
Abstract
Previous studies have used a secondary probe reaction time (RT) task to assess attentional demands of a primary task. The current study used a startling acoustic stimulus (SAS) in a probe RT paradigm to test the hypothesis that attentional resources would be directly related to limitations in response preparation. Participants performed an easy or difficult version of a continuous primary task that was either primarily motor in nature (pursuit tracking) or cognitive (counting backward). Concurrently, participants responded to an auditory cue as fast as possible by performing a wrist extension secondary movement. On selected trials, the auditory cue was replaced with a SAS (120 dB), which is thought to involuntarily trigger a prepared response and thus bypass any response initiation bottleneck that may be present when trying to perform two movements. Although startle trials were performed at a shorter latency, both non-startle and startle probe trials resulted in a delayed RT, as compared to single-task trials, consistent with reduced preparation of the secondary task. In addition, analysis of SAS trial RT when a startle indicator was present versus absent provided evidence that the secondary task was at a lowered state of preparation when engaged in the cognitive primary task as compared to a motor primary task, suggesting a facilitative effect on preparatory activation when both the primary and secondary tasks are motoric in nature.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Researchers have long used reaction time (RT) paradigms, in which a participant must react as fast as possible to an imperative stimulus (IS) by producing a given movement, to assess the processes associated with response preparation and initiation. Based on results from the use of these paradigms, neural activity related to these processes has been described using an activation model (Hanes and Schall 1996), in which response initiation occurs once a group of cortical neurons responsible for the movement reach a threshold level of neural activation (Wickens et al. 1994). In a simple RT task, in which the required response is known in advance, one can minimize the time required to reach threshold by increasing the activation level of the neurons to a sub-threshold level prior to the IS via advance preparation. Then, following the IS, the activation of these neurons is further increased to supra-threshold levels resulting in movement initiation. In these models, RT would reflect the time required to raise activation levels from preparatory levels to initiation threshold.
More recently, a loud acoustic stimulus, capable of eliciting a reflexive startle response, has been used in conjunction with a simple RT paradigm to provide additional insight into the preparatory state of the motor system. When a startling acoustic stimulus (SAS) replaces the IS in a simple RT task, responses are triggered at such a short latency (i.e., <70 ms) that it is thought that the SAS acts to trigger the response directly, bypassing normal initiation processes (see Valls-Solé et al. 2008; Carlsen et al. 2012 for reviews). Although the same preparatory processes are assumed to occur, it is thought that the SAS acts via a brainstem-mediated pathway to produce a rapid rate of activation following the IS, resulting in dramatically shorter RTs (see Carlsen et al. 2012 for a detailed description). Thus by presenting a SAS either prior to or following the IS, the response latency to the SAS can be used as an indicator to determine the activation level of the motor system at any point in the preparation or initiation processes (e.g., MacKinnon et al. 2007; Carlsen and Mackinnon 2010; Alibiglou and MacKinnon 2012; Maslovat et al. 2014a). For example, if the presentation of a SAS prior to the IS results in similar RT latencies as when the SAS is presented in conjunction with the IS, it can be concluded that preparatory activation is at a similar level irrespective of time, whereas a delayed RT may be indicative of a reduced level of preparatory activation.
When engaged in a simple RT task, the single required response allows all preparatory resources of the participant to be focused on advance preparation of the movement. While this provides insight into simplified processes of response preparation and initiation, it does not probe the limits of the human’s ability to process information, nor does it mimic daily interactions in which multiple actions are required in response to multiple stimuli. In order to address these shortfalls, researchers have employed a dual-task paradigm in which participants must perform two actions either concurrently or in close temporal proximity. In these situations, the typical finding is that performance of one task is degraded as compared to when performed alone and further decrements are observed as the task difficulty increases (Kerr 1973). This dual-task “cost” is explained by a limitation associated with the requirement to concurrently perform multiple responses, which requires a resource broadly referred to as “attention” (Kahneman 1973). For example, one common dual-task method is the probe RT paradigm (Posner and Keele 1969) in which participants are required to perform a continuous primary task and then respond as fast as possible to an IS presented during the trial. An increase in the probe RT latency is thought to be indicative of greater resource allocation to the primary task, thus resulting in less attention being allocated to the secondary RT task (Glencross 1978). Although increased RTs in dual-task paradigms are often used to infer increased attentional demands of the tasks performed, they could also be considered to be indicative of limitations in the ability to concurrently prepare multiple responses. In the previously described activation model, an increase in probe RT could be attributed to preparatory resources being used to prepare the primary task and thus an inability to prepare the secondary task to a similar level of activation as when performed in isolation. A lowered level of secondary-task activation would result in greater time to reach initiation threshold and thus a delay in RT. In this manner, the allocation of attentional resources can be reflected in preparatory levels of the primary and secondary task.
One challenge in using a probe RT task to examine preparatory levels is that it is possible that changes in RT latency may also be confounded by a response initiation “bottleneck” (Keele 1973; De Jong 1993; Bratzke et al. 2009) whereby it is difficult to produce multiple responses simultaneously, particularly when using similar effectors. For example, both Lee and Elliott (1986) and McLeod (1980) found additional dual-task constraints when the primary and secondary tasks were manual in nature, as compared to a motor primary task and secondary vocal response, a result they attributed to a competition for similar response programming resources. One way to overcome any effects of a response initiation bottleneck is to utilize an atypical initiation process that would bypass the bottleneck. This might be achieved by using a SAS in place of the IS during the probe RT task, such that it would act as an involuntary response trigger for the secondary response (Valls-Solé et al. 2008; Carlsen et al. 2012). A comparison of the response latency to SAS-induced RT during both dual-task and single-task situations would provide novel insight into the state of motor preparation of the secondary task and thus the resource demands of the primary task.
Indeed, a SAS has been successfully utilized in both a dual-task psychological refractory period (Maslovat et al. 2013) and probe RT (Begeman et al. 2007) paradigm to examine the preparatory state of the second response during processing of a primary movement. In both experiments, it was found that RT on startle trials was faster than non-startle trials, but delayed in comparison with single-task trials, a result the researchers attributed to a decreased level of preparation of the secondary task. Thus, it appears that the use of a SAS can bypass any interference between responses and provide additional information pertaining to response preparation in a dual-task environment. In the work by Begeman et al., participants were required to perform a key-press task while performing a rhythmic oscillatory movement with the contralateral hand. However, one limitation of this design is that the requirement to simultaneously perform two manual responses may result in structural interference that does not accurately reflect the limitations of preparing multiple responses (Lee and Elliott 1986). Previous work examining the effects of interference between tasks of similar versus different response effectors has suggested that multiple manual responses are produced by a single limited capacity process, whereas a manual and vocal response involves separate, independent processes (McLeod 1977). Thus, in order to accurately assess the capacity of demands of the primary task, as well as response preparation of the secondary task, it is preferable to employ a secondary task which is likely to incur less structural interference (McLeod 1980; Lee and Elliott 1986).
The purpose of the current study was to use a SAS in a dual-task probe RT paradigm to assess how motor preparatory levels of a secondary task change depending on the resource allocations required by the primary task. In order to examine these effects, participants performed in a dual-task paradigm whereby a motor response was used as the probe RT task for all trials, and the primary task was altered such that it was either primarily cognitive or motor in nature, with differing levels of difficulty. Consistent with previous dual-task studies, it was predicted that probe RT on non-startle trials would be delayed in the dual-task paradigm, as compared to when performed in isolation (i.e., single-task RT), with a greater delay when the primary task was more difficult. It was also predicted that if the delayed probe RT (i.e., dual-task cost) was due to reduced preparation of the secondary task as a result of attentional limitations, startle trials would be performed at shorter latency as compared to control trials due to a faster initiation process (Carlsen et al. 2012), but would show a similar pattern of results to non-startle trials (i.e., dual-task RT would be longer than single-task RT, with an added delay when the primary task was more difficult). Conversely, if the delayed probe RT was due to a bottleneck in response initiation processes, it was predicted that all startle trials would be performed at a similar latency due to the involuntary triggering of the secondary response bypassing the bottleneck. Furthermore, a comparison of the dual-task interference effects when the primary task was cognitive versus motor provided information pertaining to the effects of structural interference. On one hand, there is evidence for greater interference when both tasks require manual responses, as compared to a vocal response during a manual tracking task (McLeod 1977). Alternatively, there is data indicating when performing a primary pointing movement, probe RT is faster for a manual versus vocal response (Lee and Elliott 1986), although in this study single-task probe RT was also faster for the manual versus vocal response. Through the use of a SAS, the current study allowed for a novel examination of the effects of structural interference when the secondary task was initiated via a typical (voluntary; non-startle trials) and atypical (involuntary; startle trials) process.
Materials and methods
Participants
Data were collected from fifteen right-handed or ambidextrous volunteers with no sensory or motor dysfunctions. However, in order to ensure participants exhibited a reliable startle response indicator, sternocleidomastoid (SCM) activation was examined for the first 120 ms following the SAS on startle trials during the single-task testing block (see “Data reduction and dependent measures” section for details). Five participants showed SCM activation in less than half the single-task startle trials and were excluded from the data analysis (see Carlsen et al. 2011 for more details regarding recommended inclusion criteria); thus, data are presented from 10 participants (six females, four males; age 24.4 ± 6.6 years). All participants signed a written informed consent and were naïve to the hypothesis under investigation. This study was approved by the Research Ethics Board at the University of Ottawa and was conducted in accordance with the latest revision of the Declaration of Helsinki.
Apparatus, tasks, and experimental design
Participants sat facing a 24″ LCD computer screen with both forearms secured with Velcro to armrests, parallel to the floor, and the palms facing inward (see Fig. 1). The right hand was attached to a manipulandum allowing horizontal flexion and extension movements about the wrist axis (used for secondary-task performance). The left hand gripped a handle which allowed pronation–supination motion about the wrist joint in the coronal plane (used during the motor-based primary task). A speaker was placed 30 cm behind the participant, in line with the vertical axis of the head.

Experimental setup, including location of the computer screen, manipulanda, and loudspeaker. The left hand was used to perform pursuit tracking in the motor primary task via pronation/supination movements, while the right hand performed a 20° targeted wrist extension as the secondary task
Participants performed a dual-task paradigm in which the primary task was either motor or cognitive in nature and had two difficulty levels. The motor primary task was a manual pursuit tracking task in which participants were required to track the movement of a yellow circular “ball” (.7 cm diameter) on the screen with a green rectangular virtual paddle within a 38.8 cm (w) × 1.6 cm (h) rectangular boundary zone. The ball randomly moved in the horizontal plane at 5.5 cm/s, and supination/pronation movements of the handle gripped by the left hand were translated into left and right movements of the paddle, whereby the extent of the horizontally bounded zone corresponded to ±30° from the neutral central position (i.e., the paddle moved .64 cm per degree of handle rotation). Starting position for the paddle and ball was in the middle of the horizontal movable space, which was equivalent to the hand being in a neutral position (handle perpendicular to the floor). Motor task difficulty was manipulated by changing the width of the paddle, making it easier (10.0× wider than the ball, 7 cm) or harder (3.3× wider than the ball, 2.33 cm) to keep the ball within the green paddle area. The cognitive primary task was a verbalized reverse counting task in which participants were required to count backward starting from a random number between 70 and 99 displayed on the screen. The easy version of the cognitive task consisted of counting backward by two and always began on an even number, and the difficult version consisted of counting backward by sevens from either an even or odd starting number.
Participants were instructed that if and when they heard an auditory tone during the performance of the primary task, they were to perform the secondary task consisting of a targeted 20° right wrist extension movement from the neutral start position. The auditory tone was delivered via the loudspeaker (MG Electronics M58-H, frequency response 300 Hz–11 kHz, rise time <1 ms) and was either a non-startling control tone (80 dB, 25 ms, 1000 Hz) or a white noise SAS (120 dB, 25 ms, equal power from 1 Hz to 22 kHz).
Each trial began with a visual “Get Ready” warning signal which disappeared 2000 ms later, followed by a 500-ms fixed foreperiod. Following this, the primary task was initiated which was indicated by the “ball” starting to move (motor), or by the appearance of the number to start counting in reverse from (cognitive). Trials lasted for 6 s and the secondary-task probe (if presented) occurred randomly between 2000 and 4000 ms after the beginning of the primary task. Following completion of the trial, feedback was presented on the screen for 3.5 s, consisting of points earned for the primary task and secondary task on a per trial basis, as well as the total points earned across trials. Primary-task points in the motor task were given based on the amount of time the ball spent in the paddle zone [points = time (ms)/10], and in the cognitive task was the total number of verbalized answers regardless of accuracy. Points were earned in the secondary task when RT was less than 350 ms. Instructions to participants were to earn as many points as possible with equal emphasis on the primary and secondary tasks.
Participants performed one block of dual-task trials for each condition (i.e., motor easy, motor difficult, cognitive easy, cognitive difficult; total of four blocks) with block order counterbalanced. Each block included 35 trials consisting of 24 control trials, eight SAS trials, and three catch trials (where no acoustic stimulus was presented). Trials were pseudorandomized such that no two consecutive trials included a SAS, and a SAS was not presented in the first two trials in any condition. Prior to testing in each block, participants first completed one practice trial for that particular condition with a non-startle secondary-task probe in order to familiarize participants with the dual-task trials to be performed. Following the completion of all four dual-task blocks, participants performed one block of single-task simple RT trials involving only the right hand wrist extension. Twenty trials were completed of which five were startle trials, pseudorandomized as outlined in dual-task testing. This testing block was completed to determine baseline RT as well as ensure consistent startle-related activation in the SCM during startle trials. Those participants who exhibited SCM activation in less than half the SAS trials in the single-task block were excluded from the analyses as they were considered to be “low responders.” In total, the testing session lasted approximately 1 h and involved four dual-task practice trials, 140 dual-task testing trials, and 20 single-task trials.
Recording equipment
EMG activity was recorded from the right extensor carpi radialis longus (ECR; agonist) and the flexor carpi radialis (FCR; antagonist) (i.e., the prime movers for the secondary task), as well as the left sternocleidomastoid (SCM; startle indicator) using bipolar pre-amplified surface electrodes (Delsys, Bagnoli DE-2.1) connected via shielded cabling to an external amplifier system (Delsys, Bagnoli-8). A reference electrode was placed on the right lateral epicondyle. Recording sites were prepared and cleansed in order to decrease electrical impedance. The electrodes were placed parallel to the muscle fibers and attached to the skin using double-sided adhesive strips. At the start of each trial, data collection was initiated by the computer and unfiltered EMG and position data from both manipulanda were digitally sampled at 1 kHz (National Instruments Inc., PCI-6030E) for 6 s using a customized program written in LabVIEW (National Instruments Inc.) and stored for offline analysis. Verbal responses during the cognitive primary task were recorded manually by the experimenter.
Data reduction and dependent measures
Practice trials were excluded from analysis, as were trials in which an error occurred for the secondary wrist extension movement (e.g., lack of movement, six single-task trials, 63 dual-task trials). Trials were also excluded due to secondary-task anticipation (one dual-task trial), slow response (i.e., >500 ms RT; 30 dual-task trials), and outlier trials in which RT was more than two standard deviations above the participant’s mean value for each condition (eight single-task trials, 34 dual-task trials). This resulted in a total inclusion rate of 91 % of trials (1458/1600 total).
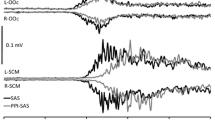
For the primary task, performance on the motor task was defined as the total amount of time the center of the ball stayed within the boundaries of the paddle whereas performance on the cognitive task was defined as the total number of verbalized responses (correct or not). For the secondary task, premotor RT was calculated as the time from the auditory cue to onset of EMG activity in the ECR. EMG onsets were defined as the first point where the rectified and filtered (25 Hz low pass elliptical filter) EMG activity first reached a value of two standard deviations above baseline levels (mean EMG activity in a 100-ms interval starting 500 ms prior to the probe) and was maintained for a minimum of 20 ms. EMG onset points were first determined using a custom LabVIEW (National Instruments Inc.) program and then were visually confirmed and manually adjusted (if necessary) to compensate for any errors due to the strictness of the algorithm. For startle trials, SCM onset was determined in a similar fashion, with reflex activation deemed to be present if onset occurred within 120 ms of the SAS (Carlsen et al. 2011).
Statistical analysis
Primary-task performance was analyzed using separate 2 difficulty (easy, difficult) × 3 stimulus (control, SAS, catch trials) repeated-measures analysis of variance (RM ANOVA) to determine whether performance on the primary task was affected by the type of secondary-task stimulus presented.
In contrast to previous studies that manipulated the secondary-task requirements (e.g., Lee and Elliott 1986), the current study altered the primary task to be either cognitive or motor-based. Due to a lack of equalization of attentional demands between the motor and cognitive primary tasks, secondary-task (RT) performance was analyzed separately for the cognitive and motor primary conditions. Thus, these analyses were completed using separate 2 stimulus (control, SAS) × 3 condition (single-task, dual-task easy, dual-task difficult) RM ANOVAs. This analysis compared secondary-task RT for SAS trials in which short-latency (<120 ms) SCM activation was present (i.e., SCM+) to control (80 dB) trials. Although secondary-task RT was the primary measure of preparation level, a complementary analysis was performed on the probability of eliciting a reflexive startle response in the SCM, as startle response incidence has been previously used as an indicator of preparation (Carlsen and Mackinnon 2010; Drummond et al. 2013). The proportion of SCM+ trials for each startle condition was subjected to an arcsine square-root transform and analyzed using separate one-way, 3 factor (Condition: single-task, dual-task easy, dual-task difficult) RM ANOVAs.
For the secondary-task analysis, only startle trials that were categorized as SCM+ were included in the analysis, as reflex-related activity in the SCM is thought to be necessary to ensure sufficient activation for response-triggering effects caused by the SAS (Carlsen et al. 2007). However, one challenge in the interpretation of startle-induced RT latencies is that it can be difficult to determine whether any reduction in RT is attributable to the triggering of a prepared response or to stimulus intensity effects (Woodworth 1938). One method used to make this distinction involves a comparison of RT latency between trials with and without SCM activation. Previous research has shown that when the required response is sufficiently prepared such that the SAS results in response triggering, SCM+ trials are performed at significantly shorter latency than SCM− trials (Carlsen et al. 2007; Honeycutt et al. 2015; Maslovat et al. 2015). Conversely, if the required response cannot be prepared to an extent where the movement is involuntarily triggered by the SAS, no significant difference is found between SCM+ and SCM− trials (Carlsen et al. 2009; Honeycutt et al. 2013). In these cases, SCM− RT values act as a control for the effects of stimulus intensity, whereby any reduction in RT compared to non-startle trials can be attributed to stimulus intensity effects. Thus, while SCM activation is necessary to evaluate response triggering by the SAS and thereby preparatory activation level, this activation does not necessitate early response triggering as reflexive startle activation can occur in the absence of a motor plan (Brown et al. 1991). This comparison was particularly important in the current study as performance in the dual-task paradigm was expected to result in fewer resources available to prepare the secondary task, and thus it was unclear whether the secondary task would be prepared to a sufficient level of activation for response triggering to occur. Thus, although separate analyses were performed for the secondary-task RT data due to a lack of equalization of attentional demands, an analysis of the relative effect of startle allowed a more direct comparison of preparation between the motor and cognitive condition. This was achieved by analyzing whether the presence of SCM activation resulted in differences in response latency between conditions. SAS trials for the dual-task motor and cognitive conditions were analyzed using a 2 SCM presence (SCM+, SCM−) × 2 task (motor, cognitive) × 2 difficulty (easy, difficult) RM ANOVA.
As there were only eight startle trials per condition, the separation of SCM+ and SCM− resulted in three missing data cells (out of 80) for the secondary-task analysis. These cells were filled using a regression-based multiple imputation technique (SPSS®, IBM Inc, Armonk, NY). In brief, five values were calculated for each missing data cell by drawing values from a probability distribution of values for each missing cell, and a mean of these five values used to fill the cell for the analysis. This method was chosen so that data from all participants could be included in the analysis while avoiding the confound of reduced variance associated with a single-filled value based on a mean or otherwise (Schafer and Graham 2002). Lastly, for one participant, there were no SCM− startle trials for any condition in the dual-task paradigm and thus data from this participant were excluded from the SCM comparison analysis. This SCM comparison analysis was not performed on the single-task trials as a large majority of trials showed SCM activation (80 vs. 61 % for dual-task trials), as would be expected when all resources were allocated to the preparation of one response, thus leading to a higher percentage of response triggering as evidenced by SCM activation.
For all analyses, in cases where sphericity was violated, Greenhouse–Geisser-corrected p values are reported. Partial eta squared (η 2 p ) values are reported as an estimate of effect size, and differences with a probability of less than .05 were considered significant. Post hoc analyses were performed using Bonferroni-corrected pairwise comparisons to determine the locus of any significant differences.
Results
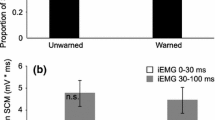
Analysis of the primary-task performance (Fig. 2a, b) confirmed a main effect for difficulty level for both the motor task, F(1, 9) = 469.56, p < .001, η 2 p = .99, and the cognitive task, F(1, 9) = 188.04, p < .001, η 2 p = .95. In addition, in the motor primary task, there was a main effect of stimulus, F(2, 18) = 8.22, p = .003, η 2 p = .48, which post hoc analysis confirmed was due to decreased performance on startle trials, compared to catch trials. In contrast, there was no significant effect of stimulus in the cognitive primary task, F(2, 18) = 4.38, p = .054, η 2 p = .33. No significant interaction effects were found [motor task, F(2, 18) = 1.76, p = .201, η 2 p = .16; cognitive task, F(2, 18) = .82, p = .455, η 2 p = .08].

Primary-task performance for the motor (top panel, a) and cognitive (bottom panel, b) task under both the easy and difficult conditions. A single asterisk denotes a main effect of difficulty level for both primary tasks, while a double asterisk denotes a significant reduction in motor performance on the startle trials (black bars) as compared to the catch (gray bars) trials. Error bars represent within-subject confidence intervals (Morey 2008)
Analysis of secondary-task performance (Fig. 3) confirmed a main effect for stimulus for both the motor primary task, F(1, 9) = 53.77, p < .001, η 2 p = .86, and the cognitive primary task, F(1, 9) = 88.90, p < .001, η 2 p = .91, with shorter RT values produced on startle trials. In addition, secondary-task RT values were affected by the testing condition for both the motor task, F(2, 18) = 18.34, p < .001, η 2 p = .67, and cognitive task, F(2, 18) = 36.56, p < .001, η 2 p = .80, which post hoc analyses confirmed was due to significantly faster RTs during single-task trials, compared to both difficulty levels of dual-task trials. Although mean RTs were longer for both startle and non-startle trials when engaged in the more difficult dual-task situation, this result did not reach statistical significance for either the motor (p = .239) or cognitive (p = .305) condition. No significant interaction effects were found [motor task, F(2, 18) = .35, p = .712, η 2 p = .04; cognitive task, F(2, 18) = 3.43, p = .054, η 2 p = .28].

Secondary-task performance during the motor (top panel, a) and cognitive (bottom panel, b) task for both startle (120 dB, SCM+ trials only) and non-startle (80 dB) trials. A single asterisk denotes a main effect of trial type, with significantly faster reaction time (RT) during the single-task condition (white bars) as compared to both the dual-task easy (gray bars) and dual-task difficult (black bars) conditions. A double asterisk denotes a main effect of reduced RT on startle trials as compared to non-startle trials. Error bars represent within-subject confidence intervals (Morey 2008)
The proportion of SAS trials that resulted in an observed burst of EMG activity in SCM (SCM+) was highest for the single-task trials (M = 80 %), as compared to all dual-task trials (motor easy, M = 55 %; motor difficult M = 61 %; cognitive easy, M = 63 %; cognitive difficult, M = 65 %). This difference in SCM+ proportion reached significance for the motor task, F(2, 18) = 4.77, p = .022, η 2 p = .35, due to a difference between the single task and motor easy dual task. For the cognitive task, no differences were found between SCM+ proportion, although a trend toward significance was present, F(2, 18) = 2.58, p = .103, η 2 p = .22.
For the comparison of secondary-task RT latency when SCM activation was present or absent (Fig. 4), the only significant effect was an SCM × task interaction, F(1, 8) = 14.31, p = .005, η 2 p = .64. Post hoc analyses confirmed this effect was due to a significantly lower RT on SCM+ trials (M = 143 ms) as compared to SCM− trials (M = 182 ms) for the motor task, whereas when participants were engaged in the cognitive task, no differences were found in RT latency for SCM+ trials (M = 191 ms) as compared to SCM− trials (M = 191 ms).

Secondary-task performance during the dual-task conditions, comparing performance on startle trials when sternocleidomastoid activation was present (SCM+) versus absent (SCM−). A single asterisk denotes a significantly faster RT on SCM+ trials for the motor conditions only. Error bars represent within-subject confidence intervals (Morey 2008)
Discussion
The purpose of the current study was to investigate the state of preparation of the secondary task in a dual-task probe RT paradigm using a SAS, which has previously been used to assess activation level of a prepared response in single-task RT paradigms (e.g., MacKinnon et al. 2007; Carlsen and Mackinnon 2010; Alibiglou and MacKinnon 2012; Maslovat et al. 2014a). This method was particularly useful as the increased activation associated with the startle reflex is thought to involuntarily trigger the prepared response via a subcortically mediated initiation process (Valls-Solé et al. 2008; Carlsen et al. 2012), thus bypassing any cortical response initiation bottleneck (see Maslovat et al. 2013 for similar rationale). In addition, the preparation of the secondary RT task was assessed when participants were engaged in either a cognitive or motor primary task with varying levels of difficulty, allowing for an examination of structural interference effects when the two tasks involved similar versus different response effectors. Analysis of the primary task indicated that the difficult version of each task led to reduced performance and that the requirement to perform a secondary task had little effect on primary-task performanceFootnote 1 (Fig. 2). This confirmed both appropriate manipulation of difficulty level and that participants allocated consistent resources on performance of the primary task regardless of whether secondary-task performance was required.
Given the similar allocation of attentional resources to the primary task across testing conditions, it was expected that RT latency for the secondary task would be an accurate measure of the preparatory activation level of the secondary response. As predicted, on non-startle trials, secondary-task RT was significantly longer when engaged in a dual-task paradigm, as compared to a single-task paradigm, irrespective of whether participants were engaged in a cognitive or motor primary task (Fig. 3). Furthermore, startle trials resulted in significantly shorter RTs but with a similar pattern of results as non-startle trials (Fig. 3). That is, even when engaged in a primary task, the secondary response was significantly speeded by the SAS, but not to the same extent as when only the single task was required. The similar magnitude RT increase observed in both startle and non-startle trials when participants engaged in a dual-task paradigm supports the assertion that the delayed RTs observed are the result of lower preparatory activation levels for the secondary task, rather than a response initiation bottleneck (Keele 1973; De Jong 1993; Bratzke et al. 2009). This is based on the prediction that if the delay in secondary-task performance was due to an initiation bottleneck, RT latency would have been similar for all startle trials (single-task and dual-task), which was clearly not the case (Fig. 3).
Further support for reduced preparatory levels during dual-task performance may be provided by examining the reflexive startle response in the SCM. Previous research has found that a decreased SCM burst amplitude may be associated with reduced preparation when comparing performance in paradigms such as choice versus simple RT (Maslovat et al. 2012) and go/no-go versus forced choice RT (Kumru et al. 2006). In the current study, the mean percentage of trials in which a SCM EMG burst was observed (SCM+) was lower for all dual-task conditions (overall M = 61 %), as compared to the single-task condition (M = 80 %), a result that reached significance for the motor primary task (p = .022) and approached significance for the cognitive primary task (p = .102). While this provides complementary evidence that participants were less engaged in preparing the wrist extension movement during the dual-task conditions, these results should be viewed with caution as the probability of eliciting a startle reflex is not always a direct measure of preparation levels. For example, some research has shown significant differences can exist in startle-elicited RTs, a result attributed to reduced preparatory activation, even though there is a consistently high probability of observing reflexive SCM activity (Maslovat et al. 2014b). Similarly, other work has shown a varying probability of response triggering by a SAS with an equal and elevated probability of SCM+ trials (Drummond et al. 2015). These results indicate that if the motor system is engaged, even at a lowered level of activation, it is still possible to consistently elicit startle-related SCM activation.
While the secondary-task performance results in the present experiment are consistent with previous probe RT findings, they can be considered and evaluated within a neural activation framework (Hanes and Schall 1996), in which attentional resources not allocated to the primary task are used to increase the activation level of the cortical neurons associated with secondary-task performance. As greater resources are required for primary-task performance, less sub-threshold preparatory activation of the secondary task can occur, resulting in a greater amount of time required to reach initiation threshold, which is reflected in lengthened response latency. In this manner, RT values would be expected to increase during dual-task performance with further increases predicted when primary-task difficulty increases. A similar explanation would account for startle trial results, whereby overall lowered RTs are due to initiation-related activation increases beginning earlier and occurring at a faster rate (Carlsen et al. 2012; Maslovat et al. 2014a); however, a lowered preparatory activation level of the secondary task would nevertheless result in increased startle probe RT latency compared to single-task trials. Thus, a similar pattern of results would be expected for startle and non-startle trials (as observed, see Fig. 3). In this manner, the utilization of a SAS in the current study provides novel insight into the preparatory state of the motor system when participants are engaged in either a motor or cognitive task.
The delayed startle RT during dual-task trials can present a challenge in terms of data interpretation as the observed response latency is within a similar range as that seen for voluntary initiation, as compared to the sub-100-ms latencies typically observed in response to a SAS in single-task studies (e.g., Valls-Solé et al. 1999; Carlsen et al. 2004). It is conceivable that the reduction in RT on dual-task startle trials is simply attributable to the more intense stimulus (Woodworth 1938), rather than triggering of a movement at a low level of preparatory activation. Therefore, trials with and without SCM activation were compared in order to differentiate stimulus intensity effects from response-triggering effects. When engaged in the motor primary task, results showed that trials with SCM activity (SCM+) exhibited significantly shorter RTs than SCM− trials (Fig. 4), indicative of response-triggering effects (Carlsen et al. 2007; Honeycutt et al. 2015; Maslovat et al. 2015). On the other hand, no differences were found between SCM+ and SCM− trials when participants were engaged in a cognitive primary task, suggesting that the reduced RT on these startle trials was more likely due to stimulus intensity effects, rather than response triggering (Carlsen et al. 2009; Honeycutt et al. 2013). The comparison of trials with and without SCM activation suggests that when participants were engaged in a motor-based primary task, they were able to also allocate sufficient attentional resources to increase activation of the motor-based secondary task to a level such that when the SAS resulted in SCM activation, the response was triggered at short latency relative to both SCM− and non-startle trials. Conversely, when participants were engaged in a cognitive-based primary task, they were unable to prepare the motor-based secondary task to a sufficient preparatory level such that even when the SAS resulted in SCM activation, no response-triggering effect was present. Although there was no direct comparison of secondary-task response latency between the motor and cognitive primary-task conditions, an examination of mean latencies provided additional (albeit indirect) support for a lowered level of secondary-task preparation when engaged in the cognitive primary task (Fig. 3). For example, the non-startle results showed the secondary-task RT latency was longer when engaged in the “easy” cognitive task (counting backward by twos; M = 253 ms) as compared to the “difficult” motor task (tracking with small paddle; M = 216 ms). This pattern of results was also present on startle trials (cognitive easy M = 176 ms versus motor difficult M = 147 ms), indicating less interference when both responses were motoric in nature, irrespective of whether the initiation process was voluntary or involuntary.
Based on the comparison of SCM+ versus SCM− trials (Fig. 4), it appears that when a primary task engages the motor system, there is a facilitative effect on the preparation of a motor-based secondary task. Conversely, when the motor system is minimally engaged in the primary task, such as only requiring a vocal response to a cognitively based process, there appears to be attenuation in the preparation of a motor-based secondary task, resulting in delayed response latency as well as a diminished response speeding effect of a SAS. This result is consistent with work by Lee and Elliott (1986), in which trials that required a vocal secondary response while engaged in a motor primary task were significantly longer than when the primary and secondary task were both manual in nature. However, the work by Lee and Elliott also found single-task probe RT was faster for the manual versus vocal response and the degree of interference in the manual–manual condition was larger than in the manual–vocal condition, leading the authors to conclude there are additional structural constraints involved in programming two manual movements. Similarly, McLeod (1977) required participants to engage in a continuous manual tracking task concurrently with a two-choice tone identification task, which required either a vocal or manual response. Results indicated that the continuous tracking task was performed significantly worse when the two-choice task required a manual response. This led the author to conclude the two manual responses involved a single limited capacity process, which provided additional interference leading to decreased performance. While these studies provide evidence for additional interference when both the primary and secondary tasks are motor-based, there are a number of differences between their experimental conditions and the current study. Specifically, the study by McLeod used a choice probe RT task, whereas the current study employed a simple probe RT task. It is possible that response selection processes are affected by dual-task requirements differently as compared to response production, resulting in greater interference when a decision is required regarding the secondary task. However, during a simple probe RT task when the secondary task is known in advance, such as that used in the current study, engagement of the motor system in the primary task may assist with preparation of a motor-based secondary task. In the work by Lee and Elliott, secondary-task demands were manipulated as compared to the current study which altered primary-task demands to be either motor or cognitive. While this experimental setup was required to use a SAS to assess secondary-task preparation, it did not allow for the equalization of attentional demands between the motor and cognitive primary tasks. Thus, further research may be warranted to continue to investigate differences in the ability to prepare dual tasks that involve varying degrees of structural interference.
Although the reported results are consistent with reduced preparation of the secondary task, RT latency was not significantly affected by difficulty level of the primary task. While it is possible that primary-task difficulty level was not sufficient to affect secondary-task performance, the primary-task performance was quite obviously affected by difficulty level (Fig. 2), making this explanation unlikely. A more likely alternative is that because the secondary probe occurred at random time points throughout the trial, it may not have always occurred when the participant was most engaged in the primary task. This explanation is consistent with an observed increased variability in performance, which likely resulted in a RT increase being nonsignificant, even though the longest RT values were observed for the most difficult motor and cognitive conditions (Fig. 3). This result highlights some of the challenges associated with assessing attentional demands using a continuous primary task as opposed to pairs of simple punctate task. Often, continuous tasks fail to show dual-task interference, which can be used as evidence for a lack of bottleneck, rather than an intermittent overlap in central processing (see Pashler 1994, for more details). The current study showed a clear effect of dual-task interference on both startle and non-startle trials (Fig. 3); however, a more rigorous control of probe timing may be necessary to accurately assess the effect of difficulty level on the attentional demands of both motor and cognitive processing.
One possible confound in the present study is that single-task performance was always assessed following dual-task performance and thus could have shown reduced RT due to practice effects. As the proportion of SCM+ startle trials during the single-task condition was used as an inclusion criterion for participants, it was necessary to have these trials at the end of the testing session to account for any habituation to the SAS (Brown et al. 1991). By performing the single-task trials last, we ensured that all included participants consistently exhibited a startle response (even after 32 SAS presentations during the dual-task blocks) allowing for accurate conclusions to be made regarding the startle-related data, including the comparison of SCM+ to SCM− trials. Although this protocol did not allow for control of practice effects, the single-task data suggest these effects would be minimal, especially as the attentional resources during the dual-task testing trials would be mostly applied toward performance of the primary task. The reported single-task RT results, even following multiple dual-task testing blocks, are somewhat longer than those seen in similar previous RT tasks in our laboratory (Maslovat et al. 2014a; non-startle RT of 127 ms compared to 155 ms in the current study, startle RT of 90 ms as compared to 96 ms in the current study).
In summary, the results of the current study provide strong and novel evidence for reduced secondary-task preparation during a dual-task probe RT paradigm. Even though the presentation of a SAS can directly trigger a response and bypass any response initiation bottleneck, startled dual-task trials still resulted in significantly delayed response latency, consistent with a diminished level of preparatory activation. Interestingly, the SAS appears to have led to response triggering of the secondary motor task when participants were engaged in a motor-based primary task (albeit delayed compared to single-task trials), but not when engaged in a cognitive-based primary task. This differential effect of the SAS suggests there may be a difference in ability to prepare multiple responses that involve motor and cognitive components, as compared to when both tasks are primarily motoric in nature.
Notes
Note there was a small but significant decrease in motor primary task performance on startle trials as compared to catch trials. This is likely due to the reflexive startle response temporarily interfering with the tracking task rather than any differences in attentional allocation.
References
Alibiglou L, MacKinnon CD (2012) The early release of planned movement by acoustic startle can be delayed by transcranial magnetic stimulation over motor cortex. J Physiol 590:919–936
Begeman M, Kumru H, Leenders K, Valls-Sole J (2007) Unilateral reaction time task is delayed during contralateral movements. Exp Brain Res 181:469–475
Bratzke D, Rolke B, Ulrich R (2009) The source of execution-related dual-task interference: motor bottleneck or response monitoring? J Exp Psychol Hum Percept Perform 35:1413–1426
Brown P, Rothwell JC, Thompson PD, Britton TC, Day BL, Marsden CD (1991) New observations on the normal auditory startle reflex in man. Brain 114:1891–1902
Carlsen AN, Mackinnon CD (2010) Motor preparation is modulated by the resolution of the response timing information. Brain Res 1322:38–49
Carlsen AN, Chua R, Inglis JT, Sanderson DJ, Franks IM (2004) Can prepared responses be stored subcortically? Exp Brain Res 159:301–309
Carlsen AN, Dakin CJ, Chua R, Franks IM (2007) Startle produces early response latencies that are distinct from stimulus intensity effects. Exp Brain Res 176:199–205
Carlsen AN, Chua R, Inglis JT, Sanderson DJ, Franks IM (2009) Differential effects of startle on reaction time for finger and arm movements. J Neurophysiol 101:306–314
Carlsen AN, Maslovat D, Lam MY, Chua R, Franks IM (2011) Considerations for the use of a startling acoustic stimulus in studies of motor preparation in humans. Neurosci Biobehav Rev 35:366–376
Carlsen AN, Maslovat D, Franks IM (2012) Preparation for voluntary movement in healthy and clincial populations: evidence from startle. Clin Neurophysiol 123:21–33
De Jong R (1993) Multiple bottlenecks in overlapping task performance. J Exp Psychol Hum Percept Perform 19:965–980
Drummond NM, Carlsen AN, Cressman EK (2013) Motor preparation is delayed for both directly and indirectly cued movements during an anticipation-timing task. Brain Res 1506:44–57
Drummond NM, Cressman EK, Carlsen AN (2015) Inhibition of motor-related activation during a simple reaction time task requiring visuomotor mental rotation. Behav Neurosci 129:160–169
Glencross DJ (1978) Response planning and the organization of speed movements. In: Nickerson RS (ed) Attention and performance VII. Erlbaum, Hillsdale, NJ, pp 107–125
Hanes DP, Schall JD (1996) Neural control of voluntary movement initiation. Science 274:427–430
Honeycutt CF, Kharouta M, Perreault EJ (2013) Evidence for reticulospinal contributions to coordinated finger movements in humans. J Neurophysiol 110:1476–1483
Honeycutt CF, Tresch UA, Perreault EJ (2015) Startling acoustic stimuli can evoke fast hand extension movements in stroke survivors. Clin Neurophysiol 126:160–164
Kahneman D (1973) Attention and effort. Prentice Hall, Englewood Cliffs
Keele SW (1973) Attention and human performance. Goodyear, Palisades
Kerr B (1973) Processing demands during mental operations. Mem Cogn 1:401–412
Kumru H, Urra X, Compta Y, Castellote JM, Turbau J, Valls-Solé J (2006) Excitability of subcortical motor circuits in Go/noGo and forced choice reaction time tasks. Neurosci Lett 406:66–70
Lee TD, Elliott D (1986) Probing the motor program: effects of output competition during movement preparation. Hum Mov Sci 5:173–183
MacKinnon CD, Bissig D, Chiusano J et al (2007) Preparation of anticipatory postural adjustments prior to stepping. J Neurophysiol 97:4368–4379
Maslovat D, Carlsen AN, Franks IM (2012) Subcortical motor circuit excitability during simple and choice reaction time. Behav Neurosci 126:499–503
Maslovat D, Chua R, Spencer HC, Forgaard CJ, Carlsen AN, Franks IM (2013) Evidence for a response preparation bottleneck during dual-task performance: effect of a startling acoustic stimulus on the psychological refractory period. Acta Psychol 144:481–487
Maslovat D, Carter MJ, Kennefick M, Carlsen AN (2014a) Startle neural activity is additive with normal cortical initiation-related activation. Neurosci Lett 558:164–168
Maslovat D, Klapp ST, Jagacinski RJ, Franks IM (2014b) Control of response timing occurs during the simple reaction time interval but on-line for choice reaction time. J Exp Psychol Hum Percept Perform 40:2005–2021
Maslovat D, Franks IM, Leguerrier A, Carlsen AN (2015) Responses to startling acoustic stimuli indicate that movement-related activation is constant prior to action: a replication with an alternate interpretation. Physiol Rep 3:e12300
McLeod P (1977) A dual task response modality effect: support for multiprocessor models of attention. Q J Exp Psychol 29:651–667
McLeod P (1980) What can probe RT tell us about the attentional demands of movement? In: Stelmach GE, Requin J (eds) Tutorials in motor behavior. North-Holland, Amsterdam, pp 579–589
Morey RD (2008) Confidence intervals from normalized data: a correction to Cousineau (2005). Tutor Quant Methods Psychol 4:61–64
Pashler HE (1994) Dual-task interference in simple tasks: data and theory. Psychol Bull 116:220–244
Posner MI, Keele SW (1969) Attentional demands of movement. In: Proceedings of the 16th congress of applied psychology. Swets and Zeitlinger, Amsterdam
Schafer JL, Graham JW (2002) Missing data: our view of the state of the art. Psychol Methods 7:147–177
Valls-Solé J, Rothwell JC, Goulart FR, Cossu G (1999) Patterned ballistic movements triggered by a startle in healthy humans. J Physiol 516:931–938
Valls-Solé J, Kumru H, Kofler M (2008) Interaction between startle and voluntary reactions in humans. Exp Brain Res 187:497–507
Wickens J, Hyland B, Anson G (1994) Cortical cell assemblies: a possible mechanism for motor programs. J Mot Behav 26:66–82
Woodworth RS (1938) Experimental Psychology. Holt, New York
Acknowledgments
Supported by a Natural Sciences and Engineering Research Council of Canada (NSERC) discovery grant awarded to ANC (RGPIN: 418361-2012). We would also like to acknowledge the assistance of two anonymous reviewers for their constructive comments on earlier versions of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Maslovat, D., Drummond, N.M., Carter, M.J. et al. Reduced motor preparation during dual-task performance: evidence from startle. Exp Brain Res 233, 2673–2683 (2015). https://doi.org/10.1007/s00221-015-4340-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-015-4340-7