BM (1987) Comparison of total mercury levels in relation to diet and molt for nine species of marine birds. Arch Environ Contam Toxicol 16(2):217–224
Cahill TM, Anderson DW, Elbert RA, Perley BP, Johnson DR (1998) Elemental profiles in feather samples from a mercury-contaminated lake in Central California. Arch Environ Contam Toxicol 35(1):75–81
Calle P, Alvarado O, Monserrate L, Cevallos JM, Calle NL, Alava JJ (2015) Mercury accumulation in sediments and seabird feathers from the Antarctic Peninsula. Mar Pollut Bull 91:410–417
Carbonell G, Bravo J, Torija C, López-Beceiro A, Fidalgo L, Hernandez-Moreno D, Soler F, Pérez-López M (2007) Contenido hepático de mercurio y plomo en cormorán moñudo (Phalacrocorax aristotelis) y alcatraz atlántico (Morus bassanus) procedentes de las costas de Galicia (España). Rev Toxicol 24(1):31–35
Carravieri A, Fort J, Tarroux A, Cherel Y, Love OP, Prieur S, Brault-Favrou M, Bustamante P, Descamps S (2018) Mercury exposure and short-term consequences on physiology and reproduction in Antarctic petrels. Environ Pollut 237:824–831
Cherel Y, Barbraud C, Lahournat M, Jaeger A, Jaquemet S, Wanless RM, Phillips RA, Thompson DR, Bustamante P (2018) Accumulate or eliminate? Seasonal mercury dynamics in albatrosses, the most contaminated family of birds, Environ Pollut 241:124–135
Costa RA, Torres J, Vingada JV, Eira C (2016) Persistent organic pollutants and inorganic elements in the Balearic shearwater Puffinus mauretanicus wintering off Portugal. Mar Pollut Bull 108:311–316
Costalago D, Garrido S, Palomera I (2015) Comparison of the feeding apparatus and diet of European sardines Sardina pilchardus of Atlantic and Mediterranean waters: ecological implications. J Fish Biol 86(4):1348–1362
Cotín J, García-Tarrasón M, Jover L, Sanpera C (2012) Are the toxic sediments deposited at Flix reservoir affecting the Ebro river biota? Purple heron eggs and nestlings as indicators. Ecotoxicology 21(5):1391–1402
Delbeke K, Joiris C, Decadt G (1984) Mercury contamination of the Belgian avifauna 1970–1981. Environ Pollut Ser B Chem Phys 7(3):205–221
DesGranges J-L, Rodrigue J, Tardif B, Laperle M (1998) Mercury accumulation and biomagnification in ospreys (Pandion haliaetus) in the James Bay and Hudson Bay Regions of Québec. Arch Environ Contam Toxicol 35(2):330–341
Direcció General de Medi Natural i d’Avaluació Ambiental (2018) Resultats del cens de corb marí emplomallat Phalacrocorax aristotelis a la Comunitat Valenciana. Any 2018. Generalitat Valenciana, Conselleria d’Agricultura, Medi Ambient, Canvi Climàtic i Desenvolupament Rural
Espín S, Martínez-López E, Gómez-Ramírez P, María-Mojica P, García-Fernández AJ (2012) Razorbills (Alca torda) as bioindicators of mercury pollution in the southwestern Mediterranean. Mar Pollut Bull 64(11):2461–2470
Espín S, García-Fernández AJ, Herzke D, Shore RF, van Hattum B, Martínez-López E, Coeurdassier M, Eulaers I, Fritsch C, Gómez-Ramírez P, Jaspers VLB, Krone O, Duke G, Helander B, Mateo R, Movalli P, Sonne C, van den Brink NW (2016) Tracking pan-continental trends in environmental contamination using sentinel raptors—what types of samples should we use? Ecotoxicology 25(4):777–801
Evans HL, Garman RH, Laties VG (1982) Neurotoxicity of methylmercury in the pigeon. Neurotoxicology 3(3):21–36
Finley MT, Stendell RC (1978) Survival and reproductive success of black ducks fed methyl mercury. Environ Pollut (1970) 16(1):51–64
Fort J, Lacoue-Labarthe T, Nguyen HL, Boué A, Spitz J, Bustamante P (2015) Mercury in wintering seabirds, an aggravating factor to winter wrecks? Sci Total Environ 527:448–454
Fundación CRAM (2019) CRAM. Fundación para conservación y recuperación de los animales marinos. Catálogo de especies. https://cram.org/catalogo-de-especies/aves-marinas/. Accessed 6 Apr 2019
García-Fernández AJ (2014) Ecotoxicology, Avian. In: Wexler P (ed) Encyclopedia of toxicology, vol 2, 3rd edn. Elsevier, London, pp 289–294
García-Fernández AJ, Espín S, Gómez-Ramírez P, Martínez-López E, Navas I (2020) Wildlife sentinels for human and environmental health hazards in ecotoxicological risk assessment. In: Roy Kunal (ed) Ecotoxicological QSARs, methods in pharmacology and toxicology. Springer Protocols, Humana Press, New York, pp 77–94
Goutner V, Furness RW, Papakonstantinou K (2000) Mercury in feathers of Audouin’s gull (Larus audouinii) chicks from northeastern Mediterranean colonies. Arch Environ Contam Toxicol 39(2):200–204
Goutner V, Becker PH, Liordos V (2011) Organochlorines and mercury in livers of great cormorants (Phalacrocorax carbo sinensis) wintering in northeastern Mediterranean wetlands in relation to area, bird age, and gender. Sci Total Environ 409(4):710–718
Goutner V, Becker PH, Liordos V (2013) Low mercury contamination in Mediterranean gull Larus melanocephalus chicks in Greece. Chem Ecol 29(1):1–10
Häkkinen I, Häsänen E (1980) Mercury in eggs and nestlings of the osprey (Pandion haliaetus) in Finland and its bioaccumulation from fish. Ann Zool Fenn 17(3):131–139
Hamer K, Phillips R, Wanless S, Harris M, Wood A (2000) Foraging ranges, diets and feeding locations of gannets Morus bassanus in the North Sea: evidence from satellite telemetry. Mar Ecol Progr Ser 200:257–264
Hemb JG (2019) Nord University. BirdID. https://www.birdid.no/bird/index.php. Accessed 6 Apr 2019
Hopkins WA, Hopkins LB, Unrine JM, Snodgrass J, Elliot JD (2007) Mercury concentrations in tissues of osprey from the Carolinas, USA. J Wildl Manag 71(6):1819–1829
Horai S, Watanabe I, Takada H, Iwamizu Y, Hayashi T, Tanabe S, Kuno K (2007) Trace element accumulations in 13 avian species collected from the Kanto area, Japan. Sci Total Environ 373(2):512–525
Houserová P, Hedbavny J, Matejicek D, Kracmar S, Sitko J, Kuban V (2005) Determination of total mercury in muscle, intestines, liver and kidney tissues of cormorant (Phalacrocorax carbo), great crested grebe (Podiceps cristatus) and Eurasian buzzard (Buteo buteo). Vet Med Czech 50:61–68
Houserová P, Kubáň V, Kráčmar S, Sitko J (2007) Total mercury and mercury species in birds and fish in an aquatic ecosystem in the Czech Republic. Environ Pollut 145(1):185–194
Ishii C, Ikenaka Y, Nakayama SM, Mizukawa H, Yohannes YB, Watanuki Y, Fukuwaka M, Ishizuka M (2017) Contamination status and accumulation characteristics of heavy metals and arsenic in five seabird species from the central Bering Sea. J Vet Med Sci 79(4):807–814
Kalisinska E, Gorecki J, Lanocha N, Okonska A, Melgarejo JB, Budis H, Rzad I, Golas J (2014) Total and methylmercury in soft tissues of white-tailed eagle (Haliaeetus albicilla) and osprey (Pandion haliaetus) collected in Poland. Ambio 43(7):858–870
Kenney LA, Kaler RS, Kissling ML, Bond AL, Eagles-Smith CA (2018) Mercury concentrations in multiple tissues of Kittlitz’s murrelets (Brachyramphus brevirostris). Mar Pollut Bull 129(2):675–680
Kitowski I, Kowalski R, Komosa A, Sujak A (2015) Total mercury concentration in the kidneys of birds from Poland. Turkish J Zool 39(4):693–701
Kojadinovic J, Le Corre M, Cosson RP, Bustamante P (2007) Trace elements in three marine birds breeding on Reunion Island (western Indian Ocean): part 1—Factors influencing their bioaccumulation. Arch Environ Contam Toxicol 52(3):418–430
Kral T, Blahova J, Doubkova V, Farkova D, Vecerek V, Svobodova Z (2017) Accumulation of mercury in the tissues of the great cormorant (Phalacrocorax carbo) from common carp. Bull Environ Contam Toxicol 98(2):167–171
Kubetzki U, Garthe S, Fifield D, Mendel B, Furness RW (2009) Individual migratory schedules and wintering areas of northern gannets. Mar Ecol Progr Ser 391:257–265
Lambertini M, Leonzio C (1986) Pollutant levels and their effects on Mediterranean seabirds. In: MEDMARAVIS, Monbailliu X (ed) Mediterranean marine Avifauna. NATO ASI Series (Series G: Ecological Sciences), vol 12. Springer, Berlin, pp 359–378
Lehel J, Gál J, Faragó S, Berta E, Andrásofszky E, Fekete SG, Mándoki M, Budai P, Kormos E, Marosán M (2013) Evaluation of mercury and lead content in the liver of the cormorant (Phalacrocorax carbo sinensis) population of Kis-Balaton, Hungary. Acta Vet Hung 61(2):187–196
Lemarchand C, Rosoux R, Pénide ME, Berny P (2012) Tissue concentrations of pesticides, PCBs and metals among ospreys, Pandion haliaetus, collected in France. Bull Environ Contam Toxicol 88(1):89–93
Leonzio C, Fossi C, Focardi S (1986) Lead, mercury, cadmium and selenium in two species of gull feeding on inland dumps, and in marine areas. Sci Total Environ 57:121–127
Lounsbury-Billie MJ, Rand GM, Cai Y, Bass OL (2008) Metal concentrations in osprey (Pandion haliaetus) populations in the Florida Bay estuary. Ecotoxicology 17(7):616–622
Louzao M, Delord K, García D, Boué A, Weimerskirch H (2012) Protecting persistent dynamic oceanographic features: transboundary conservation efforts are needed for the critically endangered balearic shearwater. PLoS ONE 7(5):e35728
Mallory ML, Provencher JF, Robertson GJ, Braune BM, Holland ER, Klapstein S, Stevens K, O’Driscoll NJ (2018) Mercury concentrations in blood, brain and muscle tissues of coastal and pelagic birds from northeastern Canada. Ecotoxicol Environ Saf 157:424–430
Mazloomi S, Esmaeili A, Ghasempoori SM, Omidi A (2008) Mercury distribution in liver, kidney, muscle and feathers of Caspian Sea common cormorant (Phalacrocorax carbo). Res J Environ Sci 2(6):433–437
Mendes P, Eira C, Torres J, Soares AMVM, Melo P, Vingada J (2008) Toxic element concentration in the Atlantic gannet Morus bassanus (Pelecaniformes, Sulidae) in Portugal. Arch Environ Contam Toxicol 55(3):503–509
Mendes P, Eira C, Vingada J, Miquel J, Torres J (2013) The system Tetrabothrius bassani (Tetrabothriidae)/Morus bassanus (Sulidae) as a bioindicator of marine heavy metal pollution. Acta Parasitol 58(1):21–25
Miles WT, Mavor R, Riddiford NJ, Harvey PV, Riddington R, Shaw DN, Parnaby D, Reid JM (2015) Decline in an Atlantic puffin population: evaluation of magnitude and mechanisms. PLoS ONE 10(7):e0131527
Misztal-Szkudlińska M, Szefer P, Konieczka P, Namieśnik J (2011) Biomagnification of mercury in trophic relation of Great Cormorant (Phalacrocorax carbo) and fish in the Vistula Lagoon, Poland. Environ Monitor Assess 176(1–4):439–449
Misztal-Szkudlińska M, Szefer P, Konieczka P, Namieśnik J (2012) Mercury in different feather types from great cormorants (Phalacrocorax carbo L.) inhabiting the Vistula Lagoon ecosystem in Poland. Bull Environ Contam Toxicol 89(4):841–844
Misztal-Szkudlińska M, Kalisińska E, Szefer P, Konieczka P, Namieśnik J (2018) Mercury concentration and the absolute and relative sizes of the internal organs in cormorants Phalacrocorax carbo (L. 1758) from the breeding colony by the Vistula Lagoon (Poland). Ecotoxicol Environ Saf 154:118–126
Mollazadeh N, Esmaili A, Ghasempouri SM (2011) Distribution of mercury in some organs of Anzali wetland common cormorant (Phalacrocorax carbo). In: 2nd international conference on environmental engineering and applications, IPCBEE, vol 17. IACSIT Press, Singapore
Monteiro LR, Furness RW, del Nevo AJ (1995) Mercury levels in seabirds from the Azores, Mid-North Atlantic Ocean. Arch Environ Contam Toxicol 28(3):304–309
Monteiro LR, Granadeiro JP, Furness RW, Oliveira P (1999) Contemporary patterns of mercury contamination in the Portuguese Atlantic inferred from mercury concentrations in seabird tissues. Mar Environ Res 47(2):137–156
Moreno R, Jover L, Diez C, Sardà-Palomera F, Sardà F, Sanpera C (2013) Ten years after the prestige oil spill: seabird trophic ecology as indicator of long-term effects on the coastal marine ecosystem. PLoS ONE 8(10):e77360
Moura JF, Tavares DC, Lemos LS, Silveira VV, Siciliano S, Hauser-Davis RA (2018a) Variation in mercury concentration in juvenile Magellanic penguins during their migration path along the Southwest Atlantic Ocean. Environ Pollut 238:397–403
Moura JF, Tavares DC, Lemos LS, Acevedo-Trejos E, Saint’Pierre TD, Siciliano S, Merico A (2018b) Interspecific variation of essential and non-essential trace elements in sympatric seabirds. Environ Pollut 242:470–479
Nam DH, Anan Y, Ikemoto T, Okabe Y, Kim EY, Subramanian A, Saeki K, Tanabe S (2005) Specific accumulation of 20 trace elements in great cormorants (Phalacrocorax carbo) from Japan. Environ Pollut 134(3):503–514
Nardiello V, Fidalgo LE, López-Beceiro A, Bertero A, Martínez-Morcillo S, Míguez MP, Soler F, Caloni F, Pérez-López M (2019) Metal content in the liver, kidney, and feathers of Northern gannets, Morus bassanus, sampled on the Spanish coast. Environ Sci Pollut Res pp 1–9
Norheim G, Frøslle A (1978) The degree of methylation and organ distribution of mercury in some birds of prey in Norway. Acta Pharmacol Toxicol 43(3):196–204
Olaso I, Gutiérrez JL, Villamor B, Carrera P, Valdés L, Abaunza P (2005) Seasonal changes in the north-eastern Atlantic mackerel diet (Scomber scombrus) in the north of Spain (ICES Division VIIIc). Mar Biol Assoc UK 85(2):415
Otero XL, de la Peña-Lastra S, Romero D, Nobrega GN, Ferreira TO, Pérez-Alberti A (2018) Trace elements in biomaterials and soils from a yellow-legged gull (Larus michahellis) colony in the Atlantic Islands of Galicia National Park (NW Spain). Mar Pollut Bull 133:144–149
Paiva VH, Tavares PC, Ramos JA, Pereira E, Antunes S, Duarte AC (2008) The influence of diet on mercury intake by little tern chicks. Arch Environ Contam Toxicol 55(2):317–328
Parslow JLF, Jefferies DJ (1977) Gannets and toxic chemicals. Br Birds 70(366):72
Parslow JLF, Jefferies DJ, Hanson HM (1973) Gannet mortality incidents in 1972. Mar Pollut Bull 4(3):41–43
Pereira MG, Lawlor A, Bertolero A, Díez S, Shore RF, Lacorte S (2019) Temporal and spatial distribution of mercury in gulls eggs from the Iberian Peninsula. Arch Environ Contam Toxicol 76(3):394–404
Rajaei F, Sari A, Bahramifar N, Ghasempouri SM, Savabieasfahani M (2010) Mercury concentration in 3 species of gulls, Larus ridibundus, Larus minutus, Larus canus, from south coast of the Caspian Sea, Iran. Bull Environ Contam Toxicol 84:716–719
Ramos R, Ramírez F, Jover L (2013) Trophodynamics of inorganic pollutants in a wide-range feeder: the relevance of dietary inputs and biomagnification in the Yellow-legged gull (Larus michahellis). Environ Pollut 172:235–242
Ribeiro A, Eira C, Torres J, Mendes P, Miquel J, Soares A, Vingada J (2009) Toxic element concentrations in the razorbill Alca torda (Charadriiformes, Alcidae) in Portugal. Arch Environ Contam Toxicol 56(3):588–595
Rumbold DG, Niemczyk SL, Fink LE, Chandrasekhar T, Harkanson B, Laine KA (2001) Mercury in eggs and feathers of great egrets (Ardea albus) from the Florida Everglades. Arch Environ Contam Toxicol 41(4):501–507
Ruus A, Øverjordet IB, Braaten HFV, Evenset A, Christensen G, Heimstad ES, Gabrielsen GW, Borgå K (2015) Methylmercury biomagnification in an Arctic pelagic food web. Environ Toxicol Chem 34(11):2636–2643
Saeki K, Okabe Y, Kim EY, Tanabe S, Fukuda M, Tatsukawa R (2000) Mercury and cadmium in common cormorants (Phalacrocorax carbo). Environ Pollut 108(2):249–255
Sanpera C, Valladares S, Moreno R, Ruiz X, Jover L (2008) Assessing the effects of the Prestige oil spill on the European shag (Phalacrocorax aristotelis): trace elements and stable isotopes. Sci Total Environ 407(1):242–249
Savinov VM, Gabrielsen GW, Savinova TN (2003) Cadmium, zinc, copper, arsenic, selenium and mercury in seabirds from the Barents Sea: levels, inter-specific and geographical differences. Sci Total Environ 306(1–3):133–158
Scheuhammer AM (1987) The chronic toxicity of aluminium, cadmium, mercury, and lead in birds: a review. Environ Pollut 46:263–295
SEO/Birdlife (2008) Enciclopedia de las Aves de España. https://www.seo.org/listado-aves-2/. Consulted 14 May 2019
Skoric S, Visnjić-Jeftic Z, Jaric I, Djikanovic V, Mickovic B, Nikcevic M, Lenhardt M (2012) Accumulation of 20 elements in great cormorant (Phalacrocorax carbo) and its main prey, common carp (Cyprinus carpio) and Prussian carp (Carassius gibelio). Ecotoxicol Environ Saf 80:244–251
Szostek KL, Becker PH (2015) Survival and local recruitment are driven by environmental carry-over effects from the wintering area in a migratory seabird. Oecologia 178(3):643–657
Szumiło-Pilarska E, Grajewska A, Falkowska L, Hajdrych J, Meissner W, Frączek T, Bełdowska M, Bzoma S (2016) Species differences in total mercury concentration in gulls from the Gulf of Gdansk (Southern Baltic). J Trace Elements Med Biol (GMS) 33:100–109
Szumiło-Pilarska E, Falkowska L, Grajewska A, Meissner W (2017) Mercury in feathers and blood of gulls from the Southern Baltic Coast, Poland. Water Air Soil Pollut 228(4):138
Triay R, Siverio M (2004) Águila Pescadora, Pandion haliaetus. In: Madroño A, González C, Atienza YJC (eds) Libro Rojo de las Aves de España. Dirección General para la Biodiversidad-SEO/BirdLife, Madrid
Tudela S, Palomera I (1997) Trophic ecology of the European anchovy Engraulis encrasicolus in the Catalan Sea (northwest Mediterranean). Mar Ecol Progr Ser 160:121–134
Vizuete J, Hernández-Moreno D, Fidalgo LE, Bertini S, Andreini R, Soler F, Míguez-Santiyán MP, López-Beceiro A, Pérez-López M (2018) Concentrations of chlorinated pollutants in adipose tissue of yellow-legged gulls (Larus michahellis) from Spain: role of gender and age. Ecotoxicol Environ Saf 164:493–499
Wolfe MF, Achwarzbach S, Sulamian RA (1998) Effect of mercury on wildlife: a comprehensive review. Environ Toxicol Chem 17(2):146–160
Zamani-Ahmadmahmoodi R, Esmaili-Sari A, Savabieasfahani M, Ghasempouri SM, Bahramifar N (2010) Mercury pollution in three species of waders from Shadegan wetlands at the head of the Persian Gulf. Bull Environ Contam Toxicol 84(3):326–330
Zamani-Ahmadmahmoodi R, Alahverdi M, Mirzaei R (2014) Mercury Concentrations in Common Tern Sterna hirundo and Slender-billed Gull Larus genei from the Shadegan Marshes of Iran, in north-western corner of the Persian Gulf. Biol Trace Element Res 159(1–3):161–166
Zillioux EJ, Porcella DB, Benoit JM (1993) Mercury cycling and effects in freshwater wetland ecosystems. Environ Toxicol Chem 12:2245–2264
Zolfaghari G, Esmaili-Sari A, Ghasempouri SM, Baydokhti RR, Hassanzade Kiabi B (2009) A multispecies-monitoring study about bioaccumulation of mercury in Iranian birds (Khuzestan to Persian Gulf): effect of taxonomic affiliation and trophic level. Environ Res 109(7):830–836
Acknowledgements
This work was supported by Fundación Séneca—Agencia de Ciencia y Tecnología de la Región de Murcia (MASCA′2014 project, 19481/PI/14). S. Espín was financially supported by Fundación Séneca (Saavedra-Fajardo Project 20031/SF/16) and by Ministerio de Ciencia, Innovación y Universidades (Juan de la Cierva-Incorporación contract, IJCI-2017-34653). The authors thank the COST Action 16224 ERBFacility for maintaining an international collaboration network for researchers. Two anonymous referees are acknowledged for their help in improving the manuscript.
Funding
This study was funded by Fundación Séneca—Agencia de Ciencia y Tecnología de la Región de Murcia (MASCA′2014 project, 19481/PI/14). S. Espín was financially supported by Fundación Séneca (Saavedra-Fajardo Project 20031/SF/16) and by Ministerio de Ciencia, Innovación y Universidades (Juan de la Cierva-Incorporación contract, IJCI-2017-34653).
Author information
Authors and Affiliations
Contributions
SA: Methodology, Formal analysis, Writing—Original Draft NB, PS-V, IN: Methodology, Formal analysis, Writing—Review and Editing PG-R, PJ, MYT-C, IV: Methodology, Writing—Review and Editing PM-M: Conceptualization, Methodology, Resources, Writing—Review and Editing, Supervision AJG-F, SE: Conceptualization, Methodology, Formal analysis, Resources, Writing—Review and Editing, Supervision, Funding acquisition.
Corresponding authors
Ethics declarations
Conflicts of interest
The authors declare that they have no conflict of interest.
Ethics Approval
Not applicable. Carcasses from dead individuals admitted in the Wildlife Rehabilitation Centre were used in this study.
Rights and permissions
About this article

Cite this article
Albertos, S., Berenguer, N.I., Sánchez-Virosta, P. et al. Mercury Exposure in Birds Linked to Marine Ecosystems in the Western Mediterranean. Arch Environ Contam Toxicol 79, 435–453 (2020). https://doi.org/10.1007/s00244-020-00768-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00244-020-00768-1