
Abstract
A sedentary lifestyle, inadequate diet, and obesity are substantial risk factors for Type 2 diabetes mellitus (T2DM) development. A major picture of T2DM is insulin resistance (IR), which causes many impairments in brain physiology, such as increased proinflammatory state and decreased brain-derived neurotrophic factor (BDNF) concentration, hence reducing cognitive function. Physical exercise is a non-pharmacological tool for managing T2DM/IR and its complications. Thus, this study investigated the effects of IR induction and the acute effects of resistance exercise (RE) on memory, neurotrophic, and inflammatory responses in the hippocampus and prefrontal cortex of insulin-resistant rats. IR was induced by a high-fat diet and fructose-rich beverage. Insulin-resistant rats performed acute resistance exercise (IR.RE; vertical ladder climb at 50–100% of the maximum load) or rest (IR.REST; 20 min). Cognitive parameters were assessed by novel object recognition (NOR) tasks, and biochemical analyses were performed to assess BDNF concentrations and inflammatory profile in the hippocampus and prefrontal cortex. Insulin-resistant rats had 20% worse long-term memory (LTM) (p < 0.01) and lower BDNF concentration in the hippocampus (−14.6%; p < 0.05) when compared to non-insulin-resistant rats (CON). An acute bout of RE restored LTM (−9.7% pre vs. post; p > 0.05) and increased BDNF concentration in the hippocampus (9.1%; p < 0.05) of insulin-resistant rats compared to REST. Thus, an acute bout of RE can attenuate the adverse effects of IR on memory and neurotrophic factors in rats, representing a therapeutic tool to alleviate the IR impact on the brain.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Type 2 diabetes mellitus (T2DM) has the highest prevalence among cases of diabetes in the world, representing about 90–95% of all cases diagnosed with the disease (Elsayed et al. 2023a). Its specific etiology is still not known, but although the individual predisposition to T2DM has a strong genetic basis, studies suggest that its increase is related to the global expansion of the sedentary lifestyle, inadequate eating patterns, and consequently, obesity (Mozaffarian et al. 2009; Chatterjee et al. 2017). The main characteristic of T2DM is insulin resistance (IR) in peripheral tissues, which results in hyperglycemia. Chronically, this metabolic disruption stresses the pancreas, causing an overproduction of insulin to maintain normoglycemia (Robertson et al. 2003).
In addition to the hyperglycemic and hyperinsulinemic condition, IR provokes several metabolic disorders, such as mitochondrial dysfunction (Chen et al. 2015; DeFronzo et al. 2015) and increased production of reactive oxygen species (Giacco et al. 2011). Besides that, IR can lead to an increase in proinflammatory cytokines, including tumor necrosis factor-alpha (TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6), and reduction of anti-inflammatory cytokines, such as the interleukin-10 (IL-10).
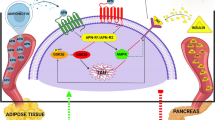
The central nervous system (CNS) also undergoes several changes resulting from insulin resistance (Sharma 2021). Among them are disruptions in the intracellular cascade of the insulin pathway (Piroli et al. 2007; Pratchayasakul et al. 2011), reduced translocation of GLUT4 to the cell membrane (Winocur et al. 2005; Piroli et al. 2007), increased proinflammatory state (Dinel et al. 2011; Puig et al. 2012) and reduction of brain neurotrophins concentrations (BDNF, VEGF, and IGF-1) (Lindqvist et al. 2006; Park et al. 2010). In the last instance, those insulin resistance-induced alterations in the CNS can collectively lead to cognitive impairments (Dinel et al. 2011; Leal et al. 2015; Park et al. 2019).
Physical exercise has been used as a non-pharmacological strategy in managing IR and its complications. It is mainly accomplished by stimulating non-insulin-dependent pathways to reduce blood glucose levels and by increased insulin sensitivity (Richter and Hargreaves 2013; Sylow et al. 2017). Growing evidence suggests resistance exercise as a therapeutic tool for type 2 diabetes (Elsayed et al. 2023b). Resistance exercise may help phenotypic alterations commonly observed in T2DM individuals, such as the decline in muscle mass and strength (Leenders et al. 2013). Resistance exercise is also promising to counteract T2DM-related cognitive dysfunction (De Sousa et al. 2020). A recent study of our group showed that a single session of resistance exercise can provoke cognitive facilitation, increasing attentional and executive function (frontal lobe-related functions) in middle-aged and older adults with T2DM (Silveira-Rodrigues et al. 2023). However, the effects of a resistance exercise session in IR animals and the mechanisms underpinning these effects in distinct brain regions remain unknown.
There is evidence that a single session of RE had positive effects on glucometabolic outcomes (Craft 2007; Pesta et al. 2017). Also, acutely, a single RE session improves cognitive function in middle-aged and older adults with T2DM (Silveira-Rodrigues et al. 2023) and mild cognitive impairment (Tsai et al. 2018). However, further studies are needed to understand the underlying mechanisms involved in cognitive function amelioration in the brain of IR rodents.
Thus, the present study investigated the acute effects of a bout of resistance exercise on memory, inflammatory status, and BDNF concentration in insulin-resistant rat’s brain. We hypothesized that an acute bout of resistance exercise would improve short and long-term memory, increase anti and reduce proinflammatory cytokines, and increase BDNF concentration in the hippocampus and prefrontal cortex of insulin-resistant rats.
Materials and methods
Animals and ethics care
Thirty-four male Wistar rats (250 g) were used in the experiments. All experimental procedures were approved by the Ethics Committee Care and Use of Laboratory Animals of the Federal University of Minas Gerais (CEUA—UFMG) and were conducted following the Brazilian National Council for the Control of Animal Experimentation (CONCEA) (protocols no. 200; 199/2021). The rats were housed in collective cages (four rats in each cage) under controlled temperature (24 °C) and light conditions (12/12 h).
Experimental design
The experimental design was organized into two experiments. In the first, 18 rats were weighed on a digital scale (FILIZOLA®) and randomized, stratified by weight, into two groups: control (CON; n = 9) and insulin resistant (IR; n = 9). After balancing, the rats were submitted to the novel object recognition (NOR) task to assess short-term (STM) and long-term (LTM) memory on three consecutive days (Bevins and Besheer 2006). After the last task day, IR group underwent the dietary induction protocol for insulin resistance (Melo et al. 2021). At the end of 60 days, the rats went through the oral glucose tolerance test (OGTT) to confirm insulin resistance. Once insulin resistance has been confirmed, 48 h after the OGTT both groups (CON and IR) performed the NOR task again to verify the effect of insulin resistance on STM and LTM. Then, the rats were euthanized by decapitation for tissue removal and subsequent biochemical analysis.
In the second experiment, the rats were habituated to the cage for 7 days and 16 animals were submitted to the NOR task. Next, the rats underwent the dietary induction protocol for insulin resistance (Melo et al. 2021). After 60 days, the rats were randomly divided into two groups: insulin resistant + rest (IR.REST; n = 8) and insulin resistant + acute resistance exercise (IR.RE; n = 8).
The IR.RE group was familiarized with the vertical ladder for three consecutive days and, 48 h later, submitted to the MWC test to determine the load of the exercise session (Neves et al. 2016; Padilha et al. 2019). Twenty-four hours after the MWC, both groups were submitted again to the NOR task, performed on three consecutive days.
On the first day, familiarization with the arena was carried out. On the second day, the second and third stages were performed for the training and the first test sessions, respectively. The experimental session (rest or acute resistance exercise) was performed one day after the third stage. The experimental sessions lasted 20 min and were performed in the morning (between 7:00 and 9:00 AM).
Immediately after the acute session, the rats were submitted to the fourth stage of the NOR task, aimed at evaluating the LTM and verifying the possible acute effects of the RE on this variable. Thirty minutes after the intervention (rest or resistance exercise), the rats were euthanized by decapitation for tissue removal and subsequent biochemical analysis (Fig. 1).

Experimental design. HAB: habituation; CON: control group; IR: insulin resistant group; NOR: novel object recognition; HFD: high-fat diet; OGTT: oral glucose tolerance test; FAM: vertical ladder familiarization; MWC: maximal weight carried test; IR.REST: insulin resistant + rest group; IR.RE: insulin resistant + acute resistance exercise group; REST: Rest intervention; RE: Acute resistance exercise intervention; LTM: Long-term memory
Diet intervention
The high-fat diet was provided in the form of pellets with a shape and texture similar to the standard chow (Presence®). It comprised 78% ground standard chow and 22% saturated fat. Thus, for each 500 g of feed, the high-fat diet consisted of 390 g of bran from the standard chow, 110 g of vegetable fat (Primor®), and 40 g of sour starch (Pacha®) dissolved in 400 mL of water. After mixing, the dough was placed in stainless steel molds and baked in an electric oven at 180 °C for 2 h. Fructose (Synth®) was dissolved in water (30% solution), and rats had ad libitum access. Food intake, fructose-rich beverage intake (Supplementary Material), and body mass of the rats were recorded throughout the intervention using a digital scale (Filizola®) (Melo et al. 2021).
Novel object recognition (NOR)
The NOR task is a consolidated test in the literature to assess short and long-term memory (Capettini et al. 2011; Crunfli et al. 2018, 2019; De Sousa et al. 2020). It was performed in four stages for three consecutive days. On the first day, familiarization with the arena was performed and consisted of positioning the rats in the center of a square box (70 cm × 70 cm × 40 cm) for free 5-min exploration.
On the second day, there were two stages. In the first one, the rats were reinserted in the arena with the presence of two identical objects (object A) for the training session. For this, they were positioned at the midpoint of the arena with the nose pointed to the opposite side of the two objects and had 10 min for free exploration (Bevins and Besheer 2006; De Sousa et al. 2020). The second stage was conducted 1 h after the training session, and the animals were placed back in the arena for the test session (STM analysis), where one of the A objects was exchanged for a new object (object B). After that the animals were reinserted in the arena to underwent 5 min of free exploration.
On the third day, the fourth and final stage (LTM analysis) was performed. For this, 24 h after the last test session, the rats were placed back in the arena in the presence of two objects, A and C, being this took the place of object B. After that, the animals had 5 min to explore freely (Capettini et al. 2011; Crunfli et al. 2018, 2019). The arena and objects used during each session were cleaned between trials with 70% ethanol, to eliminate olfactory cues. The amount of time spent exploring both the familiar object and the new object was recorded for STM and LTM analysis (De Sousa et al. 2020).
Exploratory behavior was analyzed in two ways. In the first one, a trained researcher recorded the exploration time of each object (familiar and new) with two stopwatches, one for each object, every time the animal’s nose was in contact with or close to the object (less than 1 cm) (De Sousa et al. 2020). In the second one, the exploration was analyzed using one software (Any-Maze Video Tracking System 7.16; Stoelting Co. © 2022). A Student’s t test was performed to compare the values recorded by the researcher and by the software, and no statistical difference was verified between the analyzes (p > 0.05).
The recognition index was calculated using the equation TN/(TN + TF) where TN is the time spent exploring the new object and TF is the time spent exploring the familiar object (Rojas et al. 2013). The value of the discrimination index can vary from 0 to 1 (Fonseca et al. 2013).
Oral glucose tolerance test (OGTT)
The rats were fasted for 6 h. After that, a solution of D’Glucose (1 g kg−1 of body mass, Synth®) dissolved in water (40% solution) was administered via oral gavage. Glucose concentrations were assessed by portable glucometer (Accu-Chek Performa®) through blood samples of the tail, obtained before and 15, 30, 60, 90, and 120 min after administration of the D’Glucose solution. A drop of blood was obtained through a small cut in the distal region of the tail (Melo et al. 2021).
Vertical ladder familiarization
The rats were familiarized with a vertical ladder inclined at 80° (110 cm × 18 cm, with 2 cm between the steps). The vertical ladder familiarization protocol consisted of three trials per day for 3 days. Initially, the rats were kept for 60 s in the housing chamber. Then, they were placed on the ladder at 35 cm, 55 cm, and 110 cm (bottom) from the top so that they could move to the housing chamber. Between each trial, the animals remained in the housing chamber for 60 s (Pereira et al. 2010; Cassilhas et al. 2012a).
Maximal weight carried test (MWC)
Initially, all rats (CON and IR) ascended a ladder with a load attached to their tail using adhesive tape (Scotch 3 M™), equivalent to 75% of their body weight. After successfully ascending, an additional 30 g was added, and this process was repeated until a load was reached where the rat could not climb the entire length of the ladder for three consecutive attempts. The MWC was defined as the load of the last successful climb (Neves et al. 2016; Padilha et al. 2019). MWC values were comparable in both groups (CON = 365 ± 27.45 g; IR = 346 ± 77.08 g; p = 0.562). Specifically, in IR.RE group (366 g ± 53.4 g), this test was used to determine the maximal workload and to prescribe the workload for resistance exercise sessions.
Acute resistance exercise
The rats of the IR.RE group were submitted to the acute resistance exercise. The session consisted of eight stair climbs with a 60-s interval between them. The load progressively increased throughout the session, as shown in Table 1 (adapted from Cassilhas et al. 2012b).
Euthanasia
The rats were euthanized by decapitation without prior sedation. The brain (hippocampus and prefrontal cortex) and white adipose tissues (retroperitoneal, mesenteric, and epididymal) were removed for further analysis.
Adiposity index
To determine the adiposity index (AI), the retroperitoneal (RAT), mesenteric (MAT), and epididymal (EAT) adipose tissues were weighed on a scale (Shimadzu® Model BL320H), and the following equation was used (Lacerda et al. 2015):
Biochemical analyses
The ELISA method was used to analyze interleukin 6 (IL-6), tumor necrosis factor-alpha (TNF-α), interleukin 10 (IL-10) and brain-derived neurotrophic factor (BDNF) concentrations in the hippocampus and prefrontal cortex (Rat IL-6 DuoSet ELISA; Rat IL-10 DuoSet; Rat TNF-alpha DuoSet; Human/Mouse BDNF DuoSet ELISA kit, R&D Systems, Minneapolis, MN, USA). These areas were chosen because of their relationship with learning and memory (hippocampus), attention and decision-making (prefrontal cortex). The manufacturer’s recommendations were followed.
Statistical analyses
Data are presented as mean ± standard deviation (SD). Data normality and homoscedasticity were evaluated using the Shapiro–Wilk and Levene tests, respectively.
The student’s t test was used to compare body composition values, the area under the curve (AUC) in the OGTT, BDNF, and cytokines concentrations between groups (CON vs. IR or IR.REST vs. IR.RE). The ANOVA-two-way test of repeated measures was used to analyze the OGTT, STM, and LTM, considering the factors of time (pre vs. post) and group (CON vs. IR or IR.REST vs. IR.RE). When significant interactions were found, Tukey’s post hoc test was performed to compare pairwise differences. The Pearson test was performed to investigate the correlation between BDNF concentration and long-term memory restoration. All analyzes were performed using SigmaPlot 14.5 software (SYSTAT SOFTWARE, USA), adopting a significance level of 5% (α < 0.05).
Results
Experiment 1: Body mass and adiposity index
When analyzing the variation (after 60 days of dietary induction of IR minus pre-induction) in body mass change, no significant difference was found between the IR and CON groups (IR: 124.85 ± 29.54 g vs. CON: 122.42 ± 20.32 g; p = 0.861). However, the IR group had a significantly higher adiposity index when compared to the CON group (IR: 5.23 ± 1.05% vs. CON: 4.06 ± 0.73%; p < 0.05).
Experiment 1: Oral glucose tolerance test (OGTT)
After 60 days, IR group blood glucose values were higher compared to baseline at moments 15, 30, and 60 (15 min: 182 ± 19.53 mg dL−1; 30 min: 150 ± 13.20 mg dL−1; 60 min: 140 ± 12.94 mg dL−1 vs. 0 min: 116 ± 4.79 mg dL−1; p < 0.001). Furthermore, hyperglycemia was maintained until moment 90 (90 min: 133 ± 11.21 mg dL−1 vs. 0 min: 116 ± 4.79 mg dL−1; p < 0.05), returning to the baseline value only at moment 120 (120 min: 127 ± 12.24 mg dL−1 vs. 116 ± 4.79 mg dL−1; p = 0.195).
Otherwise, the CON group increased blood glucose values at moments 15 and 30 when compared to baseline (15 min: 169 ± 21.57 mg dL−1; 30 min: 144 ± 11.32 mg dL−1 vs. 0 min: 108 ± 4.07 mg dL−1; p < 0.001), returning to the baseline value at time 60 (60 min: 119 ± 4.40 mg dL−1 vs. 0 min: 108 ± 4.07 mg dL−1; p = 0.269) (Fig. 2a).

Oral glucose tolerance test post 60 days of diet. a OGTT glucose; b AUC TTOG. Mean ± SD. * Significant difference vs. minute 0 (p < 0.05); *** significant difference vs. minute 0 (p < 0.001); # significant difference vs. CON (p < 0.05); ## significant difference vs. CON (p < 0.01). CON (n = 7); IR (n = 7)
Furthermore, the IR group had a significantly greater area under the curve (AUC) when compared to the CON group (17,060 ± 1271.84 vs. 15,105 ± 685.70 mg dL−1; p < 0.01) (Fig. 2b).
Experiment 1: Short and long-term memories
NOR task allowed us to verify that there was no statistically significant difference in the STM either in the time factor (p = 0.798), group factor (p = 0.433), or group × time interaction (p = 0.312) (Fig. 3a). However, when analyzing the LTM, it was possible to observe a decrease in the recognition index in the IR group comparing pre and post-moments (pre: 0.70 ± 0.08 vs. post: 0.56 ± 0.11; p < 0.01). Furthermore, IR exhibited a lower recognition index compared to the CON group in post-moment (IR: 0.56 ± 0.11 vs. CON: 0.70 ± 0.09; p < 0.01) (Fig. 3b). These findings suggest that IR provoked a decline in long-term memory without changing short-term memory.

Short and long-term memory pre and post-60 days of diet. a STM-NOR; b LTM-NOR. Mean ± SD. ** Significant difference vs. pre (p < 0.01); ## significant difference vs. CON-post (p < 0.01). CON (n = 9); IR (n = 9)
Experiment 1: Brain-derived neurotrophic factor (BDNF)
Assuming that BDNF and its receptors TrkB are expressed in multiple brain areas, such as the prefrontal cortex and hippocampus, we evaluated BDNF concentrations in these areas to improve the understanding of possible specific effects of IR on distinct brain regions (Neeper et al. 1996). In the hippocampus, the IR group had a significantly lower concentration of BDNF when compared to the CON group (IR: 198.23 ± 13.11 pg mL−1 vs. CON: 231.53 ± 36.04 pg mL−1; p < 0.05) (Fig. 4a). Otherwise, in the prefrontal cortex, the IR group had a significantly higher concentration of BDNF when compared to the CON group (IR: 265.73 ± 18.83 pg mL−1 vs. CON: 124.45 ± 17.57 pg mL−1; p < 0.001) (Fig. 4b). Therefore, it was possible to observe that IR does not impact the BDNF concentration in different brain areas in the same way.

BDNF concentration. a Hippocampus; b Prefrontal cortex. Mean ± SD. # Significant difference vs. CON (p < 0.05); ### significant difference vs. CON (p < 0.001). CON (n = 7); IR (n = 8)
Experiment 1: Inflammatory profile
Cytokines were analyzed in the hippocampus and prefrontal cortex areas. No statistical difference was observed between the groups in the concentration of cytokines in the hippocampus. However, the IR group had significantly higher concentrations of IL-10 and TNF-α in the prefrontal cortex when compared to the CON group (p < 0.01; p < 0.05, respectively) (Table 2).
Experiment 2: Long-term memory
The LTM was analyzed pre and post-60 days of the diet. Before evaluation, the groups underwent the second experiment, performing their respective experimental sessions (acute resistance exercise or rest). The recognition index decreased in the IR.REST group when comparing the pre- and post-moments (pre: 0.70 ± 0.08 vs. post: 0.55 ± 0.10; p < 0.01). However, the same was not observed in the IR.RE group (pre: 0.72 ± 0.05 vs. post: 0.65 ± 0.11; p = 0.095) (Fig. 5). In this way, the exercised group appeared to have their LTM restored, whereas the REST group had a decrease in their LTM.

Long-term memory pre and post-intervention. Mean ± SD. ** Significant difference vs. pre (p < 0.01); IR.REST (n = 8); IR.RE (n = 8)
Experiment 2: Brain-derived neurotrophic factor (BDNF)
The IR.RE group had a significantly higher concentration of BDNF in the hippocampus when compared to the IR.REST group (IR.RE: 214.71 ± 9.64 vs. IR.REST: 196.70 ± 14.49 pg mL−1; p < 0.05) (Fig. 6a), but not in the prefrontal cortex (p = 0.616) (Fig. 6b). Therefore, it was possible to observe that an acute bout of resistance exercise increased the concentration of BDNF in the hippocampus.

BDNF concentration. a Hippocampus; b Prefrontal cortex. Mean ± SD. # Significant difference vs. IR.REST (p < 0.05). IR.REST (n = 8); IR.RE (n = 8)
Experiment 2: Inflammatory profile
In the prefrontal cortex, no significant difference was observed between groups, however, the IR.RE group had a significantly lower concentration of IL-6 when compared to the IR.REST group (p < 0.001) (Table 3).
Experiment 2: Correlation between hippocampal BDNF and long-term memory (LTM)
Was observed a significant correlation (p = 0.040; r = 0.517) between BDNF in the hippocampus and LTM recognition index in insulin-resistant rats, demonstrating that the increase in BDNF concentration in the hippocampus, promoted by acute resistance exercise, can influence the degree of memory restoration (Fig. 7).

Correlation between BDNF concentration and performance in the LTM-NOR test after experimental situation
Discussion
Insulin resistance (IR) impaired long-term memory (LTM) and reduced BDNF concentration in the hippocampus. However, it increased BDNF and TNF-α concentrations in the prefrontal cortex of the studied rats. On the other hand, when subjected to an acute resistance exercise session, the IR rats had their long-term memory restored, showed increased BDNF concentration and reduced IL-6 concentration in the hippocampus compared to those who did not exercise.
Our results showed that insulin resistance did not affect the short-term memory of the IR group, however, it caused damage to long-term memory. This same response was observed in previous studies (Fu et al. 2017; Park et al. 2019). Neuroinflammation and transcriptional dysregulation (via CREB and BDNF) are two substantial factors underlying cognitive decline in an insulin-resistant scenery (Sharma 2021). The present study analyzed pro- and anti-inflammatory cytokines and BDNF concentration in the hippocampus and prefrontal cortex.
The IR group had a higher concentration of TNF-α in the prefrontal cortex. However, this did not reverberate in a higher proinflammatory status, inferred by the TNF-α/IL-10 ratio, in this area. Furthermore, the concentration of proinflammatory cytokines was not increased in the hippocampus. This finding differs from other studies that report neuroinflammation in the cortex (Liu et al. 2014; Cavaliere et al. 2019) and hippocampus (Jeon et al. 2012; Boitard et al. 2014; Yu et al. 2019) of insulin-resistant rodents. The studies mentioned above carried out the analysis of the concentrations of these cytokines after at least 14 weeks of consumption of diet for induction of IR/T2DM (Jeon et al. 2012; Boitard et al. 2014; Liu et al. 2014; Cavaliere et al. 2019; Yu et al. 2019). The present study analyzed cytokines after 9 weeks of the diet. This difference in diet consumption time may have influenced the condition of the inflammatory profile observed at the time of euthanasia.
In the present study, we observed a reduction in the BDNF hippocampus concentration of the IR group. Previous studies have noted the reduction of BDNF in the cortex (Liu et al. 2014; Cavaliere et al. 2019) and hippocampus (Park et al. 2010; Liu et al. 2014) of IR rodents. The hippocampus is a crucial area for memory consolidation, and BDNF plays a vital role in neuroplasticity (Egan et al. 2003; Sharma 2021), learning, and memory (Leal et al. 2015). Thus, the reduction in hippocampal BDNF concentrations can potentially explain the reduction in the LTM observed in these animals. Otherwise, in the prefrontal cortex, the observed results were not expected. The IR group showed increased BDNF concentration in the prefrontal cortex. Shin et al. (2015) observed high levels of BDNF in blood samples from individuals with Mild Cognitive Impairment (MCI), while BDNF concentrations were low in cases of severe Alzheimer’s Disease. Like this finding, the high BDNF concentration observed in the IR group’s prefrontal cortex may be due to a compensatory mechanism to counteract early neurodegeneration and/or to activate immune cells (Faria et al. 2014; Shin et al. 2015).
Both aerobic exercise and resistance exercise are recommended by the American Diabetes Association (ADA) for people living with diabetes due to their distinguished benefits (Elsayed et al. 2023b). Several studies have investigated the effects of a few weeks or months of aerobic exercise in animal models of IR/DM2 and have observed that, chronically, it was able to improve memory (Park et al. 2019; Mehta et al. 2019; Shekarchian et al. 2023), reduce proinflammatory cytokines concentration in the hippocampus (Mehta et al. 2019; Shekarchian et al. 2023) and increase BDNF concentration in the hippocampus (Park et al. 2019; Shekarchian et al. 2023) and prefrontal cortex (Shekarchian et al. 2023). De Sousa et al. (2020) showed that resistance training elicits cognitive maintenance in T2DM, whereas the non-exercise group showed a prominent decline in cognitive function. However, these studies assessed the chronic effects (i.e., multiple exercise sessions), making the acute effects (i.e., the effect of a single exercise session) on cognitive outcomes and mechanisms underlying this cognitive improvement remain not fully elucidated.
Unprecedentedly, we observed that an acute bout of RE led to an improvement in the LTM in IR animals. This result corroborates the findings of previous studies that investigated healthy individuals (Chang and Etnier 2009), T2D subjects (Silveira-Rodrigues et al. 2023), and subjects with MCI (Tsai et al. 2018). However, despite observing cognitive improvements after a RE session, Tsai et al. (2018) and Silveira-Rodrigues et al. (2023) found no change in the serum concentration of BDNF in these subjects. It should be noted that in our study, unlike those cited, the concentration of BDNF was evaluated in the animals’ brains (hippocampus and prefrontal cortex).
In addition to improving LTM, an acute bout of RE promoted increased BDNF concentration in the hippocampus. The molecular mechanisms responsible for exercise-induced BDNF elevation in the hippocampus of insulin-resistant rats are still under investigation. However, recent evidence suggests three primary mechanisms leading to exercise-induced brain BDNF production: enhanced neuronal activity, the elevation of cerebral blood flow, and the exerkine signaling emanating from peripheral tissues (Cefis et al. 2023).
Physical exercise induces an increase in neuronal activity in the hippocampus facilitated by: (1) the elevation of Ca2+ influx (Balkowiec and Katz 2002), (2) activation of glutamatergic transmission mediated by α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) and by N-methyl-D-aspartate-type (NMDA) receptors (Nishijima et al. 2012), (3) an increase in dopaminergic, noradrenergic and serotoninergic neurotransmitters (Cefis et al. 2023), and (4) activation of the signaling pathway PGC-1α/FNDC5/irisin leading to the upregulation of BDNF gene expression and memory improvement (Choi et al. 2018). Moreover, the increase in insulin-like growth factor 1 (IGF-1) synthesized from the liver and blood lactate produced by the skeletal muscle during physical exercise have the potential to cross the blood–brain barrier, modulating brain BDNF levels and promoting synaptic plasticity, neurogenesis, and cognitive function improvement (Ding et al. 2006; Müller et al. 2020).
The hippocampus has been identified as a major area of BDNF expression and preliminary studies indicate that the brain is the primary source, contributing about 80% of circulating plasma BDNF in response to exercise (Rasmussen et al. 2009), making measurements of acute exercise-induced changes in BDNF concentration relatively accessible. However, the acute effects of resistance exercise performed at different intensities and durations on BDNF concentration in the hippocampus of insulin-resistant rats have not been elucidated.
In trained adult male subjects, a single strength-training session at 80% of 1RM increased circulating BDNF levels 1 h after exercise, while no significant changes were observed in plasma BDNF after a training session at 60% of 1RM (Borges Junior et al. 2023). Conversely, a session of resistance training at 65–70% of 1RM was effective in elevating BDNF levels in older men (Arazi et al. 2021). In addition to intensity, the volume of resistance exercise can also influence BDNF concentration. In experienced resistance-trained men, an increase in BDNF was observed after both an acute bout of resistance training performed at high-intensity and a high-volume training session (Church et al. 2016). However, when the total load lifted was equalized between training sessions, a similar acute neurotrophic response was observed (Lodo et al. 2020).
Hippocampal BDNF increase has been associated with cognitive performance, specifically with memory improvement (Piepmeier and Etnier 2015). Experimental studies have demonstrated that animals deprived of BDNF in the hippocampus experience declines in cognitive performance compared to control animals (Mu et al. 1999). Conversely, the intrahippocampal administration of BDNF increases spatial memory and affects emotional behavior (Cirulli et al. 2004).
Still talking about the analyses done in the brain, we noticed a reduction in IL-6 concentration in the hippocampus after an acute bout of RE. Contrary to this result, Rasmussen et al. (2011) observed that there was an increase in IL-6 release in the human brain during and after recovery from prolonged aerobic exercise. In addition, the authors also found that mice showed an increase in IL-6 mRNA in the hippocampus 2 h after an acute bout of aerobic exercise, targeting this area as a likely source of IL-6 release in the brain. Additionally to the different type of exercise investigated (resistance exercise), in the present study we analyzed IL-6 concentrations in the hippocampus 30 min after the acute RE session. These factors may have influenced the difference in results observed between our experiment and Rasmussen et al. (2011) experiment.
In this study, we chose to investigate the effects of IR and an acute bout of RE on memory and neural outcomes. Insulin resistance typically precedes the onset of T2DM by several years (Fonseca 2009), and represents a risk factor for dementia, memory loss, and neurodegenerative diseases (Nguyen et al. 2020). Brain insulin resistance promotes neural apoptosis, brain ischemia, affects hippocampal plasticity, modulates glutamatergic neurotransmission, reduces hippocampal BDNF expression, and contributes to brain inflammatory reactions, ultimately leading to the development of cognitive impairment and memory loss (Lina et al. 2015). Thus, resistance exercise plays a crucial role in mitigating cognitive dysfunction associated with insulin resistance, thereby preventing the progression of type 2 diabetes-associated cognitive impairment (Malin et al. 2022). In the present study, we observed an increase in hippocampal BDNF after a resistance exercise session and a significant correlation between BDNF and long-term memory (LTM) recognition index in insulin-resistant rats.
The effectiveness of resistance training in improving glycemic control in type 2 diabetes became more debated in the early 2000s (Sigal et al. 2004). Currently, the ADA recommends that resistance exercise should be done two to three times a week (Elsayed et al. 2023b). Most long-term health benefits induced by physical exercise are thought to arise from adaptive changes to the acute modifications of every single bout of exercise, such as the increase in the activity and/or quantity of proteins involved in specific metabolic, physiological, and biomechanical processes (e.g., mitochondrial respiratory function, calcium cycling, contractile function/efficiency, and fuel use) (Neufer et al. 2015). Considering that T2DM is related to long-term cognitive impairment development, understanding the effects of an acute bout of resistance exercise on cognitive function is of great clinical application for recommending strength training programs for individuals living with T2DM. Future studies should investigate other forms of strength exercise prescription (volume and intensity) and their metabolic and cognitive effects on this population.
Conclusion
Insulin resistance leads to impaired long-term memory and reduced BDNF concentration in the hippocampus of rats. Furthermore, acute resistance exercise can restore long-term memory decrement and increase BDNF concentration in the hippocampus of insulin-resistant rats.
Data availability
Data are available online: (https://data.mendeley.com/drafts/7mntzk4hpw).
References
Arazi H, Babaei P, Moghimi M, Asadi A (2021) Acute effects of strength and endurance exercise on serum BDNF and IGF-1 levels in older men. BMC Geriatr 21:1–8. https://doi.org/10.1186/s12877-020-01937-6
Balkowiec A, Katz DM (2002) Cellular mechanisms regulating activity-dependent release of native brain-derived neurotrophic factor from hippocampal neurons. J Neurosci 22:10399–10407. https://doi.org/10.1523/jneurosci.22-23-10399.2002
Bevins RA, Besheer J (2006) Object recognition in rats and mice: a one-trial non-matching-to-sample learning task to study “recognition memory.” Nat Protoc 1:1306–1311. https://doi.org/10.1038/nprot.2006.205
Boitard C, Cavaroc A, Sauvant J et al (2014) Impairment of hippocampal-dependent memory induced by juvenile high-fat diet intake is associated with enhanced hippocampal inflammation in rats. Brain Behav Immun 40:9–17. https://doi.org/10.1016/j.bbi.2014.03.005
Borges Junior M, Tavares LFJ, Nagata GY et al (2023) Impact of strength training intensity on brain-derived neurotrophic factor. Int J Sports Med. https://doi.org/10.1055/a-2197-1201
Capettini SB, Moraes MFD, Prado VF et al (2011) Vesicular acetylcholine transporter knock-down mice show sexual dimorphism on memory. Brain Res Bull 85:54–57. https://doi.org/10.1016/j.brainresbull.2011.02.005
Cassilhas RC, Lee KS, Fernandes J et al (2012a) Spatial memory is improved by aerobic and resistance exercise through divergent molecular mechanisms. Neuroscience 202:309–317. https://doi.org/10.1016/j.neuroscience.2011.11.029
Cassilhas RC, Lee KS, Venâncio DP et al (2012b) Resistance exercise improves hippocampus-dependent memory. Braz J Med Biol Res 45:1215–1220. https://doi.org/10.1590/S0100-879X2012007500138
Cavaliere G, Trinchese G, Penna E et al (2019) High-fat diet induces neuroinflammation and mitochondrial impairment in mice cerebral cortex and synaptic fraction. Front Cell Neurosci 13:1–13. https://doi.org/10.3389/fncel.2019.00509
Cefis M, Chaney R, Wirtz J et al (2023) Molecular mechanisms underlying physical exercise-induced brain BDNF overproduction. Front Mol Neurosci 16:1–18. https://doi.org/10.3389/fnmol.2023.1275924
Chang YK, Etnier JL (2009) Exploring the dose-response relationship between resistance exercise intensity and cognitive function. J Sport Exerc Psychol 31:640–656. https://doi.org/10.1123/jsep.31.5.640
Chatterjee S, Khunti K, Davies MJ (2017) Type 2 diabetes. Lancet 389:2239–2251. https://doi.org/10.1016/S0140-6736(17)30058-2
Chen L, Chen R, Wang H, Liang F (2015) Mechanisms linking inflammation to insulin resistance. Int J Endocrinol 2015:1–9. https://doi.org/10.1155/2015/508409
Choi SH, Bylykbashi E, Chatila ZK et al (2018) Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer’s mouse model. Science 361(6406):eaan8821. https://doi.org/10.1126/science.aan8821
Church DD, Hoffman JR, Mangine GT et al (2016) Comparison of high-intensity vs. high-volume resistance training on the BDNF response to exercise. J Appl Physiol 121:123–128. https://doi.org/10.1152/japplphysiol.00233.2016
Cirulli F, Berry A, Chiarotti F, Alleva E (2004) Intrahippocampal administration of BDNF in adult rats affects short-term behavioral plasticity in the Morris water maze and performance in the elevated plus-maze. Hippocampus 14:802–807. https://doi.org/10.1002/hipo.10220
Craft S (2007) Insulin resistance and Alzheimer’s disease pathogenesis: potential mechanisms and implications for treatment. Curr Alzheimer Res 4:147–152. https://doi.org/10.2174/156720507780362137
Crunfli F, Mazucanti CH, De Moraes RCM et al (2018) NO-dependent Akt inactivation by S-nitrosylation as a possible mechanism of STZ-induced neuronal insulin resistance. J Alzheimers Dis 65:1427–1443. https://doi.org/10.3233/JAD-180284
Crunfli F, Vrechi TA, Costa AP, Torrão AS (2019) Cannabinoid receptor type 1 agonist ACEA improves cognitive deficit on STZ-induced neurotoxicity through apoptosis pathway and NO modulation. Neurotox Res 35:516–529. https://doi.org/10.1007/s12640-018-9991-2
De Sousa RAL, Improta-Caria AC, de Jesus-Silva FM et al (2020) High-intensity resistance training induces changes in cognitive function, but not in locomotor activity or anxious behavior in rats induced to type 2 diabetes. Physiol Behav 223:1–7. https://doi.org/10.1016/j.physbeh.2020.112998
DeFronzo RA, Ferrannini E, Groop L et al (2015) Type 2 diabetes mellitus. Nat Rev Dis Prim 1:1–23. https://doi.org/10.1038/nrdp.2015.19
Dinel AL, André C, Aubert A et al (2011) Cognitive and emotional alterations are related to hippocampal inflammation in a mouse model of metabolic syndrome. PLoS ONE 6:1–10. https://doi.org/10.1371/journal.pone.0024325
Ding Q, Vaynman S, Akhavan M et al (2006) Insulin-like growth factor I interfaces with brain-derived neurotrophic factor-mediated synaptic plasticity to modulate aspects of exercise-induced cognitive function. Neuroscience 140:823–833. https://doi.org/10.1016/j.neuroscience.2006.02.084
Egan MF, Kojima M, Callicott JH et al (2003) The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 112:257–269. https://doi.org/10.1016/S0092-8674(03)00035-7
Elsayed NA, Aleppo G, Aroda VR et al (2023a) 2. Classification and diagnosis of diabetes: standards of care in diabetes—2023. Diabetes Care 46:S19–S40. https://doi.org/10.2337/dc23-S002
Elsayed NA, Aleppo G, Aroda VR et al (2023b) 5. Facilitating positive health behaviors and well-being to improve health outcomes: standards of care in diabetes—2023. Diabetes Care 46:S68–S96. https://doi.org/10.2337/dc23-S005
Faria MC, Gonçalves GS, Rocha NP et al (2014) Increased plasma levels of BDNF and inflammatory markers in Alzheimer’s disease. J Psychiatr Res 53:166–172. https://doi.org/10.1016/j.jpsychires.2014.01.019
Fonseca VA (2009) Defining and characterizing the progression of type 2 diabetes. Diabetes Care 32(Suppl 2):S151–S156. https://doi.org/10.2337/dc09-s301
Fonseca CS, Gusmão ID, Raslan ACS et al (2013) Object recognition memory and temporal lobe activation after delayed estrogen replacement therapy. Neurobiol Learn Mem 101:19–25. https://doi.org/10.1016/j.nlm.2012.12.016
Fu Z, Wu J, Nesil T et al (2017) Long-term high-fat diet induces hippocampal microvascular insulin resistance and cognitive dysfunction. Am J Physiol Endocrinol Metab 312:E89–E97. https://doi.org/10.1152/ajpendo.00297.2016
Giacco et al (2011) Oxidative stress and diabetic complications Ferdinando NIH Public Access. NIH Public Access Author Manuscr 107:1058–1070. https://doi.org/10.1161/CIRCRESAHA.110.223545.Oxidative
Jeon BT, Jeong EA, Shin HJ et al (2012) Resveratrol attenuates obesity-associated peripheral and central inflammation and improves memory deficit in mice fed a high-fat diet. Diabetes 61:1444–1454. https://doi.org/10.2337/db11-1498
Lacerda DR, Serakides R, Ocarino NM et al (2015) Osteopetrosis in obese female rats is site-specifically inhibited by physical training. Exp Physiol 100:44–56. https://doi.org/10.1113/expphysiol.2014.082511
Leal G, Afonso PM, Salazar IL, Duarte CB (2015) Regulation of hippocampal synaptic plasticity by BDNF. Brain Res 1621:82–101. https://doi.org/10.1016/j.brainres.2014.10.019
Leenders M, Verdijk LB, van der Hoeven L et al (2013) Patients with type 2 diabetes show a greater decline in muscle mass, muscle strength, and functional capacity with aging. J Am Med Dir Assoc 14:585–592. https://doi.org/10.1016/j.jamda.2013.02.006
Lina M, Jieyu W, Yun L (2015) Insulin resistance and cognitive dysfunction. Clinica Chimica Acta. https://doi.org/10.1016/j.cca.2015.01.027
Lindqvist A, Mohapel P, Bouter B et al (2006) High-fat diet impairs hippocampal neurogenesis in male rats. Eur J Neurol 13:1385–1388. https://doi.org/10.1111/j.1468-1331.2006.01500.x
Liu Y, Fu X, Lan N et al (2014) Luteolin protects against high fat diet-induced cognitive deficits in obesity mice. Behav Brain Res 267:178–188. https://doi.org/10.1016/j.bbr.2014.02.040
Lodo L, Moreira A, Bacurau RFP et al (2020) Resistance exercise intensity does not influence neurotrophic factors response in equated volume schemes. J Hum Kinet 74:227–236. https://doi.org/10.2478/hukin-2020-0030
Malin SK, Stewart NR, Ude AA, Alderman BL (2022) Brain insulin resistance and cognitive function: influence of exercise. J Appl Physiol 133:1368–1380. https://doi.org/10.1152/japplphysiol.00375.2022
Mehta BK, Singh KK, Banerjee S (2019) Effect of exercise on type 2 diabetes-associated cognitive impairment in rats. Int J Neurosci 129:252–263. https://doi.org/10.1080/00207454.2018.1526795
Melo BP, Zacarias AC, Oliveira JCC et al (2021) Thirty days of combined consumption of a high-fat diet and fructose-rich beverages promotes insulin resistance and modulates inflammatory response and histomorphometry parameters of liver, pancreas, and adipose tissue in Wistar rats. Nutrition 91–92:1–8. https://doi.org/10.1016/j.nut.2021.111403
Mozaffarian D, Kamineni A, Carnethon M, Al E (2009) Lifestyle risk factors and new-onset DM in older adults. Arch Intern Med 169:798–807. https://doi.org/10.1001/archinternmed.2009.21.Lifestyle
Mu JS, Li WP, Bin YZ, Zhou XF (1999) Deprivation of endogenous brain-derived neurotrophic factor results in impairment of spatial learning and memory in adult rats. Brain Res 835:259–265. https://doi.org/10.1016/S0006-8993(99)01592-9
Müller P, Duderstadt Y, Lessmann V, Müller NG (2020) Lactate and BDNF: key mediators of exercise induced neuroplasticity? J Clin Med 9:1–14. https://doi.org/10.3390/jcm9041136
Neeper SA, Gómez-Pinilla F, Choi J, Cotman CW (1996) Physical activity increases mRNA for brain-derived neurotrophic factor and nerve growth factor in rat brain. Brain Res 726:49–56. https://doi.org/10.1016/0006-8993(96)00273-9
Neufer PD, Bamman MM, Muoio DM et al (2015) Understanding the cellular and molecular mechanisms of physical activity-induced health benefits. Cell Metab 22:4–11. https://doi.org/10.1016/j.cmet.2015.05.011
Neves RVP, Souza MK, Passos CS et al (2016) Resistance training in spontaneously hypertensive rats with severe hypertension. Arq Bras Cardiol 106:201–209. https://doi.org/10.5935/abc.20160019
Nguyen TT, Ta QTH, Nguyen TTD et al (2020) Role of insulin resistance in the Alzheimer’s disease progression. Neurochem Res 45:1481–1491. https://doi.org/10.1007/s11064-020-03031-0
Nishijima T, Okamoto M, Matsui T et al (2012) Hippocampal functional hyperemia mediated by NMDA receptor/NO signaling in rats during mild exercise. J Appl Physiol 112:197–203. https://doi.org/10.1152/japplphysiol.00763.2011
Padilha CS, Cella PS, Ribeiro AS et al (2019) Moderate vs. high-load resistance training on muscular adaptations in rats. Life Sci 238:116964. https://doi.org/10.1016/j.lfs.2019.116964
Park HR, Park M, Choi J et al (2010) A high-fat diet impairs neurogenesis: involvement of lipid peroxidation and brain-derived neurotrophic factor. Neurosci Lett 482:235–239. https://doi.org/10.1016/j.neulet.2010.07.046
Park HS, Park SS, Kim CJ et al (2019) Exercise alleviates cognitive functions by enhancing hippocampal insulin signaling and neuroplasticity in high-fat diet-induced obesity. Nutrients 11:1–13. https://doi.org/10.3390/nu11071603
Pereira GB, Prestes J, Leite RD et al (2010) Effects of ovariectomy and resistance training on MMP-2 activity in rat calcaneal tendon. Connect Tissue Res 51:459–466. https://doi.org/10.3109/03008201003676330
Pesta DH, Goncalves RLS, Madiraju AK et al (2017) Resistance training to improve type 2 diabetes: working toward a prescription for the future. Nutr Metab 14:1–10. https://doi.org/10.1186/s12986-017-0173-7
Piepmeier AT, Etnier JL (2015) Brain-derived neurotrophic factor (BDNF) as a potential mechanism of the effects of acute exercise on cognitive performance. J Sport Heal Sci 4:14–23. https://doi.org/10.1016/j.jshs.2014.11.001
Piroli GG, Grillo CA, Reznikov LR et al (2007) Corticosterone impairs insulin-stimulated translocation of GLUT4 in the rat hippocampus. Neuroendocrinology 85:71–80. https://doi.org/10.1159/000101694
Pratchayasakul W, Kerdphoo S, Petsophonsakul P et al (2011) Effects of high-fat diet on insulin receptor function in rat hippocampus and the level of neuronal corticosterone. Life Sci 88:619–627. https://doi.org/10.1016/j.lfs.2011.02.003
Puig KL, Floden AM, Adhikari R et al (2012) Amyloid precursor protein and proinflammatory changes are regulated in brain and adipose tissue in a murine model of high fat diet-induced obesity. PLoS ONE 7:1–16. https://doi.org/10.1371/journal.pone.0030378
Rasmussen P, Brassard P, Adser H et al (2009) Evidence for a release of brain-derived neurotrophic factor from the brain during exercise. Exp Physiol 94:1062–1069. https://doi.org/10.1113/expphysiol.2009.048512
Rasmussen P, Vedel JC, Olesen J et al (2011) In humans IL-6 is released from the brain during and after exercise and paralleled by enhanced IL-6 mRNA expression in the hippocampus of mice. Acta Physiol 201:475–482. https://doi.org/10.1111/j.1748-1716.2010.02223.x
Richter EA, Hargreaves M (2013) Exercise, GLUT4, and skeletal muscle glucose uptake. Physiol Rev 93:993–1017. https://doi.org/10.1152/physrev.00038.2012
Robertson RP, Harmon J, Tran PO et al (2003) Glucose toxicity in β-Cells: type 2 Diabetes, Good Radicals Gone Bad, and the Glutathione Connection. Diabetes 52:581–587. https://doi.org/10.2337/diabetes.52.3.581
Rojas JJ, Deniz BF, Miguel PM et al (2013) Effects of daily environmental enrichment on behavior and dendritic spine density in hippocampus following neonatal hypoxia-ischemia in the rat. Exp Neurol 241:25–33. https://doi.org/10.1016/j.expneurol.2012.11.026
Sharma S (2021) High fat diet and its effects on cognitive health: alterations of neuronal and vascular components of brain. Physiol Behav 240:113528. https://doi.org/10.1016/J.PHYSBEH.2021.113528
Shekarchian M, Peeri M, Azarbayjani MA (2023) Physical activity in a swimming pool attenuates memory impairment by reducing glutamate and inflammatory cytokines and increasing BDNF in the brain of mice with type 2 diabetes. Brain Res Bull 201:1–14. https://doi.org/10.1016/j.brainresbull.2023.110725
Shin HW, Kim H, Lee KJ (2015) Differences in BDNF serum levels in patients with Alzheimer’s disease and mild cognitive impairment. Afr J Psychiatry 18:1–5. https://doi.org/10.4172/Psychiatry.1000245
Sigal RJ, Wasserman DH, Kenny GP, Castaneda-Sceppa C (2004) Physical activity/exercise and type 2 diabetes. Diabetes Care 10:2518–2539. https://doi.org/10.2337/diacare.27.10.2518
Silveira-Rodrigues JG, Campos BT, de Lima AT et al (2023) Acute bouts of aerobic and resistance exercise similarly alter inhibitory control and response time while inversely modifying plasma BDNF concentrations in middle-aged and older adults with type 2 diabetes. Exp Brain Res. https://doi.org/10.1007/s00221-023-06588-8
Sylow L, Kleinert M, Richter EA, Jensen TE (2017) Exercise-stimulated glucose uptake-regulation and implications for glycaemic control. Nat Rev Endocrinol 13:133–148. https://doi.org/10.1038/nrendo.2016.162
Tsai CL, Ukropec J, Ukropcová B, Pai MC (2018) An acute bout of aerobic or strength exercise specifically modifies circulating exerkine levels and neurocognitive functions in elderly individuals with mild cognitive impairment. NeuroImage Clin 17:272–284. https://doi.org/10.1016/j.nicl.2017.10.028
Winocur G, Greenwood CE, Piroli GG et al (2005) Memory impairment in Obese Zucker rats: an investigation of cognitive function in an animal model of insulin resistance and obesity. Behav Neurosci 119:1389–1395. https://doi.org/10.1037/0735-7044.119.5.1389
Yu M, Huang H, Dong S et al (2019) High mobility group box-1 mediates hippocampal inflammation and contributes to cognitive deficits in high-fat high-fructose diet-induced obese rats. Brain Behav Immun 82:167–177. https://doi.org/10.1016/j.bbi.2019.08.007
Acknowledgements
The authors are thankful to the funding support agencies, by the Minas Gerais research support foundation (Fundacao de Amparo a Pesquisa de Minas Gerais, FAPEMIG). This study was also financed by the Coordenacao de Aperfeicoamento de Pessoal de Nivel Superior—Brazil (CAPES)—financial code 001 and CNPq - Brasil: Conselho Nacional de Desenvolvimento Científico e Tecnológico (financial code 310014/2021-2). The authors are also thankful for the assistance of Laboratorio de Fisiologia do Exercicio/UFMG members in data collection.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Sreedharan Sajikumar.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Berbert-Gomes, C., Ramos, J.S., Silveira-Rodrigues, J.G. et al. An acute bout of resistance exercise increases BDNF in hippocampus and restores the long-term memory of insulin-resistant rats. Exp Brain Res 242, 901–912 (2024). https://doi.org/10.1007/s00221-024-06795-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-024-06795-x