
Abstract
Rationale
Environmental enrichment (EE) could influence brain plasticity and behavior in rodents. Whether the early EE may predispose individuals to a particular social hierarchy in the social dominance tube test (SDTT) at adulthood is still unknown.
Objective
The present study directly investigated the influence of EE on competitive success in the SDTT among adult rats.
Methods
Male rats were maintained in EE from postnatal days 21 to 35. Social dominance behavior was determined by SDTT, competitive food foraging test, and mate preference test at adulthood. IBA-1 expression in the hypothalamus was examined using immunohistochemistry and western blot.
Results
EE rats were prone to become submissive during a social encounter with standard environment (SE) rats in the SDTT. No difference was found in food foraging in the competitive food foraging test between SE and EE rats. Male EE rats were more attractive than the SE to the female rats in the mate preference test. IBA-1 expression was found to be decreased in the hypothalamus of EE rats compared to SE group. Infusion of a microglia inhibitor reduced percentage of forward in SE rats in the SDTT. Infusion of DNA methyltransferase inhibitor prevented the development of subordinate status in EE rats and restored the expression of IBA-1 in the hypothalamus.
Conclusions
The results suggest that early EE did not lead to reduced social hierarchy in the male rat. However, EE caused a reduction in the percentage of forward in the SDTT, which might be associated with reduced number of microglia in the hypothalamus.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Environmental enrichment (EE) is a combination of both physical and psychological stimulation that not only consists of enriched physical environment (such as a large housing room is filled with novelties and toys that are routinely changed during the experimental period) but also includes complex social interactions (Cao et al. 2014; Nithianantharajah and Hannan 2006). It has been shown that EE has beneficial effects on brain development, cognitive functions, and the rate of recovery from neural injury (Miller et al. 2013). In rodents, early EE decreases emotional reactivity (Cao et al. 2014) and ameliorates age-dependent cognitive decline (Lores-Arnaiz et al. 2006). EE could also reduce lesion size and rescue behavioral deficits in post-stroke rats (Buchhold et al. 2007). In our previous study, we found that environmental enrichment could attenuate the anxiety-like behavior in the adult offspring of morphine-exposed rats (Li et al. 2014). This evidence indicates that EE could effectively promote the well-being of rodents and is protective against insults. However, the role of early environmental enrichment in the establishment of social dominance at adulthood is still unknown. Social dominance tube test is an assay of social dominance (SDTT) in which a forced encounter between two unfamiliar animals is observed (Garfield et al. 2011; Wang et al. 2011).
Microglia are resident macrophages of the central nervous system (CNS) and play a key role in neuroimmunomodulation. Homeostasis and development of microglia are dependent on specific survival factors, such as interleukin-34 (IL-34), colony-stimulating factor 1 (CSF-1), or transcription factors, such as PU.1, IRF8, RUNX1, and DAP12 (Erny et al. 2015). A recent study has indicated that microglia in the adult brain are physiologically dependent on colony-stimulating factor 1 receptor (CSF1R) signaling (Elmore et al. 2014). CSF1R is expressed by macrophages and microglia and has two natural ligands: CSF1 and IL-34 (Patel and Player 2009). CSF-1 and IL-34 compete for binding to the CSF-1R and are expressed in a largely non-overlapping manner by mature neurons (Chitu et al. 2016). Although the role of microglia in neurological and psychiatric diseases as well as CNS injury has been extensively investigated (Pearse and Hughes 2016), microglia also appear to be necessary for normal brain function (Kettenmann et al. 2011). The masculinization of adult rat behavior is suggested to be regulated by the activity of microglia in the medial preoptic area at birth (Lenz et al. 2013). It has been shown that inhibiting microglia activation by administration of minocycline (known to work in the brain by inhibiting activation of microglia) may promote social behavior in humans (Kato et al. 2012; Watabe et al. 2012). In mice, deficit in microglial function during early postnatal weeks is sufficient to induce impaired social behavior (Zhan et al. 2014). These two conflicting results emphasize the insufficient understanding of the role of microglia in the regulation of social behavior under normal conditions. Social hierarchy can have a significant impact on neuroimmunity in the rodent brain (Audet et al. 2010; Barnum et al. 2008). However, it remains unclear whether microglia are involved in the establishment of social hierarchy.
We undertook the present study to determine whether microglia might be involved in the effect of early environmental enrichment on the establishment of social dominance in the SDTT. We observed that EE rats were more likely to back down than their SE “opponents” in the social dominance tube test, but EE rats were found to be preferred by female rats in the mate preference test and exhibited the same level of food foraging in the competitive food foraging test. We also identified that microglia might be causally related with the percentage of forward in the social dominance tube test, which was demonstrated by pharmacological approaches. In line with the reduced expression of IBA-1 in the hypothalamus of EE, we found that expression of the genes associated with proliferation of adult microglia (CSF1 and IL-34) was reduced in the hypothalamus of EE rats, compared to SE, accompanied by increased level of DNA methylation. In weight-matched SE animals, infusion of specific microglia inhibitor reduced percentage of forward, mimicking the low probability to become dominant observed in EE animals. Conversely, infusion of DNA methyltransferase inhibitor prevented the development of a subordinate status in EE rats and restored the expression of IBA-1in the hypothalamus.
Material and methods
Animals
A total of 208 male Sprague-Dawley rats (inbred strain, Animal Center, XiangYa School of Medicine, Central South University) were used. Animals were housed in groups of four or five in an animal facility with 12-h light/dark cycles, at a constant temperature (22 °C) and humidity-controlled (50 ± 5%) animal facility, with free access to food and water. The experimental protocol was approved by the Animal Care and Use Committee of Central South University and conformed to the National Institutes of Health Guide for the Care and Use of Laboratory Animals (no. 2013-99). All efforts were made to minimize suffering as well as the number of rats used.
Environmental enrichment
After weaning (at 3 weeks of age), rats were randomly divided in two different housing conditions: (1) standard environment (SE)—rats were housed in the SE throughout the experiment and (2) environmental enrichment (EE)—rats were kept in EE conditions from postnatal days 21 to 35, according to a previous study (Arai et al. 2009; Cao et al. 2014). The EE was carried out in a large open field (150 cm × 150 cm × 50 cm) from 8:00 a.m. to 16:00 p.m., containing different toys (small house, small ladder, wheel runners, and various pet toys) and tubes that were changed every day, introducing new toys of different shapes and colors to stimulate active exploration (Li et al. 2013). All the behavioral tests were carried out at postnatal day 71.
Drugs and drug administration
Minocycline hydrochloride (Sigma-Aldrich, USA) was dissolved in normal saline and administered i.p. at 40 mg/kg once daily for 3 days (Suarez-Roca et al. 2014). The animals were assessed in the social dominance tube test 1 h after the last treatment. 5-Azacytidine (5AZ) (Sigma-Aldrich, USA) was dissolved in normal saline and administered i.p. at 0.4 mg/kg once daily for 6 days (Sales et al. 2011), and the animals were assessed in the social dominance tube test 1 h after the last treatment. All drugs were freshly prepared.
Behavioral tests
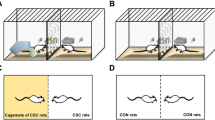
Social dominance tube test
Social dominance tube test is an assay of social dominance in which a forced encounter between two unfamiliar animals is observed (Spencer et al. 2005). The apparatus is a transparent tube 1.5 m in length and 6 cm inner diameter, a size just sufficient to permit one adult rat to pass through without reversing the direction. An acclimation process was applied to ensure that rats would readily move through the tube. Two days before the test, each rat was encouraged to move through the tube twice, and then the animal was returned to its home cage. During the test, the two weight-matched rats were released simultaneously into the opposite ends of the tube and one rat would attempt to show dominance and force the other back out of the tube. The rat that first forward from the tube within 2 min was designated the “winner.” In rare cases when no rat forward within 2 min, the test ended in a draw. Each pair of rats was tested twice, and their starting positions were swapped. In addition, another group of rats underwent three trials (paired with the same rat), with a 24-h inter-trial interval. Between each trial, the tube was cleaned with 70% ethanol. The percentage of forward was calculated from the two number of encounters. And, the Mann-Whitney U test was used to analyze the data (Shahbazian et al. 2002).
Mate preference test
The mate preference test was carried out according to a previous study (Crews et al. 2007), with some modifications. The apparatus consists of a large three-chamber glass-testing arena (120 cm × 48 cm × 54 cm). At either end of the box were small (28 × 28 × 12.5 cm) compartments, separated by wire-mesh barriers that permit exchange of visual, olfactory, and tactile cues. The large compartment was divided into three parts, the boundaries of which were marked by tape. The central part was defined as the non-social preference area; the other two parts (SE target side containing an SE rat; EE target side containing an EE rat) were preference areas. The female rats (with the same hormone level) were placed in the middle of the central part and allowed to explore the large compartment for 10 min. The amount of time that the female rat (without sexual experience) spent in one of the small compartments was recorded as soon as all four paws of the experimental animal crossed over the taped boundary line of that partner’s compartment. The event terminated as soon as one paw crossed over the boundary line back into the center compartment. The preference for mate was defined as the cumulative time spent on the SE target side or EE target side. The SE or EE target rat and the experimental female rat were not familiar with each other before the test. Experimental female rats and social target rats were used in this paradigm only once each. At the end of each test, all animals were removed, and the entire testing arena was washed with 70% ethanol to remove scent marks and residual odors.
Competitive food foraging test
The competitive food foraging test was performed according to our previous study (Li et al. 2012), with some modification. The test was carried out in an open field (150 cm × 150 cm × 50 cm). One SE rat (resident in the small plastic home cage (30 cm × 18 cm × 16 cm) for 7 days) with a metal wire cover with 250 g of standard food pellets on the wire mesh was placed in the corner of the open field, and then a test rat that had been deprived of food for 12 h was placed into the open field. Rats had free access to water during the test. The rat residing in the small home cage and the test rat came from different litters and never interacted with each other before the test. The test rat was allowed to navigate to the cage and foraged food freely from 7:00 p.m. to 9:00 p.m. The amount of food moved to the open field was calculated at 9:00 p.m.
Western blot
To exclude the effect of behavioral tests on the result, another group of rats (at postnatal day 71) that never experienced the behavioral tests were weighed and sacrificed by rapid decapitation under deep anesthesia (halothane) according to our previous study (Cao et al. 2013). The whole hypothalamus, medial prefrontal cortex, and hippocampus were removed and snap-frozen in liquid nitrogen. Frozen samples were homogenized (PRO Scientific Inc., U.S.) in a cocktail lysis buffer containing protease inhibitors and phosphatase inhibitors (Roche Applied Science, Mannheim, Germany). Samples were then centrifuged at 12,000×g for 20 min at 4 °C, and the supernatants were used for Western blot analysis. A total of 20 μg protein from each sample was loaded in and separated by 10 or 15% Bis-Tris SDS-PAGE gel. The blotted proteins were transferred to a nitrocellulose membrane. Membranes were blocked with 5% nonfat milk for 2 h at room temperature and incubated overnight with the following primary antibodies: mouse polyclonal antibody GAPDH (Boster, 1:4000), goat anti-IBA-1 (Abcam, 1:1000), mouse anti-GFAP antibody (Santa Cruz, 1:4000), mouse anti-NeuN (Millipore, 1:1000). Membranes were washed and further incubated with HRP-conjugated secondary antibodies (CWBIO, 1:1000) for 2 h at room temperature. After three rinses (10 min in each) in TBS-T buffer, immunoreactivity was detected with an ECL Western Blotting Detection Kit (CWBIO, China). Results were standardized to GAPDH control protein.
Immunohistochemistry
Expression of IBA-1 was determined immunohistochemically, as described in our previous study (Cao et al. 2013). After an overdose of 10% with chloral hydrate (Cao et al. 2013; Dai et al. 2011), the rats that never experienced the behavioral tests were sacrificed, brains were dissected and removed after transcardiac perfusion with 0.9% NaCl, followed by immersion in a solution of 4% paraformaldehyde in phosphate buffer. After dissection, brains were stored at 4 °C in 4% paraformaldehyde overnight, and then transferred to phosphate-buffered saline (PBS) containing 30% sucrose. Coronal sections (30 μm) were cut, incubated at room temperature in a blocking buffer solution containing 5% bovine serum and 0.1% Triton X-100 in 0.01 M PBS, and then incubated overnight at 4 °C with goat anti-IBA-1 antibody (abcam, 1:1000). The secondary reagents used for localization were biotinylated rabbit anti-goat IgG (CWBIO, 1:200) and ABC kit (Vector Labs). Diaminobenzidine tetrahydrochloride (DAB) (Vector Labs) was used as a peroxidase substrate.
IBA-1-immunoreactive (IBA-1-ir) cells were counted in medial preoptic area of hypothalamus, medial prefrontal cortex, and hippocampus. The number of IBA-1-ir cells was quantified on both sides of the section. IBA-1-ir was visualized using a Nikon (H600L light microscope) and HPIAS-1000 image analysis software (Tonji Qingping Company, China). Briefly, the microscope, camera, and software were background corrected and normalized to preset light levels to ensure fidelity of data acquisition. Analysis was conducted by an experimenter who was blinded to experimental treatments. Data were acquired from a minimum of three sections/brain regions/animals and the data averaged to obtain a single value per rat.
RNA isolation and qRT-PCR
Total RNA was extracted using the Trizol® reagent (Invitrogen, USA), according to the manufacturer’s instructions. The A260 nm/A280 nm absorption ratio was used to determine RNA purity. cDNA synthesis was performed with the RevertAid™ First-Strand cDNA synthesis kit (Fermentas, no. 1622) according to the manufacturer’s instructions, using 1 μg of total RNA. cDNA was stored at − 20 °C. Expression levels of genes were determined by Bio-Rad real-time PCR system employing the SYBR® Green PCR Master Mix (SsoFast™ EvaGreen® Supermix, Bio-Rad). The Gapdh gene expression was used as an internal control. Primers were designed with Primer 3 software (Supplementary materials).
A two-step PCR protocol was used according to the manufacturer’s instructions. PCR cycling conditions were as follows: 30 s at 95 °C, followed by 45 cycles at 95 °C for 10 s and 60 °C for 30 s. Samples were processed in technical duplicates, and a melting analysis was performed for each sample at the end of the PCR. Relative gene expression was determined by 2−△△Ct method according to our previous study (Cao et al. 2014).
ELISA determination
We investigated whether differences in the level of corticosterone and testosterone might explain the low competitive success of EE rats in the social dominance tube test. Thus, we examined corticosterone and testosterone levels in SE and EE rats under basal conditions and immediately following social dominance tube test. A Rat Corticosterone and Testosterone ELISA Kit (CUSABIO, China) were used to determine the serum corticosterone and testosterone levels, according to the manufacturer’s instructions. At postnatal day 71, both the SE and EE rats at basic condition or immediately after social dominance tube test were decapitated and trunk blood was collected and centrifuged for 15 min at 1000×g at 4 °C. The supernatants were collected and used to measure the corticosterone and testosterone levels of each rat. Fifty microliters of standard or sample was added to each well. Then, 50 μl of HRP-conjugate corticosterone or testosterone and 50 μl of (corticosterone or testosterone) antibody were also added to each well. All the components were mixed thoroughly and then incubated for 1 h at 37 °C. Then, they were incubated with a substrate solution for 30 min at 37 °C. Wells were then developed with tetramethylbenzidine reagent in dark and the absorbance was measured at 450 nm.
Statistical analysis
Statistical analysis was performed using software Prism 5.0 (Graphpad Soft ware, San Diego, CA). The results were presented as mean ± SEM. The behavioral tests were analyzed using the two-tailed Student’s t test or one-way or two-way ANOVA followed by post hoc Bonferroni testing. Tube test was analyzed using Mann-Whitney test. A value of p < 0.05 was considered statistically significant.
Results
Effects of environmental enrichment on the social dominance tube test at adulthood
We used social dominance tube test to explore the effect of environmental enrichment on the establishment of social hierarchy. Rats were paired such that within-pair differences in body weights between SE and EE groups were not statistically significant (t = 1.951, p > 0.05) (Fig. 1a). We found that rats that experienced environmental enrichment exposures during early time showed a significant reduction in the percentage of forward when compared to conspecific rats that were housed in standard environment (*** p < 0.001) (Fig. 1b). Importantly, we found that the subordinate character of EE rats was not only apparent from the initial interaction but also sustained throughout the three social dominance tube tests (Day 1, ***p < 0.001; Day 2, **p < 0.01; Day 3, **p < 0.01) (Fig. 1c), suggesting that the reduced percentage of forward in these animals was long-lasting. Although environmental enrichment lead to clear winner and loser effect in the social dominance tube test, this was not the case for SE rats to their SE “opponents” (p > 0.05) (Fig. 1d), indicating that early experience indeed affected the behavior of EE rats in the social dominance tube test.

Effect of early environmental enrichment on the social dominance tube test at adulthood. a No difference was observed in the weights of adult SE and EE rats, n = 40 per group. b EE rats exhibited remarkably lower percent of forward in the social dominance tube test, n = 40 pairs. c EE rats demonstrated persistently lower percent of forward in three consecutive social dominance tube tests, n = 10 pairs. d No difference was observed in the percent of forward between the SE rats in the social dominance tube test, n = 12 pairs. e No difference in the corticosterone level pre- and post-dominance tube test between SE and EE groups, but dominance tube test increased corticosterone in EE and SE, n = 7 in each group. f No difference in testosterone level pre- and post-dominance tube test between SE and EE groups, n = 7 in each group. *p < 0.05; **p < 0.01; ***p < 0.001 vs SE; ##p < 0.01 pre-tube test vs post-tube test
To investigate possible neuroendocrine mechanisms contributing to this difference in competitive success in the social dominance tube test, we also examined corticosterone and testosterone levels in SE and EE rats under basal conditions and immediately following social dominance tube test. Two-way ANOVA indicated no main effects of environmental enrichment (ANOVA, F (1, 20) = 0.2533, p > 0.05) and interaction (F (1, 22) = 0.5525, p > 0.05) on the level of corticosterone in the rats, but the tube test led to increase in the level of corticosterone in both groups (F (1, 22) = 8.437, ** p < 0.01) (Fig. 1e). Two-way ANOVA indicated no main effect of environmental enrichment (F (1, 20) = 0.04, p > 0.05) and tube test (F (1, 20) =1.66, p > 0.05) and their interaction ((F (1, 20) = 0.8198, p > 0.05) on the level of circulating testosterone (Fig. 1f).
Effects of environmental enrichment on the expression of microglia marker IBA-1 in the adult brain
Since no difference was observed in the corticosterone and testosterone levels between the EE and SE groups, we explored why EE rat exhibited reduced percent of forward in the social dominance tube test. Microglia in the hypothalamus (particularly the medial preoptic area) play a key role in the regulation of social behavior. Thus, we used immunohistochemistry to determine the expression of microglia marker IBA-1 in the medial preoptic area between SE and EE rats. Western Blot was also used to assay the expression of IBA-1 in the whole hypothalamus. We found that the number of IBA-1-ir cells in the medial preoptic area was significantly reduced in EE compared to the SE rats (t = 5.704, *** p < 0.001) (Fig. 2a, d). In addition, the results obtained from Western-Blot assay also demonstrated that EE rats showed significantly reduced expression of IBA-1 in the whole hypothalamus compared to the SE rats (t = 3.910, **p < 0.01) (Fig. 2e, f).

Effect of early environmental enrichment on the expression of IBA-1 in the brain. a Representative image of IBA-1 expression in the hypothalamus (HY), b hippocampus (HIP), and c medial prefrontal cortex (mPFC). a Representative images of IBA-1 immunohistochemical staining in the hypothalamus of SE group. b Higher magnification of inset in (a). c Representative images of IBA-1 immunohistochemical staining in the EE group. d Higher magnification of inset in (c). e Representative images of IBA-1 immunohistochemical staining in the hippocampus of SE group. f Higher magnification of inset in (e). g Representative images of IBA-1 immunohistochemical staining in the EE group. h Higher magnification of inset in (g). i Representative images of IBA-1 immunohistochemical staining in the medial prefrontal cortex of SE group. j Higher magnification of inset in (a). k Representative images of IBA-1 immunohistochemical staining in the EE group. l Higher magnification of inset in (k). Scale bar 100 μm. d Immunohistochemical analysis of expression of IBA-1 in the medial preoptic area, hippocampus, and medial prefrontal cortex (n = 5 in SE, n = 5 in EE). e Representative bands are shown for each group, using GAPDH as the loading control (n = 6 in SE, n = 6 in EE). f Shows the densitometric analysis for IBA-1. **p < 0.01; ***p < 0.001 vs SE
In order to explore whether the inhibitory effect of environmental enrichment exposure on IBA-1 expression was specifically targeted to the hypothalamus, the expression of IBA-1 in the hippocampus and medial prefrontal cortex were determined by immunohistochemistry and Western blot assay. Interestingly, no difference was observed in the number of expressions of IBA-1-ir cells in the hippocampus (t = 1.003, p > 0.05) and medial prefrontal cortex (t = 1.846, p > 0.05) between the SE and EE groups, which was also verified by Western blot assay (hippocampus (t = 1.36, p > 0.05) and medial prefrontal cortex (t = 0.5094, p > 0.05) (Fig. 2d, f).
We also explored the effects of environmental enrichment on other cell types in the hypothalamus by probing for steady-state levels of astrocyte markers (GFAPs) and neuronal markers (NeuN) via western blot. Environmental enrichment did not affect the expression of NeuN (t = 2.3, p > 0.05) and GFAP in the hypothalamus (t = 0.7383, p > 0.05) (Fig. 3a, b), suggesting that environmental enrichment might specifically reduce the number of microglia in the hypothalamus.

a Representative bands are shown for each group, using GAPDH as the loading control (n = 6 in SE, n = 6 in EE). b Shows the densitometric analysis for NeuN and GFAP
Effect of microglia inhibitor minocycline on the social dominance tube test in SE rats
Since the microglia might be positively correlated to the percentage of forward in the social dominance tube test, we tested whether reduction of microglia in the SE group affects the percentage of forward in the social dominance tube test. After 3 days of minocycline treatment, the animals exhibited significant reduction in the percentage of forward, compared with their saline opponents (*p < 0.05) (Fig. 4a). Moreover, minocycline treatment effectively suppressed the expression of IBA-1 in the hypothalamus compared to saline-treated animals, as demonstrated by immunohistochemical staining (t = 4.321, ***p < 0.001) and Western blot assay (t = 4.286, *p < 0.05) (Fig. 4b–e).

Effect of minocycline treatment on the social dominance tube test in SE rats (a). MINO-treated rats showed remarkably reduced percent of forward in the social dominance tube test, n = 10 pairs. b Representative image illustrates IBA-1 expression in the hypothalamus (HY). a Representative images of IBA-1 immunohistochemical staining in the NS group. b Higher magnification of inset in (a). c Representative images of IBA-1 immunohistochemical staining in the MINO group. d Higher magnification of inset in (c). Scale bar:100 μm. c Immunohistochemical analysis of expression of IBA-1 in the hypothalamus (n = 5 in SE, n = 5 in EE). e Representative bands are shown for each group, using GAPDH as the loading control (n = 5 in SE, n = 5 in EE). f Shows the densitometric analysis for IBA-1. *p < 0.05; ***p < 0.001 vs NS (NS, normal saline-treated group; MINO, minocycline-treated group)
Effect of environmental enrichment on genes related to microglial proliferation, the level of enzymes involved in histone modifications, and status of DNA methylation in the hypothalamus
Depletion of the microglia in the hypothalamus by environmental enrichment or minocycline treatment led to reduced percent of forward in the social dominance tube test, suggesting that this decreased competitive success could be regulated by microglia in the hypothalamus. Previous study showed that microglia were dependent on specific survival factors, including transcription factor PU.1, IRF8, RUNX1, and DAP12 or CSF1R signaling (Chitu et al. 2016). No difference in the expression of PU.1 (t = 1.204, p > 0.05), IRF8 (t = 0.3160, p > 0.05), RUNX1 (t = 0.1299, p > 0.05), and DAP12 (t = 0.2512, p > 0.05) (Fig. 5a) was observed between the SE and EE groups. However, we observed that environmental enrichment led to significantly reduced expression of CSF-1 (t = 2.477, *p < 0.05) and IL-34 (t = 2.873, *p < 0.05) compared with SE. And, CSF1R was significantly enhanced in the EE than the SE (t = 2.319, *p < 0.05) (Fig. 5b). These results suggest that the reduced expression of CSF-1 or IL-34 might cause decreased expression of IBA-1 in the hypothalamus of EE rats.

Effect of environmental enrichment on the genes related to proliferation of microglia in the hypothalamus. a No difference was observed in the expression of PU.1, IRF8, RUNX1, and DAP12mRNA between the SE (n = 6) and EE (n = 6), p > 0.05. b Reduced expression of CSF-1 and IL-34 were found in the EE (n = 6) compared with SE (n = 6); EE led to increased expression of CSF1R compared with SE. c No difference in the expression of HDAC1, HDAC2, and HDAC6 mRNA between the SE (n = 6) and EE (n = 6); p > 0.05. d EE led to increased expression of DNMT1 compared with SE. Reduced expression of TET-1 was found in the EE (n = 6) compared with SE (n = 6); *p < 0.05, **p < 0.01 vs SE
Decreased gene expression can be caused by altered chromatin remodeling, which is known to be accomplished through post-translational modifications of the amino termini of histone tails at specific residues and/or by the addition of methyl groups by DNA methyltransferases (DNMTs) to the DNA occurring mostly at CpG sites (MacDonald and Roskams 2009). No difference was observed in the expression of HDAC1 (t = 0.3613, p > 0.05), HDAC2 (t = 1.718, p > 0.05), and HDAC6 (t = 0.3085, p > 0.05) (Fig. 5c) between these two groups. We detected a significant increase in the expression of Dnmt-1 in the EE group (t = 3.535, *p < 0.05). In addition, the level of the DNA demethylation enzyme Tet-1 was significantly reduced in the EE group, compared to SE (t = 3.798, * p < 0.01) (Fig. 5d).
Effects of DNA methyltransferase inhibitors on environmental enrichment-induced changes in CSF-1 gene expression and formation of social hierarchy
Since environmental enrichment caused significant changes in the level of Dnmt1, we tested whether transcriptional repression can be relieved to restore CSF-1 and IL-34 gene expression by the Dnmt blocker 5-AZ. We found that treatment with 5-AZ significantly reduced the environmental enrichment-induced upregulation of DNMT-1 gene (t = 0.168, p > 0.05) and restored CSF-1 (t = 1.231, p > 0.05) and IL-34 mRNA (t = 1.166, p > 0.05) (Fig. 6a) levels in hypothalamus. In addition, one-way ANOVA indicated that the number of IBA-1-positive cells in the hypothalamus was affected by the 5AZ treatment (F (2, 50) = 10.23, ***p < 0.001), and the post-hoc test indicated that environmental enrichment-induced reduction in the number of microglia was prevented (t = 0.5762, p > 0.05) (Fig. 6b–e). This result was also verified by Western blot assay, expression of IBA-1 in EE rats treated with 5-AZ was comparable to that of saline-treated SE rats (t = 0.5384, p > 0.05), while saline-treated EE rats still demonstrated significantly reduced expression of IBA-1 compared to saline-treated SE rats (t = 2.458, *p < 0.05) (Fig. 6f, g). We also observed that 5-AZ-treated EE rats exhibited comparative percentage of forward in the social dominance tube test compared to saline-treated SE rats (p > 0.05) (Fig. 6h). In order to exclude the potential stress effect of injection on the present result, another group of SE and EE rats were injected daily with saline, for 6 days. In line with the results mentioned before, EE with normal saline (NS) treatment still showed significantly reduced percentage of forward compared to their saline-treated SE opponents (*p < 0.05) (Fig. 6h). Moreover, the 5-AZ specifically restored the EE rats’ behavior in the social dominance tube test, since 5AZ treatment did not affect the percentage of forward in the SE rats (p > 0.05) (Fig. 6h). These data suggest that environmental enrichment-mediated DNA hypermethylation in the hypothalamus possibly results in reduction of CSF1 and IL-34 expression.

Effects of DNA methyltransferase inhibitors on environmental enrichment-induced changes in CSF-1 gene expression and formation of social hierarchy. a No difference was observed in the expression of DNTM1, CSF1, and IL34 mRNA between the SE + NS (n = 6) and EE + 5AZ (n = 6). b Representative images of IBA-1 immunohistochemical staining in the hypothalamus of SE + NS group. c Representative images of IBA-1 immunohistochemical staining in the EE + NS group. d Representative images of IBA-1 immunohistochemical staining in the EE + 5AZ group (e). Immunohistochemical analysis of expression of IBA-1 in the medial preoptic area (n = 5 in SE + NS, n = 5 in EE + NS; n = 5 in EE + 5AZ). f Representative bands are shown for each group, using GAPDH as the loading control (n = 5 in SE + NS, n = 5 in EE + NS; n = 5 in EE + 5AZ). g Shows the densitometric analysis for IBA-1. h EE + 5AZ showed remarkably increased percent of forward in the social dominance tube test (n = 13 pairs), while 5AZ treatment had no effect on the percent of forward in the SE rats (n = 6 pairs). EE + NS still exhibited significantly reduced percent of forward in the tube test compared with the SE + NS (n = 5 pairs). *p < 0.05; **p < 0.01; ***p < 0.001 vs SE + NS (NS normal saline treated group, 5AZ 5-Azacytidine treated group)
Effects of environmental enrichment on the competitive food foraging test and mate preference test
Although EE showed reduced percent of forward in the social dominance tube test, previous studies have shown that increased percent of forward in the social dominance tube test might reflect the aggressive behavior or abnormal social behavior. Thus, we also used two more behavioral tests (competitive food foraging test and mate preference test) to determine the animals’ social competitive behavior. However, in the competitive food foraging test, we found no difference in the total amount of food foraged between the SE and EE rats (t = 0.6837, p > 0.05) (Fig. 7a). In the mate preference test, female rats spent significantly more time with the EE rat compared to the SE rat (t = 2.354, * p < 0.05) (Fig. 7b). Thus, although EE rats exhibited lower social hierarchy in the social dominance tube test, they possess the normal competitive motivation and were more attractive to female rats, compared to the SE rats.

Effect of environmental enrichment on the competitive food foraging test and mate preference test. a No difference was observed in the total amount of food foraged between the SE and EE groups (n = 8 in each group). b Female rats were more attracted toward EE rats compared to SE rats in the mate preference test (n = 8 in each group). *p < 0.05 vs SE
Discussion
In this study, we examined the effects of exposure to enriched environment, an early positive stimulation procedure, on the behavior of adult rats in the social dominance tube test. We found that environmental enrichment led to reduced percent of forward in the social dominance tube test, while these rats showed comparable food foraging under competitive condition and were more attractive to female rats. In addition, we established that the number of microglia in the hypothalamus area might be causatively correlated with the outcome of the social dominance tube test. Moreover, the reduced number of microglia in the hypothalamus of EE might be regulated through the CSF1R signaling, which was modified by DNA methylation.
When the EE rats were confronted with their SE opponents in the social dominance tube test, they exhibited a significantly higher tendency to be submissive than SE rats. This reduction in the percent of forward could not be attributed to the difference in weights, since no difference in the weight was observed between the SE and EE groups. Moreover, the EE exhibited consistently lower percent of forward compared to the SE for three consecutive tests. In addition, we found that the experience of early environmental enrichment was essential for the clear winner and loser effect in the social dominance tube test, because no difference in the percent of forward was observed between the two SE opponents. Previous studies have suggested that environmental enrichment not only increases the rodents’ affiliative behaviors (McQuaid et al. 2013; Pietropaolo et al. 2004) but also compromises the animals’ social competitive behavior (Akers et al. 2008), which is in line with the present study. Since social competitive behavior is affected by corticosterone and testosterone level (Bartolomucci et al. 2001; van den Berg et al. 2015), we investigated whether differences in the neuroendocrine response might explain the low competitive success of EE rats in the social dominance tube test. However, in the present study, we did not find any difference in the baseline or post-competitive level of corticosterone and testosterone between the EE and SE groups, suggesting that the neuroendocrine response was comparable.
The observation that adult EE and SE rats differed in competitive success in the dominance tube test demonstrates that early exposure to enriched environment may have induced permanent changes in the brain. Microglia in the hypothalamus are indispensable for masculine behavior in rat (Lenz et al. 2013), but its role in the regulation of social dominance is still unknown. In the present study, we found that environmental enrichment led to reduced number of microglia specifically in the hypothalamus, while no effect was observed on other cell types. Moreover, no difference in the expression of IBA-1 was detected in other brain regions such as medial prefrontal cortex or hippocampus. We observed that treatment with microglial inhibitor minocycline not only reduced the number of microglia in the hypothalamic of SE rats but also resulted in reduced percent of forward compared to their saline-treated opponents, suggesting that the microglia in the hypothalamic might be positively correlated with the outcome of the social dominance tube test. Because genetic constitution did not differ systematically between EE and SE rats before the environmental enrichment stimulation, we concluded that these observed changes might be epigenetically induced. We identified that the reduced expression of IBA-1 in the hypothalamus of adult EE rats might be associated with altered expression of CSF-1 and IL-34. A previous study has indicated that CSF-1 and IL-34 played a key role in the regulation of microglial proliferation in adulthood (Chitu et al. 2016). We found that environmental enrichment increased DNMT-1 level but decreased the TET-1 level in hypothalamus, suggesting that the machinery that increased DNA methylation was enhanced in this brain region. Indeed, 5-AZ treatment increased mRNA levels of CSF1 and IL-34 significantly. In line with the restored expression of CSF1 and IL-34 in the EE group, the expression of IBA-1 was restored in the EE rats, and the animals showed comparative percent of forward with the SE opponents, suggesting that the number of microglia was indeed positively correlated with the outcome of the social dominance tube test. These data, as well as the data showing that 5-AZ treatment reverses the suppressive effects of environmental enrichment on CSF-1 and IL-34 gene expression, suggest the possibility that environmental enrichment increases DNA methylation to cause suppression of CSF-1 and IL-34 genes.
Whether social dominance tube test can be used as an index of general social dominance is still debatable. Social dominance tube test is correlated with barbering and urine-marking in male mice, but there are conflicting results on whether this test correlates with aggressive behavior or competition for access to food or water (Greenberg et al. 2014; Rodriguiz et al. 2004; Wang et al. 2011). In addition, social dominance tube test might be related to other non-hierarchical social behaviors, as abnormal dominance tube test behavior has been observed in many models that show depression-like behavior and impaired social behavior such as Rett syndrome or autism disorders (Filiano et al. 2013; Jiang-Xie et al. 2014; Moretti et al. 2005; Yang et al. 2015). Some studies have failed to find a correlation between tube test dominance and more ethologically relevant measures, such as competition for food or access to female mice (Greenberg et al. 2014; Rodriguiz et al. 2004). Thus, we used competitive food foraging test and mate preference test to determine the competitive behavior of the EE rats. In the competitive food foraging test, we observed no difference in the total amount food foraged by EE and SE rats, suggesting that the EE has comparable access to resources. In line with a previous study (Mitra and Sapolsky 2012), we found that rats exposed early to enriched environment were more attractive to females than the rats housed in standard environment, suggesting that EE might lead to greater mating opportunities. Previous studies have reported that the ability to know one’s social hierarchy under different conditions was beneficial for reducing aggressive behavior, which might avoid potential injury (Wang et al. 2014; Sapolsky 2005). The animals occasionally display aggressive behavior in the social dominance tube test, which might lead to injury. It is conceivable to consider that the reduced percent of forward displayed by EE rats, in fact, constitutes an appropriate response to confrontation with a dangerous opponent that might lead to injury. Thus, we infer that the reduced percent of forward exhibited by the EE rats might not reflect decreased social dominance. In fact, it might be a self-protection strategy under “body-contacted” competitive condition. We consider that early EE experience might facilitate the establishment of social hierarchy that reduces the potential injury to the rat.
In conclusion, we found that rats which were exposed to environmental enrichment exhibited greater ability to lose in social competition than were SE rats which were housed home cage and, remarkably, this decreased competitive success among EE rats accompanied by reduced number of microglia in hypothalamus. Furthermore, the effect of EE exposure on microglia in hypothalamus was regulated by the CSF1R signaling which was in turn modulated by status of DNA methylation. By understanding their proliferative function of microglia, we may begin to shed light on the significance of CSF-1R signaling in the adult brain under normal condition, independent of the recognized role in pathology. Further, our study demonstrates that it may turn out to be a dangerous oversimplification to distinguish social hierarchy based on the social dominance tube test.
References
Akers KG, Yang Z, DelVecchio DP, Reeb BC, Romeo RD, McEwen BS, Tang AC (2008) Social competitiveness and plasticity of neuroendocrine function in old age: influence of neonatal novelty exposure and maternal care reliability. PLoS One 3:e2840
Arai JA, Li S, Hartley DM, Feig LA (2009) Transgenerational rescue of a genetic defect in long-term potentiation and memory formation by juvenile enrichment. J Neurosci 29:1496–1502
Audet M-C, Mangano EN, Anisman H (2010) Behavior and pro-inflammatory cytokine variations among submissive and dominant mice engaged in aggressive encounters: moderation by corticosterone reactivity. Front Behav Neurosci 4:156
Barnum CJ, Blandino PJ, Deak T (2008) Social status modulates basal IL-1 concentrations in the hypothalamus of pair-housed rats and influences certain features of stress reactivity. Brain Behav Immun 22:517–527
Bartolomucci A, Palanza P, Gaspani L, Limiroli E, Panerai AE, Ceresini G, Poli MD, Parmigiani S (2001) Social status in mice: behavioral, endocrine and immune changes are context dependent. Physiol Behav 73:401–410
van den Berg WE, Lamballais S, Kushner SA (2015) Sex-specific mechanism of social hierarchy in mice. Neuropsychopharmacology 40:1364–1372
Buchhold B, Mogoanta L, Suofu Y, Hamm A, Walker L, Kessler C, Popa-Wagner A (2007) Environmental enrichment improves functional and neuropathological indices following stroke in young and aged rats. Restor Neurol Neurosci 25:467–484
Cao WY, Xu Y, Luo YW, Zhong XL, Duan J, Wang XQ, Hu ZL, Li F, Dai RP, Luo XG, Zhang JY, Li CQ (2013) Activation of ERK1/2 is required for normal response of isosexual social interactions in male rats. Brain Res 1538:51–60
Cao W, Duan J, Wang X, Zhong X, Hu Z, Huang F, Wang H, Zhang J, Li F, Zhang J, Luo X, Li CQ (2014) Early enriched environment induces an increased conversion of proBDNF to BDNF in the adult rat's hippocampus. Behav Brain Res 265:76–83
Chitu V, Gokhan S, Nandi S, Mehler MF, Stanley ER (2016) Emerging roles for CSF-1 receptor and its ligands in the nervous system. Trends Neurosci 39:378–393
Crews D, Gore AC, Hsu TS, Dangleben NL, Spinetta M, Schallert T, Anway MD, Skinner MK (2007) Transgenerational epigenetic imprints on mate preference. Proc Natl Acad Sci U S A 104:5942–5946
Dai RP, Li CQ, Zhang JW, Li F, Shi XD, Zhang JY, Zhou XF (2011) Biphasic activation of extracellular signal-regulated kinase in anterior cingulate cortex distinctly regulates the development of pain-related anxiety and mechanical hypersensitivity in rats after incision. Anesthesiology 115:604–613
Elmore MR, Najafi AR, Koike MA, Dagher NN, Spangenberg EE, Rice RA, Kitazawa M, Matusow B, Nguyen H, West BL, Green KN (2014) Colony-stimulating factor 1 receptor signaling is necessary for microglia viability, unmasking a microglia progenitor cell in the adult brain. Neuron 82:380–397
Erny D, Hrabe DAA, Jaitin D, Wieghofer P, Staszewski O, David E, Keren-Shaul H, Mahlakoiv T, Jakobshagen K, Buch T, Schwierzeck V, Utermohlen O, Chun E, Garrett WS, McCoy KD, Diefenbach A, Staeheli P, Stecher B, Amit I, Prinz M (2015) Host microbiota constantly control maturation and function of microglia in the CNS. Nat Neurosci 18:965–977
Filiano AJ, Martens LH, Young AH, Warmus BA, Zhou P, Diaz-Ramirez G, Jiao J, Zhang Z, Huang EJ, Gao FB, Farese RJ, Roberson ED (2013) Dissociation of frontotemporal dementia-related deficits and neuroinflammation in progranulin haploinsufficient mice. J Neurosci 33:5352–5361
Garfield AS, Cowley M, Smith FM, Moorwood K, Stewart-Cox JE, Gilroy K, Baker S, Xia J, Dalley JW, Hurst LD, Wilkinson LS, Isles AR, Ward A (2011) Distinct physiological and behavioural functions for parental alleles of imprinted Grb10. Nature 469:534–538
Greenberg GD, Howerton CL, Trainor BC (2014) Fighting in the home cage: agonistic encounters and effects on neurobiological markers within the social decision-making network of house mice (Mus musculus). Neurosci Lett 566:151–155
Jiang-Xie LF, Liao HM, Chen CH, Chen YT, Ho SY, Lu DH, Lee LJ, Liou HH, Fu WM, Gau SS (2014) Autism-associated gene Dlgap2 mutant mice demonstrate exacerbated aggressive behaviors and orbitofrontal cortex deficits. Mol Autism 5:32
Kato TA, Watabe M, Tsuboi S, Ishikawa K, Hashiya K, Monji A, Utsumi H, Kanba S (2012) Minocycline modulates human social decision-making: possible impact of microglia on personality-oriented social behaviors. PLoS One 7:e40461
Kettenmann H, Hanisch UK, Noda M, Verkhratsky A (2011) Physiology of microglia. Physiol Rev 91:461–553
Lenz KM, Nugent BM, Haliyur R, McCarthy MM (2013) Microglia are essential to masculinization of brain and behavior. J Neurosci 33:2761–2772
Li F, Cao WY, Li MB, Xu Y, Zhang JW, Zhang JY, Luo XG, Dai RP, Zhou XF, Li CQ (2012) A simple method for detection of food foraging behavior in the rat: involvement of NMDA and dopamine receptors in the behavior. Neuroscience 205:73–80
Li S, Jin M, Zhang D, Yang T, Koeglsperger T, Fu H, Selkoe DJ (2013) Environmental novelty activates beta2-adrenergic signaling to prevent the impairment of hippocampal LTP by Abeta oligomers. Neuron 77:929–941
Li CQ, Luo YW, Bi FF, Cui TT, Song L, Cao WY, Zhang JY, Li F, Xu JM, Hao W, Xing XW, Zhou FH, Zhou XF, Dai RP (2014) Development of anxiety-like behavior via hippocampal IGF-2 signaling in the offspring of parental morphine exposure: effect of enriched environment. Neuropsychopharmacology 39:2777–2787
Lores-Arnaiz S, Bustamante J, Arismendi M, Vilas S, Paglia N, Basso N, Capani F, Coirini H, Costa JJ, Arnaiz MR (2006) Extensive enriched environments protect old rats from the aging dependent impairment of spatial cognition, synaptic plasticity and nitric oxide production. Behav Brain Res 169:294–302
MacDonald JL, Roskams AJ (2009) Epigenetic regulation of nervous system development by DNA methylation and histone deacetylation. Prog Neurobiol 88:170–183
McQuaid RJ, Audet MC, Jacobson-Pick S, Anisman H (2013) Environmental enrichment influences brain cytokine variations elicited by social defeat in mice. Psychoneuroendocrinology 38:987–996
Miller LS, Colella B, Mikulis D, Maller J, Green RE (2013) Environmental enrichment may protect against hippocampal atrophy in the chronic stages of traumatic brain injury. Front Hum Neurosci 7:506
Mitra R, Sapolsky RM (2012) Short-term enrichment makes male rats more attractive, more defensive and alters hypothalamic neurons. PLoS One 7:e36092
Moretti P, Bouwknecht JA, Teague R, Paylor R, Zoghbi HY (2005) Abnormalities of social interactions and home-cage behavior in a mouse model of Rett syndrome. Hum Mol Genet 14:205–220
Nithianantharajah J, Hannan AJ (2006) Enriched environments, experience-dependent plasticity and disorders of the nervous system. Nat Rev Neurosci 7:697–709
Patel S, Player MR (2009) Colony-stimulating factor-1 receptor inhibitors for the treatment of cancer and inflammatory disease. Curr Top Med Chem 9:599–610
Pearse DD, Hughes ZA (2016) PDE4B as a microglia target to reduce neuroinflammation. Glia 64:1698–1709
Pietropaolo S, Branchi I, Cirulli F, Chiarotti F, Aloe L, Alleva E (2004) Long-term effects of the periadolescent environment on exploratory activity and aggressive behaviour in mice: social versus physical enrichment. Physiol Behav 81:443–453
Rodriguiz RM, Chu R, Caron MG, Wetsel WC (2004) Aberrant responses in social interaction of dopamine transporter knockout mice. Behav Brain Res 148:185–198
Sales AJ, Biojone C, Terceti MS, Guimaraes FS, Gomes MV, Joca SR (2011) Antidepressant-like effect induced by systemic and intra-hippocampal administration of DNA methylation inhibitors. Br J Pharmacol 164:1711–1721
Sapolsky RM (2005) The influence of social hierarchy on primate health. Science 308:648–652
Shahbazian M, Young J, Yuva-Paylor L, Spencer C, Antalffy B, Noebels J, Armstrong D, Paylor R, Zoghbi H (2002) Mice with truncated MeCP2 recapitulate many Rett syndrome features and display hyperacetylation of histone H3. Neuron 35:243–254
Spencer CM, Alekseyenko O, Serysheva E, Yuva-Paylor LA, Paylor R (2005) Altered anxiety-related and social behaviors in the Fmr1 knockout mouse model of fragile X syndrome. Genes Brain Behav 4:420–430
Suarez-Roca H, Quintero L, Avila R, Medina S, De Freitas M, Cardenas R (2014) Central immune overactivation in the presence of reduced plasma corticosterone contributes to swim stress-induced hyperalgesia. Brain Res Bull 100:61–69
Wang F, Zhu J, Zhu H, Zhang Q, Lin Z, Hu H (2011) Bidirectional control of social hierarchy by synaptic efficacy in medial prefrontal cortex. Science 334:693–697
Wang F, Kessels HW, Hu H (2014) The mouse that roared: neural mechanisms of social hierarchy. Trends Neurosci 37:674–682
Watabe M, Kato TA, Monji A, Horikawa H, Kanba S (2012) Does minocycline, an antibiotic with inhibitory effects on microglial activation, sharpen a sense of trust in social interaction? Psychopharmacology 220:551–557
Yang CR, Bai YY, Ruan CS, Zhou HF, Liu D, Wang XF, Shen LJ, Zheng HY, Zhou XF (2015) Enhanced aggressive behaviour in a mouse model of depression. Neurotox Res 27:129–142
Zhan Y, Paolicelli RC, Sforazzini F, Weinhard L, Bolasco G, Pagani F, Vyssotski AL, Bifone A, Gozzi A, Ragozzino D, Gross CT (2014) Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior. Nat Neurosci 17:400–406
Acknowledgements
This research was supported by the National Natural Science Foundation of China (31371212 to Chang-Qi Li, 81471372 to Fang Li) and the Graduate Research and Innovation Projects of Central South University (2016zzts116 to Zhao-Lan Hu).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Electronic supplementary material
ESM 1
(DOC 49 kb).
Rights and permissions
About this article

Cite this article
Cao, WY., Hu, ZL., Xu, Y. et al. Role of early environmental enrichment on the social dominance tube test at adulthood in the rat. Psychopharmacology 234, 3321–3334 (2017). https://doi.org/10.1007/s00213-017-4717-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-017-4717-3