
Abstract
The maternal effect of microplastics (MPs) toxicity is likely influenced by age and brood of test species. This study investigated the maternal effect of polyethylene MP fragments (18.23 ± 8.02 μm) with benzophenone-3 (BP-3; 2.89 ± 0.20% w/w) on chronic toxicity to Daphnia magna over two generations. Neonate (< 24 h old) and adult (5 d old) daphnids in the F0 generation were exposed until 21 d old, then first and third brood neonates in the F1 generation were recovered in clean M4 medium for 21 d. Higher chronic toxicity and maternal effect of MP/BP-3 fragments were observed in the adult group compared with the neonate group, reducing growth and reproduction in both F0 and F1 generations. First brood neonates in the F1 generation showed a higher maternal effect of MP/BP-3 fragments than third brood ones, resulting in enhanced growth and reproduction relative to the control. This study provided insights into the ecological risk of MPs containing plastic additives in the natural environment.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Microplastic (MP) pollution is ubiquitous in the aquatic environment, from riverine and coastal waters to remote lakes and open seas (Ajith et al. 2020; Li et al. 2020; Dusaucy et al. 2021). MPs are a growing concern due to their potential harms: intestinal damage, oxidative stress, neurotoxicity, and behavioral change when ingested by organisms (De Sá et al. 2018; Botterell et al. 2019; Gola et al. 2021). Furthermore, MPs may introduce chemical additives into the environment, such as plasticizers, flame retardants, and ultraviolet (UV) stabilizers, which are known to be toxic to aquatic organisms (Cole et al. 2011; Koelmans 2015; Gunaalan et al. 2020). Previous studies have focused on plasticizers and flame retardants that are more commonly found in plastics, thus the adverse effect of UV stabilizers on aquatic organisms is relatively unknown (Gunaalan et al. 2020; Carve et al. 2021). Several studies have reported the toxicity of UV stabilizers, such as endocrine disruption, stress response, and neurotoxicity (Kim et al. 2014; Martín-Folgar et al. 2018; Tao et al. 2020). In particular, benzophenone-3 (BP-3), one of the most widely used UV stabilizers (Kim and Choi 2014; Bratkovics et al. 2015), is known to inhibit somatic growth, embryonic development, and reproduction of the water flea Daphnia magna (Im et al. 2022). Moreover, our previous studies have reported the combined effect of polyethylene (PE) MP fragments and BP-3 on acute toxicity (Na et al. 2021) and chronic toxicity (Song et al. 2021) in D. magna.
Most D. magna chronic toxicity studies with MPs have been conducted in the standard condition with neonates under 24 h old (OECD 211, 2012). However, in the real environment, populations consist of different age groups, which have different sensitivities to toxic chemicals (Muyssen and Janssen 2007; Wagner et al. 2017; Ellis et al. 2020). For instance, neonates have shown higher toxicity than adults when exposed to insecticides (Ginjupalli and Baldwin 2013), plastic additives (Shen et al. 2019), pharmaceuticals (Wagner et al. 2017), and microplastic (Eltemsah and Bøhn 2019). Therefore, chronic toxicity assessments with different age groups are required to better predict the ecological outcome of MPs.
Parental exposure to MPs may induce maternal effects that influence offspring fitness, including development, somatic growth, and reproduction (Mousseau and Fox 1998; LaMontagne and McCauley, 2001). Several studies have demonstrated the maternal effect in D .magna under MPs exposure (Martins and Guilhermino 2018; Liu et al. 2022; Song et al. 2022). For instance, Liu et al. (2022)demonstrated that parental exposure to polystyrene MPs and roxithromycin decreased the reproduction of D. magna in the subsequent generation. In addition, Song et al. (2022) reported the transgenerational effect of PE MP fragments containing BP-3 across four generations. However, there is limited knowledge about the maternal effects of MPs in D. magna at different ages and broods.
The aim of this study was to investigate the maternal effect of PE MP fragments containing BP-3 additive (MP/BP-3 fragments) on chronic toxicity to D. magna. We evaluated the difference between (1) neonate (< 24 h) and adult (5 d) exposure groups and (2) the first and third brood groups over two subsequent generations. We hypothesized that (1) the neonate exposure will induce higher chronic toxicity and maternal effect than the adult exposure and (2) the maternal effect will be higher on the first brood than on the third brood offspring.
Materials and Methods
Preparation and Physiochemical Characterization of MP/BP-3 Fragments
Virgin PE pellets and BP-3 were purchased from Sigma-Aldrich, USA. The PE pellets were washed with hexane and methanol (J.T. Baker, USA), then rinsed with distilled water to eliminate plastic additives (Lee et al. 2014). After drying in an oven (30 ℃), the BP-3 was incorporated into the PE pellets(3% w/w) in a mixing extruder (LME, Dynisco, USA). The BP-3 content was chosen based on the study by Hahladakis et al. (2018), where they reported that UV-stabilizer composition in plastic is generally less than 3%. Then, the MP/BP-3 fibers were cut into fragments (< 1 mm) and ground in a freezer mill (Freezer/Mill 6875, SPEX® Sample Prep, USA). The products were dried in a desiccator and sieved using a stainless-steel mesh (≤ 20 μm) to obtain MP/BP-3 fragments. The morphology and size of MP/BP-3 fragments were analyzed under a field emission scanning electron microscope (FE-SEM, Quanta 250 FEG, FEI, USA). The BP-3 content in MP/BP-3 fragments was analyzed by high-performance liquid chromatography (HPLC, Agilent, USA) after extraction in methanol (Song et al. 2021).
The leaching of the BP-3 additive was determined by stirring MP/BP-3 fragments (1.0 mg L− 1) in a 2 L M4 medium for 48 h. The samples (2 mL) were collected periodically, filtered with 0.45 μm PTFE syringe filters (Whatman, USA), and analyzed for BP-3 concentration using HPLC, as mentioned above.
Multigenerational Chronic Toxicity Testing of MP/BP-3 Fragments
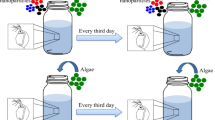
Daphnia magna clones, provided by the National Institute of Environmental Research (Republic of Korea), have been cultured since 2019 in accordance with the Organization for Economic Cooperation Development (OECD) Test Guideline 211 (OECD 2012). Daphnids were incubated in 2 L of M4 medium at 20 ± 0.1 ℃ with a light/dark cycle of 16:8 h, and the medium renewal was every 3 days. The daphnids were fed with the freshwater algae, Chlorella vulgaris (5.0 × 105 cells mL− 1; Aquanet, Republic of Korea) daily. Female neonates (< 24 h old and ≥ 3rd brood) were used to maintain the laboratory culture.
Two generational chronic toxicity tests (n = 15) were conducted in compliance with the OECD Daphnia magna reproduction test (OECD 2012). The third brood female neonates under 24 h old produced by same-aged mothers were collected from the laboratory culture for F0 generation. Each daphnid was grown in a 100-mL glass beaker containing 50 mL M4 medium under the same conditions as the culture (20 ± 0.1 ℃ and light/dark cycle 16:8 h). The daphnids were fed with 5.0 × 105 cells mL− 1 of C. vulgaris every day, and the medium was refreshed every two days. Only the parental generation (F0) was exposed to 1.0 mg L− 1 of MP/BP-3 fragments at neonate (< 24 h old) and adult (5 d old) stages. To compare the somatic growth and reproduction of daphnids at the same age (21 d old), neonate and adult groups were exposed for 21 d and 16 d, respectively. For recovery in the F1 generation, the first (F1B1) and third (F1B3) brood neonates were grown in a clean M4 medium. Growth (adult and offspring body length) and reproduction (number of offspring per female, days to first brood) were determined for 21 d.
Statistical Analysis
Statistical analyses were performed with SAS version 9.4 (SAS Institute Inc., Cary, NC, USA). One-way analysis of variance (ANOVA) and post-hoc Tukey’s honest significance test were used to test the significance of differences (p < 0.05) in neonate and adult groups. Student’s t-tests were used to measure significant differences (p < 0.05) in the first and third brood groups.
Results and Discussion
Physiochemical Properties of MP/BP-3 Fragments
The shape of synthesized MP/BP-3 fragments was irregular thin pointy flakes (Fig. 1). The size of the fragments was 18.23 ± 8.02 μm (n = 60), which is an edible size for D. magna (Rehse et al. 2016; Frydkjær et al. 2017; Canniff and Hoang 2018). The amount of BP-3 in MP/BP-3 fragments was measured as 2.89 ± 0.20% w/w. In the M4 medium, BP-3 was gradually released from MP/BP-3 fragments(Fig. 2), thus, the leaching was up to 49.1% for 48 h. The leaching of BP-3 can further aggravate the toxicity of MP fragments in D. magna (Na et al. 2021).

Scanning electron microscopy (SEM) image of (a) MP/BP-3 fragments and (b) BP-3 (2.89 ± 0.20% w/w) leaching from MP/BP-3 fragments in M4 medium (1.0 mg L− 1) at 20 ℃ for 48 h. All data are expressed as mean ± standard deviation (n = 3)
Different Chronic Toxicity and Maternal Effects in Neonate and Adult Exposure Groups
MP/BP-3 fragments significantly (p < 0.05) reduced the growth (body length) and reproduction (number of offspring per female) in the adult group of F0 generation relative to the control (Fig. 2), but not in the neonate group. Our previous study reported that pristine PE MP fragments (44.39 ± 11.16 μm) were not acutely toxic to neonates (< 24 h old) but toxic to juveniles (4 d old) (Song et al. 2021). Additionally, Liu et al. (2018) demonstrated that the oldest daphnids (21 d old) were the most sensitive to polystyrene nanoplastics compared to younger individuals (1, 4, 7, and 14 d old). Considering that the filtering and feeding rate of D. magna increase with body size (McMahon 1965), faster and greater uptake of adult daphnids may aggravate the chronic toxicity of MP/BP-3 fragments. However, the bioaccumulation kinetics of MP/BP-3 fragments in D. magna should be investigated to evaluate the toxicity difference between neonates and adult daphnids. On the other hand, D. magna, in its early life stage, may have more possibility of acclimation to MP/BP-3 stress (Klerks and Weis 1987; Dietrich et al. 2010).
Several studies have reported the negative effects of MPs on D. magna reproduction and growth (Martins and Guilhermino 2018; An et al. 2021; Trotter et al. 2021). For instance, An et al. (2021) demonstrated that polyethylene MP fragments (17.23 ± 3.43 μm) reduced the total number of offspring and adult body length in D. magna. Ingestion of MP particles can hinder the feeding activities of organisms and decrease the energy budget for reproduction and growth (Cole et al. 2013; Murphy and Quinn 2018). Moreover, the irregular shape of MP fragments can result in interstitial damage and a longer retention time in the gut (An et al. 2021; Silva et al. 2021). Recent studies emphasize the potential harm of plastic additives in MPs (Schrank et al. 2019; Zimmermann et al. 2020; Koelmans et al. 2022). BP-3 is known to act as an endocrine-disrupting chemical (Kim and Choi 2014; Wang et al. 2016), possibly by mimicking the ecdysone hormone (Ozáez et al. 2014). Ecdysone is a type of steroid hormone that regulates molting and development in insects and invertebrates, including D. magna (Pan et al. 2021). In particular, Lambert et al. (2021) observed that BP-3 significantly altered the expression of genes related to endocrine activity in D. magna, leading to a significant decrease in the number of offspring and body length. Song et al. (2021) also reported that BP-3 significantly decreased the embryonic development of D. magna.

Chronic toxicity and maternal effect of MP/BP-3 fragments in the neonate and adult Daphnia magna in terms of (a) body length and (b) number of offspring per female. F0 and F1 indicate parental and first generation, respectively, and B3 indicates the third brood. Only the F0 generation (neonate < 24 h and adult 5 d) was exposed to MP/BP-3 fragments (1.0 mg L− 1) until 21 d old, and F1 generation (third brood) was recovered in M4 medium (control) for 21 d. All data are expressed as mean ± standard deviation (n = 15). Lowercase letters indicate significant differences (p < 0.05) among the three treatment groups
Parental exposure to MP/BP-3 fragments did not influence the life-history traits of the F1 generation in the neonate group (F1B3). However, the number of offspring significantly (p < 0.05) decreased in the adult group of the F1 generation relative to the F1 control (Fig. 2b), indicating a maternal effect of MP/BP-3 fragments. Parental exposure in the adult group may influence the fitness of progeny (Beyer and Hambright 2017; Shaw et al. 2017), possibly leading to reduced reproduction in the F1 generation. In addition, the adverse effect of MPs may be exacerbated since the BP-3 additive is known to hinder development in D. magna embryos (Song et al. 2021). Moreover, the embryos (F1 generation) in the brood chamber can be directly exposed to MP/BP-3 fragments during parental exposure. Several studies have reported that MPs were found in the brood chamber (Guilhermino et al. 2021) and in developing embryos (Rosenkranz et al. 2009) of daphnids.
Different Maternal Effects in the First and Third Brood of Neonate Exposure Group
As discussed in the previous section, parental exposure of MP/BP-3 fragments to neonates did not influence the life-history traits of the third brood (F1B3). However, this exposure significantly (p < 0.05) increased the body length and offspring number of the first brood of D. magna (F1B1) relative to the control (Fig. 3). Several studies have reported increased maternal effect on the first brood of D. magna. For instance, Ellis et al. (2020) demonstrated that the first brood of offspring was the most sensitive to parental exposure to silver and titanium oxide nanomaterials. Additionally, Liu et al. (2012) reported that the third brood offspring showed enhanced recovery after parental exposure to the pesticide buprofezin compared to the first brood.

Maternal effect of MP/BP-3 fragments in the first and third brood of Daphnia magna in terms of (a) body length, (b) number of offspring per female, (c) days to first brood, and (d) offspring body length. F0 and F1 indicate parental and first generation, respectively. B1 and B3 indicate the first and third brood, respectively. Only the F0 generation (neonate < 24 h) was exposed to MP/BP-3 fragments (1.0 mg L− 1) until 21 d old, and the F1 generation was recovered in M4 medium (control) for 21 d. All data are expressed as mean ± standard deviation (n = 15). Asterisks indicate significant differences (p < 0.05) between the control and treatment groups
The first brood of D. magna is generally less stable than subsequent broods (Kim et al. 2014). The number of offspring of the F1B1 control (46.15 ± 16.09) was significantly (p < 0.05) lower than in the F1B3 control (71.2 ± 10.49), which is far lower than the number (≥ 60) recommended in the OECD guideline (OECD 2012). However, parental exposure to MP/BP-3 fragments increased the number of offspring per female (71.2 ± 13.74) to the control level in the F1B1 recovery group. Under this stress condition, D. magna may use more energy for growth and reproduction than for defense (Costantini 2014; Im et al. 2019). Indeed, the F1B1 recovery group reached primiparity significantly faster (10.6 ± 0.51 d) than the control group (12 ± 1.22 d), leading to higher somatic growth and reproduction. However, the size of the F1B1 offspring (818.27 ± 19.00 μm) was significantly smaller than that of the control group (828.95 ± 17.96 μm). Given that offspring fitness increases with size (Smith and Fretwell 1974; Moran and Emlet 2001), the above findings suggest that the first brood of D. magna responded to MP/BP-3 stress by accelerating somatic growth and reproduction, which resulted in less fit offspring.
Conclusion
Contrary to the initial hypothesis, MP/BP-3 fragments induced greater chronic toxicity and maternal effect in adults than in neonates, inhibiting the growth and reproduction in both F0 and F1 generations of D. magna. On the other hand, the maternal effect of MP/BP-3 fragments was higher in the first brood neonates than in the third brood neonates, enhancing growth and reproduction in the F1 generation. This study demonstrates different MP toxicity and maternal effects in D. magna at different ages and broods, providing insight into the ecological risk of MPs. However, further studies are required over multiple generations to identify the transgenerational effect of MPs containing plastic additives.
References
Ajith N, Arumugam S, Parthasarathy S, Manupoori S, Janakiraman S (2020) Global distribution of microplastics and its impact on marine environment—A review. Environ Sci Pollut Res Int 27(21):25970–25986. https://doi.org/10.1007/s11356-020-09015-5
An D, Na J, Song J, Jung J (2021) Size-dependent chronic toxicity of fragmented polyethylene microplastics to Daphnia magna. Chemosphere 271:129591. https://doi.org/10.1016/j.chemosphere.2021.129591
Beyer JE, Hambright KD (2017) Maternal effects are no match for stressful conditions: a test of the maternal match hypothesis in a common zooplankter. Funct Ecol 31(10):1933–1940. https://doi.org/10.1111/1365-2435.12901
Botterell ZLR, Beaumont N, Dorrington T, Steinke M, Thompson RC, Lindeque PK (2019) Bioavailability and effects of microplastics on marine zooplankton: a review. Environ Pollut 245:98–110. https://doi.org/10.1016/j.envpol.2018.10.065
Bratkovics S, Wirth E, Sapozhnikova Y, Pennington P, Sanger D (2015) Baseline monitoring of organic sunscreen compounds along South Carolina’s coastal marine environment. Mar Pollut Bull 101(1):370–377. https://doi.org/10.1016/j.marpolbul.2015.10.015
Canniff PM, Hoang TC (2018) Microplastic ingestion by Daphnia magna and its enhancement on algal growth. Sci Total Environ 633:500–507. https://doi.org/10.1016/j.scitotenv.2018.03.176
Carve M, Nugegoda D, Allinson G, Shimeta J (2021) A systematic review and ecological risk assessment for organic ultraviolet filters in aquatic environments. Environ Pollut 268:115894. https://doi.org/10.1016/j.envpol.2020.115894
Cole M, Lindeque P, Fileman E, Halsband C, Goodhead R, Moger J, Galloway TS (2013) Microplastic ingestion by zooplankton. Environ Sci Technol 47(12):6646–6655. https://doi.org/10.1021/es400663f
Cole M, Lindeque P, Halsband C, Galloway TS (2011) Microplastics as contaminants in the marine environment: a review. Mar Pollut Bull 62(12):2588–2597. https://doi.org/10.1016/j.marpolbul.2011.09.025
Costantini D (2014) Oxidative stress and hormesis in evolutionary ecology and physiology. A marriage between mechanistic and evolutionary approaches, vol 362
De Sá LC, Oliveira M, Ribeiro F, Rocha TL, Futter MN (2018) Studies of the effects of microplastics on aquatic organisms: what do we know and where should we focus our efforts in the future? Sci Total Environ 645:1029–1039. https://doi.org/10.1016/j.scitotenv.2018.07.207
Dietrich S, Ploessl F, Bracher F, Laforsch C (2010) Single and combined toxicity of pharmaceuticals at environmentally relevant concentrations in Daphnia magna–A multigenerational study. Chemosphere 79(1):60–66. https://doi.org/10.1016/j.chemosphere.2009.12.069
Dusaucy J, Gateuille D, Perrette Y, Naffrechoux E (2021) Microplastic pollution of worldwide lakes. Environ Pollut 284:117075. https://doi.org/10.1016/j.envpol.2021.117075
Ellis LA, Kissane S, Lynch I (2020) Maternal responses and adaptive changes to environmental stress via chronic nanomaterial exposure: differences in inter and transgenerational interclonal broods of Daphnia magna. Int J Mol Sci 22(1):15. https://doi.org/10.3390/ijms22010015
Eltemsah YS, Bøhn T (2019) Acute and chronic effects of polystyrene microplastics on juvenile and adult Daphnia magna. Environ Pollut 254(A):112919. https://doi.org/10.1016/j.envpol.2019.07.087
Frydkjær CK, Iversen N, Roslev P (2017) Ingestion and egestion of microplastics by the cladoceran Daphnia magna: Effects of regular and irregular shaped plastic and sorbed phenanthrene. Bull Environ Contam Toxicol 99(6):655–661. https://doi.org/10.1007/s00128-017-2186-3
Ginjupalli GK, Baldwin WS (2013) The time- and age-dependent effects of the juvenile hormone analog pesticide, pyriproxyfen on Daphnia magna reproduction. Chemosphere 92(9):1260–1266. https://doi.org/10.1016/j.chemosphere.2013.04.061
Gola D, Kumar Tyagi PK, Arya A, Chauhan N, Agarwal M, Singh SK, Gola S (2021) The impact of microplastics on marine environment: a review. Environ Nanotechnol Monit Manag 16:100552. https://doi.org/10.1016/j.enmm.2021.100552
Guilhermino L, Martins A, Cunha S, Fernandes JO (2021) Long-term adverse effects of microplastics on Daphnia magna reproduction and population growth rate at increased water temperature and light intensity: combined effects of stressors and interactions. Sci Total Environ 784:147082. https://doi.org/10.1016/j.scitotenv.2021.147082
Gunaalan K, Fabbri E, Capolupo M (2020) The hidden threat of plastic leachates: a critical review on their impacts on aquatic organisms. Water Res 184:116170. https://doi.org/10.1016/j.watres.2020.116170
Hahladakis JN, Velis CA, Weber R, Iacovidou E, Purnell P (2018) An overview of chemical additives present in plastics: Migration, release, fate and environmental impact during their use, disposal and recycling. J Hazard Mater 344:179–199. https://doi.org/10.1016/j.jhazmat.2017.10.014
Im H, Achar JC, Shim T, Jung J (2022) Elevated temperature alleviates benzophenone-3 toxicity in Daphnia magna. Aquat Toxicol 242:106047
Im H, Samanta P, Na J, Jung J (2019) Time-dependent responses of oxidative stress, growth, and reproduction of Daphnia magna under thermal stress. Bull Environ Contam Toxicol 102(6):817–821. https://doi.org/10.1007/s00128-019-02613-1
Kim HY, Yu S, Jeong TY, Kim SD (2014) Relationship between trans-generational effects of tetracycline on Daphnia magna at the physiological and whole organism level. Environ Pollut 191:111–118. https://doi.org/10.1016/j.envpol.2014.04.022
Klerks PL, Weis JS (1987) Genetic adaptation to heavy metals in aquatic organisms: a review. Environ Pollut 45(3):173–205. https://doi.org/10.1016/0269-7491(87)90057-1
Koelmans AA (2015) Modeling the role of microplastics in bioaccumulation of organic chemicals to marine aquatic organisms. A critical review. Mar anthropogenic litter 309–324. https://doi.org/10.1007/978-3-319-16510-3_11
Koelmans AA, Redondo-Hasselerharm PE, Nor NHM, de Ruijter VN, Mintenig SM, Kooi M (2022) Risk assessment of microplastic particles. Nat Rev Mater 7(2):138–152. https://doi.org/10.1038/s41578-021-00411-y
Lambert FN, Gracy HR, Gracy AJ, Yoon SH, Scott RW, Rincon DM, Vulpe CD (2021) Effects of ultraviolet-filters on Daphnia magna development and endocrine-related gene expression. Aquat Toxicol 238:105915. https://doi.org/10.1016/j.aquatox.2021.105915
LaMontagne JM, McCauley E (2001) Maternal effects in Daphnia: what mothers are telling their offspring and do they listen? Ecol Lett 4(1):64–71. https://doi.org/10.1046/j.1461-0248.2001.00197.x
Lee H, Shim WJ, Kwon JH (2014) Sorption capacity of plastic debris for hydrophobic organic chemicals. Sci Total Environ 470–471:1545–1552. https://doi.org/10.1016/j.scitotenv.2013.08.023
Li C, Busquets R, Campos LC (2020) Assessment of microplastics in freshwater systems: a review. Sci Total Environ 707:135578. https://doi.org/10.1016/j.scitotenv.2019.135578
Liu Y, Qi S, Zhang W, Li X, Qiu L, Wang C (2012) Acute and chronic toxicity of buprofezin on daphnia magna and the recovery evaluation. Bull Environ Contam Toxicol 89(5):966–969. https://doi.org/10.1007/s00128-012-0802-9
Liu Z, Cai M, Yu P, Chen M, Wu D, Zhang M, Zhao Y (2018) Age-dependent survival, stress defense, and AMPK in Daphnia pulex after short-term exposure to a polystyrene nanoplastic. Aquat Toxicol 204:1–8. https://doi.org/10.1016/j.aquatox.2018.08.017
Liu J, Yang H, Meng Q, Feng Q, Yan Z, Liu J, Zhou Z (2022) Intergenerational and biological effects of roxithromycin and polystyrene microplastics to Daphnia magna. Aquat Toxicol 248:106192. https://doi.org/10.1016/j.aquatox.2022.106192
Martins A, Guilhermino L (2018) Transgenerational effects and recovery of microplastics exposure in model populations of the freshwater cladoceran Daphnia magna Straus. Sci Total Environ 631–632:421–428. https://doi.org/10.1016/j.scitotenv.2018.03.054
McMahon JW (1965) Some physical factors influencing the feeding behavior of Daphnia magna Straus. Can J Zool 43(4):603–611. https://doi.org/10.1139/z65-060
Moran AL, Emlet RB (2001) Offspring size and performance in variable environments: field studies on a marine snail. Ecology 82(6):1597–1612. https://doi.org/10.1890/0012-9658(2001)082[1597:OSAPIV]2.0.CO;2
Mousseau TA, Fox CW (1998) The adaptive significance of maternal effects. Trends Ecol Evol 13(10):403–407. https://doi.org/10.1016/S0169-5347(98)01472-4
Murphy F, Quinn B (2018) The effects of microplastic on freshwater Hydra attenuata feeding, morphology & reproduction. Environ Pollut 234:487–494. https://doi.org/10.1016/j.envpol.2017.11.029
Muyssen BTA, Janssen CR (2007) Age and exposure duration as a factor influencing Cu and Zn toxicity toward Daphnia magna. Ecotoxicol Environ Saf 68(3):436–442. https://doi.org/10.1016/j.ecoenv.2006.12.003
Martín-Folgar R, Aquilino M, Ozáez I, Martínez-Guitarte JL (2018) Ultraviolet filters and heat shock proteins: effects in Chironomus riparius by benzophenone-3 and 4-methylbenzylidene camphor. Environ Sci Pollut Res 25:333–344. https://doi.org/10.1007/s11356-017-0416-1
Na J, Song J, Achar JC, Jung J (2021) Synergistic effect of microplastic fragments and benzophenone-3 additives on lethal and sublethal Daphnia magna toxicity. J Hazard Mater 402:123845. https://doi.org/10.1016/j.jhazmat.2020.123845
Ozáez I, Martínez-Guitarte JL, & Morcillo G (2014) The UV filter benzophenone 3 (BP-3) activates hormonal genes mimicking the action of ecdysone and alters embryo development in the insect Chironomus riparius (Diptera). Environ Pollut 192:19–26. https://doi.org/10.1016/j.envpol.2014.04.038
OECD (2012) Test No. 211: Daphnia magna reproduction test, OECD guidelines for the testing of chemicals, Sect. 2. OECD Publishing, Paris. https://doi.org/10.1787/9789264185203-en
Rehse S, Kloas W, Zarfl C (2016) Short-term exposure with high concentrations of pristine microplastic particles leads to immobilisation of Daphnia magna. Chemosphere 153:91–99. https://doi.org/10.1016/j.chemosphere.2016.02.133
Rosenkranz P, Chaudhry Q, Stone V, Fernandes TF (2009) A comparison of nanoparticle and fine particle uptake by Daphnia magna. Environ Toxicol Chem 28(10):2142–2149. https://doi.org/10.1897/08-559.1
Schrank I, Trotter B, Dummert J, Scholz-Böttcher BM, Löder MGJ, Laforsch C (2019) Effects of microplastic particles and leaching additive on the life history and morphology of Daphnia magna. Environ Pollut 255(2):113233. https://doi.org/10.1016/j.envpol.2019.113233
Shaw JLA, Judy JD, Kumar A, Bertsch P, Wang MB, Kirby JK (2017) Incorporating transgenerational epigenetic inheritance into ecological risk assessment frameworks. Environ Sci Technol 51(17):9433–9445. https://doi.org/10.1021/acs.est.7b01094
Shen C, Wei J, Wang T, Wang Y (2019) Acute toxicity and responses of antioxidant systems to dibutyl phthalate in neonate and adult Daphnia magna. PeerJ 7:e6584. https://doi.org/10.7717/peerj.6584
Silva CJM, Patrício Silva ALP, Campos D, Machado AL, Pestana JLT, Gravato C (2021) Oxidative damage and decreased aerobic energy production due to ingestion of polyethylene microplastics by Chironomus riparius (Diptera) larvae. J Hazard Mater 402:123775. https://doi.org/10.1016/j.jhazmat.2020.123775
Smith CC, Fretwell SD (1974) The optimal balance between size and number of offspring. Am Nat 108(962):499–506. https://doi.org/10.1086/282929
Song J, Kim C, Na J, Sivri N, Samanta P, Jung J (2022) Transgenerational effects of polyethylene microplastic fragments containing benzophenone-3 additive in Daphnia magna. J Hazard Mater 436:129225. https://doi.org/10.1016/j.jhazmat.2022.129225
Song J, Na J, An D, Jung J (2021) Role of benzophenone-3 additive in chronic toxicity of polyethylene microplastic fragments to Daphnia magna. Sci Total Environ 800:149638. https://doi.org/10.1016/j.scitotenv.2021.149638
Tao J, Bai C, Chen Y, Zhou H, Liu Y, Shi Q, Pan W, Dong H, Li L, Xu H, Tanguay R (2020) Environmental relevant concentrations of benzophenone-3 induced developmental neurotoxicity in zebrafish. Sci Total Environ 721:137686. https://doi.org/10.1016/j.scitotenv.2020.137686
Trotter B, Wilde MV, Brehm J, Dafni E, Aliu A, Arnold GJ, Fröhlich T, Laforsch C (2021) Long-term exposure of Daphnia magna to polystyrene microplastic (PS-MP) leads to alterations of the proteome, morphology and life-history. Sci Total Environ 795:148822. https://doi.org/10.1016/j.scitotenv.2021.148822
Wagner ND, Simpson AJ, Simpson MJ (2017) Metabolomic responses to sublethal contaminant exposure in neonate and adult Daphnia magna. Environ Toxicol Chem 36(4):938–946. https://doi.org/10.1002/etc.3604
Wang J, Pan L, Wu S, Lu L, Xu Y, Zhu Y, Guo M, Zhuang S (2016) Recent advances on endocrine disrupting effects of UV filters. Int J Environ Res Public Health 13(8):782. https://doi.org/10.3390/ijerph13080782
Zimmermann L, Göttlich S, Oehlmann J, Wagner M, Völker C (2020) What are the drivers of microplastic toxicity? Comparing the toxicity of plastic chemicals and particles to Daphnia magna. Environ Pollut 267:115392. https://doi.org/10.1016/j.envpol.2020.115392
Acknowledgements
This study was supported by a grant from the National Research Foundation of Korea (NRF), funded by the Korean government (NRF-2019R1A2C1002890).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kim, C., Song, J. & Jung, J. Maternal Effect of Polyethylene Microplastic Fragments Containing Benzophenone-3 in Different Ages and Broods of Daphnia Magna. Bull Environ Contam Toxicol 110, 66 (2023). https://doi.org/10.1007/s00128-023-03705-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00128-023-03705-9