
Abstract
Polycystic ovary syndrome (PCOS) is a prevalent gynecological-endocrinological disorder characterized by hyperandrogenism, menstrual irregularities, and metabolic disturbances. Recent research has highlighted the role of oxidative stress and chronic inflammation in exacerbating PCOS symptoms and impeding reproductive outcomes. Astaxanthin, a potent antioxidant found in marine organisms, has been suggested as a potential therapeutic intervention due to its ability to reduce oxidative stress and inflammation. This meta-analysis systematically reviews randomized controlled trials assessing the impact of astaxanthin supplementation on oxidative stress and reproductive outcomes in women with PCOS. Data from four trials were analyzed, focusing on markers of oxidative stress and reproductive health metrics. The meta-analysis utilized fixed and random-effects models to synthesize results, with heterogeneity assessed using Chi-square and I2 statistics. The findings indicate that while astaxanthin significantly improves markers of total antioxidant capacity (TAC) in follicular fluid, it does not show a consistent effect on other oxidative stress biomarkers such as malondialdehyde (MDA), catalase (CAT), or superoxide dismutase (SOD). Reproductive outcomes, including oocyte quality and the number of high-quality embryos, showed moderate improvements, although effects on fertilization rates and pregnancy outcomes were insignificant. The analysis highlights variability in study designs and dosing, suggesting a need for further research with standardized protocols and larger sample sizes. Future studies should focus on determining optimal dosing, exploring mechanistic pathways, and investigating the combined effects of astaxanthin with other interventions. Longitudinal studies are needed to assess long-term benefits and safety, and personalized approaches could enhance treatment efficacy for individuals with PCOS.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Astaxanthin (Fig. 1), a xanthophyll carotenoid predominantly found in marine organisms such as microalgae, salmon, and shrimp, has emerged as a potent antioxidant with broad therapeutic potential (Cao et al. 2023). Unlike other antioxidants, astaxanthin's molecular structure allows it to reside within the cell membrane, providing robust protection against lipid peroxidation and oxidative damage (Dang et al. 2024). Additionally, astaxanthin exhibits anti-inflammatory solid properties by modulating key signaling pathways and reducing the production of cytokines (Jabarpour et al. 2024b).

Molecular structure of astaxanthin
Polycystic ovary syndrome (PCOS) is a gynecological-endocrinological disorder that affects approximately 5–10% of women of reproductive age worldwide. It is characterized by a constellation of symptoms, including hyperandrogenism, hirsutism, acne, menstrual irregularities, polycystic ovaries, chronic anovulation, and metabolic disturbances such as insulin resistance and obesity (Norman et al. 2007). The pathophysiology of PCOS is complex and multifactorial, involving genetic, environmental, and hormonal factors (Di Lorenzo et al. 2023). Over the past few years, it has been discussed the role of chronic low-grade inflammation and increased oxidative stress in the development and progression of PCOS exacerbating the metabolic and reproductive abnormalities observed in affected women (Orisaka et al. 2023). In this context, astaxanthin might offer a promising therapeutic approach.
Biosynthesis of astaxanthin: mechanisms and pathways
Astaxanthin, a terpenoid, is synthesized from β-carotene, its primary precursor. Isoprenoids, which include carotenoids, are a diverse group of natural compounds. Carotenoids can be categorized into two groups based on their chemical structure or functional characteristics. The biosynthesis of isoprenoids typically starts from isopentenyl pyrophosphate (IPP). This process occurs through two main pathways. The first is the mevalonate (MVA) pathway, found in many eukaryotes, archaea, and actinobacteria. In this pathway, IPP is synthesized from acetyl-CoA. The second pathway is the 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway. It begins with pyruvate and glyceraldehyde 3-phosphate, progressing through 1-deoxy-D-xylulose 5-phosphate (DXP) to MEP. This pathway is crucial as it is present in almost all bacteria and photosynthetic eukaryotes' chloroplasts. In eukaryotic microbes, IPP is converted into dimethylallyl diphosphate (DMAPP) by the enzyme IPP isomerase (IDI). In contrast, in prokaryotic microbes and plant plastids, the MEP pathway is used for IPP and DMAPP synthesis, starting from pyruvate and glyceraldehyde-3-phosphate (Gupta et al. 2021; Basiony et al. 2022; Nishida et al. 2023; Göttl et al. 2024).
The formation of carotenoids includes the creation of a C20 compound called geranylgeranyl pyrophosphate (GGPP), which is synthesized from IPP and DMAPP. GGPP plays a crucial role in the carotenoid biosynthesis pathway. It serves as the starting point for the production of carotenoids, progressing through a sequence of compounds: phytoene, ζ-carotene, lycopene, β-carotene, canthaxanthin, and finally, astaxanthin (Huang et al. 2019).
In carotenoid biosynthesis, β-carotene is synthesized through the actions of β-carotene hydroxylase (BHY) and β-carotene ketolase (BKT). BHY catalyzes the 3-hydroxylation of β-carotene, while BKT is responsible for the 4-ketolation process. BHY genes are found in higher plants, whereas BKT genes are present in certain bacteria, microalgae, and yeasts. As a result, most higher plants cannot produce ketocarotenoids but can produce various hydroxylated carotenoids instead (Huang et al. 2012; Zhu et al. 2018; Bai et al. 2020). In fungi, bacteria, and algae, the intermediates between β-carotene and astaxanthin include β-cryptoxanthin, zeaxanthin, adonixanthin, equinenone, canthaxanthin, 3-hydroxyequinenone, and adonirubin. However, five key intermediates are common across these organisms: β-cryptoxanthin, zeaxanthin, adonixanthin, canthaxanthin, and adonirubin (Zhang et al. 2020). Given the above, the biosynthesis pathway can be divided into two main processes. First, the MVA and glycolytic pathways convert glucose into β-carotene and farnesyl pyrophosphate (FPP). Notably, FPP is transformed into β-carotene through the action of the enzymes crtE, crtYB, and crtI. Second, the synthesis of astaxanthin from β-carotene involves multiple branches. The final production of astaxanthin depends on the sequential actions of hydroxylation (catalyzed by crtZ) and β-carotene ketolase (catalyzed by crtW) (Ding et al. 2023).
Physicochemical properties of astaxanthin: structure, stability, and solubility
Astaxanthin belongs to the xanthophyll group and has the molecular formula C40H52O4, with a 596.84 g/mol molar mass. Its structure consists of a polyene chain linking two terminal rings. Each end of the molecule features a hydroxyl group (-OH), and it includes two asymmetric carbons at the 3 and 3' positions of the β-ionone ring. The double bonds in astaxanthin can exist in cis or trans configurations, with the transform being the most common in natural astaxanthin. Astaxanthin also has three configurational isomers, which vary among different organisms: two enantiomers, (3S, 3′S) and (3R, 3′R), and an optically inactive mesoform, (3R, 3′S). In nature, the predominantly found stereoisomers are (3R, 3′R), (3S, 3′S), (3R), and (3′S) (Ambati et al. 2014; Zhao et al. 2019; Martínez-Álvarez et al. 2020). Due to its high lipophilicity (logP = 10.3), astaxanthin has very low aqueous solubility, resulting in poor oral bioavailability. Additionally, astaxanthin is chemically unstable, particularly during processing and storage, where it is vulnerable to degradation from heat, pH changes, light, and oxygen (Qiao et al. 2018; Ghosh et al. 2023). In its structure, astaxanthin's hydroxyl groups can bind with various fatty acids, such as oleic acid or palmitic acid, to form monoesters or diesters (Brotosudarmo et al. 2020; Medoro et al. 2023). The red and pink color of astaxanthin is due to its 11 conjugated double bonds. This conjugated system gives astaxanthin its distinctive color and contributes to its potent antioxidant properties and ability to cross the blood–brain barrier (BBB) (Fakhri et al. 2019; Stachowiak and Szulc 2021). After about 120 h at 4 °C, astaxanthin degrades by 69%. Complete degradation occurs between 25 °C and 50 °C, highlighting its susceptibility to thermal denaturation. This, combined with its high hydrophobicity, underscores the challenge of maintaining its bioavailability (Yang et al. 2021; Snell and Carberry 2022). It is noteworthy that synthetic astaxanthin contains several stereoisomers, some of which do not occur naturally. These synthetic forms often have reduced bioavailability and lower technological stability. In contrast, only natural astaxanthin is considered safe for human use and consumption (Abdelazim et al. 2023; Nair et al. 2023).
Pharmacokinetics of astaxanthin: absorption, distribution, metabolism, and excretion
Astaxanthin is absorbed in the body along with fatty acids, emphasizing the connection between its absorption and the dietary components consumed with it. Diets high in carbohydrates enhance carotenoid absorption, while low-fat diets reduce it. After ingestion, astaxanthin combines with bile acids to form micelles in the intestine. These micelles are partially absorbed by intestinal mucosal cells and incorporated into chylomicrons. Chylomicrons are then processed by lipoprotein lipase, released into the lymphatic system, and eventually enter the bloodstream. The liver and other tissues rapidly clear the remnants of chylomicrons. Astaxanthin is transported to various tissues via lipoproteins, including very low-density lipoproteins (VLDL), low-density lipoproteins (LDL), and high-density lipoproteins (HDL). Additionally, astaxanthin integrates into lipid bilayers, where it helps maintain cell membrane integrity, supports the redox state, and ensures proper mitochondrial function (Visioli and Artaria 2017; Donoso et al. 2021; Raza et al. 2021; Abdol Wahab et al. 2022). The maximum concentration of astaxanthin in the blood observed 30 min after a meal is 2.4 to 3 times higher than the levels before the meal. This increase is likely due to the effects of cholesterol lipase and stimulated bile secretion from oral administration, which enhances the hydrolysis and absorption of astaxanthin esters. Astaxanthin is primarily excreted through the kidneys, but the spleen also eliminates it (Abdelazim et al. 2023). The half-life of astaxanthin ranges from 16 to 21 h. A single tolerated dose of astaxanthin in healthy volunteers was 40 mg (Niu et al. 2020; Bahbah et al. 2021).
Pharmacodynamics of astaxanthin: mechanisms of action and biological effects
Understanding the mechanisms by which astaxanthin acts in the organism allows us to come up with the idea of using this substance as a therapeutic alternative in managing various conditions. It is known that astaxanthin exhibits anti-inflammatory, antioxidant, anti-apoptotic, and anti-cancer properties. Because of these effects, astaxanthin has been studied for its potential to treat cardiovascular and neurodegenerative diseases. The first two properties are more relevant in the specific case of PCOS, whose pathophysiology involves inflammation and oxidative stress (Rizzardi et al. 2022).
Astaxanthin's crucial role in redox biochemistry is likely attributed to its unique chemical structure and outstanding quenching and scavenging capabilities. It targets singlet oxygen, superoxide, hydrogen peroxide, and hydroxyl radicals and prevents lipid peroxidation. Besides directly affecting reactive oxygen species (ROS), astaxanthin is capable of reducing oxidative damage by activating the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) pathway and the mitogen-activated protein kinase (MAPK)/extracellular signal-regulated protein kinase (ERK) pathway. These pathways promote the release of nuclear factor erythroid 2-related factor 2 (Nrf2) from Kelch-like ECH-associated protein 1. Subsequently, Nrf2 is directed to the nucleus, triggering the Nrf2 antioxidant response element (ARE) (Zarneshan et al. 2020). The PI3K/Akt pathway also increases the production of enzymes that combat oxidative stress, such as Heme oxygenase-1 (HO-1), NAD(P)H quinone oxidoreductase-1 (NQO-1), glutathione-S-transferase-α1, glutamate-cysteine ligase modifier subunit, and glutamate-cysteine ligase catalytic subunit (Wang et al. 2010; Wu et al. 2014, 2015).
The nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) is a critical transcription factor involved in regulating immune and inflammatory responses. Under normal conditions, NF-κB is kept in the cytoplasm in an inactive form bound to the inhibitor of nuclear factor-κB (IκB). Inside the nucleus, NF-κB binds to specific deoxyribonucleic acid (DNA) sequences and promotes the expression of genes involved in inflammation, producing cytokines like tumor necrosis factor-alpha (TNF-α), interleukin (IL)-1β, and IL-6 (Liu et al. 2017; Taniguchi and Karin 2018). Astaxanthin has been shown to impact this pathway in multiple ways. It can inhibit NF-κB activation by preventing phosphorylation and subsequent degradation, or it can upregulate the expression of IκB-messenger ribonucleic acid (mRNA) and protein in the cells (Yang et al. 2019). Finally, there is an interesting interrelation between astaxanthin's antioxidant and anti-inflammatory effects. As mentioned, this substance boosts the activity of Nrf2, which has the capacity to oppose the NF-κB pathway (Wardyn et al. 2015).
Implications of astaxanthin supplementation on oxidative stress and reproductive outcomes in women with PCOS: insights from included randomized clinical trials
Literature search methodology
A thorough literature search was conducted across major academic databases, including PubMed, Scopus, and Web of Science. The search strategy involved a combination of keywords such as "astaxanthin," "PCOS," "oxidative stress," and "randomized controlled trials." This search aimed to identify studies evaluating the effects of astaxanthin on oxidative stress and reproductive health in women with PCOS. Only English-language studies were considered to maintain consistency and clarity in the review process. To qualify for inclusion in the review, studies had to be randomized, placebo-controlled clinical trials involving human participants diagnosed with PCOS. The intervention under review had to be astaxanthin, with comparisons made against a placebo group. Outcomes of interest included markers of oxidative stress and reproductive health metrics. The initial search results were screened to remove duplicates and records marked as ineligible by automation tools. Subsequent screening involved evaluating titles and abstracts to select studies that met the inclusion criteria. Full-text assessments were then performed to confirm eligibility. Data extraction focused on the study design, participant demographics, intervention specifics, and reported outcomes related to oxidative stress and reproductive health. Each study was critically appraised for quality to ensure the reliability and validity of the findings. A meta-analysis was conducted for studies that provided sufficient data to synthesize the results effectively.
Results of literature search following PRISMA guidelines
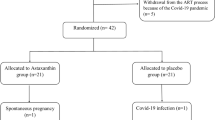
The literature search for this systematic review was conducted following the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) (Page et al. 2021) guidelines to ensure a comprehensive and rigorous approach. Initially, a total of 180 records were identified from various databases and 21 records from registers. To ensure the accuracy and relevance of the studies, duplicates were first removed, totaling 49 records. Additionally, 16 records were marked as ineligible by automation tools, and a further 89 records were excluded for other reasons, such as not meeting the predefined eligibility criteria or being irrelevant to the review topic. Following the removal of these records, 47 records proceeded to the screening phase. During this phase, 32 records were deemed unsuitable based on their titles and abstracts. The remaining 15 reports were sought for full-text retrieval to further evaluate their eligibility. Despite efforts to retrieve these reports, 11 were ultimately not accessible. Of the reports that were retrieved, four were assessed for their eligibility according to the inclusion and exclusion criteria established for this review. This careful assessment process led to the inclusion of four studies in the final review. It is important to note that no additional reports were available for the studies included in the review, which limited the scope of data available for analysis. Figure 2 presents the PRISMA flow diagram, illustrating the process of study selection and screening for this systematic review (Page et al. 2021).

PRISMA flow diagram of study selection and screening process (Page et al. 2021)
Methodology of the meta-analysis: protocol, data extraction, and statistical analysis
Data extraction was conducted using a standardized form, focusing on study design, participant demographics, intervention specifics, and reported outcomes related to oxidative stress and reproductive health. Two independent reviewers (L.F.L. and S.M.B.) extracted data, resolving discrepancies through discussion or third-party consultation. The quality of the included studies was appraised to ensure the reliability and validity of findings. A meta-analysis was performed for studies with sufficient data to synthesize results effectively, using statistical methods appropriate for the data and heterogeneity observed. To assess heterogeneity in the meta-analysis, both the Chi-square (Chi2) test and the I2 inconsistency measure were utilized. The Chi-square test evaluates whether observed differences among studies are greater than what would be expected by chance. A p-value of ≤ 0.100 from this test was considered indicative of heterogeneity, suggesting that there is variability among the studies beyond random error. Additionally, the I2 statistic was used to quantify the extent of inconsistency across studies, with values exceeding 50% signalling considerable heterogeneity. For the meta-analysis, the inverse variance method was employed to aggregate results. When substantial heterogeneity was detected, as indicated by the Chi-square test and high I2 values, a random-effects model was applied to account for variability between studies. Conversely, when heterogeneity was minimal, a fixed-effects model was used, assuming that the true effect size is consistent across studies. The effect size was quantified using the standardized mean difference (SMD), which expresses the difference between the treated and control groups in standard deviation units. This measure, along with its 95% confidence interval, was visually represented in a Forest Plot, which provides a graphical summary of the effect sizes across studies. The results of the meta-analysis were analysed and interpreted using Revman software, version 5.4. This comprehensive approach ensured a rigorous evaluation of the data, accounting for variability among studies and potential biases. In interpreting the Forest Plot, positive SMD values indicate that the intervention group had higher outcomes compared to the control group. Conversely, negative SMD values suggest that the control group exhibited superior outcomes. In the Forest Plot visualization, studies with positive SMD values will be plotted to the right, reflecting higher efficacy in the treated group, while those with negative SMD values will appear to the left, indicating better outcomes in the control group.
Impact of astaxanthin on women with PCOS: findings and results
PCOS is the most common cause of anovulation (Palomba 2021), which can lead to infertility due to potential changes in oocyte quality and endometrial receptivity. To address PCOS-related infertility, ovulation induction is often employed as a treatment for anovulatory patients (Palomba et al. 2017). Despite this approach, many women with PCOS may still struggle to conceive and may turn to assisted reproductive technologies (ART). Additionally, women with PCOS are at a higher risk for endometrial dysfunction, as well as abnormal trophoblast invasion and placentation, which can increase the likelihood of pregnancy complications such as miscarriage or other disruptions (Palomba et al. 2021). Table 1 provides a comprehensive overview following PRISMA Guidelines (Page et al. 2021) of how astaxanthin impacts oxidative stress and reproductive outcomes in women with PCOS across different randomized clinical trials. The risk of bias evaluation was performed for each included randomized clinical trial (detection, selection of the trial, and reporting of bias) according to the Cochrane Handbook for Systematic Reviews of Interventions (COCHRANE) for quality assessment (Table 2) (Higgins 2008).
Fereidouni et al. (Fereidouni et al. 2024) conducted a randomized controlled trial involving women with PCOS, administering astaxanthin (6 mg/day for 8 weeks) and comparing it to a placebo group. The study found no significant differences in oxidative stress markers such as Malondialdehyde (MDA), Catalase (CAT), Superoxide Dismutase (SOD), and ROS levels in follicular fluid (FF) between the intervention and control groups. However, the study reported improvements in reproductive parameters, including oocyte retrieval, metaphase II (M-II) oocyte count, oocyte maturity rate, and the number of frozen embryos. Despite these improvements, there were no significant differences in chemical, clinical, or multiple pregnancy rates. This suggests that while astaxanthin may enhance certain aspects of reproductive health, its effect on oxidative stress markers in FF may be limited.
In contrast, Jabarpour et al. (Jabarpour et al. 2024a) found significant reductions in MDA levels and increases in Total Antioxidant Capacity (TAC) levels following astaxanthin supplementation (12 mg/day for 8 weeks). This study highlights the potential of astaxanthin to improve oxidative stress markers, which could be beneficial in managing PCOS-related oxidative stress. However, this study did not report on reproductive health outcomes, leaving a gap in understanding the full impact of astaxanthin on fertility.
Another study by Jabarpour et al. (Jabarpour et al. 2023), involving infertile PCOS patients undergoing Intracytoplasmic Sperm Injection (ICSI), administered astaxanthin (12 mg/day for 60 days) and observed significant increases in TAC in FF. The study also reported higher rates of high-quality oocytes, embryos, and oocyte maturity, although there were no significant differences in oocyte number, fertilization rate, or overall fertility rate. These findings suggest that astaxanthin may positively influence oocyte and embryo quality, potentially improving reproductive outcomes in women with PCOS.
Lastly, Gharaei et al. (Gharaei et al. 2022) assessed astaxanthin (8 mg/day for 40 days) in women with PCOS and found significant increases in serum CAT and TAC levels, with no significant changes in serum MDA and SOD levels or FF oxidative stress markers. This study also noted an increase in Nrf2, HO-1, and NQO-1 expression, indicating an enhanced antioxidant defense mechanism. Reproductive outcomes showed significant increases in M-II oocyte and high-quality embryo rates, though chemical and clinical pregnancy rates did not differ significantly. This study underscores the potential of astaxanthin to improve certain aspects of reproductive health, despite mixed results on oxidative stress markers.
Age characteristics, BMI, PRL, LH, FSH, and AMH measures in the meta-analysis sample
The meta-analysis, incorporating data from four studies, provided a comprehensive overview of baseline characteristics for participants in both intervention and control groups. Age distribution was consistent across all studies, showing no significant differences between groups and minimal heterogeneity. Body mass index (BMI) measurements were comparable between intervention and control groups, with an overall fixed effects SMD of 0.00727, reflecting a uniform BMI profile and no notable heterogeneity. Prolactin (PRL) levels showed similar values between groups, with an overall fixed effects SMD of 0.181, indicating only minor variation and slight heterogeneity. Luteinizing hormone (LH) levels were also consistent across the groups, with a fixed effects SMD of 0.0544, demonstrating uniform levels and no significant heterogeneity. Follicle-stimulating hormone (FSH) levels were similarly comparable, with an overall fixed effects SMD of 0.107 and no observed heterogeneity. For anti-müllerian hormone (AMH) levels, the fixed effects SMD was -0.192, with consistent measurements across studies and no significant heterogeneity. These findings highlight that the baseline characteristics were similar between intervention and control groups, suggesting that the groups were well-matched at the start of the studies (Fig. 3).

Sample settings for age (1st), BMI (2nd), PRL (3rd), LH (4th), FSH (5th), and AMH (6th)
Meta-analysis of assisted reproductive outcome measures: fertilization rate, high-quality embryos, number of embryos, and M-II rate
The meta-analysis examined several fertility-related outcomes across multiple studies. For fertilization rates, data from three studies indicated a small, non-significant effect of the intervention, with mean differences close to zero and consistent results across studies. The overall SMD was 0.2, suggesting minimal impact and no significant heterogeneity. Regarding high-quality embryos, the analysis revealed a moderate positive effect of the intervention. The mean differences ranged from 0.162 to 1.108 across studies, with the overall SMD of 0.571, indicating some inconsistency but a general benefit of the intervention on the number of high-quality embryos. For the number of embryos, the meta-analysis found a modest effect of the intervention. The SMDs ranged from 0.116 to 0.654, with a consistent overall SMD of 0.393, showing a modest yet stable impact across studies and minimal variability. Finally, the assessment of the M-II rate (%) showed a moderate positive effect of the intervention, with SMDs ranging from 0.675 to 0.807 and an overall SMD of 0.733. This outcome was consistent across studies with no significant heterogeneity, suggesting a moderate improvement in the M-II rate due to the intervention (Fig. 4).

Sample settings for M-II rate (%) (1st), n° of Embryos (2nd), HighQ Embryos (3rd), and Fertilization rate (4th)
Assessment of oxidative stress biomarkers in follicular fluid
The meta-analysis evaluated various antioxidant measures in FF across multiple studies, revealing insights into the impact of interventions on these outcomes. For TAC, the combined data from Gharaei et al. (Gharaei et al. 2022) and Jabarpour et al. (Jabarpour et al. 2023) showed a significant improvement in the intervention groups. The overall SMD was 0.658 with a 95% CI ranging from 0.241 to 1.075, indicating a notable positive effect of the intervention on TAC. The results were consistent, with no significant heterogeneity across studies, suggesting that the intervention effectively increases TAC in FF. In contrast, the analysis of CAT activity from Gharaei et al. (Gharaei et al. 2022) and Fereidouni et al. (Fereidouni et al. 2024) revealed no significant overall effect of the intervention. The pooled SMD was 0.000464 with a 95% CI of -0.430 to 0.431, suggesting that CAT activity did not significantly differ between intervention and control groups. Moderate to high heterogeneity (Q = 3.5613, I2 = 71.92%) indicates variability in the results, implying that the intervention may not uniformly affect CAT activity across different studies or populations. For SOD activity, the meta-analysis combining data from Gharaei et al. (Gharaei et al. 2022), Jabarpour et al. (Jabarpour et al. 2023), and Fereidouni et al. (Fereidouni et al. 2024) showed no significant differences between groups. The overall SMD was -0.042 with a 95% CI of -0.375 to 0.291, and minimal heterogeneity (Q = 2.1633, I2 = 7.55%) suggests that the intervention does not significantly alter SOD activity. This consistency across studies, along with no significant publication bias, indicates that SOD activity is not substantially impacted by the intervention. Finally, the meta-analysis of MDA levels across studies by Gharaei et al. (Gharaei et al. 2022), Jabarpour et al. (Jabarpour et al. 2023), and Fereidouni et al. (Fereidouni et al. 2024) found no significant effect of the intervention. The combined SMDs of -0.0757 (fixed effects) and -0.0671 (random effects) with 95% CIs of -0.410 to 0.258 and -0.485 to 0.351, respectively, indicate that MDA levels were not significantly affected by the intervention. The moderate variability in results (Q = 3.1207, I2 = 35.91%) suggests that while the intervention does not have a significant impact on MDA levels, there may be some degree of variability in response among different studies. Overall, the findings suggest that while the intervention significantly enhances TAC, it does not significantly affect CAT activity, SOD activity, or MDA levels (Fig. 5).

Sample settings for MDA (1st), SOD (2nd), CAT (3rd), and TAC (4th)
Meta-analysis of oxidative stress biomarkers in blood
The meta-analysis of MDA levels, which serves as a marker of lipid peroxidation, included studies by Gharaei et al. (Gharaei et al. 2022) and Jabarpour et al. (Jabarpour et al. 2024a). Gharaei et al. (Gharaei et al. 2022) reported a mean difference of -0.147 with a standard deviation (SD) of 0.31 in the intervention group compared to controls, while Jabarpour et al. (Jabarpour et al. 2024a) reported a mean difference of -0.409 with a SD of 0.274. The combined fixed and random effects models indicated a non-significant overall effect with a standardized mean difference of -0.294 (SE = 0.205, 95% CI: -0.702 to 0.113, p = 0.155). This suggests no significant reduction in MDA levels due to the interventions. For SOD, an enzyme crucial in combating oxidative stress, the meta-analysis incorporated data from Gharaei et al. (Gharaei et al. 2022) and Jabarpour et al. (Jabarpour et al. 2024a). Gharaei et al. (Gharaei et al. 2022) found an effect size of 0.21 (SD = 0.311), while Jabarpour et al. (Jabarpour et al. 2024a) reported an effect size of 0.636 (SD = 0.278). The fixed effects model yielded a SMD of 0.447 (SE = 0.207, 95% CI: 0.0359 to 0.859, p = 0.033), indicating a significant increase in SOD levels due to the intervention. The random effects model showed similar results with a SMD of 0.446 (SE = 0.212, 95% CI: 0.0257 to 0.867, p = 0.038), suggesting a consistent beneficial effect of the intervention on SOD levels. The TAC analysis, which included Gharaei et al. (Gharaei et al. 2022) and Jabarpour et al. (Jabarpour et al. 2024a), demonstrated mixed results. Gharaei et al. (Gharaei et al. 2022) reported an effect size of 0.862 (SD = 0.325), whereas Jabarpour et al. (Jabarpour et al. 2024a) observed an effect size of 0.286 (SD = 0.272). The fixed effects model indicated a significant increase in TAC with a standardized mean difference of 0.524 (SE = 0.209, 95% CI: 0.110 to 0.938, p = 0.014). The random effects model also showed a positive effect, though with less significance (SMD = 0.547, SE = 0.286, 95% CI: -0.0222 to 1.116, p = 0.059), suggesting that the intervention may improve TAC in the blood (Fig. 6).

Sample settings for MDA (1st), SOD (2nd), and TAC (3rd)
Limitations and future research endeavours
Recently, Maleki-Hajiagha et al. (Maleki-Hajiagha et al. 2024) published a meta-analysis examining the effects of astaxanthin on reproductive outcomes in women. However, their analysis did not focus specifically on women with PCOS. Furthermore, they did not address the impact of astaxanthin on oxidative stress biomarkers in blood and FF. Our meta-analysis aimed to address these gaps in the literature by investigating the specific effects of astaxanthin in PCOS patients. We explored its influence on reproductive outcomes and its potential impact on oxidative stress markers in both blood and FF, providing a more detailed understanding of its benefits for this particular population. We explored the molecular mechanisms through which astaxanthin exerts its effects, reviewed and analyzed the current scientific evidence supporting its use in PCOS, and discussed the potential implications for improving reproductive health and ART success rates. By shedding light on the benefits of astaxanthin, we hoped to provide a comprehensive understanding of its role in managing PCOS and enhancing reproductive outcomes, thereby paving the way for future research and clinical applications in this area.
Despite the promising findings, the included studies have several limitations that warrant consideration. First, there is variability in the dosage and duration of astaxanthin supplementation across the studies. For instance, dosages ranged from 6 mg/day to 12 mg/day, and the duration varied from 40 days to 8 weeks. This variability makes it challenging to determine the optimal dosage and duration for maximum benefit. Additionally, the sample sizes in some studies were relatively small, which could limit the generalizability of the results and increase the risk of Type I and Type II errors. Another limitation is the inconsistency in the assessment of oxidative stress biomarkers. While some studies focused on serum markers like MDA, CAT, and TAC, others evaluated these markers in FF or did not report on reproductive outcomes. This lack of standardization can complicate the interpretation of results and their relevance to clinical practice. Furthermore, several studies did not report on potential confounding factors such as lifestyle, dietary habits, or concurrent medications, which could influence oxidative stress and reproductive health outcomes. The methodology of some studies, including the use of different types of controls and blinding techniques, may also affect the reliability of the findings. For example, while some studies used placebo controls, others employed triple-blind designs. The differences in study design can lead to variability in outcomes and may impact the overall assessment of astaxanthin's efficacy.
Additionally, most studies on astaxanthin's effects in women with PCOS have concentrated on its impact on oxidative stress and reproductive outcomes. While advancements have been made in understanding astaxanthin's role in managing PCOS, further research is needed to explore its effects during pregnancy in women with this condition. Specifically, it is important to investigate how astaxanthin might reduce the risk of multiple gestations, prevent miscarriage, and improve outcomes for the child (Palomba et al. 2015).
To advance the understanding of astaxanthin's impact on PCOS and its role in oxidative stress, future research should address several crucial areas with innovative approaches and robust methodologies. First, conducting multi-center, large-scale trials with diverse populations can provide more generalizable insights and identify variations in astaxanthin's efficacy across different demographic groups. Such studies should adopt a standardized protocol for dosing, duration, and assessment to ensure consistency and comparability of results. Moreover, exploring the mechanistic pathways through which astaxanthin exerts its effects could offer valuable insights. Researchers should investigate how astaxanthin interacts with specific oxidative stress pathways and its impact on cellular processes in detail. This can involve using advanced techniques like metabolomics and proteomics to uncover the molecular changes induced by astaxanthin supplementation. Another promising avenue is the combination of astaxanthin with other therapeutic interventions. Evaluating the synergistic effects of astaxanthin when used in conjunction with lifestyle modifications, such as diet and exercise, or other PCOS treatments, could provide a more holistic approach to managing the condition. Investigating how astaxanthin interacts with other medications and supplements will help in developing comprehensive treatment strategies. Longitudinal studies are also essential to determine the long-term benefits and safety of astaxanthin supplementation. Understanding how prolonged use affects not only oxidative stress but also long-term reproductive health and metabolic outcomes will be crucial for evaluating its overall efficacy and safety. Such studies should include follow-up periods extending beyond the supplementation phase to assess sustained effects and any potential delayed outcomes. Future research should also explore personalized approaches to astaxanthin therapy. Investigating genetic and phenotypic factors that may influence individual responses to astaxanthin can lead to tailored treatment plans. For instance, identifying biomarkers that predict which patients are most likely to benefit from astaxanthin could optimize treatment efficacy. Additionally, integrating patient-reported outcomes and quality of life assessments into research designs will provide a more comprehensive view of how astaxanthin affects daily functioning and well-being. Understanding the patient perspective will enhance the relevance and applicability of research findings.
Satoh et al. (Satoh and Gupta 2021) reported that astaxanthin is widely used as a dietary supplement worldwide and, to date, no adverse effects have been documented in humans. However, no study regarding astaxanthin supplementation in humans during pregnancy has been documented. Although current literature lacks information on astaxanthin toxicity during human pregnancy, studies in pregnant mice provide some insight. In toxicity tests involving repeated doses of astaxanthin at 100, 250, and 500 mg/kg body weight, no abnormalities were observed in body and organ weight, nor in hematological and biochemical parameters throughout the pregnancy (Niu et al. 2020). Additionally, administering astaxanthin crystals to pregnant female rats during the organogenesis period also showed no adverse effects on fetal development (Lin et al. 2017).
Conclusions
The systematic review and meta-analysis revealed a nuanced picture regarding the effects of astaxanthin supplementation in women with PCOS. While the overall findings suggest some benefits, there are notable limitations and areas for further investigation. Astaxanthin shows promise in enhancing TAC, which is beneficial for managing oxidative stress in PCOS. However, its impact on other oxidative stress biomarkers, such as CAT, SOD, and MDA, appears limited or inconsistent. This indicates that while astaxanthin may contribute to reducing oxidative stress, it does not uniformly affect all related biomarkers. In terms of reproductive health, astaxanthin supplementation seems to positively influence parameters such as oocyte quality, embryo quality, and the M-II oocyte rate. Nonetheless, the lack of significant differences in overall pregnancy and fertilization rates suggests that while there may be improvements in certain aspects of reproductive health, the overall impact on fertility outcomes remains unclear. The variability in dosage, duration, and study designs across the reviewed trials presents challenges in determining the optimal use of astaxanthin for PCOS management. Future research should focus on larger, more standardized trials to clarify these effects and explore the mechanisms through which astaxanthin operates. Additionally, investigating the synergistic effects of astaxanthin with other treatments and considering personalized approaches could offer a more comprehensive understanding of its potential benefits for women with PCOS.
Data availability
No datasets were generated or analysed during the current study.
References
Abdelazim K, Ghit A, Assal D, Dorra N, Noby N, Khattab SN, El Feky SE, Hussein A (2023) Production and therapeutic use of astaxanthin in the nanotechnology era. Pharmacol Rep 75:771–790
AbdolWahab NR, MeorMohdAffandi MMR, Fakurazi S, Alias E, Hassan H (2022) Nanocarrier system: State-of-the-art in oral delivery of astaxanthin. Antioxidants (Basel) 11:1676
Ambati RR, Phang SM, Ravi S, Aswathanarayana RG (2014) Astaxanthin: sources, extraction, stability, biological activities and its commercial applications–a review. Mar Drugs 12:128–152
Bahbah EI, Ghozy S, Attia MS, Negida A, Emran TB, Mitra S, Albadrani GM, Abdel-Daim MM, Uddin MS, Simal-Gandara J (2021) Molecular mechanisms of astaxanthin as a potential neurotherapeutic agent. Mar Drugs 19:201
Bai F, Gusbeth C, Frey W, Nick P (2020) Nanosecond pulsed electric fields modulate the expression of the astaxanthin biosynthesis genes psy, crtR-b and bkt 1 in Haematococcus pluvialis. Sci Rep 10:15508
Basiony M, Ouyang L, Wang D, Yu J, Zhou L, Zhu M, Wang X, Feng J, Dai J, Shen Y, Zhang C, Hua Q, Yang X, Zhang L (2022) Optimization of microbial cell factories for astaxanthin production: Biosynthesis and regulations, engineering strategies and fermentation optimization strategies. Synth Syst Biotechnol 7:689–704
Brotosudarmo THP, Limantara L, Setiyono E, Heriyanto (2020) Structures of astaxanthin and their consequences for therapeutic application. Int J Food Sci 2020:2156582
Cao Y, Yang L, Qiao X, Xue C, Xu J (2023) Dietary astaxanthin: an excellent carotenoid with multiple health benefits. Crit Rev Food Sci Nutr 63:3019–3045
Dang Y, Li Z, Yu F (2024) Recent advances in astaxanthin as an antioxidant in food applications. Antioxidants (Basel) 13:879
Di Lorenzo M, Cacciapuoti N, Lonardo MS, Nasti G, Gautiero C, Belfiore A, Guida B, Chiurazzi M (2023) Pathophysiology and nutritional approaches in Polycystic Ovary Syndrome (PCOS): A comprehensive review. Curr Nutr Rep 12:527–544
Ding YW, Lu CZ, Zheng Y, Ma HZ, Jin J, Jia B, Yuan YJ (2023) Directed evolution of the fusion enzyme for improving astaxanthin biosynthesis in Saccharomyces cerevisiae. Synth Syst Biotechnol 8:46–53
Donoso A, González-Durán J, Muñoz AA, González PA, Agurto-Muñoz C (2021) Therapeutic uses of natural astaxanthin: An evidence-based review focused on human clinical trials. Pharmacol Res 166:105479
Fakhri S, Aneva IY, Farzaei MH, Sobarzo-Sánchez E (2019) The neuroprotective effects of astaxanthin: Therapeutic targets and clinical perspective. Molecules 24:2640
Fereidouni F, Kashani L, Amidi F, Khodarahmian M, Zhaeentan S, Ardehjani NA, Rastegar T (2024) Astaxanthin treatment decreases pro-inflammatory cytokines and improves reproductive outcomes in patients with polycystic ovary syndrome undergoing assisted reproductive technology: A randomized clinical trial. Inflammopharmacology 32:2337–2347
Gharaei R, Alyasin A, Mahdavinezhad F, Samadian E, Ashrafnezhad Z, Amidi F (2022) Randomized controlled trial of astaxanthin impacts on antioxidant status and assisted reproductive technology outcomes in women with polycystic ovarian syndrome. J Assist Reprod Genet 39:995–1008
Ghosh A, Banik S, Yamada K, Misaka S, Prud’homme RK, Sato H, Onoue S (2023) Stabilized astaxanthin nanoparticles developed using flash nanoprecipitation to improve oral bioavailability and hepatoprotective effects. Pharmaceutics 15:2562
Göttl VL, Meyer F, Schmitt I, Persicke M, Peters-Wendisch P, Wendisch VF, Henke NA (2024) Enhancing astaxanthin biosynthesis and pathway expansion towards glycosylated C40 carotenoids by Corynebacterium glutamicum. Sci Rep 14:8081
Gupta AK, Seth K, Maheshwari K, Baroliya PK, Meena M, Kumar A, Vinayak V, Harish (2021) Biosynthesis and extraction of high-value carotenoid from algae. FBL 26:171–190
Higgins JP (2008) Cochrane handbook for systematic reviews of interventions version 5.0. 1. The Cochrane Collaboration. http://www.cochrane-handbook.org. Accessed date 13 Aug 2024.
Huang J, Zhong Y, Sandmann G, Liu J, Chen F (2012) Cloning and selection of carotenoid ketolase genes for the engineering of high-yield astaxanthin in plants. Planta 236:691–699
Huang D, Liu W, Li A, Wang C, Hu Z (2019) Discovery of Geranylgeranyl Pyrophosphate Synthase (GGPPS) paralogs from haematococcus pluvialis based on iso-seq analysis and their function on astaxanthin biosynthesis. Mar Drugs 17:696
Jabarpour M, Aleyasin A, Nashtaei MS, Lotfi S, Amidi F (2023) Astaxanthin treatment ameliorates ER stress in polycystic ovary syndrome patients: a randomized clinical trial. Sci Rep 13:3376
Jabarpour M, Aleyasin A, Shabani Nashtaei M, Amidi F (2024a) Astaxanthin supplementation impact on insulin resistance, lipid profile, blood pressure, and oxidative stress in polycystic ovary syndrome patients: A triple-blind randomized clinical trial. Phytother Res 38:321–330
Jabarpour M, Amidi F, Aleyasin A, Nashtaei MS, Marghmaleki MS (2024b) Randomized clinical trial of astaxanthin supplement on serum inflammatory markers and ER stress-apoptosis gene expression in PBMCs of women with PCOS. J Cell Mol Med 28:e18464
Lin Y-J, Lin J-Y, Wang D-S, Chen C-H, Chiou M-H (2017) Safety assessment of astaxanthin derived from engineered Escherichia coli K-12 using a 13-week repeated dose oral toxicity study and a prenatal developmental toxicity study in rats. Regul Toxicol Pharmacol 87:95–105
Liu T, Zhang L, Joo D, Sun SC (2017) NF-κB signaling in inflammation. Signal Transduct Target Ther 2:17023
Maleki-Hajiagha A, Shafie A, Maajani K, Amidi F (2024) Effect of astaxanthin supplementation on female fertility and reproductive outcomes: a systematic review and meta-analysis of clinical and animal studies. J Ovarian Res 17:163
Martínez-Álvarez Ó, Calvo MM, Gómez-Estaca J (2020) Recent advances in astaxanthin micro/nanoencapsulation to improve its stability and functionality as a food ingredient. Mar Drugs 18:406
Medoro A, Davinelli S, Milella L, Willcox BJ, Allsopp RC, Scapagnini G, Willcox DC (2023) Dietary astaxanthin: a promising antioxidant and anti-inflammatory agent for brain aging and adult neurogenesis. Mar Drugs 21:643
Nair A, Ahirwar A, Singh S, Lodhi R, Lodhi A, Rai A, Jadhav DA, Harish A, Varjani S, Singh G, Marchand J, Schoefs B, Vinayak V (2023) Astaxanthin as a king of ketocarotenoids: structure, synthesis, accumulation bioavailability and antioxidant properties. Mar Drugs 21:176
Nishida Y, Berg PC, Shakersain B, Hecht K, Takikawa A, Tao R, Kakuta Y, Uragami C, Hashimoto H, Misawa N, Maoka T (2023) Astaxanthin: Past, Present, and Future. Mar Drugs 21:514
Niu T, Zhou J, Wang F, Xuan R, Chen J, Wu W, Chen H (2020) Safety assessment of astaxanthin from Haematococcus pluvialis: Acute toxicity, genotoxicity, distribution and repeat-dose toxicity studies in gestation mice. Regul Toxicol Pharmacol 115:104695
Norman RJ, Dewailly D, Legro RS, Hickey TE (2007) Polycystic ovary syndrome. Lancet 370:685–697
Orisaka M, Mizutani T, Miyazaki Y, Shirafuji A, Tamamura C, Fujita M, Tsuyoshi H, Yoshida Y (2023) Chronic low-grade inflammation and ovarian dysfunction in women with polycystic ovarian syndrome, endometriosis, and aging. Front Endocrinol (Lausanne) 14:1324429
Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, Shamseer L, Tetzlaff JM, Akl EA, Brennan SE, Chou R, Glanville J, Grimshaw JM, Hróbjartsson A, Lalu MM, Li T, Loder EW, Mayo-Wilson E, McDonald S, McGuinness LA, Stewart LA, Thomas J, Tricco AC, Welch VA, Whiting P, Moher D (2021) The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 372:n71–n71
Palomba S (2021) Is fertility reduced in ovulatory women with polycystic ovary syndrome? An opinion paper. Hum Reprod 36:2421–2428
Palomba S, de Wilde MA, Falbo A, Koster MP, La Sala GB, Fauser BC (2015) Pregnancy complications in women with polycystic ovary syndrome. Hum Reprod Update 21:575–592
Palomba S, Daolio J, La Sala GB (2017) Oocyte Competence in Women with Polycystic Ovary Syndrome. Trends Endocrinol Metab 28:186–198
Palomba S, Piltonen TT, Giudice LC (2021) Endometrial function in women with polycystic ovary syndrome: a comprehensive review. Hum Reprod Update 27:584–618
Qiao X, Yang L, Zhang T, Zhou Q, Wang Y, Xu J, Xue C (2018) Synthesis, stability and bioavailability of astaxanthin succinate diester. J Sci Food Agric 98:3182–3189
Raza SHA, Naqvi SRZ, Abdelnour SA, Schreurs N, Mohammedsaleh ZM, Khan I, Shater AF, Abd El-Hack ME, Khafaga AF, Quan G, Khan R, Wang S, Cheng G, Zan L (2021) Beneficial effects and health benefits of Astaxanthin molecules on animal production: A review. Res Vet Sci 138:69–78
Rizzardi N, Pezzolesi L, Samorì C, Senese F, Zalambani C, Pitacco W, Calonghi N, Bergamini C, Prata C, Fato R (2022) Natural astaxanthin is a green antioxidant able to counteract lipid peroxidation and ferroptotic cell death. Int J Mol Sci 23:15137
Satoh T, Gupta RC (2021) Chapter 51 - Astaxanthin: health benefits and toxicity. In: Gupta RC, Lall R, Srivastava A (eds) Nutraceuticals (Second Edition). Academic Press, pp 881–889
Snell TW, Carberry J (2022) Astaxanthin bioactivity is determined by stereoisomer composition and extraction method. Nutrients 14:1522
Stachowiak B, Szulc P (2021) Astaxanthin for the food industry. Molecules 26:2666
Taniguchi K, Karin M (2018) NF-κB, inflammation, immunity and cancer: coming of age. Nat Rev Immunol 18:309–324
Visioli F, Artaria C (2017) Astaxanthin in cardiovascular health and disease: mechanisms of action, therapeutic merits, and knowledge gaps. Food Funct 8:39–63
Wang HQ, Sun XB, Xu YX, Zhao H, Zhu QY, Zhu CQ (2010) Astaxanthin upregulates heme oxygenase-1 expression through ERK1/2 pathway and its protective effect against beta-amyloid-induced cytotoxicity in SH-SY5Y cells. Brain Res 1360:159–167
Wardyn JD, Ponsford AH, Sanderson CM (2015) Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem Soc Trans 43:621–626
Wu Q, Zhang XS, Wang HD, Zhang X, Yu Q, Li W, Zhou ML, Wang XL (2014) Astaxanthin activates nuclear factor erythroid-related factor 2 and the antioxidant responsive element (Nrf2-ARE) pathway in the brain after subarachnoid hemorrhage in rats and attenuates early brain injury. Mar Drugs 12:6125–6141
Wu H, Niu H, Shao A, Wu C, Dixon BJ, Zhang J, Yang S, Wang Y (2015) Astaxanthin as a potential neuroprotective agent for neurological diseases. Mar Drugs 13:5750–5766
Yang C, Hassan YI, Liu R, Zhang H, Chen Y, Zhang L, Tsao R (2019) Anti-inflammatory effects of different astaxanthin isomers and the roles of lipid transporters in the cellular transport of astaxanthin isomers in caco-2 cell monolayers. J Agric Food Chem 67:6222–6231
Yang L, Qiao X, Gu J, Li X, Cao Y, Xu J, Xue C (2021) Influence of molecular structure of astaxanthin esters on their stability and bioavailability. Food Chem 343:128497
Zarneshan SN, Fakhri S, Farzaei MH, Khan H, Saso L (2020) Astaxanthin targets PI3K/Akt signaling pathway toward potential therapeutic applications. Food Chem Toxicol 145:111714
Zhang C, Chen X, Too HP (2020) Microbial astaxanthin biosynthesis: recent achievements, challenges, and commercialization outlook. Appl Microbiol Biotechnol 104:5725–5737
Zhao X, Zhang X, Liu H, Zhu H, Zhu Y (2019) Enzyme-assisted extraction of astaxanthin from Haematococcus pluvialis and its stability and antioxidant activity. Food Sci Biotechnol 28:1637–1647
Zhu Q, Zeng D, Yu S, Cui C, Li J, Li H, Chen J, Zhang R, Zhao X, Chen L, Liu YG (2018) From golden rice to aSTARice: Bioengineering astaxanthin biosynthesis in rice endosperm. Mol Plant 11:1440–1448
Funding
The authors declare that no external funding was received for the research, authorship, and/or publication of this article. All resources used in this study were provided by the authors' respective institutions or were self-funded. The content is solely the responsibility of the author and does not necessarily represent the official views of any affiliated institutions.
Author information
Authors and Affiliations
Contributions
Victória Dogani Rodrigues and Beatriz Leme Boaro contributed equally to the conception and design of the study, literature search, data analysis, and manuscript drafting. Victória Dogani Rodrigues and Beatriz Leme Boaro share first authorship. Lívia Fornari Laurindo was responsible for the data extraction quality assessment of included studies and contributed to the interpretation of results. Eduardo Federighi Baisi Chagas provided substantial contributions to the study design, statistical analysis, and interpretation of data. He also critically revised the manuscript for important intellectual content. Enzo Pereira de Lima critically revised the manuscript for important intellectual content. Lucas Fornari Laurindo contributed to the study conception, provided significant input in the data synthesis and meta-analysis, and helped with the drafting and revising of the manuscript. Sandra Maria Barbalho was involved in the overall supervision of the research, provided critical insights during the data interpretation phase, and contributed to the final revision of the manuscript. Lucas Fornari Laurindo and Sandra Maria Barbalho share senior authorship. The authors confirm that no paper mill and artificial intelligence was used.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary material
The original contributions presented in the study are comprehensively included in this article. All relevant data supporting the findings of this study are provided within the article and its supplementary materials. For any additional information, specific datasets, or further inquiries regarding the methodology and results, interested researchers are encouraged to contact the corresponding author. The corresponding author is available to assist with any questions and provide additional details as needed to facilitate further research and collaboration.
Lucas Fornari Laurindo and Sandra Maria Barbalho share senior authorship.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Rodrigues, V.D., Boaro, B.L., Laurindo, L.F. et al. Exploring the benefits of astaxanthin as a functional food ingredient: Its effects on oxidative stress and reproductive outcomes in women with PCOS – A systematic review and single-arm meta-analysis of randomized clinical trials. Naunyn-Schmiedeberg's Arch Pharmacol (2024). https://doi.org/10.1007/s00210-024-03432-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00210-024-03432-w