
Abstract
Past studies have revealed connections in directional programming between hands. The present study investigated whether there could also be interaction in programming proximal and distal components of a prehensile hand action. In Experiment 1, the participants performed simultaneously either a push or pull response with the left hand and the grip closure or opening with the right hand. In Experiment 2, the push and pull responses of the left hand were performed together with the precision or power grip responses of the right hand. The participants showed preference, measured in reaction times, to couple the push response with the grip opening and the precision grip, whereas the pull response was associated with the grip closure and the power grip. The study shows for the first time a systematic interaction in proximal and distal prehensile components between two hands. We propose that these effects reflect inter- and intra-limb connections between the representations that prepare the arm extension for the outward reaching, the finger extension for the grip opening, and the motor processes that prepare the precision grip. Conversely, there appear to be connections between the representations that prepare the arm flexion for the inward directed hand movements, the flexion of the thumb and the fingers for the grip closure, and flexion of four fingers for the power grip.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The present study investigates whether there exists a systematic coupling between proximal hand actions of the left hand and distal hand actions of the right hand. Regarding inter-limb coupling, previous research has revealed connections in directional programming between hands. There appears to be an interaction between homologous muscle groups of two hands; it is easier to perform symmetrical movements of two hands utilizing homologous muscle groups than asymmetrical movements that utilize non-homologous muscle groups (e.g., Swinnen et al. 1997; Serrien et al. 1999). The phenomena of mirror movements and contralateral motor irradiation have shown that corresponding effects can be also observed in the increased activity of homologous muscles of the resting hand during voluntary actions of the opposite hand (Cernacek 1961; Armatas et al. 1994).
In general, regarding intra-limb action programming, each neuron in the motor system participates in several hand actions (Schieber and Hibbard 1993). In addition, research has revealed an overlap in activation of contralateral motor representations of distal (thumb, fingers and wrist) and proximal (elbow and shoulder) forelimb movements (e.g., Stark et al. 2007). In line with this evidence, although reaching and grasping are mostly interdependent and programmed in parallel (Jeannerod 1984; Vesia et al. 2010, 2013) there also appears to be interaction in programming reach (i.e., proximal) and grasp (i.e., distal) components of a prehensile action (e.g., Dum and Strick 2005; Raos et al. 2004; Stark et al. 2007; Haggard and Wing 1995).
The grasp component of a prehensile action is typically accomplished by the precision or power-type of grip (Napier 1956). Moreover, grasping includes grip opening and closure phases (Jeannerod 1984). In addition, it could be assumed that some characteristics of reach-to-grasp actions are programmed in tighter interaction than some other characteristics. For example, given that outward directed reach-to-grasp is strongly coupled with grip opening, the maximal finger opening occurring within 60–70% of the duration of the reach (Jeannerod 1984), it could be assumed that representations that are commanding forward rather than backward hand movements are tighter integrated with the motor processes that command grip opening.
Intermanual interactions have been previously studied using a variety of behavioral methods such as a periodic-movement paradigm (e.g., Swinnen et al. 2001) and a reaction-time paradigm (Diedrichsen et al. 2001). The present study uses the bimanual choice reaction time (RT) task (i.e., the dual-action task) to investigate, for the first time, whether there could be some systematic preference to couple different types of proximal hand movements related to prehensile actions (i.e., reach forwards vs. reach backwards) with the specific grip phase (opening vs. closure: Experiment 1) and/or the specific grip type (precision vs. power: Experiment 2). The basic principle behind the present study is based on views on how mechanisms of separate hands are coupled in action planning (Rosenbaum 1980). According to the view, when the task requires choosing between two alternative responses for both hands according to a single stimulus, the action plans of two alternative responses are prepared in parallel for both hands prior to the onset of the stimulus. When then the stimulus is presented and the choice task must be carried out simultaneously for the responses of both hands, there occurs competition between alternative response options. Consequently, the time to select the required motor program for one action is affected by the response selection processes related to the other action. The more the two responses required in each trial share motor characteristics, the less there is interference between selection processes related to two actions, and consequently, the faster the actions are executed.
We predict that some characteristics of reach-to-grasp actions such as the arm and finger extension are programmed in tighter interaction than some other characteristics such as the arm extension and the finger flexion. Moreover, we assume that similarly to joint directional programming between hands (Swinnen et al. 1997; Serrien et al. 1999; Armatas et al. 1994), these common prehensile characteristics could be also programmed (to some extent) jointly for both hands. As such, according to the view presented above, we hypothesize that if there is partial overlap between processes that plan, for example, forward hand movements and grip opening, dual-action responses should be executed relatively rapidly when one hand is performing grip opening rather than grip closure while the other hand is performing the push movement.
In Experiment 1, the participants performed either the push or pull responses with their left hand using a joystick and simultaneously performed either the grip closure or opening with their right hand. Planning the push and pull actions with a joystick can be assumed to utilize the same directional codes as the forward and backward reach actions, respectively. In addition, it can be assumed that execution of the push and pull actions employ the same proximal extension–flexion motor representations as the forward and backward reach actions, respectively. Given that outward directed reach-to-grasp is commonly coupled with grip opening (Jeannerod 1984), we assumed that the participants might prefer to couple the push action of the left hand with the grip opening movement of the right hand. In contrast, the inward directed hand movements are relatively more often associated with moving the grasped object toward the body with fingers wrapped around the object (e.g., bringing an apple to the mouth). Consequently, we presumed that the participants might prefer to couple pull actions with the grip closure in this task.
In addition to grip opening and closure, grasp formation consists of a selection of the grip type; that is, preparing to grasp an object using either the precision or power grip. Consequently, if Experiment 1 explores how the proximal prehensile component (i.e., reaching inward vs. outward) is associated with the grip opening and closure processes, Experiment 2 explores how the proximal prehensile component is associated with performing the precision and power grip responses. In Experiment 2, participants performed either the push or pull responses with their left hand and simultaneously performed either the precision grip or power grip with their right hand. Due to the explorative nature of Experiment 2, we did not have specific hypotheses about how push and pull responses would be interacting with the precision and power grip.
The precision grip, most commonly used for reaching to grasp small objects, is accomplished by pressing an object between the tip of the index finger and the thumb. In contrast, the power grip is typically accomplished by flexing four fingers toward the palm base so that the thumb does not necessarily have an active role in the grip (see, for example, Stark et al. 2007). The power grip is mostly implemented by the flexor digitorum profundus (Kilbreath and Gandevia 1994; Reilly and Schieber 2001), which is responsible for flexing index, middle, ring and little finger toward the palm base. Entirely different flexor, namely the flexor pollicis longus, is responsible for flexing the thumb in the precision grip (Kilbreath and Gandevia 1994). Thus, it is possible that the grips would be differently related to the push and pull gestures. As an example, it would be possible that the motor processes that prepare the arm flexion of the left hand are more tightly integrated with the motor processes that prepare the flexion of the fingers for the power grip than the flexion of the thumb for the precision grip. This would be observed in coupling between the backward directed hand movement of the left hand and the power grip of the right hand.
Methods
Participants
In Experiment 1, nineteen naïve volunteers participated in the study (19–36 years of age; mean age = 24.4 years; 2 males). In Experiment 2, eighteen naïve volunteers participated in the study (22–31 years of age; mean age = 25.5 years; 5 males). These sample sizes were chosen based on our previous knowledge of effect sizes in these kinds of experiments (Vainio et al. 2015; Tiainen et al. 2017), and we estimated that with these sample sizes, we should reach power around 0.9. All participants were psychology students of University of Helsinki. The gender was not considered as a criterion for selecting participants because the preliminary analyses of our previous studies, in which the grip and joystick responses were similarly performed in dual-action tasks (Vainio et al. 2015; Tiainen et al. 2017) indicated that gender does not seem to play a significant role in these kinds of behavioural effects. Most of the participants were female because approximately 80% of the psychology students are female in University of Helsinki. All participants were right-handers and had normal or corrected-to-normal vision.
Compliance with ethical standards
The study was approved by the Ethical Committee of the Institute of Behavioural Sciences at the University of Helsinki and has therefore been performed in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki. Written informed consent was obtained from all participants.
Apparatus, stimuli and procedure
Each participant sat in a dimly lit room with his or her head 70 cm in front of a 19-in. CRT monitor (screen refresh rate: 85 Hz; screen resolution: 1280 × 1024). The target stimuli consisted of two horizontally oriented lines (length: 2°) from which one line was clearly thicker than the other line (thin: 0.2°; thick: 0.4°) (the double line stimulus). The thin line was either below or above the thick line (Fig. 1). In addition, both lines were presented either in green or blue color. All stimuli were displayed in the center of the monitor on a white background.

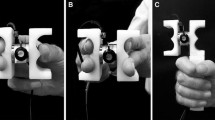
Graphical presentation of the trial structure of Experiments 1 and 2 (see text for details). In both experiments, the left-hand responses were performed with the joystick by moving it forward or backward. The right hand responses were performed with the scissors (Experiment 1) by the finger opening or closure gesture or with the grip device by the precision or power grip (Experiment 2). The scissors and the grip device were equipped with microswitches (their location is marked with the dotted circles in the picture) that registered the responses
In the trial structure (see Fig. 1), a fixation cross was presented for 400 ms at the beginning of each trial. Then, a blank screen was displayed for 400 ms. After that the target stimulus was presented 2000 ms or until a response was executed with both hands. Finally, a blank screen was displayed for 500 ms. Both experiments began with practice trials. Each participant was given as much practice as it took to perform the task fluently (i.e., fast and simultaneous left and right hand responses with a small number of errors).
In both experiments, the participants’ left-hand task was to pull or push the joystick until the end of the full motion range of the joystick (4.5 cm forward or backward) as fast and accurately as possible according to the spatial arrangement of the double line stimuli (see Fig. 1). Half of the participants responded according to the relative spatial location of the thin line (e.g., move the hand forwards if the thin line is above the thick line and backward if it is below the thick line) while half of the participants responded according to the relative location of the thick line. The joystick was always returned to the central starting position after the response. The joystick was attached steadily onto the table and located between the participant and the monitor, 30 cm away from the monitor. It was positioned horizontally 10 cm to the left of the center of the monitor.
In Experiment 1, the right hand responses were performed with the scissors (see Fig. 1) by opening or closing them at the same time with the left hand response. The scissors were equipped with two microswitches from which one registered the opening of the scissors and the other registered the closing of them. The scissors were opened and closed by the handles using index and middle finger in the one handle and the thumb in the other handle. The grip opening versus closure was performed according to the color of the double line stimulus (e.g., blue for the opening and green for the closing). The microswitches of the scissors were marked with color tapes that corresponded to the required color-movement mapping arrangements. After the opening/closure response was executed, the participants were required to return the scissors back to the half open position. Two scissor blades were attached by a screw (see Fig. 1), and twisting this screw enabled manipulating stiffness of the closing movement. When the screw was loose, there was no tactually observable resistance in the closing movement. The extent that the screw was tightened defined the extent of the resistance of the closing movement as well as at which point of the closing movement, from fully open to fully close, the resistance began. Stiffness of the scissors was adjusted so that they provided tactually observable resistance at the point from which there was approximately equal motion range (2 cm) toward the maximal open and the maximal close position of the scissors.
In Experiment 2, the right-hand responses were performed with the precision (1 × 1 × 0.7 cm) and power (10 cm long, 3 cm diameter) grip devices (see Fig. 1) by pressing one of the devices at the same time with the left hand response. The devices were equipped with an inlaid microswitch. As the switches were depressed in each device, there was noticeable tactile feedback. The participants were required to perform the precision or power grip response according to the color of the stimulus (e.g., blue for the precision grip and green for the power grip). The grip devices were marked with corresponding color tapes that corresponded to the required color-grip mapping arrangements. The grip task and the devices were adapted from the study reported by Tucker and Ellis (2001) and Vainio et al. (2008). Because both of the grip devices were held in the right hand, the contribution of the index finger—typically also involved in the power grip—was missing in the power grip task, which had to be executed using the little, ring and middle finger. Otherwise these grip actions were performed exactly in the way they are normally performed (see the descriptions in the Introduction).
The color-movement (Experiment 1) and color-grip (Experiment 2) mapping was balanced so that approximately half of the participants performed closing/precision action for the blue color, while rest of the participants performed closing/precision action for the green color, in both of the joystick response arrangements that determined push–pull responses according to the relative spatial location of the thin or thick line. In addition, in both experiments, the right hand was resting on the arm of an armchair while holding the devices.
Reaction times (RTs) of the joystick movements were measured from the onset of the target stimulus to the point of the joystick movement in which the joystick reached 100% from the motion range of the joystick to the forward or backward direction. The response was registered as an ‘error’ if joystick movement exceeded 100% of the motion range but was moved into the wrong direction. In addition, the response was registered as ‘no-response’ if the joystick movement did not reach 100% of the motion range. Regarding the close/open and precision/power responses, RTs were measured from the onset of the target stimulus to the point of the response in which the movement resulted in the pushing one of the microswitches of the device.
Statistical analysis
Experiment 1 consisted of 140 trials [35 (repetition) × 2 (left-hand: push/pull) × 2 (right-hand: close/open)]. Statistical significance of reaction time values was tested using a random intercept model (linear mixed models; see Barr et al. 2013 for a justification of using this statistical method with the current data) that treated joystick (push or pull), scissors (open or close) and mapping (between subjects) [MP1 (green/open or MP2 (green/close)] as fixed factors and subject as a random intercept. Post hoc comparisons were carried out using the Bonferroni correction. Regarding the joystick reaction times, the post hoc comparisons tested whether the joystick responses were modulated by the scissor responses; That is, whether one type of joystick response (e.g., push) was performed significantly faster/slower together with one type of scissor response (e.g., open) compared to other type of scissor response (e.g., close). Regarding the scissor reaction times, the post hoc comparisons tested whether the scissor responses were modulated by the joystick responses; That is, whether one type of scissor response (e.g., close) was performed significantly faster/slower together with one type of joystick response (e.g., pull) compared to other type of joystick response (e.g., push).
Experiment 2 consisted of 160 trials [40 (repetition) × 2 (left-hand: push/pull) × 2 (right-hand: precision/power)]. Statistical significance of reaction time values was tested using a random intercept model (linear mixed models) that treated joystick (push or pull), grip (precision or power) and mapping (between subjects) [MP1 (green/precision or MP2 (green/power)] as fixed factors and subject as a random intercept. Post hoc comparisons were carried out using the Bonferroni correction. Regarding the joystick reaction times, the post hoc comparisons tested whether the joystick responses were modulated by the grip responses; That is, whether one type of joystick response (e.g., push) was performed significantly faster/slower together with one type of grip response (e.g., precision) compared to other type of grip response (e.g., power). Regarding the grip reaction times, the post hoc comparisons tested whether the grip responses were modulated by the joystick responses; That is, whether one type of grip response (e.g., power) was performed significantly faster/slower together with one type of joystick response (e.g., pull) compared to other type of joystick response (e.g., push).
Results
Experiment 1
6.4% of the raw data containing errors were discarded from the analysis of reaction times of joystick and scissor responses. The discarded error data consisted of those responses that were incorrectly executed with the left hand (2.2%), right hand (3.8%) or with both hands (0.4%). In addition, regarding the left-hand (joystick) responses, 4.5% of trials in which the RTs were more than two standard deviations from a participant’s overall mean were discarded from the analysis. Regarding the right-hand (scissor) responses, 4.4% of trials in which the RTs were more than two standard deviations from a participant’s overall mean were additionally discarded from the analysis.
The errors were not analyzed for the joystick or scissor responses because the participants produced only few errors. The analysis of joystick responses revealed significant main effects of joystick [F(1,2359) = 6.09, p = .014] and scissor [F(1,2359) = 5.79, p = .016]. The backward responses were made faster [M = 930 ms, 95% CI (842, 1018)] than the forward responses [M = 953 ms, 95% CI (865, 1040)]. In addition, the close responses were made faster [M = 930 ms, 95% CI (843, 1018)] than the open responses [M = 952 ms, 95% CI (865, 1040)]. Joystick × scissor interaction was also obtained [F(1,2359) = 52.81, p < .001]. This interaction is presented in Fig. 2. The pairwise comparisons test showed that the push responses were performed significantly faster when the scissors were opened [M = 930 ms, 95% CI (842, 1019)] rather than closed [M = 975 ms, 95% CI (886, 1063)] (p = .001). In contrast, the pull responses were performed faster when the scissors were closed [M = 886 ms, 95% CI (798, 975)] rather than opened [M = 974 ms, 95% CI (886, 1063)] (p < .001). The interaction between joystick, scissor and mapping was not significant [F(1,2359) = 0.88, p = .348] suggesting that mapping (i.e., which color cued opening/closing responses) did not interfere with the interaction between the joystick and scissor responses.

The mean vocal reaction times for Experiment 1 as a function of the joystick (i.e., the left-hand) and the scissor (i.e., the right-hand) responses. Error bars depict the standard error of the mean. Asterisks indicate statistically significant differences (*** < . 001; **p < .01; *p < .05)
The analysis of scissor responses revealed significant main effects of scissor [F(1,2357) = 8.29, p = .004]. The close responses were made faster [M = 907 ms, 95% CI (812, 1002)] than the open responses [M = 934 ms, 95% CI (839, 1029)]. Joystick × scissor interaction was also obtained [F(1,2357) = 50.33, p < .001]. This interaction is presented in Fig. 2. The pairwise comparisons test showed that the open responses were performed faster when the joystick was pushed [M = 906 ms, 95% CI (810, 1001)] rather than pulled [M = 962 ms, 95% CI (866, 1058)] (p < .001). In contrast, the close responses were performed faster when the joystick was pulled [M = 869 ms, 95% CI (773, 965)] rather than pushed [M = 944 ms, 95% CI (849, 1040)] (p < .001). The interaction between joystick, scissor and mapping was not significant [F(1,2357) = 1.64, p = .201].
We also analyzed the reaction time difference between the left and right hand responses (joystick RTs—scissor RTs). The reaction times of the bimanual responses were closely synchronized (mean left hand lead = 20 ms, standard error = 8.5 ms). In the analysis of the reaction time difference, in which the joystick and scissors were treated as fixed factors and subject as a random intercept, did not reveal interaction between the joystick and scissor responses [F(1,2344) = 0.09, p = .766] suggesting that the synchrony between the joystick and scissor responses is similar in conditions that are hypothesized to be congruent (i.e., push-open and pull-close) and incongruent (i.e., push-close and pull-open).
Experiment 2
3.3% of the raw data containing errors were discarded from the analysis of reaction times of joystick and grip responses. The discarded error data consisted of those responses that were incorrectly executed with the left hand (1.2%), right hand (1.8%) or with both hands (0.3%). In addition, regarding the left-hand (joystick) responses, 4.6% of trials in which the RTs were more than two standard deviations from a participant’s overall mean were discarded from the analysis. Regarding the right-hand (grip) responses, 4.4% of trials in which the RTs were more than two standard deviations from a participant’s overall mean were additionally discarded from the analysis.
The errors were not analyzed for the joystick or scissor responses because the participants produced only few errors. The analysis of joystick responses revealed significant main effects of joystick [F(1,2635) = 38.25, p < .001] and grip [F(1,2635) = 4.76, p = .029]. Similarly, to Experiment 1, the backward responses were made faster [M = 759 ms, 95% CI (693, 824)] than the forward responses [M = 788 ms, 95% CI (723, 854)]. In addition, the precision responses were made faster [M = 768 ms, 95% CI (703, 834)] than the power responses [M = 779 ms, 95% CI (712, 844)]. Joystick × grip interaction was also obtained [F(1,2635) = 60.10, p < .001]. This interaction is presented in Fig. 3. The pairwise comparisons test showed that the push responses of the left hand were performed faster when the right hand was making the precision responses [M = 764 ms, 95% CI (698, 830)] rather than the power responses [M = 812 ms, 95% CI (746, 878)] (p < .001). In contrast, the pull responses were performed faster when the right hand was making the power responses [M = 745 ms, 95% CI (679, 811)] rather than the precision responses [M = 771 ms, 95% CI (706, 838)] (p < .001). The interaction between joystick, grip and mapping was not significant [F(1,2635) = 1.70, p = .192].

The mean vocal reaction times for Experiment 2 as a function of the joystick (i.e., the left-hand) and the grip (i.e., the right-hand) responses. Error bars depict the standard error of the mean. Asterisks indicate statistically significant differences (*** < . 001; **p < .01; *p < .05)
The analysis of grip responses revealed significant main effects of grip [F(1,2640) = 4.26, p = .039]. The precision responses were made faster (M = 764 ms, 95% CI [682, 845]) than the power responses (M = 774 ms, 95% CI [692, 855]). Joystick × grip interaction was also obtained [F(1,2640) = 58.72, p < .001]. This interaction is presented in Fig. 3. The pairwise comparisons test showed that the precision responses were performed faster when the joystick was pushed (M = 749 ms, 95% CI [667, 831]) rather than pulled (M = 779 ms, 95% CI [697, 860]) (p < .001). In contrast, the power responses were performed faster when the joystick was pulled (M = 751 ms, 95% CI [669, 833]) rather than pushed (M = 797 ms, 95% CI [715, 878]) (p < .001). The interaction between joystick, grip and mapping was not significant [F(1,2640) = 0.01, p = .926].
We also analyzed the reaction time difference between the left and right hand responses (joystick RTs—grip RTs). The reaction times of the bimanual responses were closely synchronized (mean left hand lead = 6 ms, standard error = 18.5 ms). The analysis was similar as in Experiment 1, with joystick and grip as the fixed factors, and did not reveal interaction between the joystick and grip responses [F(1,2610) = 0.12, p = .734].
Discussion
The present study demonstrated preference to couple the forward hand movement (i.e., proximal prehensile component) with the grip opening and the precision grip (i.e., distal prehensile components) whereas the backward hand movement was preferably coupled with the grip closure and the power grip. These interactions were observed even though the proximal and distal actions were performed with separate hands. This is the first time that the processes involved in planning of the specific proximal and distal prehensile actions are shown to interact in a systematic manner between two hands.
We propose that the push action was connected to the grip opening because both actions are related to an effector extension: in the push action, the arm is extended, whereas in the grip opening, the thumb and the fingers are extended. Similarly, we assume that the pull action was connected to the grip closure because both actions are related to an effector flexion: in the pull action, the arm is flexed, whereas in the grip closure, the thumb and the fingers are flexed. However, it should be noted that the push and pull actions are accomplished by moving the elbow as well as the shoulder. Importantly, in the push action, the elbow is extended whereas the shoulder is flexed (Lacquaniti and Soechting 1982). In contrast, in the pull action, the elbow is flexed whereas the shoulder is extended. Consequently, if the present interaction effect is indeed based on intra- and inter-limb connections between processes that prepare extension–flexion movements of proximal and distal components related to prehensile actions, it has to be exclusively related to elbow-related movement planning instead of shoulder-related movement planning. We should have observed the opposite interaction effect if the effect would operate in relation to motor processes that prepare shoulder extension–flexion movements.
Regarding the interaction between the push–pull and the precision–power responses observed in Experiment 2, similarly to the effect observed in Experiment 1, the interaction can be based on intra- and inter-limb connections between processes that prepare extension–flexion movements of proximal and distal components related to prehensile actions. According to this view, given that the power grip is accomplished by flexing four fingers toward the palm base mostly via the flexor digitorum profundus (Kilbreath and Gandevia 1994; Reilly and Schieber 2001), we propose that the pull response was preferably coupled with the power grip because there are intra- and inter-limb connections between motor representations that control elbow flexion and flexion of the four fingers.
In contrast to the power grip, the precision grip is mostly implemented by the flexor pollicis longus, which is responsible for flexing the thumb in this grip (Kilbreath and Gandevia 1994). Regarding the fingers that are typically actively flexed in the power grip (i.e., the little, ring and middle finger), the flexion of these fingers has to be inhibited in the precision grip, and in some extreme types of the pincer grip (e.g., when one is, for example, pinch gripping a needle) these fingers might be even extended (McMillan and Carin-Levy 2012). Similarly, in the current paradigm, when the task required executing the precision grip response, the accurate performance required flexor inhibition of the little, ring and middle fingers to withhold incorrectly performing the power grip. According to this view, when the participant had to select the precision grip for the response, the selection was executed relatively slowly when the precision grip response was coupled with the pull action because the pull action is associated with active flexion of the fingers, consequently favoring the power grip rather than the precision grip.
An alternative reason why the precision grip was associated with the push response assumes developmental links between outward directed reach action and the precision grip. It has been proposed that learning the precision grasp action precedes the development of hand pointing action (Leung and Rheingold 1981; Murphy and Messer 1977) in which the central component is the extension of an arm away from the body. According to this view, outward pointing action is adjusted from the precision grasp action by the end of the 1st year. Consequently, it is possible that there are developmentally driven connections between processes that prepare forward directed hand movements and the precision grip, which in turn manifest themselves in the present findings.
In previous studies that have used reaction time paradigm to explore intermanual interactions, reaction times have been associated with the onset (reaction time—RT) or the offset (movement time—MT) of, for example, the aiming movements (Diedrichsen et al. 2006; Heuer and Klein 2006). It has been previously also suggested that the congruency effects in dual-action tasks exploring intermanual interactions can be based on (1) interferences in assigning the specific stimulus to the correct response and/or (2) movement related conflicts reflecting interactions of movement parameters (e.g., Diedrichsen et al. 2006). In addition, it might be assumed, for example, that RTs are more sensitive to interferences in stimulus–response mapping processes, whereas MTs are more sensitive to movement-related conflicts. Hence, given that RTs and MTs might reflect different underlying mechanisms in the effects of intermanual interactions, the inability to report the congruency effects in relation to RTs as well as MTs can be considered as one of the potential weaknesses of the present study. The response devices of the study did not allow measuring RTs as well as MTs. In Experiments 1 and 2, the left-hand (joystick) responses were measured in relation to the offset of the movement. The right-hand responses of Experiment 1 were similarly measured in relation to offset of the opening/closing movement. In contrast, the right-hand responses of Experiment 2 were measured in relation to squeezing the precision of power grip devices. As such, the grip devices did not allow measurement of movement times. Consequently, we cannot be certain whether processes related to mapping the specific stimulus to the correct response could also have some role in the effects or whether the effects purely reflect interactions of movement parameters.
In conclusion, the present study shows a systematic interaction in proximal and distal prehensile components between two hands. The forward directed movement of the left hand was associated with the grip opening and the precision grip performed with the right hand. In contrast, the backward directed movement of the left hand was associated with the grip closure and the power grip performed with the right hand. The previous research exploring preferences to couple dual-actions simultaneously performed with two hands have mostly focused on directional aspects of the two movements showing, for example, a general preference to produce movements of two hands in the same direction in comparison to different directions (e.g., Swinnen et al. 1997; Serrien et al. 1999). The present study shows that, in addition to these inter-limb connections related to directional coding, the two hands also share motor programming processes in relation to planning proximal and distal prehensile components. Processes involved in planning the proximal prehensile action for one hand have systematic influence on processes involved in planning the distal prehensile action for another hand, and vice versa. In better detail, we propose that the motor system includes relatively tight inter- and intra-limb connections between the representations that prepare the elbow extension for the outward directed reaching, the finger extension for the grip opening, and the mechanisms that operate for preparing the precision grip. In addition, corresponding connections might be operating between the representations that prepare the elbow flexion for the inward directed arm movements, the flexion of fingers and the thumb for the grip closure, and the flexion of the four fingers for the power grip.
References
Armatas CA, Summers JJ, Bradshaw JL (1994) Mirror movements in normal adult subjects. J Clin Exp Neuropsychol 16:405–413
Barr DJ, Levy R, Scheepers C, Tily HJ (2013) Random effects structure for confirmatory hypothesis testing: keep it maximal. J Mem Lang 68:255–278
Cernacek J (1961) Contralateral motor irradiation-cerebral dominance: its changes in hemiparesis. Arch Neurol 4:165–172
Diedrichsen J, Hazeltine E, Kennerley S, Ivry RB (2001) Moving to directly cued locations abolishes spatial interference during bimanual actions. Psychol Sci 12:493–498
Diedrichsen J, Grafton S, Albert N, Hazeltine E, Ivry RB (2006) Goal-selection and movement-related conflict during bimanual reaching movements. Cereb Cortex 16:1729–1738
Dum RP, Strick PL (2005) Frontal lobe inputs to the digit representations of the motor areas on the lateral surface of the hemisphere. J Neurosci 25:1375–1386
Haggard P, Wing A (1995) Coordinated responses following mechanical perturbation of the arm during prehension. Exp Brain Res 102:483–494
Heuer H, Klein W (2006) The modulation of intermanual interactions during the specification of the directions of bimanual movements. Exp Brain Res 169:162–181
Jeannerod M (1984) The timing of natural prehension movements. J Mot Behav 16:235–254
Kilbreath SL, Gandevia SC (1994) Limited independent flexion of the thumb and fingers in human subjects. J Physiol 479:487
Lacquaniti F, Soechting JF (1982) Coordination of arm and wrist motion during a reaching task. J Neurosci 2:399–408
Leung EH, Rheingold HL (1981) Development of pointing as a social gesture. Dev Psychol 17:215
McMillan I, Carin-Levy G (2012) Tyldesley and grieve's muscles, nerves and movement in human occupation. Wiley, Blackwell
Murphy CM, Messer D (1977) Mothers, infants, and pointing: a study of a gesture. In: Schaffer HR (ed) Studies in mother-infant interaction. Academic Press, London, pp 325–354
Napier JR (1956) The prehensile movements of the human hand. Bone Jt J 38:902–913
Raos V, Umiltá MA, Gallese V, Fogassi L (2004) Functional properties of grasping-related neurons in the dorsal premotor area F2 of the macaque monkey. J Neurophysiol 92:1990–2002
Reilly KT, Schieber MH (2001) Electromyographic (EMG) activity of functional subdivisions in human multitendoned finger muscles. In: Society of Neuroscience Abstracts, vol 27, no 938.3. Washington, DC
Rosenbaum DA (1980) Human movement initiation: specification of arm, direction, and extent. J Exp Psychol Gen 109:444
Schieber MH, Hibbard LS (1993) How somatotopic is the motor cortex hand area? Science (Washington) 261:489–492
Serrien DJ, Bogaerts H, Suy E, Swinnen SP (1999) The identification of coordination constraints across planes of motion. Exp Brain Res 128:250–255
Stark E, Asher I, Abeles M (2007) Encoding of reach and grasp by single neurons in premotor cortex is independent of recording site. J Neurophysiol 97:3351–3364
Swinnen SP, Jardin K, Meulenbroek R, Dounskaia N, Hofkens-Van Den Brandt M (1997) Egocentric and allocentric constraints in the expression of patterns of interlimb coordination. J Cogn Neurosci 9:348–377
Swinnen SP, Dounskaia N, Levin O, Duysens J (2001) Constraints during bimanual coordination: the role of direction in relation to amplitude and force requirements. Behav Brain Res 123:201–218
Tiainen M, Tiippana K, Vainio M, Komeilipoor N, Vainio L (2017) Interaction in planning vocalizations and grasping. Q J Exp Psychol 70:1590–1602
Tucker M, Ellis R (2001) The potentiation of grasp types during visual object categorization. Vis Cogn 8:769–800
Vainio L, Symes E, Ellis R, Tucker M, Ottoboni G (2008) On the relations between action planning, object identification, and motor representations of observed actions and objects. Cognition 108:444–465
Vainio L, Tiainen M, Tiippana K, Komeilipoor N, Vainio M (2015) Interaction in planning movement direction for articulatory gestures and manual actions. Exp Brain Res 233:2951–2959
Vesia M, Prime SL, Yan X, Sergio LE, Crawford JD (2010) Specificity of human parietal saccade and reach regions during transcranial magnetic stimulation. J Neurosci 30:13053–13065
Vesia M, Bolton DA, Mochizuki G, Staines WR (2013) Human parietal and primary motor cortical interactions are selectively modulated during the transport and grip formation of goal-directed hand actions. Neuropsychologia 51:410–417
Acknowledgements
We are very grateful to Aleksi Rantala for helping us collecting the data. Many thanks to Martti Vainio and Kaisa Tiippana for getting involved in planning these studies. The research leading to these results has received funding from the Academy of Finland under Grant agreement numbers 1265610.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Vainio, L., Tiainen, M. Inter-limb coupling of proximal and distal hand actions. Exp Brain Res 236, 153–160 (2018). https://doi.org/10.1007/s00221-017-5114-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-017-5114-1