
Abstract
Rationale
Tobacco smoking is associated with dysregulated reward processing within the striatum, characterized by hypersensitivity to smoking rewards and hyposensitivity to non-smoking rewards. This bias toward smoking reward at the expense of alternative rewards is further exacerbated by deprivation from smoking, which may contribute to difficulty maintaining abstinence during a quit attempt.
Objective
We examined whether abstinence-induced changes in striatal processing of rewards predicted lapse likelihood during a quit attempt supported by contingency management (CM), in which abstinence from smoking was reinforced with money.
Methods
Thirty-six non-treatment-seeking smokers participated in two functional MRI (fMRI) sessions, one following 24-h abstinence and one following smoking as usual. During each scan, participants completed a rewarded guessing task designed to elicit striatal activation in which they could earn smoking and monetary rewards delivered after the scan. Participants then engaged in a 3-week CM-supported quit attempt.
Results
As previously reported, 24-h abstinence was associated with increased striatal activation in anticipation of smoking reward and decreased activation in anticipation of monetary reward. Individuals exhibiting greater decrements in right striatal activation to monetary reward during abstinence (controlling for activation during non-abstinence) were more likely to lapse during CM (p < 0.025), even when controlling for other predictors of lapse outcome (e.g., craving); no association was seen for smoking reward.
Conclusions
These results are consistent with a growing number of studies indicating the specific importance of disrupted striatal processing of non-drug reward in nicotine dependence and highlight the importance of individual differences in abstinence-induced deficits in striatal function for smoking cessation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Despite widespread and longstanding public health efforts, tobacco smoking remains a leading cause of preventable death in the developed world (CDC 2011). As many as 70 % of smokers wish to quit, but less than 5 % of quit attempts result is abstinence for more than 3 months (CDC 2002). Identifying mechanisms contributing to lapse and relapse among chronic smokers is critical for improving treatment and prevention efforts and may ultimately improve interventions for those at greatest risk for relapse.
Disordered reward processing within the mesolimbic dopamine system has been repeatedly implicated in addictive disorders, including nicotine dependence, and may be an important contributor to relapse. According to incentive sensitization theory, repeated exposure to drugs of abuse, including nicotine, results in sensitization of the mesolimbic response to drug-related cues, ultimately conferring heightened motivational incentive properties to these stimuli and leading to increased drug-seeking behavior (Robinson and Berridge 1993). At the same time, opponent process theory posits that chronic drug exposure contributes to compensatory alterations in reward processing (Koob and Le Moal 2005). causing non-drug rewards to lose their incentive value and thus fail to motivate behavior. Human neuroimaging studies comparing drug abusers to controls have consistently demonstrated heightened blood oxygenation level-dependent (BOLD) response to drug-related vs. neutral stimuli in key reward-related regions, including medial prefrontal cortex (mPFC), orbitofrontal cortex (OFC), anterior cingulate cortex (ACC), and striatum among individuals dependent on cocaine, alcohol, and nicotine (David et al. 2005; Franklin et al. 2007; Goldstein et al. 2009; Heinz et al. 2004; McClernon et al. 2005; Wrase et al. 2002). At the same time, decrements in response to non-drug rewards and positive affective stimuli have been found in the mPFC and striatum among individuals dependent on cocaine (Garavan et al. 2000; Goldstein et al. 2007; Volkow et al. 1993). opiates (Martin-Soelch et al. 2001), alcohol (Beck et al. 2009; Wrase et al. 2007). and nicotine (Martin-Soelch et al. 2003; Peters et al. 2011; Rose et al. 2012). Together, these findings suggest that chronic smoking is associated with a bias in mesolimbic processing toward smoking reward at the expense of alternative rewards that may contribute to a narrowing of the behavioral repertoire, such that individuals increasingly choose drug over other non-drug reward alternatives.
Importantly, underlying reward dysregulation may be amplified by abstinence from smoking, leading to heightened vulnerability to relapse. Evidence from animal studies suggests that acute nicotine enhances—while withdrawal from nicotine attenuates—the incentive value of other reinforcers (Besheer and Bevins 2003; Chaudhri et al. 2006; Donny et al. 2003; Thiel et al. 2009; Weaver et al. 2012). Abstinent smokers demonstrate increased desire to smoke (i.e., craving) (Shiffman and Jarvik 1976). along with diminished interested in pleasant events, evidenced by decreased attentional bias toward positive stimuli (Dawkins et al. 2006; Freeman et al. 2012; Leventhal et al. 2012; Powell et al. 2002b; Powell et al. 2004). lower ratings of attractiveness for unfamiliar faces (Attwood et al. 2009). lower ratings of anticipated enjoyment in activities; and reduced responsiveness to monetary reward (Dawkins et al. 2006; Geier et al. 2014; Powell et al. 2002a; Powell et al. 2004). Consistent with behavioral evidence, we recently demonstrated that 24-h abstinence from smoking—compared with smoking as usual—was associated with both heightened response to smoking reward (i.e., puffs of a cigarette delivered after the scan) and attenuated response to monetary reward within the ventral striatum (Sweitzer et al. 2013b)—a key component of reward circuitry thought to encode motivational salience of primary and secondary reinforcers (Bassareo and Di Chiara 1999; Delgado et al. 2000; O’Doherty 2004; Schultz 2000). Furthermore, individual differences were observed in magnitude of abstinence-induced changes in reward processing, suggesting that some smokers may be more vulnerable to these effects and, potentially, at greater risk for resumption of smoking.
Thus, abstinence-induced dysregulation of reward processing may represent an important mechanism contributing to the choice to smoke in lieu of alternative reinforcement during a quit attempt. In support of this hypothesis, Wilson and colleagues recently found that blunted ventral striatal response to monetary outcomes during a functional MRI (fMRI) reward paradigm predicted higher likelihood of smoking during a laboratory delay of smoking reinforcement task (Wilson et al. 2014). However, findings were limited by a restricted range of smoking outcomes (i.e., only five participants chose to smoke during the reinforcement task), lack of a smoking as usual condition, and examination of only one reward type. Furthermore, it is unclear whether results extend beyond the laboratory setting to a real-world quit attempt.
In the present study, we evaluated whether abstinence-induced changes in striatal responsivity to smoking and monetary rewards, previously reported for this sample, predicted smoking lapse outcomes during a 3-week quit attempt supported by contingency management (CM). CM, in which smokers are provided with monetary incentives for periods of biochemically verified abstinence, has been widely demonstrated to be effective for improving abstinence rates, particularly while incentives remain in place (Alessi et al. 2004; Dallery et al. 2007; Dunn et al. 2008; Higgins et al. 2004; Roll and Higgins 2000; Roll et al. 1996; Tidey et al. 2002). Importantly, the procedure provides a real-world corollary for assessing the trade-off between monetary and smoking rewards. We focused our investigation on the striatum because this region is central to reward processing, activation within the striatum is strongly elicited by the rewarded guessing task used here, and previously identified dissociated effects of abstinence on anticipation of smoking and monetary reward within this sample were restricted to this region. We hypothesized that smokers exhibiting greater abstinence-induced dysregulation in striatal processing of rewards, characterized by increased responsivity to smoking reward and decreased responsivity to monetary reward, would be most likely to lapse during CM.
Methods and materials
Participants
Fifty-six daily smokers were recruited as part of a study investigating genetic predictors of abstinence during a quit attempt. Inclusion criteria included being between 18 and 65 years old, smoking five or more cigarettes per day during the past year, expired carbon monoxide (CO) level of ≥8 ppm, and willingness to make a quit attempt. Exclusion criteria included self-reported psychiatric illness or significant medical illness in the past year, current heavy drug or alcohol use as determined by self-report (drug use for 10+ days or four or more drinks per day for 10+ days in past 30) or positive urine drug screen or blood alcohol >0.08, current use of any psychotropic medication or other tobacco products, interest in using smoking cessation medications, pregnancy/lactation, head trauma with loss of consciousness in the past year, claustrophobia, and any known conditions putting them at risk from exposure to high-field strength magnetic fields. Because the parent study involved a genetic component, participation was restricted to Caucasians to minimize population stratification. All participants provided informed consent as approved by the University of Pittsburgh Institutional Review Board.
Out of 56 individuals initially recruited, 11 withdrew from the study prior to completion of fMRI sessions (non-attendance, n = 5; inability to achieve abstinence for fMRI session, n = 3; alcohol intoxication, n = 1; claustrophobia, n = 1; size constraints of scanner, n = 1) and 9 were excluded from analyses due to technical issues. The 36 participants included in analyses ranged from 18 to 58 years of age (M = 34.1 years, SD = 12.4); 53 % were female. Participants smoked between 5 and 30 cigarettes per day (M = 13.6, SD = 6.2) and had a mean score of 3.5 (SD = 2.4) on the Fagerström Test for Nicotine Dependence (FTND) (Heatherton et al. 1991). Excluded participants smoked significantly more cigarettes per day [t(54) = 2.16, p < 0.05] but did not differ on the FTND. Only two participants endorsed illicit drug use in the past 30 days (marijuana and hallucinogens, on two occasions). Results were unchanged when these individuals were excluded from analyses.
Procedure
Screening session
Eligible participants attended an in-person screening. Breath and urine samples were collected to assess blood alcohol level and illicit drug use and pregnancy, respectively. Expired CO was assessed upon arrival and after smoking a cigarette in the laboratory. To prevent exclusion of participants who had not smoked recently prior to the visit, the minimum CO inclusion criterion was satisfied if either CO sample was greater than 8 ppm. Participants completed questionnaires assessing demographics, medical and psychiatric history, nicotine use history, and nicotine dependence (FTND).
Functional MRI sessions
Participants completed two identical fMRI sessions on two different days (mean of 12.8 days apart), once following smoking ad libitum (non-abstinent) and once after 24-h abstinence (abstinent). Session order was randomly assigned; 16 participants (44.4 %) completed the abstinent session first. Abstinence was verified via self-report and expired CO <8 ppm or 50 % reduction from baseline. Prior to each scan, participants underwent task training and completed subjective measures, including a four-item version of the Questionnaire on Smoking Urges (QSU-4; Toll et al. 2006) and the Minnesota Withdrawal Scale (MNWS; Hughes and Hatsukami 1986). The QSU-4 was repeated immediately after the scans, which lasted approximately 1 h. During the non-abstinent session, participants smoked a cigarette immediately prior to the scan to prevent unintended withdrawal.
Contingency management
Following completion of both fMRI sessions, participants made a smoking quit attempt using an adapted internet-based CM procedure (Dallery et al. 2007; Glenn and Dallery 2007). allowing for mobile biochemical verification of smoking abstinence. Participants were provided with computer equipment, including a netbook with webcam and a Bedfont piCO+ CO monitor (http://bedfont.com/smokerlyzer), and were instructed on how to use this equipment. Participants selected a quit date and were provided with brief smoking cessation counseling prior to beginning the procedure. During CM, participants were instructed to log in to the study website and record and submit CO samples twice daily (at least 8 h apart) throughout a 3-day baseline period and subsequent 18-day quit attempt. During the baseline period, participants smoked ad libitum and were paid $3.00 for each submitted sample; during the abstinence incentive period, only samples meeting criteria for abstinence (self-reported abstinence and CO <8 ppm or a 30 % drop from the previous sample) were reinforced. In cases where technical problems prevented video verification (5.5 % of total samples), self-reported abstinence was accepted. Reinforcement was provided according to an ascending schedule: participants received $3.00 for the first abstinent sample, and this amount increased by $0.25 for each consecutive abstinent sample. Participants also received a $5.00 bonus for every third consecutive abstinent sample. Non-abstinent samples were not reinforced and resulted in resetting of the payment schedule to $3.00. Following a lapse, reinforcement was returned to the highest previously earned level after submission of three consecutive abstinent samples. Missing samples (7.0 % of total) were not reinforced, but data were imputed based on self-report and surrounding samples for the purpose of analyses (i.e., missing samples were coded as abstinent when preceded and followed by four abstinent samples and <2-ppm increase in CO was observed following the missing sample). After achieving an initial 24-h abstinence following the quit date (i.e., two consecutive abstinent samples), participants who self-reported smoking or submitted a CO sample above the required cutoff were deemed to have lapsed. These criteria for classification of lapsers versus abstainers are consistent with previous studies and the Society for Research on Nicotine and Tobacco’s working group definition of a slip (Hughes et al. 2003; Janes et al. 2010). Subsequent analyses were aimed at differentiating those who lapsed from those maintaining continuous abstinence. In cases where coding of missing samples affected lapse classification (n = 1), analyses were rerun with that participant dropped from analyses.
Functional neuroimaging
Reward guessing task
During each fMRI scan, participants completed a rewarded guessing task (Fig. 1), during which they could earn monetary reward and puffs of a cigarette by “guessing” whether a computer-generated number would be higher or lower than 5. This task has been previously described in detail, and effects of abstinence on striatal response to monetary and puff rewards within this sample were previously reported (Sweitzer et al. 2013b). Briefly, for each trial, participants first indicated their guess with a button press and were then presented with a picture representation of the type of reward that they could earn on that trial ($0.50, a puff of a cigarette, or nothing). After the 6-s anticipation phase, the actual number was presented, followed by win or no-win feedback and a 9-s intertrial interval. The task was divided into four runs of 18 trials each, with each run lasting 6 min, 10 s. Rewards ($6 and 12 puffs) were delivered during a 1-h waiting period after the scan, during which participants were required to abstain from smoking cigarettes other than the puffs earned in the scanner.

Schematic of events within each trial of the fMRI reward task. Trials began with a 4-s presentation of a question mark, during which participants indicated their guess (higher or lower than 5) via button press. Next, an image was presented for 6 s depicting the reward that would be won if the guess was correct (stack of dollar bills = $0.50, cigarette = a puff of a cigarette, or blue notebook = no reward). The “actual” number was then presented for 500 ms, followed by 500-ms win or no-win feedback (up arrow or yellow circle, respectively). The feedback phase was followed by a 9-s intertrial interval (ITI), marked by a white fixation cross [reprinted from Sweitzer et al. 2013b, with permission from Elsevier]
Imaging acquisition and preprocessing
BOLD functional images were acquired using a 3.0-T Siemens Allegra scanner (Erlangen, Germany) with gradient EPI sequence covering 34 interleaved axial slices of 3-mm thickness with the following parameters: TR = 2 s; TE = 29 ms; flip angle = 90°; 64 × 64 matrix with FOV = 20 × 20 cm. A T1-weighted structural image was acquired using a three-dimensional volume MPRAGE pulse sequence covering 176 axial slices of 1-mm thickness. Image preprocessing was conducted using SPM8 (www.fil.ion.ucl.ac.uk/spm). Images were corrected for slice timing effects and realigned to the mean functional image. Structural images were segmented into native space grey matter and coregistered to the mean of functional images. Functional images were spatially normalized into Montreal Neurologic Institute stereotactic space and resampled to a voxel size of 2 × 2 × 2 mm. Images were smoothed with a Gaussian filter set at 6-mm full-width at half maximum (FWHM), and a high-pass filter (128 s) was applied. Artifact detection tools implemented in SPM were used to identify and adjust for image artifacts related to intensity spiking and motion as previously reported (Sweitzer et al. 2013b).
Data analysis
As in our previous report which found habituation of the striatum to monetary reward anticipation during the second half of the task (Sweitzer et al. 2013b) and consistent with prior research (Forbes et al. 2012; Forbes et al. 2009; Forbes et al. 2010; Forbes et al. 2011). analyses were restricted to the first two runs of the task to minimize habituation effects. In our previous report, first-level contrasts for the reward anticipation phase (money > neutral and puff > neutral) were submitted to a second-level random effects 2 (reward type) × 2 (abstinent vs non-abstinent condition) × 2 (scan order) ANOVA. We previously reported a significant reward type × abstinence interaction within a priori regions of interest (ROIs) of the ventral and dorsal striatum, such that abstinence was associated with heightened anticipatory activation to smoking reward and attenuated anticipatory activation to monetary reward (Sweitzer et al. 2013b). Here, we extracted the first eigenvariate from these previously identified clusters (encompassing 169 and 121 voxels for the right and left caudate head, respectively; Fig. 2) showing a significant reward type by abstinence condition interaction. This process yielded an estimate of the overall activation across voxels within each ROI (i.e., left and right striatum) for anticipation of each reward type (money, puffs) within each abstinence condition (abstinent, non-abstinent). Extracted values were exported to SPSS, and logistic regression was used to examine whether individual differences in striatal activation during abstinence, relative to non-abstinence, were predictive of lapse outcomes. Age and sex were included as covariates for all analyses.

Functional regions of interest within the right (169 voxels) and left (121 voxels) striatum exhibiting a significant reward type by abstinence condition interaction. Panel shown at x = 8
Results
Contingency management outcomes
All 36 participants achieved an initial period of 24-h abstinence following the quit date. Thirteen (36.1 %) subsequently maintained continuous abstinence throughout the procedure (including one participant for whom a missing data point was imputed as abstinent); the remaining 23 (63.9 %) lapsed during the quit attempt. Demographic predictors of lapse are presented in Table 1. Age, sex, and FTND were not significant predictors of lapse during CM. There was a trend toward smoking higher cigarettes per day associated with greater likelihood of lapse, but this difference did not reach statistical significance [odds ratio (OR) = 1.138, p = 0.06, 95 % confidence interval (CI) 0.993–1.305].
Subjective effects
As previously reported for this sample, smoking abstinence was associated with significant increases in self-reported craving (t = 6.5, p < 0.001) and withdrawal (t = 3.5, p < 0.005). During 24-h abstinence, higher levels of self-reported craving and withdrawal were positively correlated with anticipatory activation to puff reward in both the right caudate (r = 0.387 and 0.384, respectively, both ps < 0.05) and the left caudate (r = 0.402 and 0.411, respectively, both ps < 0.05). No associations were seen for monetary reward or during the non-abstinent session. We further examined whether severity of craving and withdrawal during abstinence, controlling for non-abstinent levels, were predictive of lapse outcomes. Contrary to expectations, higher craving at 24-h abstinence was associated with significantly lower likelihood of lapse (OR = 0.957, p < 0.05, CI 0.918–0.997); withdrawal was unrelated to lapse. Results were unchanged when the participant with missing data was dropped from analyses.
Neuroimaging predictors of lapse outcomes
Extracted values for money and puff anticipation during abstinent and satiated conditions from right and left caudate were entered into logistic regression models to evaluate their association with lapse outcome, with separate models created for each hemisphere. For each model, activation to smoking and monetary rewards during non-abstinence were entered during a first step to examine baseline differences between lapsers and abstainers in a satiated state. Activation to each reward type during abstinence was added in the next step to examine abstinence-induced changes in activation as predictors of lapse outcomes. When the addition of a new step significantly improved the overall model, specific predictors (i.e., monetary or smoking reward) were then examined to determine their relative contributions. Significance was set to p < 0.025 to control for multiple comparisons across two models (right and left striatum).
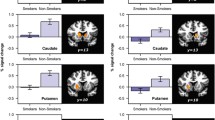
For all measures, activation during the non-abstinent condition was not significantly associated with lapse. In the right caudate, activation during abstinence significantly predicted lapse outcomes (χ 2 = 1.02, p < 0.025). Specifically, blunted anticipatory activation to monetary reward during abstinence relative to non-abstinence predicted higher likelihood of lapse (OR = 0.038, p < 0.025, CI 0.003–0.534) (Fig. 3). This corresponds to a standardized odds ratio of 4.18, indicating that a one standard deviation decrease in anticipatory reward activation is associated with an approximately fourfold increase in likelihood of lapse. This association remained significant even when controlling for behavioral predictors (i.e., abstinence-induced craving and cigarettes per day). Indeed, when behavioral and neuroimaging measures were included in the model, both craving and monetary reward anticipation during abstinence were significant predictors of lapse (both ps < 0.05). Follow-up pairwise comparisons indicated that individuals who lapsed during CM exhibited a significant decrease in anticipatory activation to monetary reward during abstinence relative to non-abstinence in the right VS (t = 4.6, p < 0.001), while no change was observed for non-lapsers (t = 0.4, ns). Abstinence-induced changes in anticipatory activation for monetary reward in the left caudate or for puff reward in the right or left caudate were not significantly predictive of lapse (all ps > 0.10). Results were unchanged when the subject with missing data was dropped from analyses.

Eigenvariate for anticipatory activation to monetary (a, b) and puff (c, d) reward extracted from clusters in left and right ventral striatum, plotted by lapse status and condition. Striped bars represent satiated condition. Error bars reflect standard error of the mean. Single asterisk indicates significant difference at p < 0.01; Double asterisks indicate significant difference at p < 0.001
Discussion
The present study examined associations between abstinence-induced changes in striatal reward processing and the ability to maintain abstinence during a quit attempt supported by CM. We hypothesized that individuals who exhibited greater blunting of striatal response to monetary reward and heightened response to smoking reward during abstinence relative to satiation would be most likely to lapse despite monetary incentives for abstinence. Our hypothesis was partially supported: we found that an attenuated response to monetary reward anticipation in the right caudate during abstinence predicted greater likelihood of lapse during CM, while response to smoking reward was not predictive of lapse outcomes.
Striatal processing of non-drug rewards has been increasingly implicated in nicotine dependence, with several human neuroimaging studies showing decreased activation among smokers relative to non-smokers or occasional smokers (Buhler et al. 2010; Lessov-Schlaggar et al. 2013; Luo et al. 2011; Martin-Soelch et al. 2003; Peters et al. 2011; Rose et al. 2012) or altered functioning during abstinence relative to satiation (Addicott et al. 2012; Rose et al. 2013; Sweitzer et al. 2013b). but see also (Buhler et al. 2010). The present results extend our previous findings for this dataset and suggest that individual differences in abstinence-induced changes in striatal function have important implications for real-world smoking behavior and cessation outcomes. Given the central role of the ventral striatum in salience attribution and subjective valuation of rewards (Kahnt et al. 2010; Knutson et al. 2001; O’Doherty et al. 2004), smokers attempting to quit may find themselves increasingly motivated to smoke against a backdrop of declining motivation for other rewards. Interestingly, although our previous work demonstrated a main effect of smoking abstinence on striatal activation to monetary reward anticipation, our current findings indicate that this effect was not observed among the subset of smokers (33 %) who were able to maintain continuous abstinence during CM. This suggests that intact reward processing during withdrawal may be a protective factor for smoking cessation, particularly in the context of reinforcement for abstinence, and highlights the importance of individual differences in mechanisms maintaining smoking behavior that may impact treatment response.
The present findings are consistent with a small number of recent studies that have examined associations between reactivity to affective or rewarding stimuli and ability to abstain from smoking. Wilson and colleagues similarly observed attenuated striatal response to monetary reward among abstinent smokers who subsequently smoked during a delay of smoking reinforcement task compared with those who abstained throughout the entire task (Wilson et al. 2014). Another recent study examined neural response to images of emotional and cigarette cues as a predictor of quit outcome among smokers interested in quitting (Versace et al. 2014). Smokers classified as low in reward sensitivity (based on lower reactivity to pleasant versus cigarette-related stimuli across several cortical regions) were significantly less likely to be abstinent from smoking at 6 months post-quit. Although initial classification of reward sensitivity was based on findings of attenuated activation to pleasant stimuli relative to cigarette cues in regions of the extended visual system, the authors also observed a similar pattern of lower reactivity to positive versus smoking stimuli within the dorsal striatum among low reward sensitivity smokers. The current results provide further support for the importance of blunted reward sensitivity as a risk factor for relapse and highlight the moderating effect of abstinence state. Importantly, associations between striatal functioning and lapse outcomes were not evident during satiation, suggesting that early abstinence may be a particular period of vulnerability. Additional work is needed to determine whether deficits in reward functioning emerging early in a quit attempt may improve with extended abstinence.
Contrary to expectations, heightened striatal activation to smoking reward during abstinence did not predict lapse outcome. This was surprising given the robust increases in activation observed at the group level (Sweitzer et al. 2013b) and the putative role of striatal activation as an index of motivational salience of smoking reward, and in light of previous studies demonstrating an association between reactivity to smoking or alcohol-related cues and quit outcomes (Grusser et al. 2004; Janes et al. 2010). However, methodological differences in task design and timing of fMRI assessment may have contributed to discrepant findings, as these prior studies utilized passive viewing block design tasks and did not examine a withdrawal state. The lack of a significant effect in the present study was not simply due to a ceiling effect restricting the range of meaningful individual differences, as variability in striatal activation to smoking reward during abstinence was positively correlated with several subjective measures, including self-reported craving, withdrawal, negative affect, and diminished positive affect. Thus, while heightened striatal activation to smoking reward did correspond to a subjective experience of increased desire to smoke, this did not translate to increased smoking behavior during CM. Interestingly, higher craving during the abstinence session was actually associated with lower likelihood of lapse during CM. This is in contrast with several studies that have demonstrated associations between increased craving and poorer abstinence outcomes in laboratory-based studies and smoking cessation trials (Bold et al. 2013; Sweitzer et al. 2013a; Van Zundert et al. 2012) but see also (Wray et al. 2013). Thus, it is possible that the availability of monetary incentives for abstinence may have enhanced extrinsic motivation to quit and thereby disrupted the association between subjective craving for cigarettes (and potentially increased striatal anticipation for smoking reward) and actual smoking behavior.
Given the recruitment of non-treatment-seeking smokers and the use of CM in the present study, future work will be needed to determine the generalizability of these findings for smoking cessation. The availability of monetary incentives was a primary motivator for abstinence among this sample. Thus, it is possible that striatal activation to monetary reward served as an index of motivation for money, which in turn directly influenced motivation to abstain from smoking during CM. It is noteworthy that the predictive relationship between striatal activation and lapse outcome was based on abstinence-induced attenuation of monetary reward anticipation, suggesting that these findings are not merely a reflection of baseline differences in desire for money. However, individual differences in valuation of money, whether due to socioeconomic status or other factors, will be an important consideration for future studies. Consideration of other non-drug rewards, such as personally relevant rewards, also deserves attention. Finally, it is unclear whether reward sensitivity may be modulated by intrinsic motivation to quit or if potential deficits in reward functioning could be alleviated by nicotine replacement therapy or other pharmacotherapy.
It is important to note that responses to smoking and monetary rewards were not necessarily independent, as response to one reward may have been influenced by its relative valuation against other available rewards. Thus, it is possible that the decrease in activation to monetary reward during abstinence simply reflected a decrease in the relative valuation of money against a more highly desired cigarette reward, rather than an actual deficit in non-drug reward processing per se. However, even if this is the case, it is notable that the relative decrease in anticipatory activation to monetary reward predicted smoking outcomes, while the presumably more direct increase in smoking reward anticipation did not. Additional limitations include the relatively small number of abstainers during CM, recruitment of only Caucasian smokers, and short duration of follow-up. Small sample size, in particular, could lead to an inflated estimate of effect size (see Button et al. 2013). particularly given that moderate test-retest reliability of BOLD signal for reward-related tasks is likely to attenuate the ability to detect a true effect (Plichta et al. 2012). Within the present dataset, estimates of test-retest reliability of striatal signal during anticipation of monetary reward (intraclass correlation = 0.45) were consistent with those obtained from previous reports, falling in the “fair to good” range (Plichta et al. 2012). Furthermore, although post hoc power analysis based on observed effect size is contraindicated (Hoenig & Heissey, 2001; Thomas 1997). our current sample size and repeated measures design provided 80 % power to detect a medium effect size, supporting the validity of the present findings. However, as with all neuroimaging studies incorporating increasingly complex designs, it will be critical for future research to provide replication in a larger, more representative sample. Effect sizes obtained here can provide a basis for powering future studies. Despite limitations, the present findings provide the first evidence that abstinence-induced changes in pre-quit striatal response to reward predict smoking behavior over the first several weeks of a quit attempt. Further work is needed to determine whether altered reward processing is associated with long-term cessation outcomes.
The present findings may have important implications for understanding vulnerability to relapse and for smoking cessation interventions. Blunted sensitivity to reward may be an important mechanism contributing to the high prevalence of smoking and higher rates of relapse among individuals with psychiatric comorbidities that are themselves associated with deficits in reward function, including depression and ADHD (Breslau et al. 1998; Kollins et al. 2005). Although the present findings demonstrate the importance of individual variability in striatal reward processing among smokers without comorbid psychopathology, future research should examine whether abstinence-induced deficits are exacerbated among individuals with other disorders. The present findings also suggest that pharmacological or behavioral treatments targeting appetitive processing, such as behavioral activation, may be beneficial as a smoking cessation strategy for vulnerable individuals. Preliminary evidence suggests that behavioral activation contributes to greater abstinence among smokers with depression compared with supportive counseling (MacPherson et al. 2010). suggesting that targeting reward deficits as a mechanism for improving cessation outcomes may be a promising area of future research.
References
Addicott MA, Baranger DA, Kozink RV, Smoski MJ, Dichter GS, McClernon FJ (2012) Smoking withdrawal is associated with increases in brain activation during decision making and reward anticipation: a preliminary study. Psychopharmacology 219:563–73. doi:10.1007/s00213-011-2404-3
Alessi SM, Badger GJ, Higgins ST (2004) An experimental examination of the initial weeks of abstinence in cigarette smokers. Exp Clin Psychopharmacol 12:276–87. doi:10.1037/1064-1297.12.4.276
Attwood AS, Penton-Voak IS, Munafo MR (2009) Effects of acute nicotine administration on ratings of attractiveness of facial cues. Nicotine Tob Res 11:44–8. doi:10.1093/ntr/ntn006
Bassareo V, Di Chiara G (1999) Modulation of feeding-induced activation of mesolimbic dopamine transmission by appetitive stimuli and its relation to motivational state. Eur J Neurosci 11:4389–97
Beck A, Schlagenhauf F, Wustenberg T, Hein J, Kienast T, Kahnt T, Schmack K, Hagele C, Knutson B, Heinz A, Wrase J (2009) Ventral striatal activation during reward anticipation correlates with impulsivity in alcoholics. Biol Psychiatry 66:734–42. doi:10.1016/j.biopsych.2009.04.035
Besheer J, Bevins RA (2003) Impact of nicotine withdrawal on novelty reward and related behaviors. Behav Neurosci 117:327–40
Bold KW, Yoon H, Chapman GB, McCarthy DE (2013) Factors predicting smoking in a laboratory-based smoking-choice task. Exp Clin Psychopharmacol 21:133–43. doi:10.1037/a0031559
Breslau N, Peterson EL, Schultz LR, Chilcoat HD, Andreski P (1998) Major depression and stages of smoking. A longitudinal investigation. Arch Gen Psychiatry 55:161–6. doi:10.1016/j.biopsych.2009.10.029
Buhler M, Vollstadt-Klein S, Kobiella A, Budde H, Reed LJ, Braus DF, Buchel C, Smolka MN (2010) Nicotine dependence is characterized by disordered reward processing in a network driving motivation. Biol Psychiatry 67:745–52. doi:10.1016/j.biopsych.2009.10.029
Button KS, Ioannidis JP, Mokrysz C, Nosek BA, Flint J, Robinson ES, Munafo MR (2013) Power failure: why small sample size undermines the reliability of neuroscience. Nature Reviews Neuroscience 14:365–76
CDC (2002) Annual smoking attributable mortality, years of potential life lost, and economic costs—United States, 1995-1999. Morbidity and Mortality Weekly Report 51:300–303
CDC (2011) Tobacco use: targeting the nation’s leading killer. At a glance 2011
Chaudhri N, Caggiula AR, Donny EC, Palmatier MI, Liu X, Sved AF (2006) Complex interactions between nicotine and nonpharmacological stimuli reveal multiple roles for nicotine in reinforcement. Psychopharmacology 184:353–66
Dallery J, Glenn IM, Raiff BR (2007) An Internet-based abstinence reinforcement treatment for cigarette smoking. Drug Alcohol Depend 86:230–8. doi:10.1016/j.drugalcdep.2006.06.013
David SP, Munafo MR, Johansen-Berg H, Smith SM, Rogers RD, Matthews PM, Walton RT (2005) Ventral striatum/nucleus accumbens activation to smoking-related pictorial cues in smokers and nonsmokers: a functional magnetic resonance imaging study. Biol Psychiatry 58:488–94
Dawkins L, Powell JH, West R, Powell J, Pickering A (2006) A double-blind placebo controlled experimental study of nicotine: I—effects on incentive motivation. Psychopharmacology (Berl) 189:355–67
Delgado MR, Nystrom LE, Fissell C, Noll DC, Fiez JA (2000) Tracking the hemodynamic responses to reward and punishment in the striatum. J Neurophysiol 84:3072–7
Donny EC, Chaudhri N, Caggiula AR, Evans-Martin FF, Booth S, Gharib MA, Clements LA, Sved AF (2003) Operant responding for a visual reinforcer in rats is enhanced by noncontingent nicotine: implications for nicotine self-administration and reinforcement. Psychopharmacology 169:68–76
Dunn KE, Sigmon SC, Thomas CS, Heil SH, Higgins ST (2008) Voucher-based contingent reinforcement of smoking abstinence among methadone-maintained patients: a pilot study. J Appl Behav Anal 41:527–38
Forbes EE, Hariri AR, Martin SL, Silk JS, Moyles DL, Fisher PM, Brown SM, Ryan ND, Birmaher B, Axelson DA, Dahl RE (2009) Altered striatal activation predicting real-world positive affect in adolescent major depressive disorder. Am J Psychiatry 166:64–73. doi:10.1176/appi.ajp.2008.07081336
Forbes EE, Olino TM, Ryan ND, Birmaher B, Axelson D, Moyles DL, Dahl RE (2010) Reward-related brain function as a predictor of treatment response in adolescents with major depressive disorder. Cogn Affect Behav Neurosci 10:107–18. doi:10.3758/CABN.10.1.107
Forbes EE, Phillips ML, Silk JS, Ryan ND, Dahl RE (2011) Neural systems of threat processing in adolescents: role of pubertal maturation and relation to measures of negative affect. Dev Neuropsychol 36:429–52. doi:10.1080/87565641.2010.550178
Forbes EE, Dahl RE, Almeida JR, Ferrell RE, Nimgaonkar VL, Mansour H, Sciarrillo SR, Holm SM, Rodriguez EE, Phillips ML (2012) PER2 rs2304672 polymorphism moderates circadian-relevant reward circuitry activity in adolescents. Biol Psychiatry 71:451–7. doi:10.1016/j.biopsych.2011.10.012
Franklin TR, Wang Z, Wang J, Sciortino N, Harper D, Li Y, Ehrman R, Kampman K, O’Brien CP, Detre JA, Childress AR (2007) Limbic activation to cigarette smoking cues independent of nicotine withdrawal: a perfusion fMRI study. Neuropsychopharmacology 32:2301–9. doi:10.1038/sj.npp.1301371
Freeman TP, Morgan CJ, Beesley T, Curran HV (2012) Drug cue induced overshadowing: selective disruption of natural reward processing by cigarette cues amongst abstinent but not satiated smokers. Psychol Med 42:161–71. doi:10.1017/S0033291711001139
Garavan H, Pankiewicz J, Bloom A, Cho JK, Sperry L, Ross TJ, Salmeron BJ, Risinger R, Kelley D, Stein EA (2000) Cue-induced cocaine craving: neuroanatomical specificity for drug users and drug stimuli. Am J Psychiatry 157:1789–98
Geier CF, Sweitzer MM, Denlinger R, Sparacino G, Donny EC (2014) Abstinent adult daily smokers show reduced anticipatory but elevated saccade-related brain responses during a rewarded antisaccade task. Psychiatry Res 223:140–7. doi:10.1016/j.pscychresns.2014.04.007
Glenn IM, Dallery J (2007) Effects of internet-based voucher reinforcement and a transdermal nicotine patch on cigarette smoking. J Appl Behav Anal 40:1–13
Goldstein RZ, Alia-Klein N, Tomasi D, Zhang L, Cottone LA, Maloney T, Telang F, Caparelli EC, Chang L, Ernst T, Samaras D, Squires NK, Volkow ND (2007) Is decreased prefrontal cortical sensitivity to monetary reward associated with impaired motivation and self-control in cocaine addiction? Am J Psychiatry 164:43–51. doi:10.1176/appi.ajp.164.1.43
Goldstein RZ, Tomasi D, Alia-Klein N, Honorio Carrillo J, Maloney T, Woicik PA, Wang R, Telang F, Volkow ND (2009) Dopaminergic response to drug words in cocaine addiction. J Neurosci 29:6001–6. doi:10.1523/JNEUROSCI.4247-08.2009
Grusser SM, Wrase J, Klein S, Hermann D, Smolka MN, Ruf M, Weber-Fahr W, Flor H, Mann K, Braus DF, Heinz A (2004) Cue-induced activation of the striatum and medial prefrontal cortex is associated with subsequent relapse in abstinent alcoholics. Psychopharmacology 175:296–302. doi:10.1007/s00213-004-1828-4
Heatherton TF, Kozlowski LT, Frecker RC, Fagerstrom KO (1991) The Fagerstrom Test for Nicotine Dependence: a revision of the Fagerstrom Tolerance Questionnaire. Br J Addict 86:1119–27
Heinz A, Siessmeier T, Wrase J, Hermann D, Klein S, Grusser SM, Flor H, Braus DF, Buchholz HG, Grunder G, Schreckenberger M, Smolka MN, Rosch F, Mann K, Bartenstein P (2004) Correlation between dopamine D(2) receptors in the ventral striatum and central processing of alcohol cues and craving. Am J Psychiatry 161:1783–9. doi:10.1176/appi.ajp.161.10.1783
Higgins ST, Heil SH, Solomon LJ, Bernstein IM, Lussier JP, Abel RL, Lynch ME, Badger GJ (2004) A pilot study on voucher-based incentives to promote abstinence from cigarette smoking during pregnancy and postpartum. Nicotine Tob Res 6:1015–20
Hoenig JM, Heisey DM (2001) The abuse of power: the pervasive fallacy of power calculations for data analysis. The American Statistician 55:1–6
Hughes JR, Hatsukami D (1986) Signs and symptoms of tobacco withdrawal. Arch Gen Psychiatry 43:289–94
Hughes JR, Shiffman S, Callas P, Zhang J (2003) A meta-analysis of the efficacy of over-the-counter nicotine replacement. Tob Control 12:21–7
Janes AC, Pizzagalli DA, Richardt S, de BFB, Chuzi S, Pachas G, Culhane MA, Holmes AJ, Fava M, Evins AE, Kaufman MJ (2010) Brain reactivity to smoking cues prior to smoking cessation predicts ability to maintain tobacco abstinence. Biol Psychiatry 67:722–9. doi:10.1016/j.biopsych.2009.12.034
Kahnt T, Heinzle J, Park SQ, Haynes JD (2010) The neural code of reward anticipation in human orbitofrontal cortex. Proc Natl Acad Sci USA 107:6010–5. doi:10.1073/pnas.0912838107
Knutson B, Fong GW, Adams CM, Varner JL, Hommer D (2001) Dissociation of reward anticipation and outcome with event-related fMRI. Neuroreport 12:3683–7
Kollins SH, McClernon FJ, Fuemmeler BF (2005) Association between smoking and attention-deficit/hyperactivity disorder symptoms in a population-based sample of young adults. Arch Gen Psychiatry 62:1142–7. doi:10.1001/archpsyc.62.10.1142
Koob GF, Le Moal M (2005) Plasticity of reward neurocircuitry and the ‘dark side’ of drug addiction. Nat Neurosci 8:1442–4. doi:10.1038/nn1105-1442
Lessov-Schlaggar CN, Lepore RL, Kristjansson SD, Schlaggar BL, Barnes KA, Petersen SE, Madden PA, Heath AC, Barch DM (2013) Functional neuroimaging study in identical twin pairs discordant for regular cigarette smoking. Addict Biol 18:98–108. doi:10.1111/j.1369-1600.2012.00435.x
Leventhal AM, Munafo M, Tidey JW, Sussman S, Monterosso JR, Sun P, Kahler CW (2012) Anhedonia predicts altered processing of happy faces in abstinent cigarette smokers. Psychopharmacology (Berl) 222:343–51. doi:10.1007/s00213-012-2649-5
Luo S, Ainslie G, Giragosian L, Monterosso JR (2011) Striatal hyposensitivity to delayed rewards among cigarette smokers. Drug Alcohol Depend 116:18–23. doi:10.1016/j.drugalcdep.2010.11.012
MacPherson L, Tull MT, Matusiewicz AK, Rodman S, Strong DR, Kahler CW, Hopko DR, Zvolensky MJ, Brown RA, Lejuez CW (2010) Randomized controlled trial of behavioral activation smoking cessation treatment for smokers with elevated depressive symptoms. J Consult Clin Psychol 78:55–61. doi:10.1037/a0017939
Martin-Soelch C, Chevalley AF, Kunig G, Missimer J, Magyar S, Mino A, Schultz W, Leenders KL (2001) Changes in reward-induced brain activation in opiate addicts. Eur J Neurosci 14:1360–8
Martin-Soelch C, Missimer J, Leenders KL, Schultz W (2003) Neural activity related to the processing of increasing monetary reward in smokers and nonsmokers. Eur J Neurosci 18:680–8
McClernon FJ, Hiott FB, Huettel SA, Rose JE (2005) Abstinence-induced changes in self-report craving correlate with event-related FMRI responses to smoking cues. Neuropsychopharmacology 30:1940–7
O’Doherty JP (2004) Reward representations and reward-related learning in the human brain: insights from neuroimaging. Curr Opin Neurobiol 14:769–76. doi:10.1016/j.conb.2004.10.016
O’Doherty J, Dayan P, Schultz J, Deichmann R, Friston K, Dolan RJ (2004) Dissociable roles of ventral and dorsal striatum in instrumental conditioning. Science 304:452–4. doi:10.1126/science.1094285
Peters J, Bromberg U, Schneider S, Brassen S, Menz M, Banaschewski T, Conrod PJ, Flor H, Gallinat J, Garavan H, Heinz A, Itterman B, Lathrop M, Martinot JL, Paus T, Poline JB, Robbins TW, Rietschel M, Smolka M, Strohle A, Struve M, Loth E, Schumann G, Buchel C (2011) Lower ventral striatal activation during reward anticipation in adolescent smokers. Am J Psychiatry 168:540–9. doi:10.1176/appi.ajp.2010.10071024
Plichta MM, Schwarz AJ, Grimm O, Morgen K, Mier D, Haddad L, Gerdes AB, Sauer C, Tost H, Esslinger C, Colman P, Wilson F, Kirsch P, Meyer-Lindenberg A (2012) Test-retest reliability of evoked BOLD signals from a cognitive-emotive fMRI test battery. NeuroImage 60:1746–58
Powell J, Dawkins L, Davis RE (2002a) Smoking, reward responsiveness, and response inhibition: tests of an incentive motivational model. Biol Psychiatry 51:151–63
Powell J, Tait S, Lessiter J (2002b) Cigarette smoking and attention to signals of reward and threat in the Stroop paradigm. Addiction 97:1163–70
Powell JH, Pickering AD, Dawkins L, West R, Powell JF (2004) Cognitive and psychological correlates of smoking abstinence, and predictors of successful cessation. Addict Behav 29:1407–26
Robinson TE, Berridge KC (1993) The neural basis of drug craving: an incentive-sensitization theory of addiction. Brain Res Brain Res Rev 18:247–91
Roll JM, Higgins ST (2000) A within-subject comparison of three different schedules of reinforcement of drug abstinence using cigarette smoking as an exemplar. Drug Alcohol Depend 58:103–9
Roll JM, Higgins ST, Badger GJ (1996) An experimental comparison of three different schedules of reinforcement of drug abstinence using cigarette smoking as an exemplar. J Appl Behav Anal 29:495–504. doi:10.1901/jaba.1996.29-495, quiz 504-5
Rose EJ, Ross TJ, Salmeron BJ, Lee M, Shakleya DM, Huestis M, Stein EA (2012) Chronic exposure to nicotine is associated with reduced reward-related activity in the striatum but not the midbrain. Biol Psychiatry 71:206–13. doi:10.1016/j.biopsych.2011.09.013
Rose EJ, Ross TJ, Salmeron BJ, Lee M, Shakleya DM, Huestis MA, Stein EA (2013) Acute nicotine differentially impacts anticipatory valence- and magnitude-related striatal activity. Biol Psychiatry 73:280–8. doi:10.1016/j.biopsych.2012.06.034
Schultz W (2000) Multiple reward signals in the brain. Nat Rev Neurosci 1:199–207. doi:10.1038/35044563
Shiffman SM, Jarvik ME (1976) Smoking withdrawal symptoms in two weeks of abstinence. Psychopharmacology (Berl) 50:35–9
Sweitzer MM, Denlinger RL, Donny EC (2013a) Dependence and withdrawal-induced craving predict abstinence in an incentive-based model of smoking relapse. Nicoting Tob Res 15:36–43. doi:10.1093/ntr/nts080
Sweitzer MM, Geier CF, Joel DL, McGurrin P, Denlinger RL, Forbes EE, Donny EC (2013b) Dissociated effects of anticipating smoking versus monetary reward in the caudate as a function of smoking abstinence. Biol Psychiatry 76:681–688. doi:10.1016/j.biopsych.2013.11.013
Thiel KJ, Sanabria F, Neisewander JL (2009) Synergistic interaction between nicotine and social rewards in adolescent male rats. Psychopharmacology 204:391–402. doi:10.1007/s00213-009-1470-2
Thomas L (1997) Retrospective power analysis. Conservation Biology 11:276–280
Tidey JW, O’Neill SC, Higgins ST (2002) Contingent monetary reinforcement of smoking reductions, with and without transdermal nicotine, in outpatients with schizophrenia. Exp Clin Psychopharmacol 10:241–7
Toll BA, Katulak NA, McKee SA (2006) Investigating the factor structure of the Questionnaire on Smoking Urges-Brief (QSU-Brief). Addict Behav 31:1231–9. doi:10.1016/j.addbeh.2005.09.008
Van Zundert RM, Ferguson SG, Shiffman S, Engels R (2012) Dynamic effects of craving and negative affect on adolescent smoking relapse. Health Psychol 31:226–34. doi:10.1037/a0025204
Versace F, Engelmann JM, Robinson JD, Jackson EF, Green CE, Lam CY, Minnix JA, Karam-Hage MA, Brown VL, Wetter DW, Cinciripini PM (2014) Prequit FMRI responses to pleasant cues and cigarette-related cues predict smoking cessation outcome. Nicotine Tob Res 16:697–708. doi:10.1093/ntr/ntt214
Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ, Dewey SL, Wolf AP (1993) Decreased dopamine D2 receptor availability is associated with reduced frontal metabolism in cocaine abusers. Synapse 14:169–77. doi:10.1002/syn.890140210
Weaver MT, Sweitzer M, Coddington S, Sheppard J, Verdecchia N, Caggiula AR, Sved AF, Donny EC (2012) Precipitated withdrawal from nicotine reduces reinforcing effects of a visual stimulus for rats. Nicotine Tob Res 14:824–32. doi:10.1093/ntr/ntr293
Wilson SJ, Delgado MR, McKee SA, Grigson PS, MacLean RR, Nichols TT, Henry SL (2014) Weak ventral striatal responses to monetary outcomes predict an unwillingness to resist cigarette smoking. Cog Affect Behav Neurosci 14:1196–207. doi:10.3758/s13415-014-0285-8
Wrase J, Grusser SM, Klein S, Diener C, Hermann D, Flor H, Mann K, Braus DF, Heinz A (2002) Development of alcohol-associated cues and cue-induced brain activation in alcoholics. Eur Psychiatry 17:287–91
Wrase J, Schlagenhauf F, Kienast T, Wustenberg T, Bermpohl F, Kahnt T, Beck A, Strohle A, Juckel G, Knutson B, Heinz A (2007) Dysfunction of reward processing correlates with alcohol craving in detoxified alcoholics. Neuroimage 35:787–94. doi:10.1016/j.neuroimage.2006.11.043
Wray JM, Gass JC, Tiffany ST (2013) A systematic review of the relationships between craving and smoking cessation. Nicotine & tobacco research : official journal of the Society for Research on Nicotine and Tobacco 15:1167–82. doi:10.1093/ntr/nts268
Acknowledgments
This research was supported by NIH grant (DA027441) to ECD. Maggie Sweitzer was supported by NSF IGERT award (0549352) and the Center for the Neural Basis of Cognition.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to report.
Rights and permissions
About this article

Cite this article
Sweitzer, M.M., Geier, C.F., Denlinger, R. et al. Blunted striatal response to monetary reward anticipation during smoking abstinence predicts lapse during a contingency-managed quit attempt. Psychopharmacology 233, 751–760 (2016). https://doi.org/10.1007/s00213-015-4152-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-015-4152-2