
Abstract
A plan to create solution-mined salt caverns for natural gas storage by discharging brine into the Shubenacadie River estuary poses a potential risk to an “endangered” stock of striped bass. Toxicity of brine made from both salt-core and artificial sea-salt “Instant Ocean” was assessed by 1-h acute toxicity tests at both 19 °C and 12 °C, the typical thermal range in June, post-spawning. The short test duration was justified given the rapid dilution of the brine in the macrotidal estuary. The median lethal concentration (LC50 1 h) 95% confidence intervals of salt-core brine at 19 °C for eggs was 51–60 parts per thousand (ppt); yolk-sac larvae 34–55 ppt; first-feeding stage larvae (6–8 mm total length, TL) 37–44 ppt, and 30–46 ppt for large larvae (14–20 mm TL). Among juveniles, the median lethal concentration was significantly higher compared to larvae: 51–58 ppt for early juveniles (4-cm fork length, FL) and 63–67 ppt for juveniles 12-cm FL. The toxicity of brine made from either Instant Ocean or salt-core was similar. At 12 °C, yolk-sac larvae salinity tolerance was 30% lower than at 19 °C, whereas other life stages exhibited a similar response to 12 °C and 19 °C. The threshold observed effect concentration (TOEC) of the salt-core ranged from 24.4 ppt on large larvae to 59.7 ppt on 12-cm juveniles. In conclusion, a very low direct threat to striped bass is estimated for the discharge of brine into the Shubenacadie River estuary.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Salt caverns have been used for storing natural gas since 1944, and by 2017, there were 104 worldwide with 17 under construction (Bays 1963; Cornot-Gandolphe 2018). The caverns are created by “solution mining”, where water is injected underground to dissolve the salt, then the brine is pumped out and discharged to the ocean, natural lagoons or lakes, artificial ponds, or into disposal wells (Simpson and Connolly 1982; Crossley 1998; Quintino et al. 2008; Stantec 2014; Lankof et al. 2016; Warren 2016). In Nova Scotia, Canada, Alton Natural Gas Storage LP plans to construct two caverns (each 3.3 × 105 m3) in the first phase of its development within the Stewiacke Formation, a stratum rich in halite (NaCl) that accrued approximately 344 million years ago (Jutras et al. 2006; Jacques Whitford 2007; MacNeil et al. 2018; T. Church, Alton Gas, personal communication April 2019). Water from the Shubenacadie River estuary will be filtered and pumped 10 km to the cavern site and then saturated brine (up to 260 parts per thousand (ppt), when operating at full capacity) will be returned to a brine pond at the discharge site for slow release during the flood tide and early ebb tide midway along a 200-m long, open-ended constructed channel (T. Church, Alton Gas, personal communication, April 2019; Fig. 1). The Shubenacadie River is macrotidal with a tidal bore, a characteristic of rivers draining into the Inner Bay of Fundy (Lynch 1982; Fig. 1a). The highly turbid tidal water (range 0–28 ppt salinity) will flush the constructed channel at an estimated velocity of 0.7–0.9 m/s, diluting the brine very quickly. Real-time monitoring and discharge control will ensure salinity 5 m from the point of discharge will be no more than 7 ppt above background or exceed 28 ppt (Alton Gas 2016; Fig. 1b). In addition to these safeguards, regulatory agencies requested the present study, because the estuary is a critical habitat for a genetically discrete population of striped bass listed as endangered by COSEWIC (COSEWIC 2012; Duston et al. 2018; Leblanc et al. 2018).

a Shubenacadie-Stewiacke River estuary, Nova Scotia, Canada. The brine discharge site is 25 river kilometers from the mouth of the Shubenacadie River (rkm 25). Eggs and estuary water were collected from the Stewiacke River (rkm 0.7 from the confluence). b Details of the brine discharge site, showing the constructed channel (adapted from Alton Gas 2018)
The effects of brine discharge on estuarine ecosystems is undocumented. In the marine environment, brine was toxic to benthic isopods and polychaetes (Eurydice pulchra, Ophelia radiate) at 60 ppt salinity (LC50 96 h exposure, local seawater 36 ppt) and had sublethal effects at 41 ppt (72 h exposure; Quintino et al. 2008). High salinity increases the diffusional influx of ions and osmotic loss of water in aquatic organisms, potentially causing a lethal elevation in the osmolality of the extracellular fluid. Conversely, freshwater is lethal to stenohaline marine fish due to the reversal of these fluxes (Evans 2010). Juvenile and adult stages of euryhaline estuarine fish, such as striped bass, easily cope with large changes in salinity by rapid adjustments in both drinking rate and active transport of ions and water across the gills and gastrointestinal tract (Madsen et al. 1994; Edwards and Marshall 2013; Kültz 2015). The salinity tolerance of the eggs and larval stages of euryhaline species, by comparison, are more uncertain as they undergo ontogenetic changes in their osmoregulatory ability, hence the need for the present study (Varsamos et al. 2005). Shubenacadie striped bass spawn in spring mostly in the Stewiacke River, approximately 5 to 11 km up-estuary from the brine discharge site (Fig. 1a). Fertilization occurs in tidal freshwater, then the early developmental stages travel back-and-forth with the tide over a salinity range of 1–15 ppt in the vicinity of the brine discharge site (Duston et al. 2018; Fig. 1a). Survival and growth of Shubenacadie striped bass was independent of salinity up to 20 ppt among eggs and larvae, and 30 ppt among early juveniles (Cook et al. 2010).
To simulate natural conditions as best possible, eggs and larvae were tested at both 19 °C and 12 °C, because they are the only stages likely to be exposed to these temperatures, and estuary water was used to dissolve salt core samples from the cavern site. Artificial sea-salt was used as reference toxicant. The 1-h test duration, approved by regulators, was conservative, because the brine will be diluted in a few seconds in the constructed channel.
Materials and Methods
Striped Bass Collection and Rearing Procedures
Newly fertilized striped bass (Morone saxatilis (Walbaum, 1792)) eggs were collected in May–June 2017 and 2018 from the Stewiacke River estuary approximately rkm 0.7 from the confluence with the Shubenacadie River (Fig. 1a). The pelagic eggs were caught late in the ebb tide (salinity < 2 ppt) with a plankton net (1-mm mesh; Aquatic Research Instruments, Hope, ID), then transported in aerated insulated containers (Igloo Cooler 19-L, Katy, TX) to the lab. Eggs hatched in 2 days in upwelling incubators (Kriesel 100-L, Aquabiotech, Quebec, QC, Canada) in 17–18 °C brackish water (2–4 ppt), a mix of Dal-AC groundwater and 30-µm sand-filtered seawater (~ 30–32 ppt; source: Marine Research Station, NRC, Ketch Harbour, NS). At 3 days post hatch (dph), each cohort of yolk-sac larvae (ca. 4-mm total length, TL) was transferred to a larger tank (1.5-m diameter, 0.25-m water depth, 800-L) at 2–6 ppt and 17–18 °C. Swim-bladder inflation at 5–7 dph was facilitated by removing oil on the water surface by using paper towel, a gentle water spray and turbidity about 150 NTU by addition of porcelain clay (6–50, Dragonfire Pottery and Supplies, Dartmouth, NS, Canada). First feeding was associated with swim-bladder inflation, at which point the temperature was raised to 20 °C. Stage I Artemia nauplii (Biomarine Aquafauna, Hawthorn, CA) were fed up to 12 dph, then stage II nauplii enriched with Algamac 3050 (Biomarine Aquafauna) up to 33 dph. Weaning onto a dry diet began at 30 dph (Gemma 0.3 mm, Skretting, Saint Andrews, NB, Canada), progressing as body size increased to commercial salmonid diets (Nutra 0.5 and 1.0 mm, Skretting; Vita 1.5 and 2.0 mm, Ewos, Surrey, BC, Canada). Rearing tanks from about 15 dph onwards were 500-L volume (1 m diameter) in a small recirculation system (Aquabiotech). Salinity was maintained at 2 ppt until the fish reached approximately 5 cm TL, then freshwater. Photoperiod was L:D 24:0 until August, then simulated natural daylength (Latitude 45 °N). Light intensity at the water surface up to 10 dph was 0 lx, then approximately 30 lx (Light meter 840022, Sper Scientific, Scottsdale, AZ).
Acute Brine Toxicity Tests
Brine (ca. 100 ppt) was prepared from eleven salt core samples each 1-m long (10-cm diameter) extracted in summer 2014 from 883 to 933 m depth at the planned cavern site (Alton, NS; Core hole 08–01, 45°12′04.9″ N, 63°16′11.6″ W). Reference toxicant brine was made from Instant Ocean (Spectrum Brands, Blacksburg, VA). To produce test brine as close as possible to discharge brine, both salts were dissolved in estuary water collected from within 3 km of the discharge point (Fig. 1a). Late in the ebb tide, to avoid salinities > 5 ppt, the highly turbid estuary water was pumped into a 1,400-L tank (82124649, Global Industries, ON, Canada) on a ¾ ton truck using a semi-trash pump (6RLAG-2LST, Red Lion, Oklahoma, OK). At the lab, the water stood for 12 h to allow the sediment to settle, then was pumped (MD12, Danner Supreme, Islandia, NY) from the truck through a one-micron filter (#Bag1, Pentair Aquatic Ecosystems, Miami, FL) into a 1,000-L insulated storage tank (Insulated Container #3501, Xactics Canada, Cornwall, ON, Canada). Brine was produced by mixing 210 L of filtered estuary water with 28 kg of either crushed salt core or Instant Ocean in a 250-L Xactic tank with vigorous aeration (airstone ALR23, Sweetwater, Pentair Aquatic Eco-Systems, Miami, FL). For 24 h, during dissolution, brine was heated to 36 °C (n = 3, Top Light Excel 300 W, Rena, Charlotte, NC), then cooled to the test temperature, either 19 °C or 12 °C (chiller: Cyclone AE5DA, AquaLogic, San Diego, CA).
Static non-renewal toxicity tests, 1-h duration, were conducted on six developmental stages: eggs, yolk-sac larvae (3–4 dph), first-feeding larvae (6–7 dph), large larvae (14–20 dph), and early juvenile (ca. 54 dph, 4-cm fork length, FL) and juvenile (ca. 134 dph, 12-cm FL). Each test comprised of six test concentrations and a control at the same salinity as the rearing water (2–6 ppt), each in triplicate. The endpoint was mortality. The 1-h test duration was highly conservative compared to the planned brine dilution rate in the estuary, and provided adequate time to operate the 21 test vessels. Mortalities during the 1-h test were not removed, because they posed no immediate threat to water quality. At each life stage, an initial “range-finder” test was run at 15, 25, 35, 45, 55, and 100 ppt, as agreed with the Department of Fisheries and Oceans and approved by Nova Scotia Environment (NSE 2016). In these “ranger-finder” tests, the time of mortality at the 100-ppt test concentration was recorded. Then, to determine the median lethal concentration (LC50) more accurately, a narrower range of salinities were tested (Table 1). Salinity (based on conductivity and temperature), temperature, oxygen concentration, and pH were recorded with handheld meters (Pro2030, YSI, Yellow Springs, OH; Orion 9107BNMD, Orion Star A121, Thermo Scientific, Beverly, MA). Light intensity at test vessel level was 20 lx for eggs and larvae and 300 lx for juveniles. Fish were transferred without acclimation from their rearing tank to the test vessels. Mortality after the 1-h exposure among swimming stages was assessed in each of the test vessels. Assessing egg mortality required a dissecting microscope; dead eggs have an opaque chorion. The acceptability criterion for each test was mortality in the control of < 30% among eggs and larvae and < 10% among juveniles.
Test vessel design and volume increased with fish size. Eggs were tested in a set of four tissue culture plates (6-wells, each 15.5 mL, Falcon, Corning Life Sciences, Oneota, NY), following Kupsco et al. (2017). Using a small spoon, 20 eggs were placed in each well containing 10 mL of test solution. Yolk-sac and first-feeding larvae were tested in 30-mL plastic mesh baskets (Café Cup, Spark Innovators, Fairfield, NJ). Three replicate baskets were secured with tape in each of seven food-grade, white, plastic 2-L containers (QS21, Ropak, Springhill, NS, Canada) filled to the brim with the test solution. Using a small plastic spoon, 20 larvae were counted into 25 mL of rearing water in a 90-mL specimen cup (02-1104, Fisher Scientific, Ottawa, ON, Canada). Then, the entire contents were gently poured into each basket; the brine in the 2-L container was stirred briefly to homogenize the test solution. To account for the small dilution due to addition of rearing water, salinity was measured after each test, and this value was used for LC50 estimation. The same procedure was used to transfer 20 large larvae to their test vessels, a 2-L container (QS21) filled with 1.5 L of test solution. Ten juveniles, both size classes, were transferred using an aquarium dip net from a bucket containing rearing water into 15 L of test solution in each of twenty-one 26-L clear plastic totes (ClearView Latch, Sterilite, Townsend, MA). Oxygen saturation was > 90% in all tests; additional aeration was not needed.
The median lethal concentration (LC50 1 h) was estimated using the Trimmed Spearman-Kärber Method with the “tsk” package in R software (Hamilton et al. 1977, 1978; Stone 2015). The trim level with the lowest standard deviation was selected. Egg mortality among controls (14–21%) was corrected by the Henderson–Tilton equation (Henderson and Tilton 1955). The difference in the LC50 between two tests was considered statistically significant if their respective 95% confidence intervals did not overlap. To compare the effect of the type of brine (SC or IO) at 19 °C, a two-way ANOVA (life stage and salt source) was conducted using the Proc Mixed model with Bonferroni adjustment (SAS 9.4, Institute Inc., Cary, NC). Yolk-sac larvae were excluded from this analysis, because only a single datum was available for the IO brine. A second two-way ANOVA (life stage and temperature) was conducted to compare the effect of temperature (12 °C or 19 °C) when SC brine was used. The “No Observed Effect Concentration” (NOEC) was estimated for each test comparing the survival at the different test concentrations with the control by an ANOVA, Proc Mixed model with Bonferroni adjustment (SAS). In the cases where the normality requirement was not met, the Kruskal–Wallis test was conducted in SAS. The “Threshold Observed Effect Concentration” (TOEC) was determined for each test condition, based on the geometric mean of the NOECs (EC 2005). To analyze the difference between two TOECs (geometric means), the NOECs were log transformed and a two-sample t test was conducted in SAS (Zar 2014). The number of pairwise comparisons was reduced from 120 to 55, because four test conditions only had a single datum and one test condition had zero valid NOEC values. The slope of the survival curves for each life stage and test condition was calculated by a linear regression between highest test concentration with 100% survival and the lowest test concentration with 0% survival, including all the intermediate points (SAS). Then each pair of slopes were compared by Student’s t test.
The chemical composition of the dilution water (1-µm filtrated estuary water) and brine samples were evaluated by Standard Water Analysis and Total Metals (AGAT Laboratories, Dartmouth, NS). Due to ion interference, not all the samples were analyzed to the same detection limit (Gros 2013). Parameters were standardized to the lowest Reported Detection Limit possible that allowed a comparison between samples. Of the 31 water-quality parameters measured, only 19 are reported here; the others were considered irrelevant to the study. Of the total metals, 12 of 26 analyzed are presented; the concentration of the others was considered too low to warrant inclusion. Halite (%) of each sample was calculated from the proportion of sodium chloride relative to the total ionic composition. Salinity (ppt) of each sample was calculated from the chloride concentration (Wooster et al. 1969). The difference in the mean concentration of each parameter between brine made from either salt core or Instant Ocean was analyzed by a two-sample t test using SAS. All protocols were reviewed and approved by the Faculty’s Animal Care and Use Committee (ACUC 2016-52, 2017-44, 2018-45).
Results
Water Quality
The chemical composition of the dilution water exhibited some variation due to natural fluctuations in the tide and freshwater run-off (see Shubenacadie River estuary water at Tables 2 and 3). Nevertheless, it was consistently very hard (mean 398 mg/L), well buffered (total alkalinity 40 mg/L), oligotrophic (nitrite < 50 mg/L, ammonia 0.17 mg/L), and slightly saline (total dissolved solids (TDS) 1858 mg/L, 1.6 ppt; Table 2). After filtration to 1 micron, the water remained turbid (210 NTU); true color 26 TCU (Table 2). Total metals were highly correlated with turbidity, indicating they were bound to particulates and biologically inactive.
Salt core (SC) brine and Instant Ocean (IO) brine were similar in mean chloride concentration, conductivity, TDS and calculated salinity (Table 2). Salt core brine was significantly higher than IO brine in calculated halite (94 vs. 81%), sodium (53 vs. 40 mg/L), and uranium (1.7 vs. 0.4 µg/L) and was significantly lower in magnesium (− 98%), potassium (− 97%), boron (− 91%), hardness (− 89%), strontium (− 84%), total alkalinity (− 78%), barium (− 75%), sulphate (− 71%), molybdenum (− 51%), calcium (− 44%), nickel (− 30%), and pH (− 5%; Tables 2 and 3).
In test solutions at 19 °C, the pH of the SC brine was significantly lower than IO brine (mean ± SE, 7.52 ± 0.10 vs. 8.07 ± 0.05), but dissolved oxygen was similar (98 ± 1% vs. 97 ± 1%). Tests at 12 °C were conducted with SC brine only, the parameters were similar to those at 19 °C, pH 7.64 ± 0.05, and dissolved oxygen 100 ± 1%.
Salinity Tolerance
Survival curves for five of the six life stages had a similar shape. The exception was eggs, which exhibited a significantly lower slope in the three test conditions (Fig. 2). The slope of eggs, first-feeding larvae, large larvae, and juvenile did not vary between the test conditions (P > 0.05). The slope of the survival curves of yolk-sac larvae, by contrast, was significantly higher in SC brine at 19 °C (sudden acute mortality; Fig. 2a) than at 12 °C (Fig. 2c; P = 0.044). Finally, the slope of the survival curve of early juveniles was higher in IO brine 19 °C (Fig. 2b) than in SC brine 19 °C (Fig. 2a; P = 0.038).

Mean survival of striped bass early life stages exposed for 1-h to salt core (SC) brine at 19 °C (a), Instant Ocean brine at 19 °C (b), and SC brine at 12 °C (c). The life stages were: eggs, yolk-sac larvae (3–4 days post-hatch, dph); first-feeding larvae (6 dph); large larvae (14–15 dph); early juvenile (57 dph); and juvenile (130–143 dph)
The TOEC following 1-h exposure ranged from 25.8 to 59.7 ppt depending on stage of development, brine type, and temperature (Table 4). In SC brine at 19 °C, large larvae and eggs were the most vulnerable life stages (TOEC 24.4 and 30.8 ppt, respectively; Table 4). Early juveniles were significantly less tolerant than the 12-cm juveniles (TOEC 45.1 vs. 59.7 ppt, respectively; Table 4). In IO brine at 19 °C and SC brine 12 °C, there was no significant difference among the life stages (Table 4).
Median Lethal Concentration
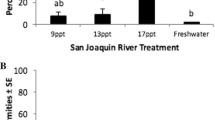
The mean of the median lethal concentration (LC50 1 h) at 19 °C estimated for the six developmental stages ranged between 34 and 65 ppt, with similar values for both SC and IO brine (Fig. 3). Salinity tolerance was highest among egg and juvenile stages and lowest among larval stages (Fig. 3). Salinity tolerance at 12 °C among eggs was similar to 19 °C, but among yolk-sac and first-feeding larvae was reduced significantly at the lower temperature (Fig. 3).

Median lethal concentration (LC50 1 h) of brine made from either salt core or Instant Ocean on early life stages of striped bass: eggs, yolk-sac larvae (3–4 days post-hatch, dph), first-feeding larvae (6–8 dph), large larvae (14–20 dph), early juvenile (43–57 dph), and juvenile (130–143 dph) exposed to 19 °C and 12 °C. Mean of the LC50 by stage and test condition. Error bar = SE of the mean. Means sharing the same lowercase letter are not significantly different between life stage and brine type at 19 °C. Means sharing the same capital letter are not significantly different between life stage and temperature when exposed to SC brine. One-way ANOVA with Bonferroni adjustment (α 0.05)
Salt core brine toxicity (LC50 1 h) on eggs at 19 °C was 54.3 ± 0.0 ppt (mean ± SE; tests 1 and 2; Table 5). Yolk-sac larvae (3–4 dph) tolerance to SC brine was similar to eggs; mean LC50 was 53.5 ± 1.5 ppt with a significant difference between tests 5 and 6 (Table 5). First-feeding larvae (6 dph) were significantly more sensitive to SC brine than both eggs and yolk-sac stages; their mean LC50 was 42.1 ± 0.9 ppt (tests 8 and 9; Table 5). Large larvae 14–15 dph were the least tolerant of all developmental stages; their mean LC50 was 34.3 ± 3.4 ppt (tests 12 and 13; Table 5). The tolerance of early juveniles to SC brine was similar to eggs and yolk-sac larvae; mean LC50 was 56.0 ± 1.6 ppt (tests 17 and 18; Table 5). The larger 12-cm juveniles (130–143 dph) were even more tolerant; the mean LC50 was 65.0 ± 0.6 ppt (tests 23 to 25; Table 5).
Instant Ocean brine toxicity at 19 °C was similar to SC brine at 19 °C. Among eggs, mean ± SE LC50 1 h was 60.1 ± 4.9 ppt compared with 54.3 ppt in SC brine (tests 3 and 4; Table 5). Among yolk-sac larvae (3 dph), the LC50 of IO brine was 51.9 ppt (test 7; Table 5), similar to the 53.5 ppt for SC brine. First-feeding larvae (8 dph) mean LC50 in IO brine was 44.4 ± 2.0 ppt compared with 42.1 ppt in SC brine. Large larvae (12–20 dph) LC50 in IO brine ranged widely from 46.1 to 57.3 ppt in four tests (14–16; Table 5), the overall mean was 50.6 ± 3.4 ppt. Early juveniles, 4-cm long, also exhibited significant variability in their LC50s. The overall mean was 60.0 ± 3.0 ppt (tests 19–22; Table 5), but among large juvenile the LC50s were consistent, mean 64.3 ± 0.3 ppt (tests 26–28; Table 5).
At 12 °C, the mean ± SE LC50 1 h of the SC brine on eggs was 56.9 ± 0.5 ppt (tests 29 and 30; Table 6), similar to 19 °C, 54.3 ppt (Table 5). By contrast, yolk-sac larvae (3–4 dph) salinity tolerance was reduced considerably at 12 °C vs. 19 °C, 38.1 ± 1.3 ppt (tests 31 and 32; Table 6) versus 53.5 ppt. First-feeding larvae (7 dph) LC50 was independent of temperature: 38.8 ± 1.0 ppt at 12 °C (tests 33 and 34; Table 6) and 42.1 ppt at 19 °C. Finally, large larvae (16–20 dph) exhibited better survival at 12 °C than at 19 °C; 41.8 ± 3.0 ppt (tests 35 and 36; Table 6) versus 34.3 ppt (Table 5).
Timing of Mortality in 100-ppt Brine
Yolk-sac larvae (3 dph, 5-mm TL) exposed to 100-ppt SC brine began to die after 3 min, and 100% mortality was reached at 5.5 min (Fig. 4). First-feeding larvae (6 dph, 6.5-mm TL) reached 50% mortality at 4 min and 100% at 5.75 min (Fig. 4). Large larvae (13 dph, 10-mm TL, exposed to IO brine) mortality started around 3.5 min and reached 100% at 12 min (Fig. 4). Early juveniles (4 cm FL) started to die after 10 min, mortality was 20% at 13 min, and reached 100% at 20.5 min (Fig. 4). Juveniles (12 cm FL) in 100-ppt brine started to die at 9 min and reached 100% mortality at 19.5 min (Fig. 4).

Mortality rate (%) of striped bass early life stages exposed to 100-ppt salt core brine at 19 °C, except large larvae expose to Instant Ocean brine: yolk-sac larvae (3 days post-hatch, dph); first-feeding larvae (6 dph); large larvae (13 dph); early juveniles (57 dph); and juveniles (129 dph)
Discussion
The engineering design considers that at 5 m from the discharge, the salinity will be no higher than 7 ppt above background and never higher than 28 ppt (Fig. 1b). After the water leaves the mixing channel, it will be quickly fully mixed with the river water. Based on these design criteria, the discharge of brine into the Shubenacadie River estuary poses a very low direct threat to striped bass, because the TOECs for five of the six early life stages were higher than the 28-ppt threshold at the normal spring water temperature (18–20 °C). Large larvae were the most sensitive, but even their TOEC was 24.4 ppt. In colder conditions (12 °C), the TOECs were closer to the 28-ppt threshold, but the quick dilution in the estuary reduces the threat observed in the 1-h test exposure. Further safeguards include the 24-days cessation of brine release following detection of the first striped bass egg, and the compulsory 20-ppt maximum at 5 m distance from point of discharge when eggs and larvae are present after the 24-days cessation. The significant changes in salinity tolerance from egg to juvenile can be attributed to the ontogenic changes in the osmoregulatory mechanisms, a common characteristic among euryhaline teleosts. Ionic composition differences between the two sources of salt were associated with the salt core brine being relatively more toxic to eggs and large larvae, but the other four stages were unaffected. Low temperature is recognized as a lethal factor for yolk-sac stage striped bass larvae and was confirmed by the greater toxicity of the brine at 12 °C compared with 19 °C.
Eggs were highly tolerant to the brine due to a largely impermeable vitelline epithelium and ionocytes (aka mitochondria-rich cells, chloride cells), specialized ion pumping cells that protect the developing embryo in all teleosts examined (Guggino 1980; Rombough 2007). Ionocytes become functional at gastrulation, the developmental stage of the eggs tested, maintaining the osmotic homeostasis of the extracellular fluid (Guggino 1980; Kaneko et al. 2008). From fertilization through blastulation, by comparison, salinity tolerance is lower since the ionocytes are absent. Among all striped bass stocks, spawning and water hardening occurs in freshwater, and survival is greatly reduced if they are exposed to brackish water as low as 2 ppt (Turner and Farley 1971). In the Shubenacadie River estuary, the risk of exposure of newly fertilized eggs the brine discharge is very low. Following spawning in the Stewiacke River in freshwater, the pelagic eggs are transported down-estuary on the ebb-tide at approximately 2 km/h (Duston, unpublished data), reaching the brine discharge site in approximately 3–4 h, by which time they are fully hardened. Moreover, any threat is further reduced, because brine discharge will be stopped for 24 days after detection of the first egg, and when larvae are present the allowable upper salinity threshold 5 m from the point discharge will be 20 ppt (Alton Gas 2015; DFO 2016; NSE 2016). From fertilization to hatch, approximately 48 h, the eggs are transported up- and down-estuary with the tide, distributed mostly between 0.5 and 15 ppt in the main channel of the estuary (Duston et al. 2018). Despite this distribution pattern, the upper salinity tolerance appears to be greater than 15 ppt, because survival to hatch was independent of salinity up to 20 ppt, and reduced significantly only at 30 ppt (Cook et al. 2010). Salinity tolerance of eggs from U.S. stocks, by comparison, appears to be lower than Shubenacadie River stock (Lal et al. 1977; Winger and Lasier 1994; Cook et al. 2010).
Yolk-sac larvae (3–4 dph, 5.5 mm TL) salinity tolerance at 19 °C was similar to eggs, because they share the same impermeable epithelium and associated ionocytes (Hirai et al. 2000). The subsequent decrease in salinity tolerance around the first-feeding stage, confirming Winger and Lasier (1994), is associated with the yolk-sac decreasing in both size and surface area, reducing its capacity for osmoregulation (Rombough 2007). The rudimentary gills and gastrointestinal tract begin to contribute to ion and water balance at first-feeding in all teleosts studied but are unable to fully compensate for the loss of yolk-sac osmoregulatory capacity (Rombough 2007). By 11 dph, striped bass gill filaments have functional ionocytes, which begin to actively excrete sodium (Na+) and chloride (Cl−; Hirai et al. 2002). Also, larval marine fish commence drinking saltwater and extract water across the intestinal wall, actively excrete Na+ and Cl− at the gills and magnesium (Mg2+) and sulfate (SO42−) via the primitive urinary system (Guggino 1980; Varsamos et al. 2005; Edwards and Marshall 2013). The increase in salinity tolerance through the larval stage reported here was likely due the increase in density of ionocytes on the skin and gill filaments quantified from 11 to 41 dph by Hirai et al. (2002). Transformation from larva to juvenile occurs during this period, around 25–36 mm TL in striped bass, associated with acquisition of definitive organs and adult morphology and structures (Otwell and Merriner 1975; Lal et al. 1977; Hardy 1978).
The metamorphosis from larva to juvenile can temporarily reduce osmoregulatory capacity in some species (Varsamos et al. 2005); but at its completion, the juveniles are better adapted to tolerate changes in salinity, as demonstrated by the 4- and 12-cm long striped bass exhibiting a LC50 1 h of 54–66 ppt (Fig. 3), and surviving in 100 ppt SC several minutes longer than larvae (Fig. 4). Similarly, survival and growth of Shubenacadie River early juveniles (6–9 cm FL) was independent of salinity between 1 to 30 ppt (Cook et al. 2010), and 114-dph juveniles from other stocks easily tolerated transfer to full seawater (34 ppt; Lal et al. 1977). When juvenile striped bass are transferred directly from freshwater to seawater (30 ppt), the structure of their ionocytes is modified quickly to maintain osmotic balance (King and Hossler 1991). The ability of juvenile striped bass to tolerate rapid increases in salinity is due to a high abundance of gill Na+/K+/ATPase and Na+/K+/2Cl− cotransporter that are “dormant” in freshwater but are activated immediately when the fish are exposed to seawater (Madsen et al. 1994; Tipsmark et al. 2004). Moreover, the rapid response is facilitated by insulin-like growth factor 1 and 2 and “epidermal growth factor” receptors present in the gill lamellae of striped bass are absent in other teleosts (Madsen et al. 2007; Tipsmark et al. 2007).
Differences in the ion composition between the salt core brine (SC) and the Instant Ocean brine (IO), specifically the very low potassium (K+) and magnesium (Mg2+) in the salt core, had no effect on the relative toxicity of the two brines at 19 °C in most of the life stages (Fig. 3). Moreover, the mixing of the brine with estuary water in the constructed channel will be very rapid, homogenizing the ions. Salt core composition is dictated by the long-term oscillation of the ocean ionic make-up over the past 600 My (Horita et al. 2002; Lowenstein et al. 2003; Holt et al. 2014). There is no evidence the very low levels of either K+ or Mg2+ in the SC brine would pose a threat to aquatic organisms. The K+ concentration in the SC brine was fourfold higher than the lower-lethal level to larval Gulf killifish (Fundulus grandis), and Mg2+ concentration was 30% greater than the lower-level that interfered with Na+/K+-ATPase activity (Fisher et al. 2013, 2015). Potassium ions are essential for ionocyte function and needed for the ion exchange by the Na+/K+-ATPase and Na+/K+/2Cl− cotransporter (Fisher et al. 2013). Magnesium and Ca2+ to a greater extent serve to reduce the ionic permeability of the epithelium. Their deficiency produced an osmotic shock following transfer of teleosts to hypertonic media due to a rapid influx of Na+ and dehydration (Lemm et al. 1993; Dolomatov et al. 2012; Fisher et al. 2015).
The interaction between temperature and salinity is important for the survival of striped bass early life stages, with a low of 12 °C identified as a lethal factor among U.S. stocks (Otwell and Merriner 1975; Morgan et al. 1981; Rutherford and Houde 1995). Poor egg survival at 12 °C in the Chesapeake Bay, estimated by Rutherford and Houde (1995), was in contrast to the similar acute salinity tolerance of eggs at 12 and 19 °C (Fig. 3). The vulnerability of yolk-sac larvae to low temperature, as evidenced by the 28% decrease in the median lethal brine concentration at 12 versus 19 °C (Fig. 3), supports data on U.S. stocks (Dey 1981; Morgan et al. 1981; Rutherford and Houde 1995). The cold tolerance of yolk-sac larvae of Shubenacadie stock, however, appears to be superior to U.S. stocks (Cook et al. 2010). Among feeding larvae, the independence in salinity tolerance between 12 and 19 °C reported here contrasts with reduced survival at 10 to 14 versus 16 °C in a 6-day trial in Instant Ocean (Cook et al. 2010). The longer exposure may have caused more disruption to osmoregulation than our 1-h tests because of delayed effects on Na+/K+/ATPase, an integral component of the salt pump (Donaldson et al. 2008).
In conclusion, the discharge of brine from the salt cavern in the mixing channel should not present any direct threat to the early life stages of striped bass, because it will be diluted to no more than 28-ppt (or 20 ppt when eggs or larvae are present) at 5 m from the point of release due to the high mixing and dilution due to the macrotidal conditions in the estuary.
References
Alton Gas - Alton Natural Gas Storage LP (2015) Exposure pathway assessment framework for aquatic and non-aquatic species in relation to the Alton Gas Natural Gas Storage cavern development at the river site. http://altonnaturalgasstorage.ca/doc/spacial-and-temporal-aquatic-species-monitoring-dec-9-2015.pdf. Accessed 15 Jan 2019
Alton Gas - Alton Natural Gas Storage LP (2016) Review of the Conestoga-Rovers & Associates third party review findings. http://altonnaturalgasstorage.ca/doc/Alton-CRA-Response-May-2016.pdf. Accessed 7 May 2017
Alton Gas - Alton Natural Gas Storage LP (2018) Young striped bass have very high salinity tolerance. Newsletter June 2018. https://s3.amazonaws.com/alton-docs/Alton+salinity+tolerance+report.pdf. Accessed 8 Aug 2018
Bays CA (1963) Use of salt solution cavities for underground storage. First Symposium on Salt. Northern Ohio Geol Soc, pp 564–578. http://www.worldsaltsymposium.org/download/use-of-salt-solution-cavities-for-underground-storage-5/. Accessed 6 Sept 2018
Cook AM, Duston J, Bradford RG (2010) Temperature and salinity effects on survival and growth of early life stage Shubenacadie River striped bass. Trans Am Fish Soc 139:749–757. https://doi.org/10.1577/T08-173.1
Cornot-Gandolphe S (2018) Underground gas storage in the world - 2018 status. Cedigaz Insights 31:1–17. https://www.cedigaz.org/. Accessed 5 Jan 2019
COSEWIC - Committee on the Status of Endangered Wildlife in Canada (2012) COSEWIC assessment and status report on the striped bass Morone saxatilis in Canada. 2012. https://wildlife-species.canada.ca/species-risk-registry/virtual_sara/files/cosewic/sr_bar_raye_striped_bass_1213a_e.pdf. Accessed 11 June 2018
Crossley NG (1998) Conversion of LPG salt caverns to natural gas storage “A TransGas experience”. J Can Petrol Technol 37:37–47. https://doi.org/10.2118/98-12-03
Dey WP (1981) Mortality and growth of young-of-the-year striped bass in the Hudson River estuary. Trans Am Fish Soc 110:151–157. https://doi.org/10.1577/1548-8659(1981)110%3c151:MAGOYS%3e2.0.CO;2
DFO - Department of Fisheries and Oceans - Canada (2016) Review of a method for identifying a window of principle striped bass (Morone saxatilis) spawning in the Shubenacadie River estuary. DFO Can Sci Advis. Sec Sci Resp 026:1–8. http://publications.gc.ca/collections/collection_2016/mpo-dfo/Fs70-7-2016-026-eng.pdf. Accessed 22 March 2017
Dolomatov SI, Zukow W, Novikov YN, Muszkieta R, Bulatowicz I, Dzierzanowski M, Kazmierczak U, Strojek K (2012) The regulation of osmotic and ionic balance in fish reproduction and in the early stages of ontogeny. Russ J Mar Biol 38:365–374. https://doi.org/10.1134/S1063074012050057
Donaldson MR, Cooke SJ, Patterson DA, Macdonald JS (2008) Cold shock and fish. J Fish Biol 73:1491–1530. https://doi.org/10.1111/j.1095-8649.2008.02061.x
Duston J, Manríquez-Hernández J, MacInnis GM, Reesor CM, Astatkie T (2018) Striped bass early life history in the macrotidal Shubenacadie River. Trans Am Fish Soc 147:919–938. https://doi.org/10.1002/tafs.10076
EC - Environment Canada (2005). Guidance document on statistical methods for environmental toxicity tests. Reference method EPS 1/RM/46 with June 2007 amendments. http://publications.gc.ca/collections/collection_2012/ec/En49-7-1-46-eng.pdf. Accessed 3 September 2019
Edwards SL, Marshall WS (2013) Principles and patterns of osmoregulation and euryhalinity in fishes. In: McCormick SD, Farrell AP, Brauner CJ (eds) Euryhaline fishes. Fish physiology, vol 32. Academic Press, London, pp 1–44. https://doi.org/10.1016/b978-0-12-396951-4.00001-3
Evans DH (2010) A brief history of the study of fish osmoregulation: the central role of the Mt. Desert Island Biological Laboratory. Front Physiol. https://doi.org/10.3389/fphys.2010.00013
Fisher C, Bodinier C, Kuhl A, Green C (2013) Effects of potassium ion supplementation on survival and ion regulation in Gulf killifish Fundulus grandis larvae reared in ion deficient saline waters. Comp Biochem Physiol. A Mol Integr Physiol 164:572–578. https://doi.org/10.1016/j.cbpa.2013.01.002
Fisher C, Bodinier C, Kuhl A, Green C (2015) Application of physiological tests to determine specific monovalent and divalent ion supplementation for culture of marine species. Bull Fish Res Agen 40:97–107
Gros N (2013) Ion chromatographic analyses of sea waters, brines and related samples. Water 5:659–676. https://doi.org/10.3390/w5020659
Guggino WB (1980) Salt balance in embryos of Fundulus heteroclitus and F. bermudae adapted to seawater. Am J Physiol 238:R42–R49. https://doi.org/10.1152/ajpregu.1980.238.1.R42
Hamilton MA, Russo RC, Thurston RV (1977) Trimmed Spearman-Karber method for estimating median lethal concentrations in toxicity bioassays. Environ Sci Technol 11:714–719. https://doi.org/10.1021/es60130a004
Hamilton MA, Russo RC, Thurston RV (1978) Correction. Trimmed Spearman-Karber method for estimating median lethal concentrations in toxicity bioassays. Environ Sci Technol 12:417. https://doi.org/10.1021/es60140a017
Hardy Jr JD (1978) Development of fishes of the mid-Atlantic bight. An atlas of egg, larval and juvenile stages, vol III. Aphredoderidae trough Rachycentridae. Biological Services Program. Fish Wildlife Series. US Department of the Interior, USA
Henderson CF, Tilton EW (1955) Tests with acaricides against the brown wheat mite. J Econ Entomol 48(2):157–161. https://doi.org/10.1093/jee/48.2.157
Hirai N, Tanaka M, Tagawa M, Secor DH (2000) Osmoregulatory responses in striped bass Morone saxatilis larvae: survival, growth, yolk absorption, and development of chloride cells in body skin. In: International congress on the biology of fish, fish migration and passage, symposium proceedings, pp 83–95
Hirai N, Tagawa M, Kaneko T, Secor DH, Tanaka M (2002) Freshwater adaptation in Japanese sea bass and striped bass: A comparison of chloride cell distribution during their early life history. Fish Sci 68(sup1):433–434
Holt NM, García-Veigas J, Lowenstein TK, Giles PS, Williams-Stroud S (2014) The major-ion composition of Carboniferous seawater. Geochim Cosmochim Acta 134:317–334. https://doi.org/10.1016/j.gca.2014.03.009
Horita J, Zimmermann H, Holland HD (2002) Chemical evolution of seawater during the Phanerozoic: Implications from the record of marine evaporites. Geochim Cosmochim Acta 66:3733–3756. https://doi.org/10.1016/S0016-7037(01)00884-5
Jacques Whitford - Jacques Whitford Environment Limited (2007) Environmental registration for the proposed Alton Natural Gas Storage Project. Final report. Project N° 1012229. https://www.novascotia.ca/nse/ea/Alton/Section1-4.pdf. Accessed 9 April 2016
Jutras P, Ryan RJ, Fitzgerald R (2006) Gradual encroachment of a rocky shoreline by an invasive sea during the Mississippian at the southeastern margin of the Maritimes Basin, Nova Scotia, Canada. Can J Earth Sci 43:1183–1204. https://doi.org/10.1139/e06-094
Kaneko T, Watanabe S, Lee KM (2008) Functional morphology of mitochondrion-rich cells in euryhaline and stenohaline teleosts. Aqua Biosci Monogr 1:1–62. https://doi.org/10.5047/absm.2008.00101.0001
King JAC, Hossler FE (1991) The gill arch of the striped bass (Morone saxatilis). IV. Alterations in the ultrastructure of chloride cell apical crypts and chloride efflux following exposure to seawater. J Morphol 209:165–176. https://doi.org/10.1002/jmor.1052090204
Kültz D (2015) Physiological mechanisms used by fish to cope with salinity stress. J Exp Biol 218:1907–1914. https://doi.org/10.1242/jeb.118695
Kupsco A, Sikder R, Schlenk D (2017) Comparative development toxicity of desalination brine and sulfate-dominated saltwater in euryhaline fish. Arch Environ Contam Toxicol 72:294–302. https://doi.org/10.1007/s00244-016-0354-9
Lal K, Lasker R, Kuljis A (1977) Acclimation and rearing of striped bass larvae in sea water. Calif Fish Game 63:210–218
Lankof L, Polański K, Ślizowski J, Tomaszewska B (2016) Possibility of energy storage in salt caverns. AGH Drill Oil Gas 33:405–415. https://doi.org/10.7494/drill.2016.33.2.405
Leblanc NM, Andrews SN, Avery TS, Puncher GN, Gahagan BI, Whiteley AR, Curry RA, Pavey SA (2018) Evidence of a genetically distinct population of striped bass within the Saint John River, New Brunswick, Canada. N Am J Fish Manag 38:1339–1349. https://doi.org/10.1002/nafm.10242
Lemm CA, Herman RL, Lemarie DP, Arzapalo A (1993) Effects of diet and environmental salinity on the growth, mortality, and tissue structure of juvenile striped bass. J Aquat Anim Health 5:294–305. https://doi.org/10.1577/1548-8667(1993)005%3c0294:EODAES%3e2.3.CO;2
Lowenstein TK, Hardie LA, Timofeeff MN, Demicco RV (2003) Secular variation in seawater chemistry and the origin of calcium chloride basinal brines. Geology 31:857–860. https://doi.org/10.1130/G19728R.1
Lynch DK (1982) Tidal bores. Sci Am 247:146–156
MacNeil LA, Pufahl PK, James NP (2018) Deposition of a saline giant in the Mississippian Windsor Group, Nova Scotia, and the nascent Late Paleozoic Ice Age. Sed Geol 363:118–135. https://doi.org/10.1016/j.sedgeo.2017.10.010
Madsen SS, McCormick SD, Young G, Endersen JS, Nishioka RS, Bern HA (1994) Physiology of seawater acclimation in the striped bass, Morone saxatilis (Walbaum). Fish Physiol Biochem 13:1–11. https://doi.org/10.1007/BF00004114
Madsen SS, Jensen LN, Tipsmark CK, Kiilerich P, Borski RJ (2007) Differential regulation of cystic fibrosis transmembrane conductance regulator and Na+, K+-ATPase in gills of striped bass, Morone saxatilis: effect of salinity and hormones. J Endocrinol 192:249–260. https://doi.org/10.1677/JOE-06-0016
Morgan RP II, Rasin VJ Jr, Copp RL (1981) Temperature and salinity effects on development of striped bass eggs and larvae. Trans Am Fish Soc 110:95–99. https://doi.org/10.1577/1548-8659(1981)110%3c95:TASEOD%3e2.0.CO;2
NSE - Nova Scotia Environmental Department (2016) Approval to operate - brine storage pond. N° 2008-061384-A03. http://altonnaturalgasstorage.ca/doc/industrial-approval-esp-canada0001.pdf. Accessed 6 June 2016
Otwell WS, Merriner JV (1975) Survival and growth of juvenile striped bass, Morone saxatilis, in a factorial experiment with temperature, salinity and age. Trans Am Fish Soc 3:560–566. https://doi.org/10.1577/1548-8659(1975)104%3c560:SAGOJS%3e2.0.CO;2
Quintino V, Rodrigues AM, Freitas R, Ré A (2008) Experimental biological effects assessment associated with on-shore brine discharge from the creation of gas storage caverns. Estuar Coast Shelf Sci 79:525–532. https://doi.org/10.1016/j.ecss.2008.05.004
Rombough P (2007) The functional ontogeny of the teleost gill: which comes first, gas or ion exchange? Comp Biochem Physiol. A Mol Integr Physiol 148:732–742. https://doi.org/10.1016/j.cbpa.2007.03.007
Rutherford ES, Houde ED (1995) The influence of temperature on cohort-specific growth, survival, and recruitment of striped bass, Morone saxatilis, larvae in Chesapeake Bay. Fish Bull 93:315–332
Simpson F, Connolly GM (1982). Underground storage potential of the bedded salt deposits of Saskatchewan: project outline and rationale. Government of Saskatchewan. 1982; Summer volume, pp 112–119. http://publications.gov.sk.ca/details.cfm?p=5709. Accessed 26 November 2018
Stantec - Stantec Consulting Ltd (2014) Canadian Environmental Assessment Agency - Project description. Strathcona salt cavern storage project. Project number 123511289. https://ceaa-acee.gc.ca/050/documents/p80082/99638E.pdf. Accessed 26 Nov 2018
Stone BR (2015) Calculates the ED50 (effective dose) from a dose-response curve using the Trimmed Spearman-Karber method. https://rdrr.io/rforge/tsk/man/tsk.html. Accessed 2 Sept 2018
Tipsmark CK, Madsen SS, Borski RJ (2004) Effect of salinity on expression of branchial ion transporters in striped bass (Morone saxatilis). J Exp Zool 301A:979–991. https://doi.org/10.1002/jez.a.119
Tipsmark CK, Luckenbach JA, Madsen SS, Borski RJ (2007) IGF-I and branchial IGF receptor expression and localization during salinity acclimation in striped bass. Am J Physiol Regul Integr Comp Physiol 292:R535–R543. https://doi.org/10.1152/ajpregu.00915.2005
Turner JL, Farley TC (1971) Effects of temperature, salinity, and dissolved oxygen on the survival of striped bass eggs and larvae. Calif Fish Game 57:268–273
Varsamos S, Nebel C, Charmantier G (2005) Ontogeny of osmoregulation in postembryonic fish: a review. Comp Biochem Physiol A Mol Integr Physiol 141:401–429. https://doi.org/10.1016/j.cbpb.2005.01.013
Warren JK (2016) Solution mining and salt cavern usage. In: Warren JK (ed) Evaporites. A geological compendium, 2nd edn. Springer, Cham, pp 1303–1374
Winger PV, Lasier PJ (1994) Effects of salinity on striped bass eggs and larvae from the Savannah River, Georgia. Trans Am Fish Soc 123:904–912. https://doi.org/10.1577/1548-8659(1994)123%3c0904:EOSOSB%3e2.3.CO;2
Wooster WS, Lee AJ, Dietrich G (1969) Redefinition of salinity. Z Geophys 35:611–613. https://doi.org/10.23689/fidgeo-2856
Zar JH (2014) Biostatistical analysis, 5th edn. Pearson New International Edition, Harlow
Acknowledgements
This study was funded by a Research Grant from Alton Natural Gas Storage LP and a summer student was partially funded by Employment and Social Development Canada (Canada Summer Jobs Program, Grant 015219264) and ECO Canada (Co-op Program). S. Qiu and Q. Liu provided technical assistance during the tests. Thanks to J. Strang (GIS Centre, University Libraries, Dalhousie University) for making the map.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Manríquez-Hernández, J., Breau, H.M. & Duston, J. Acute Toxicity of Salt Cavern Brine on Early Life Stages of Striped Bass (Morone saxatilis). Arch Environ Contam Toxicol 78, 124–136 (2020). https://doi.org/10.1007/s00244-019-00684-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00244-019-00684-z