
Abstract
The Lower Ordovician (late Tremadocian–early Floian) Fenhsiang and the overlying Hunghuayuan Formations at the Chenjiahe section in the Three Gorges area of Hubei Province, South China, include four types of reef: microbe-dominated (lithistid sponge–stromatolite and lithistid sponge–calcimicrobial) reefs, and skeletal-dominated (lithistid sponge–bryozoan and bryozoan–pelmatozoan) reefs. The microbe-dominated reefs are characterized by the dominance of microbial sediments that encrusted and bound the surfaces of sponges to reinforce the reef frameworks. In contrast, the skeletal-dominated reefs are distinguished by bryozoans that encrusted frame-building sponges and pelmatozoans, and that grew downward to fill the open spaces available within the frameworks. A series of these reefs shows a temporal succession in reef type, with a decline in the lithistid sponge–stromatolite reefs and an increase in the lithistid sponges and receptaculitids within the lithistid sponge–calcimicrobial reefs in the Hunghuayuan Formation; the lithistid sponge–bryozoan reefs are common in both the Fenhsiang and Hunghuayuan Formations. These features of the Chenjiahe reefs are in marked contrast to other coeval reefs on the Yangtze Platform and elsewhere. Skeletal-dominated reefs first developed in the Three Gorges and adjacent areas, located on the central part of the platform. Likewise, lithistid sponges and receptaculitids first developed in the Three Gorges area and then expanded their range. In contrast, stromatolites declined over time, but remained abundant on a marginal part of the platform. The spatial–temporal distributions of these reefs on the Yangtze Platform reflect the initiation of the Great Ordovician Biodiversification Event and its consequences, although influenced by local environmental conditions. The Three Gorges area was a center for the development of skeletal-dominated reefs, which were established earlier here than elsewhere in the world. These reef types and their spatial–temporal successions provide invaluable clues to the earliest evolution of skeletal-dominated reefs and their ensuing development during the Middle–Late Ordovician.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Ordovician saw the Great Ordovician Biodiversification Event, which brought about a marked increase in the biodiversity of marine organisms (Harper 2006). Reefs are defined as calcareous deposits created by essentially in-place sessile organisms (Riding 2002). Reef-building sponges, bryozoans, and corals flourished during this period and were widespread during the time of a transition from microbe- to skeletal-dominated reefs. The skeletal-dominated reef ecosystems became dominant in the latest Middle–Late Ordovician. In contrast, Early Ordovician reefs were dominated by microbial components, together with lithistid sponges and receptaculitids (e.g., Church 1974; Toomey and Nitecki 1979; Pratt and James 1989; Cañas and Carrera 1993).
Early Ordovician reefs in South China (e.g., the Three Gorges, Songzi, Dongzhi and Shitai areas) have been studied previously. For example, stromatolite reefs have been reported from the Shitai area of Anhui Province, and lithistid sponge–stromatolite reefs from the Dongzhi area of Anhui Province (Li et al. 2004b; Adachi et al. 2009; Cao et al. 2009). Several other reef communities have been recognized (e.g., sponge Archaeosphia, receptaculitid Calathium–Archaeosphia, Calathium–cyanobacteria, bryozoan Batostoma, and pelmatozoan–Batostoma communities) based on field observations at the Huanghuachang section of the Three Gorges area, Hubei Province (Zhu et al. 1995, 2006). These reefs have also been discussed from the viewpoints of local environmental conditions and temporal changes in depositional environment and sea level (Li et al. 1997, 2004b; Xiao et al. 2003). The significance of the Early Ordovician reefs has also been examined in terms of transitions in reef construction and the Great Ordovician Biodiversification Event (Adachi et al. 2009, 2011). Adachi et al. (2011) recently showed that the oldest known bryozoan reefs from the Fenhsiang Formation at the Chenjiahe section of the Three Gorges area, Hubei Province represented a very early attempt by skeletal organisms to construct frameworks, and marked a pioneering step in the development of skeletal-dominated reefs, pre-dating the Middle–Late Ordovician reef development.
At the Chenjiahe section in the Three Gorges area, bryozoan reefs occur not only in the late Tremadocian Fenhsiang Formation, but also in the latest Tremadocian–early Floian Hunghuayuan Formation. It is therefore possible to reconstruct secular changes in the distribution of bryozoan reefs throughout the Fenhsiang and overlying Hunghuayuan Formations along with the changes in other types of reef, to clarify the early evolution of skeletal-dominated reefs. Such an analysis would provide insights into the subsequent diversification of skeletal reef organisms and the factors that enabled the Ordovician diversification. Here, we first outline the compositions of the reef-building organisms at the Chenjiahe section, and then document the modes of reef construction and temporal changes in the distribution of each reef type. Finally, we discuss the characteristics of Early Ordovician reef development at the Chenjiahe section, also considering coeval reefs at other sites, especially in terms of the geobiological significance of the early evolution of skeletal-dominated reefs.
Methods
Individual reefs were investigated in detail at various scales in the field, in polished slabs, and under the microscope. In the field, reef facies, reef morphologies, and structures were documented. About 150 samples, including non-reef limestones, were collected and examined. Rock samples were cut perpendicular to bedding, and polished slabs and oriented thin-sections were prepared. Both large (7.5 × 10 cm) and small (5.7 × 5 cm) thin-sections were prepared for analysis by optical microscope. The relative abundances of reef components (lithistid sponges, bryozoans, pelmatozoans, pulchrilaminids, receptaculitids and microbial sediments) were estimated by point-counting (300 points) using large thin-sections.
Geological setting
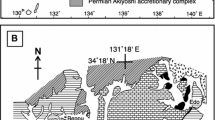
During the Early Ordovician, the South China paleoplate was covered by an extensive epeiric sea on the Yangtze Platform (Fig. 1a). Extensive shallow-marine carbonates prevailed in offshore areas, with terrigenous clastics deposited in inshore areas close to the western oldland (Zhan and Jin 2007). The reef-forming limestones described here were sampled from the Fenhsiang and Hunghuayuan Formations at the Chenjiahe section in the Three Gorges area, located about 3 km south of Fenhsiang town, Yichang City of Hubei Province, South China (Fig. 1b). This area was situated in the central part of the Yangtze Platform during the Ordovician, and it is widely known for its continuous, fossiliferous Ordovician succession. The Fenhsiang Formation at Chenjiahe conformably overlies the early Tremadocian Nantsinkuan Formation, and the Hunghuayuan Formation is overlain by the early Floian Dawan Formation (Fig. 2). The Fenhsiang Formation at the Three Gorges area belongs to the conodont Paltodus deltifer Zone to the lowest part of the Serratognathus diversus Zone, indicating a late Tremadocian age, whereas the Hunghuayuan Formation belongs to the upper part of the Serratognathus diversus Zone to the Prioniodus honghuayuansis Zone, indicating a latest Tremadocian–early Floian age (An 1987; Webby et al. 2004; Liu et al. 2010).

Location of the Chenjiahe section in the Three Gorges area of Hubei Province, South China. a Paleogeographic map during the Early Ordovician (modified from Zhan and Jin 2007). b Index map of the Chenjiahe section

Log of the Chenjiahe section, showing the lithology, reef horizons (numbers 1–42), relative abundances of the main reef components and reef types
Lithofacies descriptions
The Chenjiahe succession, as examined in this study, is subdivided into five lithostratigraphic units (Fig. 2; Table 1). Unit 1 (13 m thick, uppermost Nantsinkuan Formation) is characterized by thin- to medium-bedded oolitic grainstone and subordinate bioclastic packstone/grainstone with cross-bedding. The bioclasts include abundant trilobites and echinoderms, with subordinate brachiopods and cephalopods.
Unit 2 (24 m thick, lower Fenhsiang Formation) comprises medium-bedded, cross-bedded bioclastic packstone/grainstone and yellowish grey shale in its upper part. The lower and middle parts of this unit contain abundant reefs that grade laterally into bioclastic packstone/grainstone (Figs. 3a, 4a). The bioclasts include trilobites, echinoderms, and cephalopods. Brachiopods become abundant in the upper part of the unit. Nuia, a problematic microorganism (Toomey 1967), appears in the upper part of the first unit, and then increases in abundance in Unit 2, forming Nuia grainstone in several horizons.

Lithistid sponge–stromatolite reefs (reef horizons 3 and 3′). a Dome-shaped reef surrounded by bioclastic limestone. The pen for scale is 15 cm long. b Enlargement of the area indicated by a rectangle in a, showing a small, nodular stromatolite developed on sponges. The coin is 2.3 cm across. c Stromatolites and sponges intergrown with each other. The coin is 2.3 cm across. d Inter-stromatolite spaces filled with micrite containing bioclasts. Scale 2 mm. e Photomicrograph showing a small, domal stromatolite produced by accumulations of micritic layers, including Girvanella (white arrow) and micritic layers containing sporadic bioclasts (black arrows). Scale 2 mm. f Enlargement of the area indicated by a rectangle in e. Scale 0.5 mm. Sp lithistid sponge, Re receptaculitid

Lithistid sponge–calcimicrobial reefs. a Outcrop photograph of a small dome-shaped reef surrounded by bioclastic limestone. The hammer is 27 cm long. b Photograph of a cut slab. Scale 2 cm. c Filamentous calcimicrobe Girvanella. Scale 0.5 mm. d Line-drawing of a thin-section. Microbial micrite with laminar texture encrusts and binds the sponges. Scale 1 cm. e Detail of an area in d (the rectangle labeled e). Dome-shaped micrite with laminar texture encrusting sponges. Scale 2 mm. f Pulchrilaminid (pu) encrusts the lower surface of platy sponges. Scale 5 mm. Mi microbial encrustation, Sp lithistid sponge, Re receptaculitid, Bi bioclast, Ma matrix
Unit 3 (18 m thick, upper Fenhsiang Formation) is characterized by medium-bedded peloidal packstone/grainstone and subordinate bioclastic packstone/wackestone, as well as yellow-grey shale. The bioclasts within the packstone/wackestone include dominant brachiopods and echinoderms, as well as subordinate trilobites and cephalopods. The shale includes dominant brachiopods and ramose bryozoans, as well as graptolites (Acanthograptus). Rare small reefs are found in the middle part of this unit.
Unit 4 (19 m thick, Hunghuayuan Formation) is marked by medium-bedded bioclastic packstone/grainstone with well-developed reefs (Figs. 5a, b, 6a, b, 7a), bioturbated lime mudstone, and flat-pebble conglomerate. This unit includes abundant bioclasts, such as brachiopods, echinoderms, trilobites, and cephalopods and their micritized grains. The bioclastic packstone/grainstone commonly contains rounded and sub-rounded micritic intraclasts that were derived from lithistid sponge-bearing reefs.

Lithistid sponge–calcimicrobial reefs (a–b reef horizon 39). a Small reefs accumulated one after another and were finally covered by bioclastic limestone. Filling sediments indicate interruptions to reef growth. The hammer is 27 cm long. b Detail of a (indicated by the rectangle in a). Receptaculitids are recognizable on the outcrop surface. The coin is 2.3 cm across. c Photograph of a cut slab, in which sponges and receptaculitids predominate. Scale 3 cm. d Photomicrograph showing dome-shaped micrite with a laminar texture, encrusting the surfaces of sponges. e Transverse section of sponge with a rounded outline. Scale 5 cm. f Receptaculitids encrusting sponges. Scale 5 mm. Mi microbial encrustation, Sp lithistid sponge, Re receptaculitid

Lithistid sponge–bryozoan (b) and lithistid sponge–calcimicrobial (c–f) reefs. a The lithistid sponge–bryozoan reef developed on bioclastic limestone, followed by the lithistid sponge–calcimicrobial reef, with abundant platy sponges (reef horizons 9–11). The hammer is 27 cm long. b Lithistid sponge–bryozoan reef containing abundant downward-growing bryozoans (arrows). The coin is 2.3 cm across. c Abundant platy sponges on a outcrop surface. The coin is 2.3 cm across. d Photomicrograph of platy sponges. Scale 2 mm. e Line-drawing of a thin-section. Dome-shaped micrite encrusting the surfaces of platy sponges, whereas receptaculitids encrust the lower surfaces of the sponges. Scale 2 mm. f Detail of e (indicated by the rectangle in e) showing microbial encrustation with laminar texture. Scale 2 mm. Mi microbial encrustation, Sp lithistid sponge, Re receptaculitid, Br bryozoan, Pu pulchrilaminid, Bi bioclast

Lithistid sponge–bryozoan reefs. a Reefs (reef horizon 16) with abundant bryozoans, eroded and overlain by bioclastic limestone. The hammer is 27 cm long. b Photograph of a cut slab (reef horizon 9) in which bryozoans encrusting sponges are apparent. Scale 2 cm. c Line-drawing of a thin-section (reef horizon 3), containing abundant bryozoans. Scale 1 cm. d Photomicrograph of b, showing bryozoans on the lower surfaces of sponges, growing downward and laterally. Scale 3 mm. e Bryozoans with ragged margins, representing stacked domal sub-colonies. Scale 5 mm. f Pulchrilaminids encrusting sponges and bryozoans. Scale 5 mm. Sp lithistid sponge, Br bryozoan, Pu pulchrilaminids. Arrows indicate the growth direction of reef-builders
The final unit (>2 m thick, lowermost Dawan Formation) is made up of thin-bedded bioclastic grainstone with abundant glauconite grains. The middle and upper parts of the formation include thin-bedded nodular argillaceous limestone, mudstone and shale (Zhan and Jin 2007).
The depositional environment of the above units is briefly summarized in Table 1.
Reef-building organisms
Lithistid sponges
Most of the reefs studied include lithistid sponges in various degrees of preservation; however, only a few taxonomic studies have examined lithistid sponges from South China. For example, Liu et al. (1997) studied Early Ordovician lithistid sponges from Hubei Province, which predominantly belong to the family Anthaspidellidae and include four genera: Anthaspidella, Archaeoscyphyia, Rhopalocoelia and Jianghania. The lithistid sponges from the Chenjiahe section show a variety of growth forms, including small cylindrical (less than 1 cm in diameter), obconical, large massive (at least 7 cm in diameter) and platy (3–9 mm in thickness) (Figs. 4, 5, 6). The modes of preservation of these lithistid sponges vary from well-delineated sponge bodies with regular lattice-like spicular networks to dark micritic peloidal bodies with partly preserved, aligned sponge bodies (Adachi et al. 2012; Fig. 6). These various degrees of preservation are inferred to reflect the effects of bacterial decay of the sponge soft tissue and their concurrent calcification (e.g., Adachi et al. 2009, 2012).
Receptaculitids
The receptaculitid Calathium is a common component of Early Ordovician reefs from North America, the Argentine Precordillera, and South China (Webby 2002). Most workers regard it as a receptaculitid alga (Nitecki et al. 2004), although some consider it a sponge (Wood 1999; Liu et al. 2005). Receptaculitids occur throughout the Fenhsiang and Hunghuayuan Formations; however, they increase in abundance towards the upper parts of the Hunghuayuan Formation. Receptaculitids are easily recognized at outcrop; most grew in situ, but some have toppled over from their growth position. They exhibit double-walled conical forms (at least 3 cm in height) in longitudinal section and circular forms (0.5–8.0 cm in diameter) in transverse section (Fig. 5). The walls have pores and spine-like structures. Root-like structures or outgrowths for attachment are well developed in some specimens, and these receptaculitids attach to lithistid sponges (Fig. 5f).
Bryozoans
The oldest known bryozoans have previously been reported from the Fenhsiang Formation in the Three Gorges and Songzi areas of Hubei Province, South China (Xiao et al. 1993; Zhu et al. 2006; Adachi et al. 2011, 2012). For a systematic description of these bryozoans, see Xia et al. (2007). An encrusting bryozoan (Nekhorosheviella) from the Chenjiahe section is dominant at several reef horizons in the Fenhsiang and Hunghuayuan Formations. The bryozoans are composed mainly of hemispherical or sheet-like sub-colonies (0.25–0.50 mm in height; Figs. 6b, 7, 8c) that are stacked on top of one another, forming cylindrical or massive growth forms (up to 5 cm in height) with ragged margins. The bryozoans do not directly encrust the bioclastic sediments, but attach to the lithistid sponges and pelmatozoan echinoderms, and preferentially grow downward and laterally. The morphological characteristics and paleoecology of these reef-building bryozoans have been studied in detail by Adachi et al. (2012). The ramose bryozoan Orbiramus is rarely present as bioclasts.

Bryozoan–pelmatozoan reefs (reef horizon 3). a Outcrop surface of a reef in which pelmatozoans are apparent. The coin is 2.3 cm across. b Line-drawing of a thin-section, showing pelmatozoans encrusting bioclastic sediments (Bi). Scale 5 mm. c Pelmatozoans and downward-growing bryozoans encrusting each other. Scale 5 mm. d Dense pelmatozoan encrustation. Scale 3 mm. Br bryozoan, Pe pelmatozoan. Arrows indicate the growth direction of reef-builders
Pulchrilaminids
Pulchrilaminids (Pulchrilamina spinosa) are less abundant than other organisms in reef limestones, making up <5 % of an average area. An Early Ordovician (early Floian) Pulchrilamina was previously reported from North America as a large domical to irregularly shaped skeletal frame-builder (Toomey and Ham 1967), and has been taxonomically assigned to a microbial structure or to the Coelenterata (Cloud and Barnes 1948; Toomey and Ham 1967). However, Pulchrilamina is now tentatively assigned to the stromatoporoids based on their structure (Stearn et al. 1999). The Pulchrilamina described in this study has a thin, wavy, laminar form (0.05–0.40 mm in thickness), or a low domal form (3 mm thick) with ragged margins (Fig. 7e). Obscure laminae are found within the domes. The upper surfaces of the laminae bear randomly arranged spines (0.03–0.06 mm in diameter, 0.2–0.8 mm in height). Pulchrilamina does not directly encrust the bioclastic substrates, but encrusts the surfaces of reef-building lithistid sponges, bryozoans, and pelmatozoans. In some cases, the pulchrilaminids, together with bryozoans, encrust the lower surfaces of the platy lithistid sponges (Fig. 4f), and grew downward into empty spaces within frameworks.
Pelmatozoans
Pelmatozoan fragments are commonly found throughout the reefs and the inter-reefal bioclastic sediments of the Fenhsiang and Hunghuayuan Formations. Small discoidal or crescentic pelmatozoan holdfasts (2–10 mm in diameter) are dominant, especially in the bryozoan–pelmatozoan reefs (for details, see below; Fig. 8). These holdfasts are unique to crinoids and blastozoans.
Calcimicrobes
Girvanella is the only recognizable calcimicrobe in the Chenjiahe section (Figs. 3f, 4c), where it occurs as a non-branching filamentous calcimicrobe with a thin micritic wall and an external diameter of 10–15 μm. It generally consists of a randomly tangled arrangement of filaments. Aggregations of Girvanella, combined aggregations, and micrite encrust and/or bind the skeletal reef-building organisms and micritic substrates. Girvanella is also involved in the formation of stromatolites, as described below (Fig. 3e, f).
Reef types and their characteristic modes of construction
Lithistid sponge–microbial reefs
Adachi et al. (2011) studied the Fenhsiang reefs at the Chenjiahe section in the Three Gorges area, and divided them into three types (lithistid sponge–microbial, lithistid sponge–bryozoan and bryozoan–pelmatozoan) based on the main reef-building organisms and their roles in reef construction. Although typical stromatolites are not found in the Hunghuayuan Formation at the Chenjiahe section, the lithistid sponge–microbial reefs are further subdivided into lithistid sponge–stromatolite reefs and lithistid sponge–calcimicrobial reefs in the present study (Table 2).
Lithistid sponge–stromatolite reefs
The lithistid sponge–stromatolite reefs occur only in the lower part of the Fenhsiang Formation (reef horizons 3 and 3′; Fig. 2). Each reef occurs as a small dome-shaped structure (several tens of centimeters to several meters in height and width) surrounded by bioclastic limestone (Fig. 3a). These reefs are characterized by stromatolites that are microbialites with mesoscopic laminated textures. The stromatolites (3–6 cm in width and 5–7 cm in height) have a small nodular or columnar outer shape, with an obscure laminar texture on outcrop surfaces (Fig. 3). Examination under an optical microscope reveals that the laminae of the stromatolites are defined by irregularly repeated accumulations of micritic layers, including abundant Girvanella and micritic layers with sporadic bioclasts (Fig. 3e, f). The stromatolites formed directly on lithistid sponges or bioclastic sediments. The lithistid sponges occur sporadically intergrown within the stromatolite frameworks. Inter-stromatolite and inter-lithistid sponge spaces are filled with micrite containing bioclasts (Fig. 3d). The bioclasts include sponge spicules, pelmatozoans, and trilobites. These modes of occurrence of stromatolites are similar to those of the Lower Ordovician Hunghuayuan reefs in Anhui Province, South China (Adachi et al. 2009), which are interpreted to have formed by micrite precipitation and/or the trapping and binding of lime mud.
Construction of the reefs: Lithistid sponge–stromatolite reefs were produced by an association of stromatolites and lithistid sponges. The biofilms, including filamentous microbes, first encrusted the bioclastic substrates or lithistid sponges and then stabilized these substrates, growing upward and laterally to eventually form microbial frameworks (stromatolites) that resulted from repeated changes in microbial activity (i.e., micrite precipitation alternating with the trapping and/or binding of micrite). Stabilized substrates also contributed to the formation of hardgrounds for further attachment and colonization by lithistid sponges and pelmatozoans.
Lithistid sponge–calcimicrobial reefs
The lithistid sponge–calcimicrobial reefs are the most common reef type in both the Fenhsiang and Hunghuayuan Formations (Fig. 2), occurring as dome-shaped structures that range in height and width from several tens of centimeters to several meters. Laterally, the reefs grade into surrounding bioclastic limestones (Fig. 4a). These reefs were constructed by microbial encrustations (average area, 13 %), lithistid sponges with cylindrical or obconical forms (16 %), receptaculitids (7 %), and rare pulchrilaminids, bryozoans and pelmatozoans (Fig. 4). The lithistid sponges and receptaculitids show an upward increase in abundance within the Hunghuayuan Formation. In particular, large massive lithistid sponges and receptaculitids are abundant (total average area <50 %) in the upper part of the Hunghuayuan Formation (reef horizons 38 and 39; Fig. 5), along with sporadic bryozoans, pelmatozoan holdfasts and pulchrilaminids. These lithistid sponges and receptaculitids mostly grew in situ; however, they do not usually encrust upon each other. Despite the abundance of these reef-builders, they did not act alone in constructing a rigid, three-dimensional skeletal framework. Small domal and columnar microbial encrustations, made up of peloids and dense micrites including Girvanella, developed on the surfaces of lithistid sponges, bryozoans, pelmatozoans and pulchrilaminids (Figs. 4, 5d). Inter-skeletal reef-builders are also filled with micrites containing bioclasts such as trilobites, brachiopods, pelmatozoans, sponge spicules, cephalopods and gastropods, which are covered by Girvanella-bearing micrites. Trilobites are the dominant bioclasts in the Fenhsiang Formation, whereas brachiopods and gastropods are increasingly abundant in the Hunghuayuan Formation.
Lithistid sponges with platy growth forms flourished below the boundary between the Fenhsiang and Hunghuayuan Formations, and continued to contribute to reef development in the Hunghuayuan Formation (reef horizons 8–14, 25–41). The upper and lower surfaces of the lithistid sponges provided attachment sites for encrusting organisms. Their upper surfaces are typically encrusted by domal and columnar micrites (Fig. 6e, f) that are characterized by irregular, wavy, laminar textures and that include rare bioclasts. In contrast, the lower surfaces are sporadically encrusted by pulchrilaminids and bryozoans showing evidence of downward growth (Fig. 4f).
Construction of the reefs: The lithistid sponge–calcimicrobial reefs were produced by lithistid sponges and in part by receptaculitids, which initially colonized the substrate. The lithistid sponges, especially the cylindrical and obconical forms, and receptaculitids may have acted as bafflers that trapped sediment within the frameworks of the reefs. More importantly, Girvanella-bearing micrites encrusted the bioclastic micrites among the lithistid sponges, thereby stabilizing the underlying sediments and binding the skeletal reef-builders to enhance the rigidity of the reefs. Bacterial decay of the sponge soft tissues was the main control on sponge preservation and on micrite formation within the sponges, as reported previously for Lower Ordovician reefs of Anhui Province (Adachi et al. 2009).
The abundance of lithistid sponges and receptaculitids varies in the lithistid sponge–calcimicrobial reefs, as does the growth form of the lithistid sponges (e.g., cylindrical or platy). The development of lithistid sponges with different growth forms within this type of reef may have been related to subtle changes in the habitat, such as changes in water currents or sedimentation, rather than to differences in the mode of reef construction. Waisfeld et al. (1993) considered that Early Ordovician discoid, domal, or laminar sponges with central cavities were unable to live in environments with moderate to high rates of sedimentation. The occurrence of abundant platy lithistid sponges in the present study section may suggest a low rate of sedimentation.
Lithistid sponge–bryozoan reefs
Lithistid sponge–bryozoan reefs are recognized in both the Fenhsiang and Hunghuayuan Formations (reef horizons 1, 9, 16, 21, 30 and 33; Fig. 2). The initial reef development in this section is marked by the first appearance of lithistid sponge–bryozoan reefs, and the bryozoans are mostly developed in the lower part of the Fenhsiang Formation (horizon 2). The reefs occur as small dome-shaped structures, 1.0–3.0 m in width and 0.5–2.0 m in height. The reefs were developed directly on and are surrounded by bioclastic limestones that contain pelmatozoans, trilobites, and brachiopods. Locally, the reefs are eroded and overlain by bioclastic limestones (Fig. 7a). The reefs show a succession from lithistid sponge bafflestones to lithistid sponge–bryozoan framestones. The lithistid sponges with cylindrical or massive growth forms are dominant in the lower bafflestones. The upper framestones are dominated by the bryozoan Nekhorosheviella (average area, 24 %), subordinate lithistid sponges (average area, 18 %) and microbial encrustations (average area, 6 %), with rare receptaculitids, pelmatozoans and pulchrilaminids. The bryozoans encrust the surfaces of lithistid sponges as scaffolds and are encrusted in turn by other bryozoans, producing irregular frameworks. The most remarkable characteristic of these reefs is the predominance of bryozoans, which grew downward and laterally (Fig. 7). Inter-frameworks are filled with micrite that contains bioclasts such as brachiopods, trilobites, cephalopods, pelmatozoans and gastropods. In the reefs of the Hunghuayuan Formation, in which the platy lithistid sponges are dominant, the bryozoans uniquely encrust the lower surfaces of the lithistid sponges (Fig. 6b).
Construction of the reefs: The lithistid sponge–bryozoan reefs were produced mainly by lithistid sponges, which were the first to colonize the bioclastic substrates. These sponges may have acted as bafflers that trapped sediment, and they provided attachment sites for encrusting bryozoans that grew downwards and laterally. The bryozoans made full use of the vacant spaces available for growth, producing rigid skeletal frameworks. However, the lithistid sponges that acted as scaffolds were not necessarily preserved in the final products, because the degree of preservation of the sponges is highly variable (Adachi et al. 2012).
Bryozoan–pelmatozoan reefs
The bryozoan–pelmatozoan reefs are only observed in one horizon (reef horizon 3; Fig. 2) as small dome-shaped structures of at least 20–30 cm in width and 5–10 cm in height (Fig. 8a). Pelmatozoans (mainly holdfasts) and bryozoans (Nekhorosheviella) are the main components of the reef, making up 10–60 % (average, 38 %) and 25–40 % (average, 32 %), respectively. Pelmatozoans with discoidal or crescentic shapes encrust each other (Fig. 8c, d). Cylindrical boreholes (0.3–0.7 mm in diameter) belonging to the ichnogenus Trypanites are common in the pelmatozoan skeletons. In contrast, the bryozoans occur as stacked hemispherical sub-colonies that form domal growths with ragged margins (3–15 mm high). The pelmatozoans grew directly on the underlying bioclastic substrates (pelmatozoans and trilobites, with rare brachiopods and bryozoans) (Fig. 8b), whereas the bryozoans encrust individual pelmatozoans. This mode of occurrence of pelmatozoans and bryozoans, and the mode of construction, are similar to those reported for Middle Ordovician pelmatozoan-dominated patch reefs (Pratt 1989).
Construction of the reefs: The bryozoan–pelmatozoan reefs were constructed by pelmatozoans that were the first to settle on the bioclastic substrate, acting as bafflers that trapped sediment. The bryozoans then encrusted the surfaces of the pelmatozoans, using them as scaffolds, and extended downward and laterally to occupy the inter-spaces between the densely aggregated pelmatozoans. These reefs, with rigid three-dimensional frameworks built mainly by skeletal organisms, are considered one of the first skeletal-dominated reefs.
Geobiological significance of Early Ordovician reefs in the Three Gorges area
The Fenhsiang and Hunghuayuan Formations at the Chenjiahe section contain various microbe- and skeletal-dominated reefs with the following stratigraphic distributions: (1) lithistid sponge–calcimicrobial reefs are common where the lithistid sponges and receptaculitids become increasingly abundant in the Hunghuayuan Formation; (2) lithistid sponge–stromatolite reefs have a limited occurrence, restricted to the Fenhsiang Formation; and (3) lithistid sponge–bryozoan reefs are abundant in the Fenhsiang and Hunghuayuan Formations. Bryozoans show no upward increase in abundance throughout the Chenjiahe section, but were prolific when they first appeared, occupying niches as the main skeletal reef-builders at that time.
In comparing the above characteristics with those of three other reef associations that developed in contrasting environmental settings, at (1) the Shitai section in the Shitai area of Anhui Province, South China; (2) the Jianxin section in the Dongzhi area of Anhui Province; and (3) the Hunghuayuan section in the Tongzi area of Guizhou Province on the Yangtze Platform, the following unique modes of reef development can be identified at the Chenjiahe section. (1) In the late Tremadocian, the stromatolite-bearing (stromatolite or lithistid sponge–stromatolite) reefs were still common in all areas (Fig. 9). In contrast, after the latest Tremadocian, the stromatolite-bearing reefs declined markedly and only remained in the Dongzhi area, located on a marginal part of the platform (e.g., Li et al. 2004b; Adachi et al. 2009; Cao et al. 2009) (Fig. 9). (2) Whereas the calcimicrobe Girvanella is common everywhere, Renalcis and Epiphyton are only found in the Dongzhi area (Adachi et al. 2009). (3) During the late Tremadocian to early Floian, the skeletal-dominated reefs, such as the lithistid sponge–bryozoan reefs, were limited to the Three Gorges area and its surroundings (e.g., Zhu et al. 2006; Adachi et al. 2011); however, the skeletal reef-builders, such as the lithistid sponges, receptaculitids and bryozoans, expanded their habitats from the Three Gorges area, located in a central part of the platform, to the Dongzhi and Tongzi areas, forming the lithistid sponge–calcimicrobial reefs (e.g., Adachi et al. 2009; Cao et al. 2009) (Fig. 9). Therefore, the Three Gorges area was a center for the development of skeletal-dominated reefs, whose reef-builders then spread to other areas.

Index map showing reef-bearing localities on the Yangtze Platform during the Early Ordovician (late Tremadocian–early Floian). Approximate stratigraphic ranges of individual reefs are also shown (below). The Chenjiahe section in the Three Gorges area was a center for the development of skeletal-dominated reefs
The Early Ordovician reefs of the Yangtze Platform, including the Chenjiahe section, show a transition from microbe-dominated reefs (from stromatolite, to lithistid sponge–stromatolite, to lithistid sponge–calcimicrobial reefs) to skeletal-dominated (lithistid sponge–bryozoan and bryozoan–pelmatozoan) reefs. This transition in reef construction reflects the initiation of the Ordovician diversification in South China. The mode of reef development in each area may have been closely related to the local environment upon the platform (e.g., water depth, water energy, water temperature, nutrient content and siliciclastic input), which would have affected the growth and diversity of marine organisms. The Three Gorges area was situated in a somewhat shallower-water setting than the marginal platform (e.g., the Dongzhi and Shitai areas). The rate of subsidence in the marginal belt was much higher than that in the central part of the platform, especially during the deposition of the Fenhsiang and coeval reefs (Li et al. 2004b). These shallow-water environments were favorable for skeletal reef-builders.
Li et al. (2004a) examined the spatial and temporal distributions of Early–Middle Ordovician acritarchs on the Yangtze Platform, reporting that diversity increased over time in the offshore shelf region in the Three Gorges area, in contrast to low diversity in nearshore environments. The high primary productivity in the Three Gorges area would have brought rich food supplies and provided favorable habitats for filter and suspension feeders. Thus, the Three Gorges area became a suitable environment for the development of skeletal reef-builders and for the establishment of skeletal-dominated reefs.
In contrast, the marked decline in stromatolite-bearing reefs on the Yangtze Platform during the late Tremadocian to early Floian is attributable to the increased activities of skeletal benthic organisms. In fact, their decline at the Chenjiahe section was approximately synchronous with an increase in intensive bioturbation and the replacement of Cambrian-type shell-beds that are dominated by trilobites, by Paleozoic-type shell-beds that are dominated by brachiopods (Liu et al. 2010). However, stromatolite-bearing reefs were still abundant in the marginal platform areas, where the diversification of skeletal organisms may have been delayed compared with the central part of the platform.
The skeletal-dominated (lithistid sponge–bryozoan) reefs were first established in the late Tremadocian Fenhsiang Formation, succeeded by early Floian Hunghuayuan reefs in the Three Gorges area. Elsewhere in the world, no skeletal-dominated reefs had developed by the Middle–Late Ordovician (e.g., Webby 2002), with the exception of stromatoporoid reefs in west Texas and Oklahoma (early Floian; Toomey and Ham 1967) and in Argentina (Dapingian; Keller and Flügel 1996). Skeletal-dominated reefs finally reached their peak in the Middle–Late Ordovician. Microbe-dominated reefs, both lithistid sponge–stromatolite and lithistid sponge–calcimicrobial types, were still dominant globally during the Early Ordovician. It is significant that a novel reef ecosystem was initiated in South China during the Early Ordovician (late Tremadocian) with the introduction of skeletal-dominated reefs, earlier than elsewhere (Adachi et al. 2011). What allowed the skeletal-dominated reefs to develop earlier in South China? In this work, basic information has been provided on the Early Ordovician reef successions at the Chenjiahe section in the Three Gorges area, where the features of the oldest skeletal-dominated reefs (including their origin, temporal distribution and modes of reef construction) are well recorded. The results presented here indicate that further research is required into the interactions between the diversification of marine organisms and paleoenvironments, in order to explain the different timings of the appearance and development of skeletal-dominated reefs among paleoplates, the establishment and subsequent development of skeletal-dominated reefs, and the factors that triggered the Great Ordovician Biodiversification Event.
Conclusions
-
1.
Early Ordovician reefs of the Fenhsiang and Hunghuayuan Formations at the Chenjiahe section in the Three Gorges area occur as four types: (1) lithistid sponge–stromatolite, (2) lithistid sponge–calcimicrobial, (3) lithistid sponge–bryozoan, and (4) bryozoan–pelmatozoan, based on the main reef-builders and their roles in reef construction. The former two reefs are characterized by microbial sediments that encrust the inter-sponge sediments to stabilize the underlying bioclastic sediments and/or encrust the surfaces of the sponges to reinforce the frameworks. In contrast, the latter two reefs are distinguished by bryozoans that encrust the sponges and pelmatozoans, and that fill the inter-frameworks by growing downward and laterally, culminating in the construction of a rigid skeletal framework by encrusting and/or binding with each other.
-
2.
The stratigraphic distribution of the Chenjiahe reefs shows the following characteristics. The lithistid sponge–calcimicrobial reefs are the most common, and the lithistid sponges and receptaculitids show an increase in abundance over time, especially in the Hunghuayuan Formation. In contrast, the lithistid sponge–stromatolite reefs only occur in the lower parts of the Fenhsiang Formation. Whereas the lithistid sponge–bryozoan reefs (skeletal-dominated reefs) occur in both the Fenhsiang and Hunghuayuan Formations, bryozoans are especially abundant in the lower parts of the Fenhsiang Formation.
-
3.
During the Early Ordovician (late Tremadocian–early Floian), the Three Gorges area, located on the central part of the Yangtze Platform was a center for the development of skeletal-dominated reefs. The skeletal reef-builders, such as lithistid sponges, receptaculitids and bryozoans, expanded their habitats from the central part of the platform to surrounding areas. The stromatolite-bearing reefs declined earlier in the Three Gorges area than in other areas, despite their continued dominance on a marginal part of the platform. These patterns of reef development on the Yangtze Platform during the initial stage of the Ordovician diversification help us to understand the subsequent rise and fall of the skeletal reef-builders.
-
4.
This study on the nature of the earliest skeletal-dominated reefs provides invaluable clues as to why skeletal-dominated reefs first appeared in South China, and how they increased their range diachronously towards other paleoplates via a worldwide transition in reef construction.
References
Adachi N, Ezaki Y, Liu J, Cao J (2009) Early Ordovician reef construction in Anhui Province, South China: a geobiological transition from microbial- to metazoan-dominant reefs. Sed Geol 220:1–11
Adachi N, Ezaki Y, Liu J (2011) Early Ordovician shift in reef construction from microbial to metazoan reefs. Palaios 26:106–114
Adachi N, Ezaki Y, Liu J (2012) The oldest bryozoan reefs: an unusual mode of growth for Early Ordovician reefs. Lethaia 14:14–23
An T (1987) Early Paleozoic conodonts from South China. Peking University Press, Beijing, p 238 (in Chinese with English summary)
Cañas F, Carrera M (1993) Early Ordovician microbial-sponge-receptaculitid bioherms of the Precordillera, Western Argentina. Facies 29:169–178
Cao J, Liu J, Ezaki Y, Adachi N (2009) Lower Ordovician reefs in the Honghuayuan Formation at Dongzhi, Anhui: microbial reefs just prior to the Ordovician biodiversification. Acta Scientiarum Naturalium Universitatis Pekinensis 45:279–288 (in Chinese with English abstract)
Church SB (1974) Lower Ordovician patch reefs in western Utah. Brigham Young University. Geol Stud 21:41–62
Cloud PE Jr, Barnes VE (1948) The Ellenburger group of central Texas, vol 4621. University of Texas Publication, Austin, p 473
Harper DAT (2006) The Ordovician biodiversification: setting an agenda for marine life. Palaeogeogr Palaeoclimatol Palaeoecol 232:148–166
Keller M, Flügel E (1996) Early Ordovician reefs from Argentina: stromatoporoid vs stromatolite origin. Facies 34:177–192
Li B, Zhu Z, Xiao C, Hu M (1997) The biocommunity evolution and depositional environment transition of the Lower Ordovician Fenxiang Formation in the Western Hubei area. Acta Sedimentol Sin 15:97–102 (in Chinese with English summary)
Li J, Servais T, Yan K, Zhu H (2004a) A nearshore-offshore trend in the acritarch distribution of the Early-Middle Ordovician of the Yangtze platform, South China. Rev Palaeobot Palynol 130:141–161
Li Y, Kershaw S, Mu X (2004b) Ordovician reef systems and settings in South China before the late Ordovician mass extinction. Palaeogeogr Palaeoclimatol Palaeoecol 205:235–254
Liu B, Rigby JK, Yanwen J, Zhu Z (1997) Lower Ordovician lithistid sponges from the Eastern Yangtze Gorge Area, Hubei, China. J Paleontol 71:194–207
Liu B, Shu Z, Li X (2005) A discussion several problems of calathid fossils. Acta Palaeontol Sin 44:267–282 (in Chinese with English summary)
Liu J, Ezaki Y, Adachi N, Zhan R (2010) Evidence for decoupling of relative abundance and biodiversity of marine organisms in the initial stage of the GOBE: a preliminary study on shellbeds of the Lower Ordovician in South China. J Earth Sci 21:44–48
Nitecki MH, Webby BD, Spjeldnaes N, Zhen Y (2004) Receptaculitids and algae. In: Webby BD, Paris F, Droser ML, Percival IG (eds) The Great Ordovician Biodiversification Event. Columbia University Press, New York, pp 336–347
Pratt BR (1989) Small Middle Ordovician patch reefs, Laval Formation (Chazy Group) Caughnawaga Montreal Area, Quebec. In: Geldsetzer HHJ, James NP, Tebbutt GE (eds) Reefs, Canada and adjacent areas, Memoir, vol 13. Canadian Society of Petroleum Geologists, Calgary, pp 218–223
Pratt BR, James NP (1989) Early Ordovician thrombolites, St. George Group, western Newfoundland. In: Geldsetzer HHJ, James NP, Tebbutt GE (eds) Reefs, Canada and adjacent areas, Memoir, vol 13. Canadian Society of Petroleum Geologists, Calgary, pp 231–240
Riding R (2002) Structure and composition of organic reefs and carbonate mud mounds: concepts and categories. Earth Sci Rev 58:163–231
Stearn CW, Webby BD, Nestor H, Stock CW (1999) Revised classification and terminology of Palaeozoic stromatoporoids. Acta Palaeontol Pol 44:1–70
Toomey DF (1967) A problematical micro-organisms from the El Paso Group (Lower Ordovician) of West Texas. J Paleontol 40:1304–1311
Toomey DF, Ham EW (1967) Pulchrilamina, a new mound-building organism from Lower Ordovician rock of west Texas and southern Oklahoma. J Paleontol 41:981–987
Toomey DF, Nitecki MH (1979) Organic buildups in the Lower Ordovician (Canadian) of Texas and Oklahoma. Fieldiana Geol 2:1–181
Waisfeld BG, Sánchez TM, Benedetto JL, Carrera MG (1993) Early Ordovician (Arenig) faunal assemblages from western Argentina: biodiversification trends in different geodynamic and palaeogeographic settings. Palaeogeogr Palaeoclimatol Palaeoecol 196:343–373
Webby BD (2002) Patterns of Ordovician reef development. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic reef patterns. SEPM Special Publication 72, Tulsa, pp 129–179
Webby BD, Cooper RA, Bergström SM, Paris F (2004) Stratigraphic framework and time slices. In: Webby BD, Paris F, Droser ML, Percival IG (eds) The Great Ordovician Biodiversification Event. Columbia University Press, New York, pp 41–47
Wood R (1999) Reef evolution. Oxford University Press, Oxford, p 414
Xia F, Zhang S, Wang Z (2007) The oldest bryozoans: new evidence from the late Tremadocian (Early Ordovician) of East Yangtze Gorges. J Paleontol 81:1308–1326
Xiao C, Jiang Y, Liu B, Zhu Z (1993) A discovery of Batostoma genus of Early-Middle age of Early Ordovician and a study on its geologic function and ecology in Middle Yangtze area. Chin Sci Bull 38:1314–1315 (in Chinese)
Xiao C, Zhu Z, Li X (2003) Sequence stratigraphy of Lower Ordovician reef-bearing strata in the Middle Yangtze platform. Geol China 30:274–280
Zhan R, Jin J (2007) Ordovician–Early Silurian (Llandovery) stratigraphy and palaeontology of the Upper Yangtze Platform. Science Press, Beijing, p 169
Zhu Z, Jiang Y, Liu B (1995) Paleoecology of late Tremadocian reef bearing strata in western Hubei province of China. In: Cooper JD, Droser ML, Finney SC (eds) Ordovician odyssey. Short papers for the 7th international symposium Ordovician system, Las Vegas, Pacific section SEPM, pp 427–728
Zhu Z, Hu M, Liu B, Xiao C, Yang W, Li X (2006) The Early-Middle Ordovician bioherms of China. Geological Publishing House, Beijing, p 178 (in Chinese)
Acknowledgments
We thank Maurice Tucker, Brian Pratt, and Martin Keller for critical comments that improved this manuscript. This research was supported by a Grant-in-Aid from the Japan Society for the Promotion of Science Fellows, the China Postdoctoral Science Foundation, and the Fujiwara Natural History Foundation, as well as funded by grants from the Scientific Research Fund of the Japan Society for the Promotion of Science (21340154, 22654062), the National Natural Science Foundation of China (40972020, 40825006), and the State Key Laboratory of Palaeobiology and Stratigraphy (113104).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Adachi, N., Liu, J. & Ezaki, Y. Early Ordovician reefs in South China (Chenjiahe section, Hubei Province): deciphering the early evolution of skeletal-dominated reefs. Facies 59, 451–466 (2013). https://doi.org/10.1007/s10347-012-0308-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10347-012-0308-2