
Abstract
Migration is one of several marine vertebrate traits increasingly affected by human encroachment. The Adriatic Sea is an important foraging and wintering site for loggerhead turtles (Caretta caretta), and one of the Mediterranean regions where they are most heavily impacted, particularly by fisheries bycatch. Conservation measures concern foraging and wintering areas and nesting grounds, and must, therefore, be informed by the natal origin of individuals in these mixed aggregates. Genetic diversity was investigated among 488 loggerheads found stranded or incidentally captured in fishing gear across the Adriatic and the origin of individuals was assessed through mixed-stock analysis based on comparison of 755 bp of mitochondrial DNA control region sequence. In addition, we highlight the importance of the Gulf of Manfredonia (Apulia, Italy), and of the northeastern Adriatic—a previously genetically undescribed foraging ground. No significant genetic divergence was recorded among sampling areas, between turtles sampled in cold and warm months or between adults and juveniles. The distribution of turtles across the Adriatic Sea appeared not to depend on individual origin. Rookeries in western Greece and Crete provided the most important contributions to the Adriatic mixed stock. In particular, the Greek populations were the most abundant locally in the Gulf of Manfredonia, so they are likely to experience an even greater impact than previously thought because of the severe fishery bycatch levels in this area. This study also provides an example of how substantial increases in sample sizes permit a relatively comprehensive testing of genetic structure across groupings in foraging aggregations.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Patterns of dispersal and migration in marine vertebrates are known to embrace long-distance movements to breeding and nesting areas, foraging, and wintering grounds. Unfortunately, connectivity between habitats that play important roles in the life cycle of an organism can be disturbed or disrupted by a plethora of anthropogenic threats (Halpern et al. 2007). Bycatch, the incidental capture of non-target species in fisheries, is a prevalent cause of mortality particularly in the Mediterranean Sea, where marine species like sea turtles migrate on a regular basis from foraging to nesting grounds of great importance for population survival (Casale and Margaritoulis 2010; Fortuna et al. 2010; Coll et al. 2012; Luschi and Casale 2014).
The Adriatic Sea is one of the main foraging areas in the Mediterranean basin for marine wildlife, and in particular the loggerhead sea turtle Caretta caretta. The shallow (< 100 m) waters of the northern Adriatic are highly productive (Agostini and Bakun 2002) and a main neritic foraging and wintering ground for C. caretta (Margaritoulis et al. 2003; Casale et al. 2004; Lazar et al. 2004; Casale et al. 2010; Lazar et al. 2011; Luschi and Casale 2014). The southern Adriatic is mostly oceanic, has a wide depression more than 1200 m deep (Cushman-Roisin et al. 2001), and is regarded as an important area for early life stage development, although all size classes are present throughout (Casale et al. 2010; Casale and Mariani 2014). An additional, important neritic foraging area has recently been described in the Gulf of Manfredonia (Italy), off the Apulian coast, and directly west of the South Adriatic trench. This region is frequented by turtles of different size classes showing a certain degree of site fidelity (Casale et al. 2012; Casale and Simone 2017).
Describing the connectivity between natal areas and foraging grounds represents a major challenge in sea turtle research and conservation. A variety of different tools have been adopted to investigate the long-range migration of chelonians, including capture-mark-recapture, satellite tracking, and genetic analysis. Molecular tools, in particular, can provide a thorough understanding of the natal origin of foraging stocks and, among others, give insights into how anthropogenic threats affect wild populations occurring in specific marine areas and/or nesting grounds (Hamann et al. 2010; Rees et al. 2017). Furthermore, genetic structure was among the main biological parameters involved in the recent designation of regional management units (RMUs) as groups of nesting populations on possible, independent evolutionary trajectories (Wallace et al. 2010).
Flipper tagging and satellite tracking studies of loggerheads breeding in Greece have shown the importance of the Adriatic foraging grounds for the Greek nesting populations (Lazar et al. 2004; Zbinden et al. 2008), but comparable studies from other Mediterranean nesting sites are lacking. Similarly, population genetic studies conducted on C. caretta sampled in the Adriatic Sea have demonstrated a prevalence of individuals from Greece (Garofalo et al. 2013; Clusa et al. 2014). Molecular tracking of sea turtles is mainly based on interindividual variation at mitochondrial DNA (mtDNA) sequences, which occur at different frequencies in different rookeries—a result of population structuring due to natal homing (Bowen et al. 2005). Given this rookery baseline description, mixed-stock analysis (MSA) can then be used to estimate the relative contribution of different natal sites to foraging areas (e.g., Bass et al. 2004; Bowen et al. 2007; Bjorndal and Bolten 2008). However, loggerhead mtDNA population structure is rather weak in the Mediterranean, probably because of relatively recent colonization events from the Atlantic Ocean (Clusa et al. 2013). For this reason, MSA may have low power when using a relatively small sample size, which is less likely to include rare and more informative mtDNA haplotypes (Bolker et al. 2003). The previous attempts to characterize the genetic population structure of loggerhead turtles sampled in the Adriatic Sea were based on a relatively small sample size and did not include individuals from the northeastern Adriatic waters and the Gulf of Manfredonia as major loggerhead foraging sites (Casale et al. 2012).
A thorough assessment of the natal origin of loggerheads foraging in the Adriatic basin may have profound implications for our understanding of migratory patterns of turtles in the Mediterranean Sea. Different migration routes and displacement observed in tagged and satellite-tracked adult turtles may, in fact, depend on their rookeries of origin or on other factors such as ocean currents or different foraging strategies (Zbinden et al. 2011). Although juvenile movements and dispersal are mostly driven by marine currents, natal homing has been observed in juvenile sea turtles that show site fidelity to pelagic feeding areas near their rookery of origin or areas at similar latitudes (Avens et al. 2003; Bowen et al. 2004; Monzón-Argüello et al. 2009). If dispersal to foraging grounds depends on where individual turtles come from (i.e., location and/or size of rookery of origin), then genetic structuring is expected, for turtles from different rookeries would feed in different coastal areas. If, on the other hand, the migration of turtles from different rookeries to feeding sites is based on factors other than individual origin, then no genetic structure should be recorded across the Adriatic Sea. Moreover, there might be seasonal and/or body size differences when turtles of different origin congregate at specific foraging grounds.
In this study, we assessed the natal origin of the Adriatic foraging stock through MSA, using a comprehensive data set composed of loggerhead samples collected across the Adriatic Sea as a result of bycatch and stranding events off and along the coasts of Croatia, Slovenia, northern Italy, and the southern Italian region of the Gulf of Manfredonia. We conducted a population genetic analysis to characterize previously undescribed mtDNA haplotypes and investigate possible differences among turtles from the Adriatic foraging population. Using a relatively large sample size, we attempted an optimization of Bayesian MSA by (1) adopting a population abundance prior (Pella and Masuda 2001) to account for the relatively higher contribution of rookeries with a large number of nests, rather than relying on haplotype frequencies only; and (2) relaxing the assumption of equal possibility of reaching the foraging area using priors based on geographical distance between known Mediterranean rookeries and the Adriatic Sea.
Intensive human activity in the Adriatic Sea, including fisheries, shipping, and pollution, is considered a potential threat to sea turtle populations (Franzellitti et al. 2004; Casale 2011; Lazar and Gračan 2011). Comprehensive knowledge of the composition of the Adriatic foraging stock will help to inform decision-making processes aimed at identifying marine protected areas and mitigating the anthropogenic impact on sea turtle survival. An assessment of migratory routes from the main contributing rookeries can provide background data for revising regional policies on the use of fishing gear particularly harmful to loggerheads, such as set nests and bottom trawlers—responsible for most bycatch and likely one of the main factors affecting survival rates of C. caretta in the Adriatic Sea (Casale et al. 2004, 2012, 2015).
Materials and methods
Sample collection
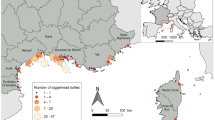
A total of 488 C. caretta samples were collected between 1995 and 2015 in the Adriatic Sea off or along the coasts of Slovenia, Croatia, and northern and southern Italy (Fig. 1). Blood and skin biopsies were taken from live bycaught turtles, while muscle and skin were obtained from necropsies of stranded or floating individuals. Sea turtles sampled alive were tagged before release to avoid pseudoreplication. Our sample set included both juveniles and adults with the curved carapace length ranging from 8.5 to 86.0 cm (see Electronic Supplementary Material Fig. S1 for the body size distribution of turtles sampled in the Adriatic Sea). Blood samples were stored at − 20 °C, while tissue samples were preserved in 95% ethanol at 4 °C or ambient temperature.

Map and geographical coordinates of the study area. Samples were collected from live bycaught turtles, as well as stranded and floating individuals, either dead or alive, in the Adriatic Sea (grey areas). Black circles show the locations of Mediterranean rookeries considered in this study as potential natal sites. Black triangles mark the location of the occasional Italian nesting sites of Tuscany (T), Campania (C), and Sicily (S) described in our study. Dashed circles include the rookeries pooled for mixed-stock analysis. Arrows show the main sea surface currents
Genetic analysis
High-molecular-weight DNA was isolated using either a standard phenol/chloroform extraction protocol (Sambrook and Russel 2001), a NucleoSpin Tissue kit (Macherey–Nagel), or a QIAamp DNA Mini Kit (Qiagen). A fragment of the mtDNA was amplified by polymerase chain reaction (PCR) using the LTEi9 light-strand primer and H950 heavy-strand primer designed by Abreu-Grobois et al. (2006). The H950 primer anneals to the control region. The LTEi9 primer anneals between the threonine tRNA and proline tRNA genes, and in combination with H950 allows PCR amplification of an 887 bp mtDNA fragment. PCR was conducted in a total volume of 15 μl with 100 ηg of total DNA, 1 × PCR buffer, 1.5 mM MgCl2, 200 μM of each dNTP, 0.5 μM of each primer, and 0.5 units of Taq DNA polymerase. Thermal profiles consisted of an initial denaturation step of 5 min at 94 °C, followed by 35 cycles of 45 s at 94 °C, 45 s at 55 °C, and 1 min and 30 s at 72 °C, with a final extension step of 10 min at 72 °C. Amplicons were cycle-sequenced using BigDye Terminator v3.1 chemistry (Applied Biosystems) according to the manufacturer’s protocol, and cycle sequencing reactions were resolved on Applied Biosystems 3100 Avant and 3130xl genetic analyzers.
Data analysis
Raw sequence chromatographs from both strands were edited and aligned to the loggerhead turtle mtDNA sequence using GENEIOUS 8.0.4 (Biomatters Ltd). The consensus sequence consisted of 755 bp of the mtDNA control region after trimming of tRNA gene sequences. We also trimmed sequences down to 380 bp to match the short mtDNA control region fragment described by Norman et al. (1994) and compared sequence diversity parameters to values reported for C. caretta in the previous studies. Mitochondrial haplotypes were classified by comparison to the mtDNA sequences available on the Archie Carr Center for Sea Turtle Research database (ACCSTR; http://accstr.ufl.edu/files/cclongmtdna.pdf). Relationships among haplotypes were inferred using an unrooted haplotype network based on the TCS inference approach of statistical parsimony implemented in POPART (Leigh and Bryant 2015). The method links haplotypes with the smaller number of differences as defined by a 95% confidence criterion (Templeton et al. 1992; Clement et al. 2000).
Mitochondrial control region sequence diversity was inferred by the number of haplotypes (k), number of polymorphic sites (p), haplotype diversity (h), and nucleotide diversity (π) using ARLEQUIN 3.5.2.2 (Excoffier and Lischer 2010). The extent of mtDNA control region differentiation was investigated by principal coordinate analysis (PCoA) of a standardized pairwise haplotype distance matrix using GenAlEx 6.503 (Peakall and Smouse 2012). We then assessed the average number of pairwise differences in haplotype frequencies among turtles using the Φ-statistics of Excoffier et al. (1992) implemented in ARLEQUIN 3.5.2.2. The statistical significance of the ΦST values was obtained after 10,000 haplotype permutations. Following a preliminary pattern of haplotype clustering recovered by PCoA, Φ-statistics was performed by pooling samples according to sampling region, sampling season, and body size. The Adriatic Sea can be divided into a northern, central, and southern basin—each with distinct topographic, bathymetric, and dynamic features (Artegiani et al. 1997; Cushman-Roisin et al. 2001). Similarly, the eastern coast is generally high and rocky, whereas the western coast is low and mostly sandy. Such differences may be mirrored by latitudinal and longitudinal patterns of genetic divergence as recorded in several studies on marine vertebrates of the Adriatic (e.g., Garoia et al. 2004; Gaspari et al. 2015; Ruggeri et al. 2016). Geographical subdivision, therefore, considered a southern Adriatic region (southern Italy) and a northern group (Slovenia, Croatia, northern Italy), as well as a western Adriatic (Italy) and eastern Adriatic (Slovenia and Croatia) area. The seasonal partitioning divided samples collected during relatively colder months (November–May) from those obtained during warmer periods (June–October), the latter also being the nesting season for most Mediterranean populations. The differences between warmer and colder months in terms of loggerhead sea turtle distribution as well as the frequency of by-catch events were reported for the Adriatic Sea by Lazar et al. (2002). Moreover, sea temperature is known to affect the migratory strategies of C. caretta from nesting beaches to foraging grounds, as well as their seasonal habitat choice (e.g., Hawkes et al. 2007; Schofield et al. 2009). Cold months were defined as those showing average water temperatures lower than the average annual temperature recorded in the Adriatic Sea across 34 locations in Italy, Slovenia, and Croatia from 2013 to 2015 (available at http://seatemperature.info/adriatic-sea-water-temperature.html). Warm months were those with average temperatures equal to or higher than the annual average. Finally, sequences were analyzed by dividing our sample set into two sea turtle body size classes defined according to the individual ability of swimming independently of water currents. Body size can affect the swimming ability of turtles and, therefore, their distribution under different sea-current speeds (Revelles et al. 2007). The mean speed of Adriatic currents, 0.30 m s−1 (Poulain 2001), was used to calculate the critical swimming speed (Ucrit) which a turtle of a certain body size can sustain without enduring muscular fatigue as follows: Ucrit = 1.763 SCL − 0.262 (Revelles et al. 2007; Monzón-Argüello et al. 2009), where SCL is the straight carapace length (Bolten 1999). Turtles with an SCL of 32 cm or larger were defined as individuals capable of swimming independently of sea currents, while turtles with an SCL of less than 32 cm were defined as juveniles (Casale et al. 2005) whose movements were more likely to have been affected by currents. Conversion from curved to straight carapace length was conducted following the method described by Casale et al. (2017).
An MSA was performed using the Bayesian approach implemented in the MIXSTOCK package in R (Bolker and Okuyama 2014). The package compares mtDNA sequences from one or more mixed populations with baseline haplotypes from source populations (rookeries). We used the foraging ground centric approach to estimate the posterior distribution with 95% confidence intervals of unknown proportions of loggerhead turtle rookeries in the Adriatic foraging stock (i.e., the proportion of Adriatic sea turtle stock originating from distinct rookeries) (Pella and Masuda 2001; Bolker et al. 2003).
We assessed the composition of the Adriatic stock using a baseline of 10 rookeries from the Atlantic Ocean and 13 rookeries from the Mediterranean Sea (Fig. 1; Electronic Supplementary Material Table S1). Baseline sequences from the island of Zakynthos and the bays of Kyparissia and Lakonikos in the Peloponnese Peninsula were pooled into a single western Greece group according to Carreras et al. (2014) and non-significant pairwise ΦST values (ranging from − 0.01 to − 0.04; P > 0.05). Only short mtDNA control region sequences of less than 500 bp were available from the small rookeries of Lampedusa (Italy) and the Kuriat islands (Tunisia) (Laurent et al. 1998; Chaieb et al. 2010), so these rookeries were not included in the baseline. Haplotypes from single Mediterranean nests were also not part of our MSA for they could not be ascribed to a geographically defined rookery with a specific haplotype frequency. A few Mediterranean sites that receive sporadic nesting have the haplotype CC-A10.4 (Garofalo et al. 2016a; Maffucci et al. 2016), previously found with a 0.1% frequency in the Central East Florida rookery (Shamblin et al. 2012) and likely to occur in other Mediterranean rookeries. Samples carrying the CC-A10.4 haplotype were also excluded from the MSA in order not to overestimate the Atlantic contribution in a baseline where this haplotype is found in one Atlantic rookery only.
An MSA was also performed by pooling rookeries into Mediterranean, Northwest Atlantic, and Northeast Atlantic RMUs (Wallace et al. 2010; Naro-Maciel et al. 2014). Subsequent analyses were conducted by (1) removing the contribution of the Atlantic nests from the baseline and (2) pooling 8 out of the 13 Mediterranean rookeries into four clusters, which included the two sites in Libya, Dalaman, and Dalayan in Turkey, middle, and eastern Turkey and the sites in Lebanon and Israel, respectively (Fig. 1; Electronic Supplementary Material Table S2). A similar subdivision of rookeries was proposed by Shamblin et al. (2014), whereby rookeries in Lebanon, Israel, Cyprus, eastern, and Middle Turkey were pooled into a single reference group. A ΦST test of pairwise differences in haplotype frequencies performed using ARLEQUIN 3.5.2.2 recorded no significant differences between rookeries pooled for MSA (Electronic Supplementary Material Table S3).
Priors were based on either of the following: (1) equal contribution from each rookery; (2) contribution weighted by population abundance; (3) contribution weighted by shortest swimming distance between rookeries and foraging ground; or (4) contribution weighted by both abundance and distance. The average number of nests per year was used as an estimate of population abundance (Electronic Supplementary Material Table S1). In the first MSA, where all 23 individual rookeries were considered, we used an equal contribution prior. On the other hand, all four different priors were used after pooling the eight rookeries from Libya, Lebanon, Israel, and Turkey into four distinct clusters. We also performed an MSA using the 380 bp mtDNA control region sequences to compare data obtained using mtDNA fragments of different length. As for the MSA based on the longer, 775 bp sequence fragment, the contribution of the Atlantic rookeries was close to zero (Electronic Supplementary Material Fig. S3). Subsequent analyses were, therefore, conducted following the same procedure used for the longer mtDNA control region sequence. We used a number of Markov chains equal to the number of source populations with 200,000 iterations and a 50% burn-in. Convergence of Markov chains was assessed according to the Gelman–Rubin criterion whereby a value lower than 1.2 for all parameters is taken as a general rule of thumb to test for between-chain variance being relatively smaller than within-chain variance, and for chains to converge (Gelman and Rubin 1992).
Results
Mitochondrial DNA control region sequence diversity
A total of 14 haplotypes were characterized in the C. caretta Adriatic foraging stock (Table 1). Four haplotypes (CC-A2.1, CC-A3.1, CC-A10.4, and CC-A20.1) were found in both Atlantic and Mediterranean rookeries and accounted for 91.6% of the Adriatic turtles. Among these, CC-A2.1 was the most abundant but the least informative as it is almost ubiquitous in Mediterranean and Atlantic rookeries. Haplotype CC-A10.4 is found in occasional Tyrrhenian nesting sites in the Mediterranean Sea (Garofalo et al. 2016a; Maffucci et al. 2016; Fig. 1). Two out of three individuals carrying haplotype CC-A10.4 were sampled in the southern Adriatic, off the Apulian coast. Eight exclusively Mediterranean haplotypes (CC-A2.8, CC-A2.9, CC-A6.1, CC-A26.1, CC-A29.1, CC-A31.1, CC-A32.1, and CC-A53.1) accounted for 8.2% of the total Adriatic sequences. Eleven (2.3%) loggerheads carried haplotype CC-A2.8, which is specific to the Cretan rookery, while 10 (2.0%) individuals had haplotypes CC-A6.1 and CC-A32.1 endemic to western Greece. Two individuals sampled in Slovenia carried haplotype CC-A29.1, found only in the Israeli rookery. Haplotype CC-A26.1, exclusive to Sirte (Libya), was recorded in three individuals from the northern Adriatic and one individual from the Apulian waters. We found only one turtle with Turkish haplotype CC-A53.1, recovered in Slovenian waters. Similarly, only one haplotype CC-A31.1, assigned to Calabrian and Greek rookeries and recently found in occasional nesting sites in Sicily (Garofalo et al. 2016b), was found in the northwestern Adriatic. Only 0.2% of the foraging stock (one juvenile turtle sampled along the Apulian coast) carried the Atlantic haplotype CC-A1.1.
One new haplotype was recorded in one individual sampled off the coast of Slovenia. The new haplotype differed from CC-A2.1 for one A-to-G transition at nucleotide site 15,648 of the published C. caretta mitochondrial genome sequence (Drosopoulou et al. 2012) (Fig. 2). The new sequence was named CC-A73.1 after submission to ACCSTR and deposited in GenBank under accession number MF182628. Haplotype CC-A73.1 was not considered in the MSA as its rookeries of origin have yet to be identified.

Unrooted haplotype network of C. caretta estimated by statistical parsimony analysis. The areas of the circles are proportional, on a logarithmic scale, to the number of samples carrying a specific haplotype (see Table 1). White and black circles refer to haplotypes recorded in Mediterranean and Atlantic rookeries, respectively. Grey circles are haplotypes found in both Mediterranean and Atlantic rookeries. The white, thick circle is the haplotype recorded in this study and not yet assigned to a rookery. Small solid circles are intermediate haplotype states that were not observed in our data set
The number of haplotypes, number of polymorphic sites, and haplotype and nucleotide diversities were significantly higher in turtles sampled in the northern versus the southern Adriatic. Similarly, genetic diversity indices were higher in the eastern than in the western Adriatic (Table 2; Electronic Supplementary Material Table S4). Eight polymorphic sites were recorded in turtles from the Apulian waters after removal of a single individual carrying haplotype CC-A1.1, which differed by 41 mutations from CC-A2.1 (see Electronic Supplementary Material Table S5 for genetic diversity measures obtained with the full data set). Haplotype and nucleotide diversities calculated for the short (380 bp) control region fragment were 0.22 ± 0.00 and 0.0006, respectively, for the northern Adriatic and 0.20 ± 0.00 and 0.0013, respectively, for the southern Adriatic. Molecular diversity indices were also significantly higher in turtles sampled during warmer months than in the colder season. A lower number of haplotypes and polymorphic sites were observed in adults versus juveniles, which accounted for 15% of our sample set. These values were recorded after removing the one individual carrying haplotype CC-A1.1 from the analysis.
The first and second coordinates of the PCoA explained 67.5 and 11.8% of the total variance, respectively. A pattern of haplotype differentiation similar to the one described by the statistical parsimony network was recovered, whereby CC-A26.1 and CC-A32.1 were the most divergent from the other haplotypes. However, the PCoA failed to recover a clear clustering between haplotypes exclusive of either the eastern (CC-A20.1, CC-A29.1, CC-A53.1, and CC-A73.1) or western (CC-A10.4, CC-A31.1, and CC-A32.1) Adriatic, and between haplotypes recorded in adults (CC-A31.1) or juveniles only (all others but CC-A2.1 and CC-A3.1). There were no haplotypes found only in warm or cold months or only in the northern or southern Adriatic (Electronic Supplementary Material Fig. S4). There was no significant differentiation in the average number of pairwise differences in haplotype frequencies as computed using Φ-statistics between the North and South Adriatic groups, West and East Adriatic groups, winter and summer groups, or body size clusters (Table 3). MSA was, therefore, conducted by pooling all samples in a single Adriatic stock.
Adriatic loggerhead turtle stock composition
The mixed-stock analysis performed using a baseline of 23 rookeries and equal contribution prior revealed a contribution of the Atlantic rookeries close to zero. However, the analysis had a shrink factor higher than 1.2 and reported relatively large confidence intervals for Mediterranean rookeries (Electronic Supplementary Material Table S6, Fig. S2). In contrast, the MSA of the relative contribution of three distinct RMUs to the Adriatic stock exhibited a shrink factor < 1.2 and very narrow confidence intervals. The analysis showed a remarkable discrepancy between contribution estimates of Atlantic rookeries, all very close to zero, and contribution proportions of Mediterranean rookeries (Fig. 3).

Contribution proportions and 95% confidence intervals of the Mediterranean, Northwest Atlantic, and Northeast Atlantic regional management units to the Adriatic loggerhead turtle foraging stock estimated using an equivalent-contribution prior
The MSA performed without Atlantic rookeries satisfied the Gelman–Rubin criteria, with shrink factors < 1.2, and showed good chain convergence, albeit with wide confidence intervals recorded. According to the MSA with priors weighted by population abundance and distance from the rookeries, the western Greece rookery had the highest contribution to the Adriatic mixed stock (P = 0.38, 95% CI 0.14–0.67), followed by Crete (P = 0.18, 95% CI 1.65E-06 to 0.51) and western Turkey (P = 0.18, 95% CI 3.81E-20 to 0.47). Much lower contributions were recorded for rookeries from the remaining Turkish sites, Libya, Lebanon, and Israel (Fig. 4; Electronic Supplementary Material Table S7). A similar pattern was observed when using an equal prior, as well as a prior based on contribution weighted by either population abundance or distance only (Electronic Supplementary Material Figs. S5–S7). Western Greece was the most represented, while there was a slight increase in the contribution to the Adriatic stock from the rookeries of Israel and Lebanon when using an equal prior (Electronic Supplementary Material Table S8).

Contribution proportions and 95% confidence intervals of nine rookeries and groups of rookeries from the Mediterranean Sea to the Adriatic loggerhead turtle foraging stock estimated using priors based on contribution weighted by both population abundance and distance between rookeries and foraging grounds. Acronyms of rookeries as in Fig. 1
Analysis of the shorter, 380 bp mtDNA control region fragment recovered a similar pattern of rookery contribution to the Adriatic stock except for Crete, which had a contribution approximately 10% lower than the one recorded using the longer control region sequence (Electronic Supplementary Material Figs. S8–S11). This is because the short sequence does not include a polymorphism of haplotype CC-A2.8, which is distinctive of Cretan rookeries.
Discussion
The Adriatic Sea is one of the main Mediterranean European fishing grounds for both pelagic and demersal fisheries, in addition to being a very important feeding ground for C. caretta. This overlap highlights the urgent need for management actions to balance the exploitation of marine resources with the protection of vulnerable loggerhead turtles and their critical habitats across the Adriatic Sea. Describing the origin of loggerhead turtles frequenting the Adriatic and their genetic characterization can be particularly useful in the design of effective fishery management plans.
This study investigated the population genetic diversity of loggerhead turtles across the Adriatic Sea, using a comprehensive sample set from Slovenia, Croatia, and Italy. Stranded and bycaught individuals in Italian waters were obtained from the northern regions and from the southern area of the Gulf of Manfredonia—a particularly important foraging ground of conservation concern. Haplotype and nucleotide diversities were within the range of values recorded in the previous studies, despite such studies having been based on very small numbers of loggerheads sampled in the Adriatic Sea. For instance, Clusa et al. (2014) reported lower haplotype and nucleotide diversity values for juvenile C. caretta sampled in both the northern and southern Adriatic basins. Garofalo et al. (2013) recorded higher diversity values in the North Adriatic foraging ground, although the comparison was based on the short (380 bp) fragment of the mtDNA control region. The genetic diversity recovered in our study was, nevertheless, lower than the diversity values described for loggerheads sampled in other western and eastern Mediterranean foraging sites, probably because the Adriatic is geographically a semi-enclosed basin (Garofalo et al. 2013; Clusa et al. 2014).
Differences in genetic diversity measures were observed between the North and South Adriatic, West and East Adriatic, as well as between turtles of different body sizes or sampled in distinct seasons. While such differences may have been due to a discrepancy in sample size for the majority of comparisons, the eastern Adriatic had higher measures than the western Adriatic, despite the lower number of turtles sampled. Genetic divergence based on pairwise differences in haplotype frequencies was nil or negligible among sampling regions, between turtles sampled in winter and summer, and between adults and juveniles. The absence of a clear genetic structure across the Adriatic basin strongly suggests the absence of a genetic distinctiveness of turtles foraging across an area extending from the northernmost part of the Adriatic Sea, genetically described here for the first time, to the southern Gulf of Manfredonia. The observed lack of genetic differentiation supports the hypothesis that loggerhead turtles do not disperse across the Adriatic foraging ground based specifically on their natal origin.
MSA based on RMUs recorded a negligible contribution from Atlantic rookeries to the Adriatic stock, corroborating the previous studies on loggerhead turtle distribution in the Mediterranean basin (Garofalo et al. 2013; Clusa et al. 2014). On the other hand, nesting sites in western Greece and Crete were the most important rookeries contributing to the Adriatic foraging population. Geographical proximity to the Adriatic basin, sea currents, and size of the Greek populations increase the likelihood of presence of adults, juveniles, and hatchlings from Greek rookeries in the Adriatic Sea (Casale et al. 2007; Casale and Mariani 2014). Moreover, eastern Mediterranean currents flow counterclockwise from North Africa to Turkey and the Balkan Peninsula (Fig. 1). Waters from the Aegean and Ionian Sea reach the northern Adriatic along the Croatian coastline and then flow back southwards to the Ionian Sea along the Italian peninsula (Millot and Taupier-Letage 2005; Cushman-Roisin et al. 2001). A likely migratory route to the Adriatic is, therefore, provided by sea currents flowing northwards from the Greek nesting grounds (Hays et al. 2010). Capture–mark–recapture and telemetry data from adult loggerhead turtles breeding in Mediterranean rookeries, and particularly in Greece, support genetic evidence of the extensive use of the Adriatic as a foraging ground for individuals from Greek nesting areas (Lazar et al. 2004; Luschi and Casale 2014). The fact that C. caretta from both the northern and the southern Adriatic foraging areas mainly originate from Greece is of particular conservation concern for the Greek population, which is likely to be affected by unmanaged Adriatic anthropogenic threats in greater proportion than previously estimated (Casale et al. 2004, 2010; Fortuna et al. 2010). In addition, increased numbers of nesting individuals or hatchlings from Greece would come under threat while feeding in the Adriatic, nullifying the conservation gains from the protection of nesting beaches.
Western Turkey had the third highest posterior probability of contributing to the Adriatic stock, along with the Libyan rookeries. The Libyan coastline is largely unsurveyed for the presence of nesting sites (Hamza 2010), meaning that the actual contribution to foraging areas might, in fact, be underestimated if a limited number of nests per year is used as the population abundance prior in the MSA. The Levantine coast (LBIS) nesting site had a surprisingly medium–high contribution, mainly as a result of two turtles recovered from the coast of Slovenia, in the northeastern Adriatic, carrying haplotype CC-A29.1, only recorded in Israeli rookeries to date. This haplotype might also occur in other, larger rookeries that have not yet been sampled or are yet undiscovered, in which case the LBIS contribution to the Adriatic stock would be relatively lower. In fact, although rookeries from Cyprus and the southern and western Turkish coasts (pooled in the TKME and DYDL clusters, respectively) are closer to the Adriatic Sea than the Israeli nesting sites and noticeably more abundant (see Electronic Supplementary Material Table S1), they provided a similar or smaller contribution than the Levantine rookeries. The possibility that haplotype CC-A29.1 could originate from rookeries other than the Israeli ones implies that unknown or poorly sampled nesting areas might be important to loggerhead recruitment yet to be quantified.
Contribution from Calabrian nesting sites was close to zero despite the relatively short geographical distance from the Adriatic Sea. This is most probably due to the small rookery size of Calabria and the sea currents flowing southwards along the Ionian coast of Italy, hindering migration to the Adriatic basin (Millot and Taupier-Letage 2005). Moreover, the rookery from Calabria shares haplotype CC-A31.1 with both western Greece and an occasional nesting site recently described in Sicily (Garofalo et al. 2016b). This makes detection of a Calabrian contribution to the Adriatic stock even more difficult. The same is true for another rare haplotype, CC-A10.4, described only in occasional nesting sites along Tyrrhenian beaches (Garofalo et al. 2016a; Maffucci et al. 2016). The three individuals carrying CC-A10.4 are likely to originate from Mediterranean colonies where CC-A10.4 has yet to be described rather than from Tyrrhenian rookeries. The CC-A10.4 haplotype may be part of the CC-A10 haplogroup (based on sequencing of 380 bp of the mtDNA control region) recorded in Greece by Laurent et al. (1998) but never observed afterwards in the same rookery that was also the major source of our Adriatic mixed stock.
In this study, we tried to optimize the MSA resolution primarily using a relatively large sample size. Although approximately 90% of the Adriatic stock included the most common CC-A2.1 and CC-A3.1 haplotypes, we found additional rare, rookery-specific sequences with respect to the previous studies based on much smaller sample sizes. We recorded haplotypes CC-A26.1 and CC-A29.1 so far described in the Libyan rookery of Sirte and the Israeli nesting sites, respectively. Haplotype CC-A20.1, which occurs in several Atlantic rookeries and only in one Mediterranean nesting site (Calabria, southern Italy), was recorded in one turtle from the northeastern Adriatic. Moreover, one turtle sampled in the northern Adriatic, off the coast of Slovenia had haplotype CC-A53.1, which is specific to eastern and central Turkish rookeries and never previously reported in the Adriatic (Yilmaz et al. 2012; Garofalo et al. 2013; Clusa et al. 2014).
We further improved the MSA performance in detecting mixed stocks from the Mediterranean baseline by removing and pooling baseline rookeries. We found that using nine single and clustered rookeries (namely the Libyan, Levantine, Cypriot, eastern, western and westernmost Turkish, Cretan, western Greek, and Calabrian) resulted in a relatively more informed baseline than using a set of reference sequences from 13 separate nesting grounds. Shamblin et al. (2014) pooled rookeries from Israel, Lebanon, Cyprus, and eastern and central Turkey into one single cluster despite significant ΦST values differentiating the Levantine and Cypriot nesting sites. Our scheme was similar to the one presented by Shamblin et al. (2014) except for retaining Cyprus, the two eastern Turkish sites (MTU and ETU) and the Levantine sites (LEB and ISR) as three distinct groups. The optimal grouping and/or splitting of rookery sites for baselines in MSA can, indeed, vary with geographical scale as well as conservation priorities. For instance, there may be compelling reasons to treat rookeries as demographically distinct for management purposes even in the absence of evidence of genetic differentiation. Formia et al. (2006) recommended recognition of Bioko and Ascension Island as demographically distinct rookeries despite lack of genetic divergence, given that they are distant by approximately 3000 km. A lack of marker resolution can make decisions on baseline structure even more complex, particularly at finer scale, such as in the eastern Mediterranean basin. Our pooling scheme considered geographically and ecologically close rookeries with similar genetic profiles, as recovered by Φ-statistics. This resulted in a relatively finer definition of some of the natal ground contributions to the Adriatic stock. For instance, the first MSA conducted with a baseline of 13 Mediterranean rookeries resulted in relatively wide confidence intervals, particularly for the contribution of the nesting sites in Misurata. Pooling of the Libyan rookeries drastically reduced the confidence intervals of the posterior probability of contribution to the Adriatic stock. Finally, priors weighted by population abundance (average number of nests per year) and length of migratory routes resulted in more realistic contributions from small, geographically distant rookeries. Approximately 20 and 10% of the Adriatic stock was found to include turtles from Lebanon and Israel, respectively, when using equal priors and priors weighted for population abundance. These values were reduced to less than 10% when using priors weighted for both abundance and geographical distance between nesting and foraging grounds.
MSA of Mediterranean foraging grounds is, nonetheless, hampered by two fundamental issues. First, the Mediterranean baseline is not a comprehensive one. For instance, frequencies of certain haplotypes were either large with respect to rookery size or inconsistent with the distance between sampling area and nesting sites. This suggests that haplotypes such as CC-A29.1 and CC-A10.4 might also originate at rookeries other than the ones described to date. Moreover, our finding of a novel mtDNA control region haplotype implies that more nesting areas need to be sampled (or sample sizes increased) to attain an inclusive baseline for sample assignment, particularly along the largely unexplored coasts of Libya. Second, the resolution power of currently available mtDNA control region haplotypes seems inadequate to accurately describe unique rookeries and their fine-scale contributions to foraging grounds. Analysis of both mtDNA sequences and microsatellite allele frequencies can better recover patterns of genetic structure among nesting grounds (Carreras et al. 2007). For instance, an integrated approach proved to be a powerful tool for MSA and individual assignment of loggerhead turtle from western Mediterranean feeding grounds to either Atlantic or Mediterranean RMUs, whereby a 35% increase in robust assignment was recovered when using both markers rather than mtDNA alone (Carreras et al. 2011). Studies on interindividual variation using additional mtDNA and nuclear markers should, therefore, be explored, to better inform our understanding of rookery dispersal and distribution throughout the Mediterranean (Duchene et al. 2012; Novelletto et al. 2016).
Notwithstanding the need for improved genetic sampling at several Mediterranean rookeries, our results confirm a high degree of connectivity between the Adriatic and western Greece and Crete, and, therefore, the importance of collaborative management efforts throughout the range of these populations. While migratory routes to and from the Adriatic deserve concerted conservation attention, genetic results indicate that foraging loggerheads do not exhibit preferential habitat use within Adriatic sub-regions. However, further research is needed to assess whether they may be able to shift foraging zones in response to local threats or whether depletion in more threatened areas such as the northern Adriatic and the Gulf of Manfredonia may be compensated by shifts in distributions of the foraging population.
References
Abreu-Grobois FA, Horrocks J, Formia A, Dutton PH, LeRoux R, Vélez-Zuazo X, Soares L, Meylan P (2006) New mtDNA Dloop primers which work for a variety of marine turtle species may increase the resolution of mixed stock analyses. In: Frick M, Panagopoulou A, Rees AF, Williams K (eds) Book of Abstracts. Twenty Sixth Annual Symposium on Sea Turtle Biology and Conservation, 2006. International Sea Turtle Society, Athens, Greece
Agostini VN, Bakun A (2002) `Ocean triads’ in the Mediterranean Sea: physical mechanisms potentially structuring reproductive habitat suitability (with example application to European anchovy, Engraulis encrasicolus). Fish Oceanogr 11:129–142. https://doi.org/10.1046/j.1365-2419.2002.00201.x
Artegiani A, Bregant D, Paschini E, Pinardi N, Raicich F, Russo A (1997) The Adriatic Sea general circulation. Part I: air-sea interactions and water mass structure. J Phys Oceanogr 27:1492–1514
Avens L, Braun-McNeill J, Epperly S, Lohmann KJ (2003) Site fidelity and homing behavior in juvenile loggerhead sea turtles (Caretta caretta). Mar Biol 143:211–220. https://doi.org/10.1007/s00227-003-1085-9
Bass AL, Epperly SP, Braun-McNeill J (2004) Multi-year analysis of stock composition of a loggerhead turtle (Caretta caretta) foraging habitat using maximum likelihood and Bayesian methods. Conserv Genet 5:783–796. https://doi.org/10.1007/s10592-004-1979-1
Bjorndal KA, Bolten AB (2008) Annual variation in source contributions to a mixed stock: implications for quantifying connectivity. Mol Ecol 17:2185–2193. https://doi.org/10.1111/j.1365-294X.2008.03752.x
Bolker B, Okuyama T (2014) Mixed stock analysis in R: getting started with the mixstock package. https://r-forge.r-project.org/R/?group_id=189
Bolker B, Okuyama T, Bjorndal K, Bolten A (2003) Sea turtle stock estimation using genetic markers: accounting for sampling error of rare genotypes. Ecol Appl 13:763–775. https://doi.org/10.1890/1051-0761(2003)013[0763:Stseug]2.0.Co;2
Bolten AB (1999) Techniques for measuring sea turtles. In: Eckert KL, Bjorndal KA, Abreu-Grobois FA, Donnelly M (eds) Research and management techniques for the conservation of sea turtles. IUCN/SSC Marine Turtle Specialist Group Publication No. 4, Consolidated Graphic Communications, Blanchard, PA, pp 110–114
Bowen BW, Bass AL, Chow S, Bostrom M, Bjorndal KA, Bolten AB, Okuyama T, Bolker BM, Epperly S, Lacasella E (2004) Natal homing in juvenile loggerhead turtles (Caretta caretta). Mol Ecol 13:3797–3808. https://doi.org/10.1111/j.1365-294X.2004.02356.x
Bowen BW, Bass AL, Soares L, Toonen RJ (2005) Conservation implications of complex population structure: lessons from the loggerhead turtle (Caretta caretta). Mol Ecol 14:2389–2402. https://doi.org/10.1111/j.1365-294X.2005.02598.x
Bowen BW, Grant WS, Hillis-Starr Z, Shaver DJ, Bjorndal KA, Bolten AB, Bass AL (2007) Mixed-stock analysis reveals the migrations of juvenile hawksbill turtles (Eretmochelys imbricata) in the Caribbean Sea. Mol Ecol 16:49–60. https://doi.org/10.1111/j.1365-294X.2006.03096.x
Carreras C, Pascual M, Cardona L, Aguilar A, Margaritoulis D, Rees A, Turkozan O, Levy Y, Gasith A, Aureggi M (2007) The genetic structure of the loggerhead sea turtle (Caretta caretta) in the Mediterranean as revealed by nuclear and mitochondrial DNA and its conservation implications. Conserv Genet 8:761–775. https://doi.org/10.1007/s10592-006-9224-8
Carreras C, Pascual M, Cardona L, Marco A, Bellido JJ, Castillo JJ, Tomás J, Raga JA, Sanfélix M, Fernández G, Aguilar A (2011) Living together but remaining apart: atlantic and Mediterranean loggerhead sea turtles (Caretta caretta) in shared feeding grounds. J Hered 102:666–677. https://doi.org/10.1093/Jhered/Esr089
Carreras C, Rees AF, Broderick AC, Godley BJ, Margaritoulis D (2014) Mitochondrial DNA markers of loggerhead marine turtles (Caretta caretta) (Testudines: Cheloniidae) nesting at Kyparissia Bay, Greece, confirm the western Greece unit and regional structuring. Sci Mar 78:115–124. https://doi.org/10.3989/scimar.03865.27b
Casale P (2011) Sea turtle by-catch in the Mediterranean. Fish Fish 12:299–316. https://doi.org/10.1111/j.1467-2979.2010.00394.x
Casale P, Margaritoulis D (eds) (2010) Sea turtles in the Mediterranean: distribution, threats and conservation priorities. IUCN, Gland
Casale P, Mariani P (2014) The first ‘lost year’of Mediterranean sea turtles: dispersal patterns indicate subregional management units for conservation. Mar Ecol Prog Ser 498:263–274. https://doi.org/10.3354/meps10640
Casale P, Simone G (2017) Seasonal residency of loggerhead turtles Caretta caretta tracked from the Gulf of Manfredonia, south Adriatic. Mediterr Mar Sci 18:4–10. https://doi.org/10.12681/mms.1663
Casale P, Laurent L, De Metrio G (2004) Incidental capture of marine turtles by the Italian trawl fishery in the north Adriatic Sea. Biol Conserv 119:287–295. https://doi.org/10.1002/aqc.841
Casale P, Freggi D, Basso R, Argano R (2005) Size at male maturity, sexing methods and adult sex ratio in loggerhead turtles (Caretta caretta) from Italian waters investigated through tail measurements. Herpetol J 15:145–148
Casale P, Freggi D, Basso R, Vallini C, Argano R (2007) A model of area fidelity, nomadism, and distribution patterns of loggerhead sea turtles (Caretta caretta) in the Mediterranean Sea. Mar Biol 152:1039–1049. https://doi.org/10.1007/s00227-007-0752-7
Casale P, Affronte M, Insacco G, Freggi D, Vallini C, Pino d’Astore P, Basso R, Paolillo G, Abbate G, Argano R (2010) Sea turtle strandings reveal high anthropogenic mortality in Italian waters. Aquat Conserv 20:611–620. https://doi.org/10.1002/aqc.1133
Casale P, Simone G, Conoscitore C, Conoscitore M, Salvemini P (2012) The Gulf of Manfredonia: a new neritic foraging area for loggerhead sea turtles in the Adriatic Sea. Acta Herpetol 7:1–12. https://doi.org/10.13128/Acta_Herpetol-9897
Casale P, Freggi D, Furii G, Vallini C, Salvemini P, Deflorio M, Totaro G, Raimondi S, Fortuna C, Godley BJ (2015) Annual survival probabilities of juvenile loggerhead sea turtles indicate high anthropogenic impact on Mediterranean populations. Aquat Conserv 25:690–700. https://doi.org/10.1002/aqc.2467
Casale P, Freggi D, Rigoli A, Ciccocioppo A, Luschi P (2017) Geometric morphometrics, scute patterns and biometrics of loggerhead turtles (Caretta caretta) in the central Mediterranean. Amphibia-Reptilia 38:145–156. https://doi.org/10.1163/15685381-00003096
Chaieb O, El Ouaer A, Maffucci F, Bradai MN, Bentivegna F, Said K, Chatti N (2010) Genetic survey of loggerhead turtle Caretta caretta nesting population in Tunisia. Mar Biodivers Rec 3:e20. https://doi.org/10.1017/S175526721000014X
Clement M, Posada D, Crandall K (2000) TCS: a computer program to estimate gene genealogies. Mol Ecol 9:1657–1659. https://doi.org/10.1046/j.1365-294x.2000.01020.x
Clusa M, Carreras C, Pascual M, Demetropoulos A, Margaritoulis D, Rees AF, Hamza AA, Khalil M, Aureggi M, Levy Y (2013) Mitochondrial DNA reveals Pleistocenic colonisation of the Mediterranean by loggerhead turtles (Caretta caretta). J Exp Mar Biol Ecol 439:15–24. https://doi.org/10.1016/j.jembe.2012.10.011
Clusa M, Carreras C, Pascual M, Gaughran SJ, Piovano S, Giacoma C, Fernández G, Levy Y, Tomás J, Raga JA (2014) Fine-scale distribution of juvenile Atlantic and Mediterranean loggerhead turtles (Caretta caretta) in the Mediterranean Sea. Mar Biol 161:509–519. https://doi.org/10.1007/s00227-013-2353-y
Coll M, Piroddi C, Albouy C, Ben Rais Lasram F, Cheung WWL, Christensen V, Karpouzi VS, Guilhaumon F, Mouillot D, Paleczny M, Palomares ML, Steenbeek J, Trujillo P, Watson R, Pauly D (2012) The Mediterranean Sea under siege: spatial overlap between marine biodiversity, cumulative threats and marine reserves. Global Ecol Biogeogr 21:465–480. https://doi.org/10.1111/j.1466-8238.2011.00697.x
Cushman-Roisin B, Gačić M, Poulain P-M, Artegiani A (eds) (2001) Physical oceanography of the Adriatic Sea. Past, present and future. Kluwer Academic Publishers, Dordrecht
Drosopoulou E, Tsiamis G, Mavropoulou M, Vittas S, Katselidis KA, Schofield G, Palaiologou D, Sartsidis T, Bourtzis K, Pantis J, Scouras ZG (2012) The complete mitochondrial genome of the loggerhead turtle Caretta caretta (Testudines: Cheloniidae): Genome description and phylogenetic considerations. Mitochondr DNA 23:1–12. https://doi.org/10.3109/19401736.2011.637109
Duchene S, Frey A, Alfaro-Nunez A, Dutton PH, Gilbert MTP, Morin PA (2012) Marine turtle mitogenome phylogenetics and evolution. Mol Phylogenet Evol 65:241–250. https://doi.org/10.1016/j.ympev.2012.06.010
Eckert KL, Bjorndal KA, Abreu-Grobois FA, Donnelly M (eds) (1999) Research and management techniques for the conservation of sea turtles. IUCN/SSC Marine Turtle Specialist Group Publication No. 4, Consolidated Graphic Communications, Blanchard
Excoffier L, Lischer HE (2010) Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour 10:564–567. https://doi.org/10.1111/j.1755-0998.2010.02847.x
Excoffier L, Smouse PE, Quattro JM (1992) Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131:479–494
Formia A, Godley BJ, Dontaine JF, Bruford MW (2006) Mitochondrial DNA diversity and phylogeography of endangered green turtle (Chelonia mydas) populations in Africa. Conserv Genet 7:353–369. https://doi.org/10.1007/s10592-005-9047-z
Fortuna CM, Vallini C, Filidei E Jr, Ruffino M, Consalvo I, Di Muccio S, Gion C, Scacco U, Tarulli E, Giovanardi O, Mazzola A (2010) By-catch of cetaceans and other species of conservation concern during pair trawl fishing operations in the Adriatic Sea (Italy). Chem Ecol 26:65–76. https://doi.org/10.1080/02757541003627662
Franzellitti S, Locatelli C, Gerosa G, Vallini C, Fabbri E (2004) Heavy metals in tissues of loggerhead turtles (Caretta caretta) from the northwestern Adriatic Sea. Comp Biochem Phis C 138:187–194. https://doi.org/10.1016/j.cca.2004.07.008
Garofalo L, Mastrogiacomo A, Casale P, Carlini R, Eleni C, Freggi D, Gelli D, Knittweis L, Mifsud C, Mingozzi T (2013) Genetic characterization of central Mediterranean stocks of the loggerhead turtle (Caretta caretta) using mitochondrial and nuclear markers, and conservation implications. Aquat Conserv 23:868–884. https://doi.org/10.1002/aqc.2338
Garofalo L, Marsili L, Terracciano G, Baini M, Senese M, Mancusi C, Lorenzini R (2016a) Welcome to the North! Genetic makeup of loggerhead Caretta caretta females from the northernmost Italian nesting site. In: Menegon M, Rodriguez-Prieto A, Deflorian MC (eds) Atti XI Congresso Nazionale Societas Herpetologica Italica. Ianieri Edizioni, Trento, Italy. https://doi.org/10.13140/rg.2.2.14077.15849
Garofalo L, Insacco G, Scaravelli D, Lorenzini R (2016b) On the connectivity between nesting colonies of the loggerhead turtle Caretta caretta in the Ionian Region. In: Menegon M, Rodriguez-Prieto A, Deflorian MC (eds) Atti XI Congresso Nazionale Societas Herpetologica Italica. Ianieri Edizioni, Trento, Italy, pp 135–136. https://doi.org/10.13140/rg.2.2.34209.81766
Garoia F, Guarniero I, Piccinetti C, Tinti F (2004) First microsatellite loci of red mullet (Mullus barbatus) and their application to genetic structure analysis of Adriatic shared stock. Mar Biotechnol 6:446–452. https://doi.org/10.1007/s10126-004-3045-x
Gaspari S, Holcer D, Mackelworth P, Fortuna C, Frantzis A, Genov T, Vighi M, Natali C, Rako N, Banchi E, Chelazzi G, Ciofi C (2015) Population genetic structure of common bottlenose dolphins (Tursiops truncatus) in the Adriatic Sea and contiguous regions: implications for international conservation. Aquat Conserv 25:212–222. https://doi.org/10.1002/aqc.2415
Gelman A, Rubin DB (1992) Inference from iterative simulation using multiple sequences. Stat Sci 7:457–472
Halpern BS, Selkoe KA, Micheli F, Kappel CV (2007) Evaluating and ranking the vulnerability of global marine ecosystems to anthropogenic threats. Conserv Biol 21:1301–1315
Hamann M, Godfrey MH, Seminoff JA, Arthur K, Barata PCR, Bjorndal KA, Bolten AB, Broderick AC, Campbell LM, Carreras C (2010) Global research priorities for sea turtles: informing management and conservation in the 21st century. Endanger Species Res 11:245–269. https://doi.org/10.3354/esr00279
Hamza AA (2010) Libya. In: Casale P, Margaritoulis D (eds) Sea turtles in the Mediterranean: distribution, threats and conservation priorities. IUCN, Gland, pp 157–170
Hawkes L, Broderick AC, Coyne MS, Godfrey MH, Godley BJ (2007) Only some like it hot—quantifying the environmental niche of the loggerhead sea turtle. Divers Distrib 13:447–457. https://doi.org/10.1111/j.1472-4642.2007.00354.x
Hays GC, Fossette S, Katselidis KA, Mariani P, Schofield G (2010) Ontogenetic development of migration: lagrangian drift trajectories suggest a new paradigm for sea turtles. J R Soc Interface 7:1319–1327. https://doi.org/10.1098/rsif.2010.0009
Laurent L, Casale P, Bradai MN, Godley BJ, Broderick GG, Schroth W, Schierwater B, Levy AM, Freggi D, Abd El-Mawla EM (1998) Molecular resolution of marine turtle stock composition in fishery bycatch: a case study in the Mediterranean. Mol Ecol 7:1529–1542. https://doi.org/10.1046/j.1365-294x.1998.00471.x
Lazar B, Gračan R (2011) Ingestion of marine debris by loggerhead sea turtles, Caretta caretta, in the Adriatic Sea. Marine Poll Bull 62:43–47. https://doi.org/10.1016/j.marpolbul.2010.09.013
Lazar B, Borboroglu PG, Tvrtković N, Žiža V (2002) Temporal and spatial distribution of the loggerhead sea turtle, Caretta caretta, in the eastern Adriatic Sea: a seasonal migration pathway? In: Seminoff JA (compiler) Proceedings of the twenty-second annual symposium on sea turtle biology and conservation. NOAA Technical Memorandum NMFS-SEFSC-503, Miami, pp 283–284
Lazar B, Margaritoulis D, Tvrtković N (2004) Tag recoveries of the loggerhead sea turtle Caretta caretta in the eastern Adriatic Sea: implications for conservation. J Mar Biol Assoc UK 84:475–480. https://doi.org/10.1017/S0025315404009488h
Lazar B, Gračan R, Katić J, Zavodnik D, Jaklin A, Tvrtković N (2011) Loggerhead sea turtles (Caretta caretta) as bioturbators in neritic habitats: an insight through the analysis of benthic molluscs in the diet. Mar Ecol 32:65–74. https://doi.org/10.1111/j.1439-0485.2010.00402.x
Leigh JW, Bryant D (2015) POPART: full-feature software for haplotype network construction. Methods Ecol Evol 6:1110–1116. https://doi.org/10.1111/2041-210X.12410
Luschi P, Casale P (2014) Movement patterns of marine turtles in the Mediterranean Sea: a review. Ital J Zool 81:478–495. https://doi.org/10.1080/11250003.2014.963714
Maffucci F, Corrado R, Palatella L, Borra M, Marullo S, Hochscheid S, Lacorata G, Iudicone D (2016) Seasonal heterogeneity of ocean warming: a mortality sink for ectotherm colonizers. Sci Rep UK 6:Article no. 23983 https://doi.org/10.1038/srep23983
Margaritoulis D, Argano R, Baran I, Bentivegna F, Bradai MN, Camiñas JA, Casale P, De Metrio G, Demetropoulos A, Gerosa G (2003) Loggerhead turtles in the Mediterranean: present knowledge and conservation perspectives. In: Bolten AB, Wutherington BE (eds) Loggerhead sea turtles. Smithsonian Institution Press, Washington DC, pp 175–198
Millot C, Taupier-Letage I (2005) Circulation in the Mediterranean sea. In: Saliot A (ed) The Mediterranean Sea. Springer, Berlin, pp 323–334 https://doi.org/10.1007/b107143
Monzón-Argüello C, Rico C, Carreras C, Calabuig P, Marco A, López-Jurado LF (2009) Variation in spatial distribution of juvenile loggerhead turtles in the eastern Atlantic and western Mediterranean Sea. J Exp Mar Biol Ecol 373:79–86. https://doi.org/10.1016/j.jembe.2009.03.007
Naro-Maciel E, Gaughran SJ, Putman NF, Amato G, Arengo F, Dutton PH, McFadden KW, Vintinner EC, Sterling EJ (2014) Predicting connectivity of green turtles at Palmyra Atoll, central Pacific: a focus on mtDNA and dispersal modelling. J R Soc Interface 11:20130888. https://doi.org/10.1098/rsif.2013.0888
Norman JA, Moritz C, Limpus CJ (1994) Mitochondrial DNA control region polymorphisms: genetic markers for ecological studies of marine turtles. Mol Ecol 3:363–373. https://doi.org/10.1111/j.1365-294X.1994.tb00076.x
Novelletto A, Testa L, Iacovelli F, Blasi P, Garofalo L, Mingozzi T, Falconi M (2016) Polymorphism in mitochondrial coding regions of Mediterranean loggerhead turtles: evolutionary relevance and structural effects. Physiol Biochem Zool 89:473–486. https://doi.org/10.1086/688679
Peakall R, Smouse PE (2012) GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 282:537–2539
Pella J, Masuda M (2001) Bayesian methods for analysis of stock mixtures from genetic characters. Fish B-NOAA 99:151
Poulain P-M (2001) Adriatic Sea surface circulation as derived from drifter data between 1990 and 1999. J Mar Syst 29:3–32
Rees AF, Carreras C, Broderick AC, Margaritoulis D, Stringell TB, Godley BJ (2017) Linking loggerhead locations: using multiple methods to determine the origin of sea turtles in feeding grounds. Mar Biol 164:30. https://doi.org/10.1007/s00227-016-3055-z
Revelles M, Carreras C, Cardona L, Marco A, Bentivegna F, Castillo JJ, de Martino G, Mons JL, Smith MB, Rico C, Pascual M, Aguilar A (2007) Evidence for an asymmetrical size exchange of loggerhead sea turtles between theMediterranean and the Atlantic through the Straits of Gibraltar. J Exp Mar Biol Ecol 349:261–271. https://doi.org/10.1016/j.jembe.2007.05.018
Ruggeri P, Splendiani A, Occhipinti G, Fioravanti T, Santojanni A, Leonori I, De Felice A, Arneri E, Procaccini G, Catanese G, Tičina V, Bonanno A, Nisi Cerioni P, Giovannotti M, Grant WS, Caputo Barucchi V (2016) Biocomplexity in populations of European anchovy in the Adriatic Sea. PLoS One 11:e0153061. https://doi.org/10.1371/journal.pone.0153061
Sambrook JF, Russel DW (eds) (2001) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor
Schofield G, Bishop CM, Katselidis KA, Dimopoulos P, Pantis JD, Hay GC (2009) Microhabitat selection by sea turtles in a dynamic thermal marine environment. J Anim Ecol 78:14–21. https://doi.org/10.1111/j.1365-2656.2008.01454.x
Shamblin BM, Bolten AB, Bjorndal KA, Dutton PH, Nielsen JT, Abreu-Grobois FA, Reich KJ, Witherington BE, Bagley DA, Ehrhart LM (2012) Expanded mitochondrial control region sequences increase resolution of stock structure among North Atlantic loggerhead turtle rookeries. Mar Ecol Prog Ser 469:145–160. https://doi.org/10.3354/meps09980
Shamblin BM, Bolten AB, Abreu-Grobois FA, Bjorndal KA, Cardona L, Carreras C, Clusa M, Monzón-Argüello C, Nairn CJ, Nielsen JT (2014) Geographic patterns of genetic variation in a broadly distributed marine vertebrate: new insights into loggerhead turtle stock structure from expanded mitochondrial DNA sequences. PLoS One 9:e85956. https://doi.org/10.1371/journal.pone.0085956
Templeton AR, Crandall KA, Sing CF (1992) A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genet Mol Biol 132:619–633
Wallace BP, DiMatteo AD, Hurley BJ, Finkbeiner EM, Bolten AB, Chaloupka MY, Hutchinson BJ, Abreu-Grobois FA, Amorocho D, Bjorndal KA (2010) Regional management units for marine turtles: a novel framework for prioritizing conservation and research across multiple scales. PLoS One 5:e15465. https://doi.org/10.1371/journal.pone.0015465
Yilmaz C, Turkozan O, Bardakcı F, White M, Kararaj E (2012) Loggerhead turtles (Caretta caretta) foraging at Drini Bay in Northern Albania: genetic characterisation reveals new haplotypes. Acta Herpetol 7:155–162. https://doi.org/10.13128/Acta_Herpetol-10286
Zbinden JA, Aebischer A, Margaritoulis D, Arlettaz R (2008) Important areas at sea for adult loggerhead sea turtles in the Mediterranean Sea: satellite tracking corroborates findings from potentially biased sources. Mar Biol 153:899–906. https://doi.org/10.1007/s00227-007-0862-2
Zbinden JA, Bearhop S, Bradshaw P, Gill B, Margaritoulis D, Newton J, Godley BJ (2011) Migratory dichotomy and associated phenotypic variation in marine turtles revealed by satellite tracking and stable isotope analysis. Mar Ecol Prog Ser 421:291–302. https://doi.org/10.3354/meps08871
Acknowledgements
This study was funded by the University of Florence, the Italian Department of Fishery and Aquaculture (BYCATCH Project—Ministry of Agriculture, Food and Forestry), the Italian National Institute for Environmental Protection and Research, the European Union NETCET project (IPA Inter-Adriatic), and the Slovenian Research Agency under Grant P1-0386. We thank Fondazione Cetacea ONLUS, the Veneto stranding network (Coordinamento Tartarughe marine del Litorale Veneto), WWF (Area Marina Protetta di Miramare, Trieste), and F. Zuppa and M. Tempesta from APS DelTa for support and for providing samples from the northwestern Adriatic. We are also grateful to V. Angelini, E. Filidei jr, L. Mizzan, G. Palmisano, P. Perlasca, S. Pari, and E. Zanetti for their help with sample collection and to P. Blasi and L. Testa for the assistance during lab work. All collaborating fishermen are particularly acknowledged.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Ethical approval
All applicable international, national, and institutional guidelines for the care of animals were followed. This work was conducted under all necessary permits. Live bycaught sea turtles were released after sample collection according to the IUCN/SSC Marine Turtle Specialist Group guidelines (Eckert et al. 1999). None of the procedures used in the study met the criteria which would define them as “experiments” according to article 2 of the Council of European Communities Directive 86/609/EEC on the protection of animals used for experimental and other scientific purposes. Approval by the institutional ethics committee (Commissione per l’Etica della Ricerca di Ateneo, University of Florence) was not required.
Additional information
Responsible Editor: C. Eizaguirre.
Reviewed by B. Shamblin, Y. Tikochinski and an undisclosed expert.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Tolve, L., Casale, P., Formia, A. et al. A comprehensive mitochondrial DNA mixed-stock analysis clarifies the composition of loggerhead turtle aggregates in the Adriatic Sea. Mar Biol 165, 68 (2018). https://doi.org/10.1007/s00227-018-3325-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-018-3325-z