
Abstract
Although phosphatized bryozoans have been described recently from the early Cambrian, the first unequivocal bryozoan fossils with hard skeletons are known from the Ordovician. Recent discoveries of bryozoans in the early Ordovician (Tremadocian) of South China have greatly expanded our understanding of the diversification of these colonial lophophorates. In particular, the Fenhsiang Formation of Late Tremadocian age (Migneintian) in Hubei Province is proving to be particularly rich in bryozoans. Here we record 24 species, including several yet to be formally described, belonging to 18 genera and four palaeostomate suborders (Esthonioporata, Cystoporata, Trepostomata, and Cryptostomata). Bryozoan diversity in the Fenhsiang Formation matches levels more typical of younger faunas of Middle Ordovician age. The presence of diverse and morphologically disparate taxa close to the base of the Ordovician suggests rapid diversification following the first appearance of bryozoans with calcified skeletons, and/or the existence of as yet unknown biomineralized bryozoans in the Cambrian.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bryozoans have recently been described for the first time from the Cambrian (Zhang et al. 2021), confirming molecular clock analyses that implied the divergence of bryozoans from other Metazoa in the late Neoproterozoic or early Cambrian (Erwin et al. 2011; Erwin 2015).
The early Cambrian species Protomelission gatehousei Brock and Cooper, 1993 has been shown to possess features known from both stenolaemates and ctenostomes. It formed an erect bifoliate colony comprising box-shaped zooids with an unmineralized body plan. Known from Australia and China, this species may potentially represent a stem-group bryozoan (Zhang et al. 2021).
There is still however a 35 Myr gap until the first known appearance of bryozoans with mineralized skeletons (Ma et al. 2015; Taylor et al. 2013). The absence of bryozoans with hard skeletons from the Cambrian fossil record is anomalous given the fact that most species belonging to this benthic colonial phylum have skeletons of low-magnesium calcite that are diagenetically stable, which accounts for the rich post-Cambrian fossil record of these suspension-feeding lophophorates.
The oldest known bryozoan with a calcified skeleton is Prophyllodictya simplex Ma et al., 2015 from the Nantsinkuan Formation (Cressagian) of South China. This has erect bifoliate branches of a seemingly advanced morphological grade that is inconsistent with the ‘weedy’ morphology previously expected of a primitive bryozoan (Larwood and Taylor 1979; Taylor 2020). The poor record of early bryozoan evolution of bryozoans may in part be explained by most early bryozoans lacking mineralized skeletons, as in Protomelission, while their absence from the classic Cambrian Lägerstatten could be due to the thinness of their organic cuticles compared to those of the arthropods and worms that dominate in these muddy sediments not well-suited for bryozoan colonization (Taylor 2020).
Here we review the composition and importance of the oldest known, multispecies bryozoan fauna. Although yet to be fully described, this fauna from the Fenhsiang Formation (Late Tremadocian, Migneintian Stage) is important in showing the presence of taxonomically diverse and morphologically disparate bryozoans soon after the first appearance of calcified bryozoans in the fossil record. This points to an extremely rapid radiation of bryozoans in the earliest Ordovician, substantial gaps in the fossil record of bryozoans from the early Cambrian to early Ordovician, or a combination of these two.
Geological setting
The bryozoans used in this study have been collected over a period of 10 years from the Fenhsiang Formation of Hubei Province and Chongqing, South China (Fig. 1), and are deposited in the Nanjing Institute of Geology and Palaeontology (abbreviated NIGPAS). The investigated area is covered by a well-defined, continuous sequence of Ordovician sedimentary rocks, widely exposed around the Huangling Anticline in the Three Gorges area of Hubei Province. The Fenhsiang Formation is c. 70 m thick in the south and c. 20 m thick in the west of the anticline, and has been subdivided into the Lower Limestone and Upper Limestone members by Zeng et al. (1987). The upper Limestone Member contains limestones intercalated with shales, which are a rich source of bryozoans. The age of the Fenhsiang Formation is interpreted to be late Tremadocian (Migneintian) based mainly on conodonts (Xia et al. 2007; Zhang et al. 2009; Wu et al. 2020). Deposition occurred on the Yangtze Platform at a palaeolatitude estimated as 19°S.

a Locations of Fenhsiang Formation sections in the Ordovician of South China. b Outline of lithostratigraphy of the Pengshui section showing the distribution of bryozoans in the Fenhsiang Formation. Fm., formation; HHY, Honghuayuan
The Fenhsiang Formation of the Chenjiahe section in Pengshui conformably overlies the early Tremadocian Nantsinkuan Formation and is overlain by the early Floian Hunghuayuan Formation. It consists of dark-grey to grey skeletal and peloidal limestones and shales belonging to the Upper Limestone Member, which contains lithistid sponges, bryozoans, pelmatozoan echinoderms, graptolites, and microbialites (Zeng et al. 1987). Also present are the oldest known metazoan reefs that were formed by lithistid sponges and esthonioporate bryozoans (Adachi et al. 2011, 2012; Cuffey et al. 2013), indicating a shallow water depositional environment or shoal on the inner shelf and platform margin. Fossil preservation in the reefal facies indicates in situ deposition without significant post-mortem transportation.
Fenhsiang bryozoan fauna
A total of 24 species distributed among 18 genera have been found in the Fenhsiang Formation (Table 1). This must be regarded as a minimum estimate of diversity as it is anticipated that further collecting will yield additional taxa. All of the Fenhsiang bryozoans are palaeostomates (Ma et al. 2014) and can be assigned to four (Esthonioporata, Cystoporata, Trepostomata, and Cryptostomata) of the five Early Palaeozoic orders in this superorder (Fig. 2); only the order Fenestrata is not yet recorded in the Fenhsiang Formation (the oldest fossil fenestrate remains Alwynopora from the Dapingian: Taylor and Curry 1985). Seven of the 24 species have been described formally. Detailed descriptions of the undescribed species will be given in a future publication.

Stratigraphical ranges of the major bryozoan groups that occur in the Fenhsiang Formation (Tremadocian) of South China and elsewhere in early and middle Ordovician. Black circles indicate previously known earliest occurrences from Baltics; solid grey circles indicate previously known earliest occurrences from North America; triangles indicate previously known earliest occurrences from Novaga Zemlya, Russia; stars indicate new occurrences reported here from South China. Based on the data of Ernst et al. (2014), Koromyslova (2011), Fedorov et al. (2017), Koromyslova and Fedorov (2021), and the current study
The Fenhsiang Formation bryozoan fauna contains equal numbers (seven species) of cystoporates, trepostomes, and cryptostomes, with fewer esthonioporines (four species). Cystoporates have not been reported previously from the Fenhsiang Formation and their occurrence in this formation is the earliest fossil record for the order. The majority of the Fenhsiang bryozoans represent the oldest known occurrences of their genera, pre-dating Floian and Dapingian examples from the Latorp and Volkhov horizons of the Leningrad region which contain well-known examples of early bryozoans (Koromyslova 2011).
Esthonioporata
Esthonioporates are morphologically primitive among palaeostomates in lacking differentiation between endozones and exozones (Ma et al. 2014). The oldest known esthonioporates globally occur in the Fenhsiang Formation in which three species of Nekhorosheviella and one of Dianulites can be recognized (Fig. 3a). The first of these genera is important in constructing small reefs comprising bifurcating columns formed of stacked domes (Adachi et al. 2011, 2013). Esthonioporata is the least diverse order of bryozoans in the Tremadocian of South China and is better represented in the slightly younger (Floian) deposits of Baltica (Pushkin and Popov 1999; Koromyslova 2011).

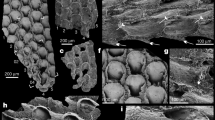
Thin sections showing examples of the diverse bryozoans from the Fenhsiang Formation (Tremadocian) of South China. a Esthoniopore Dianulites sp. NIGP 177,964. b Fistuliporine cystoporate Profistulipora sp. NIGP 177,965. c Ceramoporine Ceramopora sp. NIGP 177,966. d Ceramoporine cystoporate Haplotrypa sp. NIGP 177,967. e Ceramoporine Amsassipora sp. NIGP 177,968. f Amplexoporine trepostome Orbignyella sp. NIGP 177,969. g Ptilodictyine cryptostome Prophyllodictya sp. NIGP 177,970. h Rhabdomesine Vercoclema sp. NIGP 177,971. i Ptilodictyine Trepocryptopora sp. NIGP 177,972. j Trepostome Goldfussitrypa sp. NIGP 177,973. k Arthrostylid rhabdomesine Nematopora ovalis. NIGP 177,974. K Scales: a, f–h, j, 500 µm; b–e, 200 µm; i, k, 100 µm. NIGP (= NIGPAS), Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences
Cystoporata
Cystoporates were interpreted by Ma et al. (2014) as the sister group of esthonioporates in a cladistic analysis. They are one of the most numerous and diverse suborders in the Fenhsiang Formation, but most are known only in the Pengshui section. The Fenhsiang Formation cystoporates have colony-forms varying from discoidal and encrusting to hemispherical and massive. Of the two cystoporate suborders, Fistuliporina are more diverse than Ceramoporina and include species of Anolotichia, Hennigopora, Profistulipora, and Fistulipora (Fig. 3b; Fig. 4a-c), the latter comprising species of Ceramopora, Amsassipora, and Haplotrypa (Fig. 3c-e). Prior to the current study, the earliest cystoporates were recorded from the Floian of Russia (Astrova 1965; Koromyslova 2011) and North America (Ross 1966).

Thin sections showing vesicles (a–c), mesozooecia (d), and exilazooecia (e) in bryozoans from the Fenhsiang Formation. a Profistulipora sp. NIGP 177,975. b Hennigopora sp. NIGP 177,976. c Fistulipora sp. NIGP 177,977. d Undetermined halloporine trepostome NIGP 177,978. e Orbignyella sp. NIGP 177,979. Scale bars: a, 200 µm; b, c, e, 100 µm; d, 500 µm
The Fenhsiang ceramoporines encompass forms with unevenly thickened walls (e.g., Haplotrypa) (Fig. 3d), and others showing differentiation of endozones and exozones (Amsassipora) (Fig. 3e). Fistuliporines from the Fenhsiang Formation exhibit a variety of vesicular skeletons, including vesicles of tubular form in Anolotichia (Fig. 4a) and Profistulipora (Fig. 3b), and box-like shape in Hennigopora sp. (Fig. 4b), as well as examples containing styles in their roofs as in a species of genus Fistulipora (Fig. 4c). Previously, Profistulipora from the Floian of Novaya Zemlya with tube-like vesicular skeletons containing dense and relatively regular diaphragms was considered to be the most primitive morphology (Astrova 1965; Yaroshinskaya 1967), with other morphologies not appearing until the Late Ordovician. The earlier occurrence of disparate vesicle morphologies synchronously in the Fenhsiang Formation questions this interpretation.
Trepostomata
Trepostomes are the most important and diverse bryozoan order globally in the Early Palaeozoic. Both trepostome suborders (Halloporina and Amplexoporina) are present in the Fenhsiang Formation. These include the endemic amplexoporine genus Orbiramus of which three species were described by Xia et al. (2007). In the current study, we also found in the Fenhsiang Formation several other amplexoporine (Orbignyella, Goldfussitrypa, and Monotrypa) (Fig. 3f, j), as well as halloporine genera (Bythopora, and one undeterminated Halloporina) (Fig. 4d), pre-dating their first occurrences elsewhere in the world.
Cryptostomata
Cryptostomes were found to be the sister group of trepostomes in the cladistic study of Ma et al. (2014). Bryozoans with transitional morphologies between these two orders are evident in the cryptostome order Intraporidae (Goryunova and Lavrentjeva 1993). Cryptostomata is traditionally divided into two suborders, Ptilodictyina and Rhabdomesina, both of which were found to be diverse and locally abundant in the Fenhsiang Formation. The rhinidictyid ptilodictyine species Prophyllodictya prisca Xia in Xia et al., 2007 was described previously by Xia et al. (2007). In the current study, a second species of Prophyllodictya with large ‘capillaries’ in the exozonal walls of the autozooecia was found in the Pengshui section (Fig. 3g). The intraporid ptilodictyine genus Treptocryptopora, first reported in Middle Ordovician rocks of central China (Yang 1957), is represented in the Fenhsiang Formation by a species having fewer diaphragms and exilazooecia than the Middle Ordovician species (Fig. 3i). Two rhabdomesine genera found in the Fenhsiang Formation are worth highlighting. The presence of Veroclema in the Fenhsiang Formation (Fig. 3h) extends its range and palaeogeographical distribution from the Llandoverian (Lower Silurian) of the USA to the Tremadocian of Central China. More common in the Fenhsiang Formation are arthrostylid rhabdomesines, including Nematopora (Fig. 3k), pre-dating the previously recorded first appearance of this family in the Middle Ordovician of the Baltic area.
Discussion
The Fenhsiang Formation of South China contains the richest bryozoan fauna known from the Tremadocian. With 24 species distributed between 18 genera, its diversity more than doubles the number of Tremadocian taxa used in previously published analyses of Ordovician bryozoan dynamics (Ernst 2018; Hageman and Ernst 2019) (Fig. 5). The formation is notably more diverse than another early bryozoan fauna, the slightly younger Latorp Horizon (Floian) of the Leningrad region (Baltica) in which seven genera and 13 species were recorded by Koromyslova (2011). Several major taxa of bryozoans have their oldest known occurrences in the Fenhsiang Formation. The formation contains the earliest examples of the palaeostomate orders Esthonioporata, Cystoporata, and Trepostomata, as well as the earliest cryptostomes of the order Rhabdomesina. The only palaeostomate order not found so far is Fenestrata. In addition, no examples are known in the Fenhsiang Formation of the stenolaemate order Cyclostomata or of the gymnolaemate order Ctenostomata, both of which occur in younger stages of the Ordovician.

Range-through bryozoan generic diversity in the Ordovician, with the new data from this study added to that of Ernst (2018). Hirn, Hirnantian
The newly revealed diversity of bryozoans in the Fenhsiang Formation is paralleled by significant morphological disparity. Colony-forms include six, possibly seven, of the nine major colony-forms distinguished for bryozoans by Taylor and James (2013): encrusting, dome-shaped, palmate, foliose, robust ramose, delicate ramose, and possibly articulated. The missing colony-forms are free-living (which is unknown among Palaeozoic bryozoans) and fenestrate. Space-filling polymorphs present among the Fenhsiang bryozoans include exilazooecia and mesozooecia, while interzooecial vesicular skeleton is also present in some species.
Hageman and Ernst (2019) noted the poor sampling of Tremadocian bryozoans based on the occurrence of numerous families and genera indicating the presence of ghost lineages in the earliest Ordovician. The newly reported taxa in the Fenhsiang bryozoan fauna only add to the number of ghost lineages. There are three non-exclusive explanations for the sudden appearance of diverse and disparate bryozoans in the Early Ordovician fossil record. The first is that bryozoans possessing calcified skeletons diversified extremely rapidly during the earliest Ordovician, with the swift appearance of numerous colony- and zooid-level morphological traits, many of which define higher taxa. The second postulates the presence of undiscovered calcified bryozoans in the Cambrian, which is plausible in view of the difficulty of observing bryozoans in well-lithified carbonates especially when these are dolomitized as on the Yangtze Platform (Taylor 2020, p. 214). A third factor could be the multiple acquisition of calcite skeleton in bryozoans consequent upon geochemical changes in the marine environment. However, the transition from aragonite to calcite seas predated the appearance of diverse bryozoans with calcareous skeletons, having occurred early in the Cambrian (Zhuravlev & Wood 2008; Porter 2010). Further fossil finds are needed to evaluate the roles of these three factors.
The spectacular diversification of bryozoans during the Ordovician forms part of the ‘Great Ordovician Biodiversification Event’ (GOBE) (see Harper 2006) that involved numerous marine invertebrate groups, correlated with increased tiering and biotic interactions among benthic suspension feeders, and saw the emergence of Sepkoski’s (1984) ‘Paleozoic evolutionary fauna’. For bryozoans, taxonomic and morphological diversification was shown by Hageman and Ernst (2019) to have increased sharply at the Early-Middle Ordovician transition. Our new data concerning the Fenhsiang bryozoan fauna points to the possibility of an earlier onset of rapid diversification. These findings are consistent with a study of sclerobionts associated with bryozoans from the Fenhsiang Formation (Ma et al. 2021) showing the existence of complex ecological interactions in the Tremadocian at a very early stage in the GOBE.
References
Adachi N, Ezaki Y, Liu J (2011) Early Ordovician shift in reef construction from microbial to metazoan reefs. Palaios 26:106114
Adachi N, Ezaki Y, Liu J (2012) The oldest bryozoan reefs: a unique Early Ordovician skeletal framework construction. Lethaia 45:14–23
Adachi N, Liu J, Ezaki Y (2013) Early Ordovician reefs in South China (Chenjiahe section, Hubei Province): deciphering the early evolution of skeletal-dominated reefs. Facies 59:451–466
Astrova GG (1965) A new order of Paleozoic Bryozoa. Int Geol Rev 7:1622–1628
Brock GA, Cooper BJ (1993) Shelly fossils from the Early Cambrian (Toyonian) Wirrealpa, Aroona Creek, and Ramsay Limestones of South Australia. J Paleontol 67:758–787
Cuffey RJ, Xiang CT, Zhu ZD, Spjeldnaes N, Hu ZX (2013) The world’s oldest-known bryozoan reefs: Late Tremadocian, mid-Early Ordovician; Yichang. Central China Bryozoan Studies 143:13–27
Ernst A (2018) Diversity dynamics of Ordovician Bryozoa. Lethaia 51:198–206
Ernst A, Bogolepova OK, Hubmann B, Golubkova EY, Gubanov AP (2014) Dianulites (Trepostomata, Bryozoa) from the Early Ordovician of Severnaya Zemlya. Arctic Russia Geol Mag 151(02):328–338
Erwin DH, Laflamme M, Tweedt SM, Sperling EA, Pisani D, Peterson KJ (2011) The Cambrian conundrum: early divergence and later ecological success in the early history of animals. Science 334:1091–1097
Erwin DH (2015) Early metazoan life: divergence, environment and ecology. Philos Trans R Soc Lond B Biol Sci 370(1684):20150036. https://doi.org/10.1098/rstb.2015.0036
Fedorov PV, Koromyslova AV, Martha SO (2017) The oldest bryozoans of Baltoscandia from the lowermost Floian (Ordovician) of north-western Russia: two new rare, small and simple species of Revalotrypidae. PalZ 91:353–373
Hageman SJ, Ernst A (2019) The last phylum: occupation of Bryozoa morpho-ecospace (colony growth habits) during the early phase of the Great Ordovician Biodiversification Event. Palaeogeogr Palaeoclimatol Palaeoecol 534:109270
Harper DAT (2006) The Ordovician biodiversification: setting an agenda for marine life. Palaeogeogr Palaeoclimatol Palaeoecol 232:148–166
Koromyslova AV (2011) Bryozoans of the Latorp and Volkhov horizons (Lower-Middle Ordovician) of the Leningrad region. Paleontol J 45:887–980
Koromyslova AV, Fedorov PV (2021) The oldest bifoliate cystoporate and two other bryozoan taxa from the Dapingian (Middle Ordovician) of north-western Russia. J Paleontol 95:24–39
Larwood GP, Taylor PD (1979) Early structural and ecological diversification in the Bryozoa. In: House, M.R. (editor) Origin of major invertebrate groups: 209–234. Academic Press, London
Ma JY, Buttler CJ, Taylor PD (2014) Cladistic analysis of the ‘trepostome’ Suborder Esthonioporina and the systematics of Palaeozoic bryozoans. Studi Trent Sci Nat 94:153–161
Ma JY, Taylor PD, Xia FS, Zhan RB (2015) The oldest known bryozoan: Prophyllodictya (Cryptostomata) from the lower Tremadocian (Lower Ordovician) of Liujiachang, south-western Hubei, central China. Palaeontology 58:925–934
Ma JY, Taylor PD, Buttler CJ (2021) Sclerobionts associated with Orbiramus from the Early Ordovician of Hubei, China, the oldest known trepostome bryozoan. Lethaia 54(4):443–456
Porter SM (2010) Calcite and aragonite seas and the de novo acquisition of carbonate skeletons. Geobiology 8:256–277
Pushkin VI, Popov LE (1999) Early Ordovician bryozoans from north-western Russia. Palaeontology 42:171–189
Ross JP (1966) Early Ordovician entoproct from Oklahoma. Okla Geol Notes 26:218–224
Sepkoski JJ (1984) A kinetic model of Phanerozoic taxonomic diversity. III. Post-Paleozoic Families and Mass Extinctions Paleobiology 10:246–267
Taylor PD (2020) Bryozoan paleobiology. (Wiley Blackwell, Hoboken, Chichester). 1–-320
Taylor PD, Berning B, Wilson MA (2013) Reinterpretation of the Cambrian ‘bryozoan’ Pywackia as an octocoral. J Paleontol 87:984–990
Taylor PD, Curry GB (1985) The earliest known fenestrate bryozoan, with a short review of Lower Ordovician Bryozoa. Palaeontology 28:147–158
Taylor PD, James NP (2013) Secular changes in colony-forms and bryozoan carbonate sediments through geological history. Sedimentology 60(5):1184–1212
Ulrich EO (1890) Part II. Palaeozoic Bryozoa. In Palaeontology of Illinois. Section VI. Rep. of the Geol. Surv.of Illinois 8: 283–688.
Wu RC, Liu JB, Calner M, Gong FY, Lehnert O, Luan XS, Li LX, Zhan RB (2020) High-resolution carbon isotope stratigraphy of the Lower and Middle Ordovician succession of the Yangtze Platform, China: implications for global correlation. J Geol Soc 177:537–549
Xia FS, Zhang SG, Wang ZZ (2007) The oldest bryozoans: new evidence from the late Tremadocian (Early Ordovician) of East Yangtze Gorges. J Paleontol 81:1308–1326
Yaroshinskaya AM (1967) The family Ceramoporidae: main evolutionary trends. Int Geol Rev 9(2):182–185
Yang KC (1957) Some Bryozoa from the upper of the Lower Ordovician of Liangshan, Southern Shensi (including a new genus). Acta Palaeontol Sinica 5(1):1–15
Zeng, QL, Lai CG, Xu GH, Ni SZ, Zhou TM, Xiang LW, Wang XF, Li ZH (1987) Ordovician system, p. 43–-142. In Yichang Institute of Geology and Mineral Resources, Ministry of Geology and Mineral Resources, P. R. China (ed.), Biostratigraphy of the Yangtze Gorge area (2). Early Paleozoic Era. Geological Publishing House, Beijing, China. (In Chinese with English abstract)
Zhang SG, Xia FS, Yan HJ, Wang ZZ (2009) Horizon of the oldest known bryozoans (Ordovician). Palaeoworld 18(1):67–73
Zhang ZL, Zhang ZF, Ma JY, Taylor PD, Strotz LC, Jacquet SM, Skovsted CB, Chen FF, Han J, Brock GA (2021) Fossil evidence unveils an early Cambrian origin for Bryozoa. Nature 599:251–255
Zhuravlev AY, Wood RA (2008) Eve of biomineralization: controls on skeletalmineralogy. Geology 36:923–926
Acknowledgements
This article is a contribution to IGCP 653 ‘The onset of the Great Ordovician Biodiversity Event’. We thank two anonymous reviewers and the associate editor Julien Denayer for their constructive suggestions that improved the text.
Funding
This study was supported by the National Natural Science Foundation of China (NSFC: 41972019, 41472008) and the Strategic Priority Research Program (B) of the Chinese Academy of Sciences (XDB26000000).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Julien Denayer
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Ma, J., Taylor, P.D., Buttler, C.J. et al. Bryozoans from the Early Ordovician Fenhsiang Formation (Tremadocian) of South China and the early diversification of the phylum. Sci Nat 109, 21 (2022). https://doi.org/10.1007/s00114-022-01791-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00114-022-01791-z