
Abstract
Understanding patterns of beta diversity (heterogeneity) and its components (substitution and subset) is important for managing freshwater ecosystems and the research on these topics has increased over the last decade. However, there has been limited empirical research exploring the drivers of these components of beta diversity in ponds from agricultural landscapes, in which hydroperiod length is often a key structural driver. The purpose of our work was to examine the contribution of substitution (species replacement among sites without changes in richness) and subset (species differences among sites when species-poor sites constitute subsets of those with a greater number of taxa) structuring processes on macrophyte beta diversity patterns of sixteen permanent and ten temporary Mediterranean ponds. We also aimed to test the structuring role of local environmental factors and geographic isolation and to determine whether there are differences in the community heterogeneity and the relative importance of the substitution and subset fractions of beta diversity between permanent and temporary ponds. We found that temporary ponds were environmentally and biologically less heterogeneous and supported lower richness than permanent waterbodies. In both pond types, beta diversity almost entirely reflected patterns of species substitution rather than subsets. Local environmental conditions, and not among-pond distance, were the main drivers of macrophyte community structure. Overall, our results suggest that local environmental variables operating through niche processes were the primary mechanisms driving macrophyte beta diversity patterns, thus highlighting the importance of environmental heterogeneity for maintaining pond community diversity in agricultural landscapes. Accordingly, conservation initiatives and ecosystem management strategies should include permanent and temporary pond clusters comprising wide environmental gradients in their efforts to ensure high levels of regional biodiversity.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Ponds have been historically neglected and knowledge on their functioning has lagged behind compared to larger aquatic ecosystems such as lakes and rivers (De Meester et al. 2005). However, these ponds are now recognized as important features in landscapes throughout the world. Evidence from Europe and America suggests that, despite their small surface area compared to larger freshwater and terrestrial habitats, these ecosystems constitute biodiversity islands supporting high richness of invertebrate and aquatic plant species, as well as extensive food webs (Davies et al. 2008; Jeffries 2008). This is particularly relevant in the Mediterranean region, where ponds contribute significantly to regional biodiversity thanks to their abundance and their high spatial and temporal variability (Grillas et al. 2004; Florencio et al. 2009; Oertli et al. 2009; Van den Broeck et al. 2015). Unfortunately, more than half of European ponds have disappeared during the past century (CEC 1995). The rapid loss of biodiversity due to agricultural expansion, urban development, fragmentation and partial or total destruction of the habitat brings an urgent need for more scientific research in order to understand patterns of species diversity and the forces controlling community composition and assemblage structure (Scheffer et al. 2006).
Recently, it has been recognized that a better knowledge of patterns of diversity is essential for managing and preserving aquatic ecosystems (Alahuhta et al. 2017). Species diversity consists of three main components across different spatial scales: alpha or local diversity, gamma or regional diversity and beta diversity or community dissimilarity. Beta diversity describes among-site spatial variation in community composition (Anderson et al. 2011), and it has been connected with two different but complementary processes: turnover (species replacement among sites without changes in richness) and nestedness (species differences among sites when species-poor sites constitute subsets of those with a greater number of taxa) (Baselga 2010; Legendre 2014). This beta partitioning is based on an incidence measure of dissimilarity. A similar approach based on abundance data has recently been proposed by Baselga (2013, 2017). In this approach, variation in assemblage composition (beta diversity) can be separated into balanced variation in species abundance, analogous to turnover in incidence-based patterns, and abundance gradients, equivalent to species nestedness.
The partitioning of beta diversity into substitution (whether expressed as species turnover or balanced variation in abundance) and subset patterns (species nestedness or abundance gradients) offers a powerful approach to guide regional conservation strategies (Socolar et al. 2016). If substitution patterns are dominant, conservation efforts should probably be directed to preserve heterogeneous sets of ponds with different community compositions. By contrast, strong subset patterns among sites may require conservation actions that prioritize species-rich sites over species-poor sites (Socolar et al. 2016).
The relative contribution of substitution and subset fractions to beta diversity may be driven by many forces such as local conditions, species-specific dispersal abilities and spatial processes (Tonkin et al. 2015). One of the most important among them might be hydroperiod length, which is considered as the principal constraint of aquatic habitats, limiting the development of species (Della Bella et al. 2008; Zokan and Drake 2015).
The coexistence of temporary and permanent pond provides environmental heterogeneity at a regional scale, which can in turn promote the coexistence in a region of species with differing degrees of tolerance to water stress (e.g. Alahuhta et al. 2017; De Meester et al. 2005; Della Bella et al. 2008; Rolon et al., 2008). It is well known that macrophyte diversity patterns and assemblage structure also respond to many other drivers, including specific local conditions and regional landscape factors (e.g. Oertli et al. 2009; O’Hare et al. 2012). Many of the environmental forces structuring diversity patterns and composition of macrophyte assemblages are regarded as deterministic (e.g. pond size, altitude, land uses and water chemistry). In such cases, a niche-selection filtering (i.e. species sorting) from the regional species pool plays the major role in shaping local community composition (Chase 2003, 2007). Drought in particular is known to generate deterministic processes. It works as an environmental filter which shapes community structure (Chase and Leibold 2003; Chase 2007). In fact, deterministic niche selective forces might be more prominent in temporary ponds.
The overall purpose of this study was to explore differences in macrophyte diversity and community composition patterns between temporary and permanent ponds in an agricultural landscape. A more specific aim was to identify the environmental drivers responsible for these patterns and to determine whether hydroperiod (temporary vs permanent) influences among-pond community heterogeneity and the contribution of substitution and subset patterns to beta diversity. It was hypothesized that local richness would be higher in permanent systems. Likewise, beta diversity was expected to be higher among permanent ponds because of their higher environmental heterogeneity. Furthermore, the relative influence of local environmental factors and among-pond distances on the structure of macrophyte communities was analysed in temporary and permanent ponds. Local factors were expected to be more important than among-pond distances in both temporary and permanent systems due to the small spatial extent of the study and the good dispersal capabilities of macrophytes.
Materials and methods
Study area
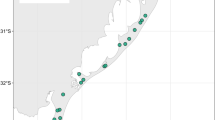
This study was conducted in 26 ponds belonging to a pond complex associated with the Castile Channel in a steppe area of the Northern Iberian Plateau (Spain). Altitudes range between 750 and 800 m.a.s.l. All the ponds lie in an agricultural landscape and are affected by a number of disturbances, in particular sedimentation, livestock activities, and diffuse pollution. They have an anthropogenic origin, arising from the construction of the Castile Channel in the eighteenth century as a result of water filtering from the channel or because the flow of water from small streams and irrigation canals was blocked by the Channel bank (Junta de Castilla and León 2010). Nowadays, all these ponds function like natural systems, although some of them are subjected to excavation or have been partially invaded by croplands. Their mean area is 3.2 ha, but most of them are between 1 and 2.5 ha. Maximum depth is usually between 0.6 and 2 m (mean 1.2). The climate in this area is Continental-Mediterranean, with a wide seasonal variation in temperature since summers are usually hot and dry and winters are primarily cold. Rains are more frequent in autumn and spring. This climate dynamic leads to a seasonal filling regime in most ponds, with strong intra-annual water level fluctuations.
Sixteen of the ponds do not usually dry up although they experience low water levels in summer, the remaining ten ponds dry up every year, typically in summer. These ponds are referred to as permanent and temporary ponds, respectively (Fig. 1). Their hydrological regime is closely related to the Castile Channel. Some of the permanent ponds are connected to the Channel through leakages. Both the permanent and temporary ponds are fed by water inputs from streams, irrigation channels or surface runoff from the catchment (Junta de Castilla and León 2010). Temporary ponds usually remain dry over July, August and September, however, this period may differ among ponds depending on their depth. Furthermore, there are interannual variations, since the ponds depend on spring rainfalls. Table 1 summarizes some environmental variables measured in the study ponds.

Inset map of Spain and location of the 26 study ponds along the Castile Channel
Sampling
Macrophyte communities were sampled once in June or July 2005 using transects. The number of transects in each pond ranged between two and eight, depending on the pond area, the development of the shore (Jensén 1977) and vegetation heterogeneity. The average numbers of transects performed in the permanent and temporary ponds were 4.3 and 4.4, respectively. Quadrats (0.5 m × 0.5 m) were placed at varying intervals of 0–5 m along each transect depending on the vegetation heterogeneity. Percentage coverage of each species was quantified within each unit. The mean coverage of each species in a pond was calculated as the sum of coverages of the species in all sampling units divided by the total number of sampling units used in the pond. Nomenclature followed Flora Iberica (Castroviejo et al. 1986–2013), Cirujano et al. (2008, 2014), and Fernández-Aláez et al. (2012).
Several water samples were randomly taken from the whole water column at different locations in each pond along a shore-centre transect using a core and were combined and mixed to make a composite sample. Conductivity, pH, temperature, dissolved oxygen and turbidity were measured in situ using field probes (Model LF 323, Model 330i and Model OXI 320, respectively, WTW) and a portable turbidimeter (Model 2100P, Hach). The composite samples were analysed in laboratory for total phosphorus (TP), soluble reactive phosphorus (SRP), nitrate (NO3−-N), ammonium (NH4+-N), total suspended solid (TSS), alkalinity, chloride, sulphate, calcium, magnesium, sodium, potassium and chlorophyll “a”. Samples were preserved at 4 °C until they were analysed. All analyses were carried out according to standard methods (APHA 1989). All the physical and chemical variables were measured on the same date as the vegetation sampling.
Pond area was measured on images available in SIGPAC (the Spanish Geographical Information System for Agricultural Parcels, https://www.sigpac.jcyl.es/visor/) and maximum pond depth was determined by measuring depth at each vegetation sampling unit.
Data analysis
To explore possible differences in macrophyte community composition between temporary and permanent ponds a non-metric multidimensional scaling (NMDS) analysis was performed on the Bray–Curtis similarity matrix. A SIMilarity of PERcentages Analysis (SIMPER) was conducted to identify which species were more frequent in temporary and permanent ponds and therefore contributed to the dissimilarities between the two types of ponds (cut off criteria = 90% of cumulative contribution to overall dissimilarity).
Alpha and gamma diversity (measured as species richness) were determined separately for each pond type. Analysis of Variance (ANOVA) was used to test whether pond types (permanent and temporary ponds) differed in their local species richness. The data were checked for normality and homoscedasticity using Shapiro Wilk’s and Levene’s tests, respectively.
The effect of log-transformed environmental variables on local species richness in temporary and permanent ponds was also investigated using stepwise multiple regression with forward selection (cut-off for variable selection p < 0.05). Multicollinearity between pairs of environmental variables was examined using Pearson’s correlation coefficient and the variables highly intercorrelated (Pearson correlation coefficient > 0.7) were removed. Hence, environmental variables used in step-wise multiple regression were pH, conductivity, SRP, TP, ammonium, potassium, chlorophyll a, turbidity, pond area, and maximum depth.
A non-parametric Permutational Multivariate Analysis of Variance (PERMANOVA, Anderson 2001) was used to test whether pond types (permanent and temporary) differed in their variability in species composition. PERMANOVA compares the variability within groups against the variability between groups and calculates a Pseudo-F statistic and its corresponding P value based on permutations (999 replicates) of species compositional data. A significant PERMANOVA Pseudo-F value may indicate differences in dissimilarity between groups (differences in within-group centroids in multivariate space), differences in the within-groups dispersion (mean distances of members to their group centroid), or both. To test whether pond types differed in their within-type dispersion, or beta diversity, a complementary analysis, the test of homogeneity of multivariate dispersion (PERMDISP, Anderson et al. 2006) was performed. This test compares between-group differences in the distance from ponds to their group centroid using analysis of variance. The null hypothesis of no differences in beta diversity between two pond groups was assessed using 999 permutations of residuals (Anderson et al. 2006). Both PERMANOVA and PERMDISP were calculated based on Bray–Curtis dissimilarity. Similarly, PERMANOVA and PERMDISP based on Euclidean distances of standardized environmental variables were used to test if permanent and temporary ponds differed in their environmental heterogeneity.
The abundance-based beta diversity partitioning approach proposed by Baselga (2013, 2017) was used to evaluate if variation in community composition within each pond type was more closely linked to balanced variation in abundance (i. e. substitution) between ponds or to abundance gradients in which one assemblage is a subset of another (i.e. subsets). These fractions are analogous to the replacement and nestedness components of incidence-based dissimilarity (Baselga 2010). To partition beta diversity based on Bray–Curtis matrix for each pond type, two approaches were used. First, the overall multiple-site dissimilarity was assessed, considering the total beta diversity (βBC), as well as its two components, the balanced variation in abundance (βBC.BAL) and abundance gradient (βBC.GRA) (Balselga 2017). In order to reduce the bias caused by the different number of ponds (16 permanent vs 10 temporary) of each group, a resampling procedure was used for permanent ponds, taking 100 random samples of 10 ponds and computing the average beta diversity values (Baselga 2010). Second, to determine beta diversity partitioning between pairs of ponds a pairwise dissimilarity procedure was used, in which the pairwise dissimilarity index (βbc) was fractioned into balanced variation in abundances (βbc.bal) and abundance gradients (βbc.gra) (Baselga 2013). Thus, three pairwise dissimilarity matrices were built: Bray Curtis dissimilarity, balanced variation and abundance gradients.
Next, for each group of ponds, RELATE (comparative Mantel-type tests on similarity matrices) routine with 9999 permutations of sample data was used in order to explore possible relationships between biotic dissimilarity computed as βbc, βbc.bal and βbc.gra and the spatial and environmental distances (Euclidean distance). This routine uses Spearman rank correlation and incorporates a significance test derived by permutation to test the null hypothesis of complete absence of match between the two matrices (Clarke and Warwick 2001). Previously, the BIOENV analysis (Clarke and Ainsworth 1993) was used to obtain “the best” environmental distance matrix from all log-transformed environmental variables, such that the combined variables maximized the correlation between environmental and biological dissimilarity. The matrix of spatial distances contained pairwise geographic distances between all ponds calculated from latitude and longitude.
ANOVA and stepwise multiple regression were conducted with SPSS v.21, while beta diversity partitioning was performed using the R package betapart version 1.4 (Baselga et al. 2017) for R (R Core Team 2013). The remaining statistical analysis (NMDS, SIMPER, PERMANOVA, PERMDISP, BIOENV, RELATE) were carried out with PRIMER7 software, including the PERMANOVA + add-on.
Results
Macrophyte species richness and composition in temporary and permanent ponds
Sixty-two species (gamma diversity) were identified in the study area. Twenty-eight of them were emergent and thirty-four were hydrophytes (twenty-nine rooted submerged, three rooted floating-leaved, one free submerged and one free floating macrophyte species). A total of fifty-nine species were recorded in permanent ponds and thirty-four in temporary ponds.
The species × sites NMDS analysis segregated permanent from temporary ponds (stress 0.21) and corroborated the initial division in permanent and temporary ponds (Fig. 2). SIMPER revealed a taxonomic dissimilarity of 67.45% between the two types of ponds. The taxa contributing most to this dissimilarity were Schoenoplectus lacustris (L.) Palla, Phragmites australis (Cav) Trin ex Steud, Persicaria amphibia L. (Gray), Drepanocladus aduncus (Hedw.) Warnst, Typha domingensis Pers., and Hippuris vulgaris L. (Table 2). The average similarities among permanent and among temporary ponds were 32.49 and 45.25%, respectively. S. lacustris and P. australis were the most frequent and abundant species in permanent ponds and four hydrophyte species were frequent and exclusive of them: Lemna minor L., Myriophyllum spicatum L., Chara hispida var. major (Vaill. ex Hy) R.D. Wood, and Ceratophyllum demersum L. Furthermore, other aquatic species such as Myriophylum verticillatum L., Nitella mucronata (A. Braun) F. Miquel, Nitella translucens (Persoon) C. Agardh, Potamogeton crispus L., Potamogeton lucens L., Ranunculus peltatus Schrank, Zannichellia palustris L. and Zannichellia peltata Bertol. were only found in permanent ponds. The most frequent and abundant species in temporary ponds were S. lacustris and D. aduncus. Furthermore, two species of bryophytes were only found in temporary ponds (Amblystegium serpens Schimp., Brachytecium rutabulum (Hedw.) Schimp.).

Non-Metric Multidimensional Scaling plot of macrophyte community structure in permanent (black diamonds) and temporary (grey circles) ponds
Mean alpha diversity, measured as local species richness, was significantly (p < 0.01) higher in permanent ponds (mean ± sd, 18.9 ± 7.5; range 7 to 40 species) than in temporary ponds (13.1 ± 3.4; range 8 to 20) (Fig. 3).

Comparison of macrophyte species richness (α diversity) of permanent and temporary ponds. Inset are the results from the Analyses of Variance (ANOVA)
The most important environmental variables influencing species richness were pond area (positively correlated) and TP (negatively correlated) in both types of ponds (Table 3). These two variables explained 64.51% and 84.62% of the variation in species richness among permanent and temporary waterbodies, respectively.
Community and environmental heterogeneities in temporary and permanent ponds
PERMANOVA showed significant differences (p < 0.05) in abundance-based beta diversity of macrophyte community between temporary and permanent ponds. (Table 4). The differences between the two pond types were due to compositional differences (location of temporary and permanent ponds in multivariate space) and to differences in the relative dispersion of ponds within each group (PERMDISP F = 7.32, p = 0.028), which was lower for temporary ponds (Fig. 4A). In addition, significant differences in the environmental conditions between the pond types and in their multivariate dispersion (PERMDISP, F = 4.41, p = 0.047) were found. Permanent ponds were significantly more heterogeneous in their environmental conditions than temporary ponds (Table 4, Fig. 4B).

Heterogeneity of macrophyte communities (a) and environmental conditions (b) visualized in an ordination space of principal coordinate analysis based on Bray–Curtis dissimilarity and Euclidean distances. Vectors connect particular ponds with their centroids
Partitioning beta diversity. Relative contribution of environmental and spatial factors to community structuring
Using the multiple site framework and the pairwise dissimilarity approach proposed by Baselga (2013, 2017), the dissimilarities in species composition and abundance among ponds (βBC, βbc) were found to be slightly higher in permanent than in temporary ponds (Table 5). Both, pairwise and multiple-site measures revealed that among-assemblage dissimilarities were explained (in both types of ponds) by balanced variation in abundance (βbc.bal, βBC.BAL), analogous to spatial turnover, ─rather than abundance gradients (βbc.gra, βBC.GRA), analogous to nestedness—in both types of ponds (Table 5).
The RELATE test conducted on the variable sets selected by BIOENV (Table 6) revealed that variation in species abundance (beta diversity) and its two components were significantly correlated with local environmental variability (p < 0.05) in permanent ponds. However, we considered that the relationship between environmental factors and abundance gradients was of little relevance because of the small contribution of this component to global beta-diversity. None of the biological dissimilarity matrices were significantly related to spatial distances between ponds. Similarly, in temporary ponds, both Bray–Curtis dissimilarity and the balanced variation in abundance were linked to environmental heterogeneity, but no significant correlation between spatial distances and Bray–Curtis dissimilarity or its two components was detected (Table 6).
Discussion
Macrophyte species richness and composition in temporary and permanent ponds
Hydroperiod is known to be one of the key factors controlling macrophyte assemblage composition and diversity patterns (Maltchick et al. 2007; Rolon et al. 2008; Van Geest et al. 2005). Drought is the principal environmental filter for macrophytes living in temporary waters (Welborn et al. 1996), especially in Mediterranean regions (Grillas and Roché 1997). This is apparent in our study in the differences between permanent and temporay ponds. Permanent ponds in the study area supported richer macrophyte assemblages than temporary ones as previously found by Della Bella et al. (2008), Gioria et al. (2010), and Rolon et al. (2008). Average pond area and number of transects per pond were similar in both pond types. Thus, it can be assumed that higher richness values in permanent ponds were not due to pond size or sampling effort. Presence of water throughout the year allowed for the development in permanent ponds of hydrophytes with long life cycles requiring permanent submergence, like Myriophyllum verticillatum L., Ch. hispida, Groenlandia densa (L.) Fourr, Zannichellia palustris L., Zannichellia. peltata Bertol, M. spicatum or C. demersum, absent from the temporary ponds in the study area. These findings are consistent with Pätzig et al. (2012), who found that hydrophytes are positively linked to hydroperiod length. However, some hydrophyte species survived in temporary ponds thanks to their fast-growing cycles (e.g. Ranunculus peltatus Schrank and Ranunculus trichophyllus Chaix) and their capability to develop terrestrial forms and to live on wet soils (e.g. P. amphibia, Potamogeton natans L.)
Environmental drivers of macrophyte community richness
Pond area and trophic status (measured as TP concentration) were the variables best explaining the variation in species richness in both types of ponds. The positive relationship between area and richness of aquatic plants is well known (e. g. Bosiacka and Pienkowski 2012; Della Bella et al. 2008). In fact, the species-area relationship is considered one of the most robust generalizations in ecological theory (He and Legendre 2002). Likewise, trophic status has often proved to be a good predictor of macrophyte richness in wetlands (e.g. Akasaka et al. 2010; Lougheed et al. 2001). Waters with high nutrient content are usually species-poor (Della Bella et al. 2008; Jeppesen et al. 2000). However, small water bodies naturally tend to be more nutrient-rich than large lakes while supporting higher species richness (Davies et al. 2008; Williams et al. 2004). In the study area a significant decline in macrophyte species richness with increasing TP in both temporary and permanent ponds was observed. As shown by other authors (Della Bella et al. 2008; Vestergaard and Sand-Jensen 2000), the highest richness values were found in mesotrophic or moderately eutrophic ponds (45–80 μg L−1 TP), that is, in the lower end of the available trophic gradient in the study area.
Community and environmental heterogeneities in temporary and permanent ponds
In spite of the small spatial scale of the study, Bray–Curtis dissimilarity revealed a high spatial variability in macrophyte assemblages in both pond groups. This supports the assumption that ponds are environmentally heterogeneous and thus strongly contribute to freshwater diversity at the regional level (Williams et al. 2004). It also agrees with the perception that macrophyte assemblages comprise a heterogeneous group of species with varying adaptations and tolerances to flooding and soil saturation or hydrological variations (Cronk and Fennessy 2001). Our results, however, point to the fact that such assemblage heterogeneity is not equally high in permanent and temporary ponds.
Hydroperiod has been mentioned as a variable strongly influencing chemical characteristics of aquatic habitats (Escalera-Vázquez and Zambrano 2010). As expected, chemical characteristics were more heterogeneous across permanent than temporary ponds. Such high environmental heterogeneity in permanent ponds was likely caused by differences in the relative contribution of inflows with different origin (leakage from the Castile Channel, rainfall, irrigation channels, and surface runoff). For example, the water coming from the Castile Channel is of higher quality than that from farmland runoff, small streams or irrigation channels, more affected by intense agricultural activity around the ponds. These chemical differences among permanent ponds are enhanced by hydrological differences since volume reduction in summer was not the same for all the ponds. Given the strong influence of chemical characteristics on composition and richness of aquatic plant assemblages (Alahuhta 2015; Fernández-Aláez et al. 2018; Vestergaard and Sand-Jensen 2000), the increased beta diversity in permanent ponds can be seen as a natural consequence of differential response to the environmental gradients of species with different ecological requirements (Chase and Leibold 2003).
In spite of the differences in flooding duration that can be expected among temporary ponds, they were less heterogeneous than permanent ponds in both environmental and biological characteristics, as similarly found by Chase (2003) and Lopes et al. (2014). Periodical drought has been postulated as a factor imposing environmental filters with strong capacity to select species from the regional pool (Chase 2003, 2007; Welborn et al. 1996) resulting in more similar niche-assembled communities (Chase 2007) and in an inverse relationship between beta diversity and disturbance intensity (Chase 2003, 2007).
Partitioning beta diversity. Relative contribution of environmental and spatial factors to community structuring
The overall beta diversity originated almost entirely from balanced variation in abundance in both pond types. This substitution pattern is equivalent to the species turnover often reported for studies of beta diversity in macrophyte assemblages based on incidence-based measures of dissimilarity (Alahuhta et al. 2017; Boschilia et al. 2016). Furthermore, the results obtained in the study area reinforce the idea stressed by Baselga (2010) that beta diversity in southern Europe responds to substitution rather than subset patterns.
On the other hand, for a good understanding of metacommunity structure it is essential to disentangle the relative contribution of environmental and spatial components to the variation of communities. High values of beta diversity, as in this study, could be attributed to deterministic processes. Under this niche-based view, environmental filters along with biotic interactions would be responsible for the community composition (Leibold et al. 2004). In contrast to the indications made by Padial et al. (2014), no significant relationship between geographical distances and assemblage heterogeneity was found. Such a result may be anticipated given the small spatial scale of the study (maximum distance between ponds around 70 km) and the absence of geographical barriers. Besides, the propagules of passive dispersers, such as macrophytes, can be dispersed over long distances by different vectors (Soomers et al. 2013; Soons et al. 2008).
Local environmental variables were the dominant drivers structuring overall beta diversity, as well as substitution and subset patterns in the two pond types. Relationships were more significant in temporary ponds. Therefore, species sorting, operating through niche processes, was the dominant mechanism responsible for the dynamics of macrophyte assemblages and the high beta diversity among ponds separated by tens of kilometers (regional scale). Similar results have often been reported for macrophyte assemblages (e.g. Alahuhta et al. 2015; O’Hare et al. 2012) and other biological communities (Heino and Tolonen 2017; Vanormelingen et al. 2008).
The environmental factors responsible for the spatial patterns of assemblage heterogeneity were different for each pond type. In permanent ponds, the variation in macrophyte beta diversity and in the patterns of spatial substitution were mainly explained by ionic content (calcium, alkalinity, conductivity) and SRP. High ionic content in aquatic habitats is common in agricultural areas (Atkinson et al. 2011; Kissoon et al. 2013) and its role in shaping macrophyte assemblages is well known (e.g. Penning et al. 2008; Rolon et al. 2008; Vesstergaard and Sand-Jensen 2000). The relevance of calcium and alkalinity, in particular, is due to the differences among macrophyte species in the forms of carbon they assimilate (Maberly and Madsen 2002). Conductivity is closely related to alkalinity and calcium and is globally recognized as a good predictor of macrophyte composition (Rolon et al. 2008). By contrast, in temporary ponds, the spatial beta diversity patterns were closely related to trophic status, as deduced from the fact that the variables included in the best environmental distance matrices were turbidity, TSS, ammonium, potassium and TP. Runoff from agricultural lands together with the effect of livestock activities and grazing are probably contributing to increase turbidity and nutrient concentration, especially at the end of spring when the ponds, just before drying up, have very low depth and volume. Our results seem to agree with previous research (Fernández-Aláez et al. 2018; Lougheed et al. 2001; Penning et al. 2008) on the fact that land use, particularly agriculture and livestock, can cause substantial changes in the abundance and composition of aquatic vegetation.
In conclusion, we found that temporary ponds were environmentally and biologically less heterogeneous and supported lower richness than permanent ponds. Furthermore, they rarely contributed species not present in permanent ponds. Macrophyte beta diversity patterns were mainly driven by species substitution and not by subset patterns in both permanent and temporary ponds. This suggests that pond clusters including both temporary and permanent habitats comprising wide environmental gradients within a region are essential for a high regional biodiversity (Baselga 2010). At the scale of our study, and in the absence of geographical barriers, spatial effects on beta heterogeneity were negligible. Under these conditions, the substitution patterns among ponds are mostly explained by species sorting. Therefore, local environmental conditions impact the degree of uniqueness of local macrophyte communities and hence the importance of environmental heterogeneity for maintaining community diversity.
References
Akasaka M, Takamura N, Mitsuhashi H, Kadono Y (2010) Effects of land use on aquatic macrophyte diversity and water quality of ponds. Freshw Biol 55:909–922
Alahuhta J (2015) Geographic patterns of lake macrophyte communities and species richness at regional scale. J Veg Sci 26:564–575
Alahuhta J, Kosten S, Akasaka M, Auderset D, Azzella M, Bolpagni R, Heino J (2017) Global variation in the beta diversity of lake macrophytes is driven by environmental heterogeneity rather than latitude. J Biogeogr Biogeogr 44:1758–1769
Alahuhta J, Rääpysjärvi J, Hellsten S, Kuoppala M, Aroviita J (2015) Species sorting drives variation of boreal lake and river macrophyte communities. Commun Ecol 16:76–85
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Aust Ecol 26:32–46
Anderson MJ, Crist TO, Chase JM, Vellend M, Inouye BD, Freestone AL, Sanders NJ, Cornell HV, Comita LS, Davies KF, Harrison SP, Kraft NJB, Stegen JC, Swenson NG (2011) Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist. Ecol Lett 14:19–28
Anderson MJ, Ellingsen KE, McArdle BH (2006) Multivariate dispersion as a measure of beta diversity. Ecol Lett 9:683–693
APHA (1989) Standard methods for the examination of water and wastewater, 17th edn. American Public Health Association, Washington DC
Atkinson CL, Golladay SW, First MR (2011) Water quality and planktonic microbial assemblages of isolated wetlands in an agricultural landscape. Wetlands 31:885–894
Baselga A (2010) Partitioning the turnover and nestedness components of beta diversity. Glob Ecol Biogeogr 19:134–143
Baselga A (2013) Separating the two components of abundance-based dissimilarity: balanced changes in abundance vs. abundance gradients. Methods Ecol Evol 4:552–557
Baselga A (2017) Partitioning abundance-based multiple-site dissimilarity into components: balanced variation in abundance and abundance gradients. Methods Ecol Evol 8:799–808
Baselga A, Orme CDL, Villéger S, De Bortoli J, Leprieur F (2017) betapart: partitioning beta diversity into turnover and nestedness components. R package version 1.4. https://CRAN.R-project.org/package=betapart
Boschilia SM, de Oliveira EF, Schwarzbold A (2016) Partitioning beta diversity of aquatic macrophyte assemblages in a large subtropical reservoir: prevalence of turnover or nestedness? Aquat Sci 78:615–625
Bosiacka B, Pienkowski P (2012) Do biogeographic parameters matter? Plant species richness and distribution of macrophytes in relation to area and isolation of ponds in NW Polish agricultural landscape. Hydrobiologia 689:79–90
Castroviejo S et al. (Eds.) (1986–2013) Flora Iberica: Plantas vasculares de la Península Ibérica e Islas Baleares, vols. 1–8, 10–15, 17–18, 20, 21. Real Jardín Botánico C.S.I.C, Madrid, Spain.
CEC (1995) Wise use and conservation of wetlands, COM (95) 18 a Final. Office for Official Publications of the European Communities, Luxembourg
Chase JM (2003) Community assembly: when should history matter? Oecologia 136:489–498
Chase JM (2007) Drought mediates the importance of stochastic community assembly. Proc Natl Acad Sci USA 104:17430–17434
Chase JM, Leibold MA (2003) Ecological niches: linking classical and contemporary approaches. University of Chicago Press, Chicago
Cirujano S, Cambra J, Sánchez-Castillo PM, Meco A, Flor-Arnau N (2008) Flora Iberica, Algas Continentales: Carófitos (Characeae). Real Jardín Botánico, CSIC, Madrid, Spain
Cirujano S, Meco A, García P, Chirino M (2014) Flora acuática española Hidrófitos vasculares. Real Jardín Botánico, CSIC, Madrid
Clarke KR, Ainsworth M (1993) A method of linking multivariate community structure to environmental variables. Mar Ecol Prog Ser 92:205–219
Clarke KR, Warwick RM (2010) Change in marine communities: an approach to Statistical Analysis and Interpretation, 2nd edn. Primer-E Ltd, Plymouth
Cronk JK, Fennessy MS (2001) Wetland plants: biology and ecology. Lewis Publisher, New York
Davies BR, Biggs J, Williams P, Whitfield M, Nicolet P, Sear D, Bray S, Maund S (2008) Comparative biodiversity of aquatic habitats in the European agricultural landscape. Agr Ecosyst Environ 125:1–8
De Meester L, Declerck S, Stoks R, Louette G, Van de Meutter F, de Bie T, Michels E, Brendonck L (2005) Ponds and pools as model systems in conservation biology, ecology and evolutionary biology. Aquatic Conserv Mar Freshw Ecosyst 15:715–725
Della Bella V, Bazzanti M, Dowgiallo MG, Iberite M (2008) Macrophyte diversity and physico-chemical characteristics of Tyrrhenian coast ponds in central Italy: implications for conservation. Hydrobiologia 597:85–95
Escalera-Vázquez LH, Zambrano L (2010) The effect of seasonal variation in abiotic factors on fish community structure in temporary and permanent pools in a tropical wetland. Freshw Biol 55:2557–2569
Fernández-Aláez C, Fernández-Aláez M, García-Criado F, García-Girón J (2018) Environmental drivers of aquatic macrophyte assemblages in ponds along an altitudinal gradient. Hydrobiologia 812:79–98
Fernández-Aláez C, Fernández-Aláez M, Santiago NF, Núñez G, Aboal M (2012) ID-tax. Catálogo y claves de identificación de organismos del grupo macrófitos utilizados como elementos de calidad en las redes de control del estado ecológico. Ministerio de Agricultura, Alimentación y Medio Ambiente, Madrid, Spain
Florencio M, Serrano L, Gómez-Rodríguez C, Millán A, Díaz-Paniagua C (2009) Inter and intra-annual variations of macroinvertebrate assemblages are related to the hydroperiod in Mediterranean temporary ponds. Hydrobiologia 634:167–183
Gioria M, Schaffers A, Bacaro G, Feehan J (2010) The conservation value of farmland ponds: predicting water beetle assemblages using vascular plants as a surrogate group. Biol Cons 143:1125–1133
Grillas P, Gauthier P, Yavercovski N, Perennou C (2004) Mediterranean Temporary Pools, Volume 1 Issues relating to conservation, functioning and management. Station biologique de la Tour du Valat, Arles, France
Grillas P, Roché J (1997) Vegetation of temporary marshes. Ecology and Management, Station Biologique de la Tour du Valat, Arles, France
He F, Legendre P (2002) Species diversity patterns derived from species-area models. Ecology 83:1185–1198
Heino J, Tolonen KT (2017) Ecological drivers of multiple facets of beta diversity in a lentic macroinvertebrate metacommunity. Limnol Oceanogr 62:2431–2444
Jeffries M (2008) The spatial and temporal heterogeneity of macrophyte communities in thirty small, temporary ponds over a period of ten years. Ecography 31:765–775
Jensén S (1977) An objective method for sampling the macrophyte vegetation in lakes. Vegetatio 33:107–118
Jeppesen E, Jensen JP, Søndergaard M, Lauridsen T, Landkildehus F (2000) Trophic structure, species richness and biodiversity in Danish lakes: changes along a phosphorus gradient. Freshw Biol 45:201–213
de Castilla J, y León, (2010) Plan de Gestión de la Zonas Húmedas del Canal de Castilla. Inventario y diagnóstico. Patrimonio Natural de Castilla y León, Valladolid, Spain, Tomo I
Kissoon LTT, Jacob DL, Hanson MA, Herwig BR, Bowe SE, Otte ML (2013) Macophytes in shallow lakes: relationships with water, sediment and watershed characteristics. Aquat Bot 109:39–48
Legendre P (2014) Interpreting the replacement and richness difference components of beta diversity. Glob Ecol Biogeogr 23:1324–1334
Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, Hoopes MF, Holt RD, Shurin JB, Law R, Tilman D, Loreau M, Gonzalez A (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecol Lett 7:601–613
Lopes PM, Bini LM, Declerck SAJ, Farjalla VF, Vieira LCG, Bonecker CC, Lansac-Toha AA, Esteves FA, Bozelli RL (2014) Correlates of zooplankton beta diversity in tropical lake systems. PLoS ONE. https://doi.org/10.1371/journal.pone.0109581
Lougheed VL, Crosbie B, Chow-Fraser P (2001) Primary determinants of macrophyte community structure in 62 marshes across the Great Lakes basin: latitude, land use, and water quality effects. Can J Fish Aquat Sci 58:1603–1612
Maberly SC, Madsen TV (2002) Freshwater angiosperm carbon concentrating mechanisms: processes and patterns. Funct Plant Biol 29:393–405
Maltchik L, Rolon AS, Schott P (2007) Effects of hydrological variation on the aquatic plant community in a floodplain palustrine wetland of Southern Brasil. Limnology 8:23–28
O’Hare MT, Gunn IDM, Chapman DS, Dudley BJ, Purse BV (2012) Impacts of space, local environment and habitat connectivity on macrophyte communities in conservation lakes. Divers Distrib 18:603–614
Oertli B, Céréghino R, Hull A, Miracle R (2009) Pond conservation: from science to practice. Hydrobiologia 634:1–9
Padial AA, Ceschin F, Declerck AAJ, De Meester L, Boncker CC, Lansac-Tôha FA, Rodrigues L, Rodrigues LC, Train S, Velho LFM, Bini LM (2014) Dispersal ability determines the role of environmental, spatial and temporal drivers of metacommunity structure. PLoS ONE. https://doi.org/10.1371/journal.pone.0111227
Pätzig M, Kaletta T, Glemnitz M, Berger G (2012) What governs macrophyte species richness in kettle hole types? A case study from Northeast Germany. Limnologica 42:340–354
Penning WE, Mjelde M, Dudley B, Hellsten S, Hanganu J, Kolada A, van den Berg M, Poikane S, Phillips G, Willby N, Ecke F (2008) Classifying aquatic macrophytes as indicators of eutrophication in European lakes. Aquat Ecol 42:237–251
Rolon AS, Lacerda T, Maltchik L, Guadagnin DL (2008) Influence of area, habitat and water chemistry on richness and composition of macrophyte assemblages in southern Brazilian wetlands. J Veg Sci 19:221–228
Scheffer M, Van Geest GJ, Zimmer K, Jeppesen E, Søndergaard M, Butler MG, Hanson MA, Declerck S, De Meester L (2006) Small habitat size and isolation can promote species richness: second-order effects on biodiversity in shallow lakes and ponds. Oikos 112:227–231
Socolar JB, Gilroy JJ, Kunin WE, Edwards DP (2016) How should beta-diversity inform biodiversity conservation? Trends Ecol Evol 31:67–80
Soomers H, Karssenberg D, Soons MB, Verweij PA, Verhoeven JTA, Wassen J (2013) Wind and water dispersal of wetland plants across fragmented landscapes. Ecosystems 16:434–451
Soons MB, van der Vlugt C, van Lith B, Heil GW, Klaassen M (2008) Small seed size increases the potential for dispersal of wetland plants by ducks. J Ecol 96:619–627
Tonkin JD, Stoll S, Jähnig SC, Haase P (2015) Contrasting metacommunity structure and beta diversity in an aquatic-floodplain system. Oikos 125:686–697
Van den Broeck M, Waterkeyn A, Rhazi L, Grillas P, Brendonck L (2015) Assessing the ecological integrity of endorheic wetlands, with focus on Mediterranean temporary ponds. Ecol Ind 54:1–11
Van Geest GJ, Coops H, Roijackers RMM, Buijse AD, Scheffer M (2005) Succession of aquatic vegetation driven by reduced water-level fluctuations in floodplain lakes. J Appl Ecol 42:251–260
Vanormelingen P, Cottenie K, Michels E, Muylaert K, Vyverman W, De Meester L (2008) The relative importance of dispersal and local processes in structuring phytoplankton communities in a set of highly interconnected ponds. Freshw Biol 53:2170–2183
Vestergaard O, Sand-Jensen K (2000) Aquatic macrophytes richness in danish lakes in relation to alkalinity, transparency, and lake area. Can J Fish Aquat Sci 57:2022–2031
Welborn GA, Skelly DK, Werner EE (1996) Mechanisms creating community structure across a freshwater habitat gradient. Annu Rev Ecol Syst 27:337–363
Williams P, Whitfield M, Biggs J, Bray S, Fox G, Nicolet P, Sear D (2004) Comparative biodiversity of rivers, streams, ditches and ponds in an agricultural landscape in Southern England. Biol Cons 115:329–341
Zokan M, Drake JM (2015) The effect of hydroperiod and predation on the diversity of temporary pond zooplankton communities. Ecol Evolut 5:3066–3074
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Fernández-Aláez, M., García-Criado, F., García-Girón, J. et al. Environmental heterogeneity drives macrophyte beta diversity patterns in permanent and temporary ponds in an agricultural landscape. Aquat Sci 82, 20 (2020). https://doi.org/10.1007/s00027-020-0694-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00027-020-0694-4