
Abstract
Impoundments alter connectivity, sediment transport and water discharge in rivers and floodplains, affecting recruitment, habitat and resource availability for fish including benthic invertivorous fish, which represent an important link between primary producers and higher trophic levels in tropical aquatic ecosystems. We investigated long-term changes to water regime, water quality, and invertivorous fish assemblages pre and post impoundment in three rivers downstream of Porto Primavera Reservoir in south Brazil: Paraná, Baía and Ivinhema rivers. Impacts were distinct in the Paraná River, which is fully obstructed by the dam, less evident in the Baía River which is partially obstructed by the dam, but absent in the unimpounded Ivinhema River. Changes in water regime were reflected mainly as changes in water-level fluctuation with little effect on timing. Water transparency increased in the Paraná River post impoundment but did not change in the Baía and Ivinhema rivers. Changes in fish assemblages included a decrease in benthic invertivorous fish in the Paraná River and a shift in invertivorous fish assemblage structure in the Baía and Paraná rivers but not in the unimpounded Ivinhema River. Changes in water regime and water transparency, caused by impoundment, directly or indirectly impacted invertivorous fish assemblages. Alterations of fish assemblages following environmental changes have consequences over the entire ecosystem, including a potential decrease in the diversity of mechanisms for energy flow. We suggest that keeping existing unimpounded tributaries free of dams, engineering artificial floods, and intensive management of fish habitat within the floodplain may preserve native fish assemblages and help maintain functionality and ecosystem services in highly impounded rivers.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Most of the largest rivers and drainage basins in the world have been dammed (Nilsson et al. 2005; Winemiller et al. 2016). Impoundments, which are lentic bodies of water created by dams, alter the continuity in matter, energy, and nutrient transfer in rivers (Vannote et al. 1980; Ward and Stanford 1983), including modifications in the extent and timing of sediment and nutrient transport, and water discharge (Ward and Stanford 1995a; Friedl and Wüest 2002). Changes in water transparency, nutrient and sediment loads, channel morphology and granulometry are some long-term physico-chemical effects of dams on environments downstream (Ward and Stanford 1995b; Brandt 2000; Manyari and Carvalho Jr. 2007; Wohl et al. 2015), potentially leading to long-term oligotrophication (Ney 1996; Stockner et al. 2000). Changes in hydrology also alter connectivity between floodplains and channels, affecting many ecological processes (Bunn and Arthington 2002; Thomaz et al. 2007). Furthermore, reduction in nutrients typical of reservoir cascades (i.e. a series of impoundments in a river or/and its tributaries) can alter the structure of the primary producer community (Barbosa et al. 1999) and the abundance of consumers, including zooplankton, macroinvertebrates and fish (Sampaio et al. 2002; Hudon et al. 2012; Braghin et al. 2015; Lima et al. 2016; Santos et al. 2016).
In many large rivers, impoundments have changed hydrology by reducing the variability in annual discharge and flood pulses (e.g. Souza Filho et al. 2004; Agostinho et al. 2009), a major driver of ecological processes in associated floodplains (Junk et al. 1989; Junk and Wantzen 2004). Dams have also reduced sediment and nutrient transport leading to reduced turbidity and less dynamic bedload movements (Roberto et al. 2009; Stevaux et al. 2009). The fish community is especially affected by these changes because changing flooding patterns can alter resource availability (Mérona et al. 2005; Luz-Agostinho et al. 2008; Abujanra et al. 2009), reproductive success and recruitment (Agostinho et al. 2004b; Oliveira et al. 2014; Angulo-Valencia et al. 2016), and ultimately fish species composition and abundance.
As an important subset of the fish community, benthic invertivorous fish represent one of the links between primary producers and higher trophic levels in tropical aquatic ecosystems (Hahn et al. 2004; Kramer et al. 2013, 2015). These fish feed on benthic resources that include insect larvae, mollusks and other macro and microinvertebrates (Fugi et al. 2001), which are reportedly affected by changes in sediment granulometry, habitat composition and spatial and temporal heterogeneity (Quinn and Hickey 1990; Takeda and Fujita 2004). Studies have reported the effect of impoundments on detritivores (e.g. Mérona et al. 2001), which are fish that also exploit the bottom, but feed on different resources than invertivorous fish, especially particulates (Hahn et al. 2004).
Considering that the food resources and habitats of invertivorous fish are directly affected by changes caused by river impoundment, we investigated the long-term impact of impoundment on the structure of benthic invertivorous fish assemblages in the floodplain below a major dam. To this end, we examined changes in descriptors of water regime, water quality and assemblage structure of invertivorous fish pre and post-impoundment, in impounded and unimpounded rivers. We expected most descriptors of water regime, water quality and fish assemblages to differ pre and post impoundment in two rivers affected by the impoundment but not in the unimpounded river. Understanding changes in hydrology and biotic communities following river impoundment is not only important to inform assessment of anthropogenic impacts, but also to design conservation strategies to preserve native fish assemblages and floodplain ecosystem services, even in an altered state (Affonso et al. 2015).
Methods
Study area
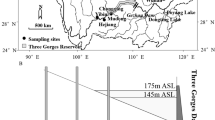
The Upper Paraná River Basin drains approximately 891,000 km2 in the most populated and industrialized region in Brazil, and it is also the most highly impounded basin in South America. This basin includes a 230-km long floodplain in its southwestern section, between the Engineer Sérgio Motta Dam (which impounded the Porto Primavera Reservoir) and the Itaipu Dam, reaching up to 20 km wide in the western margin of the Paraná River. This floodplain stretch has a high diversity of habitats that includes the alluvial plain with numerous secondary channels, floodplain lakes and sections of the main channels of the Paraná River, Baía River and Ivinhema River (Fig. 1). The Paraná River main channel has three other large impoundments upstream of the study area, and most of its tributaries are heavily dammed (Agostinho et al. 2004c; Stevaux et al. 2009), constituting a reservoir cascade. The Baía River parallels the Paraná River and it is also bisected by Porto Primavera, but no other impoundments exist in its main channel. Unlike in the Paraná River, no water is discharged from Porto Primavera Reservoir into the Baía River, but several small unimpounded tributaries contribute to its flow below the reservoir. Water from the Paraná River spills into the Baía during floods or reaches its lower stretch through reverse flow (Comunello et al. 2003). Conversely, the Ivinhema River has its lower stretches inserted in a state conservation unit, and is one of the few remaining dam-free rivers in the Upper Paraná River Basin. In 2006–2010, the mean discharge of the Paraná River in the study area was 8998 m3/s, and the mean discharge in the Ivinhema River was 363 m3/s. No discharge data are available for the Baía River, but discharge is smaller than the Ivinhema River.

Location of sampling sites in the Ivinhema River (unimpounded), Baía River and Paraná River (both impounded), in relation to Porto Primavera Reservoir. Only the lower gauging station of the Ivinhema River is shown
The flood dynamics in the upper Paraná River floodplain are complex. The wet season usually occurs from November to May, but more than one flood pulse can occur during this phase (Agostinho et al. 2009). The flood pulse is critical in terms of maintaining the high biodiversity of the floodplain (Junk et al. 1989; Agostinho et al. 2004c, 2009). The dry season usually occurs from June to October. The annual fluctuations in water level are on average about 2.5 m (Agostinho et al. 2000), but fluctuations as large as 7.5 m have been recorded as well as years with almost no flooding. Overall, the Paraná River exerts the largest influence on the water level in the floodplain; however, local precipitation both in the Baía River and in the Ivinhema River drainage basins also influence floodplain water levels (Souza Filho et al. 2004; Thomaz et al. 2004). Indeed, most floods (40%) occur due to rises in the Paraná River; however, often both the Paraná and Ivinhema rivers cause floods (30%), and the Ivinhema River alone (Comunello et al. 2003) causes some floods (28%).
The floodplain presents a great diversity of habitats. Lentic environments include floodplain lakes; some connect to the river all year, while others are isolated and connected indirectly to the river by groundwater or only during flooding events. Lakes differ in water quality depending on which river they connect to as well as connectivity dynamics (Rocha and Thomaz 2004; Thomaz et al. 2004). The main channels of the Ivinhema, Baía, and Paraná rivers represent lotic environments.
Porto Primavera Reservoir, the largest in the Paraná River and built to support hydropower, navigation and flood control, bisects the Paraná and Baía rivers. The dam was closed in 1998, but some water control had already begun in the early 1990s (BCD 2009). Filling of the reservoir was completed in March 2001. The reservoir has the fifth largest area in Brazil (approximately 2250 km2), extends 240 km upstream of the dam, is 19-m deep at the dam and operates as a run-of-the-river reservoir.
Water regime
Water level data were obtained through the HidroWeb online platform of the National Water Agency (ANA) at one gauging station in the Paraná River (Porto São José Jusante, code 64575003), and two stations in the Ivinhema River (UHE Itaipu Ivinhema, code 64617000, lower station—Fig. 1; Fazenda Ipacaraí, code 64614000, upper station). There are no agency gauging stations in the Baía River. A complete set of daily water levels was available for the Paraná River station from 1974 to 2015. In the Ivinhema River, data were available from 1974 to 2010 at the lower station and from 1984 to 2015 at the upper station. According to year, and according to wet and dry seasons, daily water levels were used to compute eight hydrological descriptors (see definitions in Table 1). Water level was represented by 7-day maximum (7max), 7-day minimum (7min), 30-day maximum (30max), 30-day minimum (30min), median, and seasonal range. The 7- and 30-day maxima and minima were calculated using a moving average procedure (TNC 2009). Median is the seasonal median water level in that year. Seasonal range is the difference between 7-day maximum and 7-day minimum in each season, and reflects the variability of water level within a season. Timing descriptors were represented by seasonal date of maximum level (season day max) and seasonal date of minimum level (season day min). Seasonal dates were computed by transforming Julian days to wet season day (i.e. 1–212, starting on November 1st) and dry season day (i.e. 1–153, starting on June 1st). These descriptors were computed with IHA version 7.1 (Indicators of Hydrological Alteration, Richter et al. 1998).
Water quality and fish
Data on water quality and fish densities were obtained in lotic and lentic floodplain habitats below Porto Primavera Reservoir pre (1986–1988) and post (2000–2015) impoundment, in the Paraná, Baía and Ivinhema river systems. The pre-impoundment sampling period included monthly samples totaling 24 months, whereas the post-impoundment period included quarterly samples totaling 62 months. Each month, water quality and fish samples were collected within the river channel and in floodplain lakes, except in the Paraná River where there was no sampling in lakes pre impoundment. Driven by fiscal cycles, the number of river channel sites sampled each sampling month over this 18-year effort varied from 3 to 4 (mean 3.2 sites/sampling month), and the number of floodplain lakes varied from 4 to 13 (mean 7.2 sites/sampling month).
We collected fish data at each site and recorded water quality simultaneously at most sites. Clusters of eight gill nets each with different mesh sizes (3, 4, 6, 8, 10, 12, 14 or 16 cm between opposite knots) were fished at each site (each net was 20 m long, 1.45–1.70 m deep). Nets were deployed for 24 h in each site, with fish removed every 8 h. Fish collected were identified to species and counted. Water conductivity and transparency (Secchi depth) were measured onsite, and total nitrogen, total phosphorous, and chlorophyll-a were measured in the laboratory from filtered water samples stored at − 20 °C, following the methodology in Mackereth et al. (1978).
The fish assemblage of the Upper Paraná River includes over 182 fish species (Agostinho et al. 2004a; Graça and Pavanelli 2007) We focused on six benthic invertivorous fish species (Fig. 2): Callichthys callichthys (Linnaeus, 1758), Hoplosternum littorale (Hancock, 1828), Iheringichthys labrosus (Lütken, 1874), Pimelodus maculatus Lacepède, 1803, Satanoperca pappaterra (Heckel, 1840) and Trachydoras paraguayensis (Eigenmann & Ward, 1907). These species comprised approximately 14% of the fish collections in 1986–1988 and 8% in 2000–2015. Most of these invertivorous fish have a specialized diet (Hahn et al. 2004). However, two species tend to be more generalists. Satanoperca pappaterra consumes detritus and plant material as well as benthic invertebrates (Hahn and Cunha 2005; Novakowski et al. 2016), and P. maculatus has benthic organisms as a major component of its diet, but also feeds on small fish and plant matter (Lobon-Cerviá and Bennemann 2000; Lima-Junior and Goitein 2003). Satanoperca pappaterra was probably introduced into the study area by aquaculture or fish farming, in the second half of the twentieth century before we began our study (Langeani et al. 2007; Ortega et al. 2015). Pimelodus maculatus migrates moderately long distances for reproduction and feeding (Makrakis et al. 2012), while I. labrosus and T. paraguaysensis migrate short-distances laterally (Bailly et al. 2008; Holzbach et al. 2009); the other study species are not reported to migrate (Mol 1996; Hostache and Mol 1998; Verba et al. 2011).

Illustration of the six invertivorous fish species included in this study. Sizes are drawn to scale of the mean standard lengths recorded during this study
Data analysis
We used a two-way permutation MANOVA (PERMANOVA) to test if water regime indexed by the eight hydrological descriptors changed between periods (i.e. pre and post impoundment) while accounting for possible differences in season (i.e. wet and dry). Temporal correlation in the data does not affect PERMANOVA because randomly permuting the years removes any inherent temporal correlation (Anderson 2001). Each gauging station was analyzed separately. We used all available water years after 1999 to represent post impoundment, while pre impoundment was represented by 1974–1990 water years in the Paraná River and Ivinhema lower station, and 1983–1990 in Ivinhema upper station. The period between 1991 and 1998 was not considered due to possible influences of dam construction. We used a greater number of pre-impoundment years for hydrological analysis than for fish assemblage analysis because more data were available, allowing for a more robust comparison. All hydrological descriptors, except those associated with timing and seasonal range, were log-transformed to reduce skewness, and all variables were normalized (zero mean and unit variance) before computing the resemblance matrix with Euclidean distances.
We assessed changes in water quality and fish using three-way PERMANOVA analyses that tested for differences between time periods in the three rivers, and accounted for any effects of season (i.e. wet and dry) and habitat type (i.e. lentic and lotic). Total nitrogen, total phosphorous, Secchi depth and chlorophyll-a were log-transformed to reduce skewness and all five water quality variables were normalized before the analysis. A resemblance matrix using Euclidean distances was assembled to perform the water quality PERMANOVA.
For fish, we first conducted a permutational ANOVA, with the same factors described above, for total abundance (total catch rate) of all invertivorous fish combined. Next, we performed a PERMANOVA to test for shifts in the six species assemblage. Analyses were applied to catches standardized to 1000 m2 of the gill net cluster by season, water year, river, and habitat type. All catch rates were log-transformed to reduce skewness and the resemblance matrix computed using the Bray-Curtis dissimilarity metric. In the PERMANOVA, a dummy variable was used to include samples containing no invertivorous fish species (11 out of 218 samples) (Clarke et al. 2006) and we used type III sum of squares to account for the unbalanced design. If the PERMANOVA identified a significant (p ≤ 0.05) period × river effect, we performed Principal Coordinate Analysis (PCoA) to visualize differences, using the same resemblance matrices of the respective PERMANOVA. All analyses were performed in the software PRIMER 6.0 with the add-on PERMANOVA, and we report pseudo-F (F) values for the main test and pseudo-t (t) values for pairwise comparisons; 999 unrestricted permutations of raw data were used in the main test as well as in the post-hoc tests for factors and interactions.
Results
Water regime
Water regime changed in the Paraná River pre and post impoundment (F = 5.8, p < 0.01), but not in the upper (F = 0.6, p = 0.59) or lower (F = 2.1, p = 0.08) Ivinhema River. These three analyses also showed significant effects of season (p ≤ 0.01) but no significant (p ≥ 0.41) period × season interactions, suggesting that seasonality did not change post impoundment. Every description of water level decreased in the Paraná River after dam construction (Table 1); however, the timing of seasonal maxima and minima did not change (Fig. 3a). The greatest decreases were observed with median water levels, maximum values of wet season and seasonal range of the wet season (Table 1). However, high overlap of data (Fig. 3a) and dispersion of data (Fig. 3b, c) suggest that discharge varies greatly among years in the same period. Although reductions in water level are apparent, water regimes post dam may resemble those pre dam in some years.

a Ordination plot of water regime descriptors in the Paraná River pre impoundment (1974–1990) and post impoundment (1999–2015) in the wet and dry seasons. Vectors represent correlations of water regime descriptors with PCoA axis 1 and 2. b PCoA 1 axis values for samples pre and post impoundment in wet and dry seasons. c PCoA 2 axis values for samples pre and post impoundment in wet and dry seasons. In b, c squares/circles represent means and whiskers represent 95% confidence intervals
Water quality
Water quality changed with period (F = 3.7, p = 0.01), river (F = 12.9, p < 0.01) and habitat type (F = 12.3, p < 0.01), but not season (F = 2.5, p = 0.07). The river × habitat type interaction was statistically significant (F = 5.5, p < 0.01) indicating that the magnitude of the difference between lentic and lotic environments differed in all three rivers (p < 0.01 for all combinations). The river × period interaction was not statistically significant (F = 1.6, p = 0.14) indicating overall differences between periods were similar among rivers. Secchi depth increased in the Paraná River post impoundment but not in the Baía and Ivinhema rivers (Table 2). Conductivity increased and total phosphorous decreased in all three rivers, while chlorophyll-a and total nitrogen did not change.
Invertivorous fish assemblage
In all, we collected 12,878 invertivorous fish. The average total catch rate of invertivorous fish differed between periods, and type of habitat (Table 3), while the effects of river and season were not statistically significant. The interaction terms period × river and river × habitat (Table 3) were statistically significant. Post-hoc comparisons showed that average catch rate of all invertivorous fish combined did not change pre and post dam in the Ivinhema River (t = 0.1, p = 0.90) nor in the Baía River (t = 1.6, p = 0.10) but it decreased from 57.5 to 16.5 fish/1000 m2 of gillnet in the Paraná River (t = 4.1, p < 0.01).
Considering both periods, T. paraguayensis represented 31.3% of the total invertivorous fish catch, H. littorale represented 23.3%, I. labrosus 20.7%, P. maculatus 18.3%, S. pappaterra 5.9% and C. callichthys 0.5%. Invertivorous fish assemblages differed between periods, rivers (Fig. 4a), and habitat types but not between seasons (Table 3). The statistically significant interaction period × river indicated that the invertivorous fish assemblage changed differently across rivers after dam construction. Post-hoc multiple comparisons showed that assemblage differed pre and post impoundment in the Paraná (t = 4.9, p < 0.01, Fig. 4d) and Baía rivers (t = 1.9, p = 0.02, Fig. 4c) but not in the Ivinhema River (t = 1.4, p = 0.10, Fig. 4b). The river × habitat type interaction was statistically significant, suggesting that the difference between lentic and lotic environments differed among rivers. The most marked changes occurred for T. paraguayensis, which decreased drastically in the Baía and Paraná rivers, and S. pappaterra, which increased in the Paraná River (Table 2). Iheringichthys labrosus and P. maculatus also decreased in the impounded rivers (Fig. 4c, d; Table 2). The lotic environment in the Paraná River post dam had the smallest catch rate for five of the six species (Table 2).

Ordination plots of the invertivorous fish assemblage a in all three river systems, not separated according to pre and post periods; b in the unimpounded Ivinhema River separated by period; c in the impounded Baía River separated by period; and d in the impounded Paraná River separated by period. Vectors represent Pearson correlations of fish species densities with PCoA axis 1 and 2, and were moved from the center of the graph to improve visualization of text
Discussion
We identified changes to the water regime and invertivorous fish assemblages below the Porto Primavera Reservoir. Changes were evident in the Paraná River, which is distinctly impacted by this impoundment, less obvious in the Baía River which is only partially influenced by the dam, but largely indistinguishable in the unimpounded Ivinhema River. Changes in water regime were reflected by variables descriptive of water-level magnitude, whereas variables descriptive of water-level timing showed little modification. Changes in fish assemblages were reflected by a decrease in catch rate of invertivorous fish in the Paraná River, and a shift in species relative abundances in the Baía and Paraná rivers, but not in the unimpounded Ivinhema River.
Changes in water quality were more ambiguous. A distinct change to water quality directly attributable to impoundment was water transparency, which increased in the Paraná River post dam (also reported by Roberto et al. 2009) but did not change in the Baía and Ivinhema rivers. The other water quality metrics did not change at all or also changed in the unimpounded river. It is unclear whether changes to the unimpounded river were induced by long-term land use changes in the Ivinhema River catchment (Arai et al. 2012) or simply by sampling error attributable to the high variability associated with sampling rather ephemeral water quality metrics. We suggest that changes in water regime and water transparency after the impoundment of Porto Primavera directly or indirectly affected invertivorous fish assemblages in the impounded rivers and have consequences through the entire aquatic ecosystem.
Water regime in the Paraná River changed after the impoundment of Porto Primavera Reservoir, shifting towards lower 7- and 30-day maxima and lower 7- and 30-day minima. Souza Filho et al. (2004), Souza Filho (2009), and Rocha (2010) have reported additional changes through examination of other water metrics, including decreased flood duration and modified discharge-water level relationships. The observed reductions in water levels can lead to loss of aquatic habitat through various mechanisms including (1) decreased connectivity among diverse floodplain habitats (Ward and Stanford 1995b; Agostinho et al. 2004c; Thomaz et al. 2007); and (2) loss of habitat volume through shallower depths (Brandt 2000) and shrinking lateral span. Such changes can cause long-term shifts in invertivorous fish assemblages as well as the entire fish community (Agostinho et al. 2001, 2008).
Differences in the pre and post impoundment structure of benthic invertivorous fish assemblages in the impounded and unimpounded rivers suggest an effect attributable to the change in water regime. The species whose catch rate decreased (namely T. paraguayensis, I. labrosus and P. maculatus) undertake either small or moderate reproductive migrations cued by flooding, stage lateral migrations to feed, and their larvae and juveniles depend on increasing water levels to access nursery areas in backwaters and lakes (Vazzoler 1996; Agostinho et al. 2000, 2001, 2004a; Bailly et al. 2008). Conversely, the only species that increased, particularly in the Paraná River, S. pappaterra, is an equilibrium strategist (i.e. low fecundity, brood protection, non-seasonal spawning; Winemiller 1989; Vazzoler 1996; Verba et al. 2011), thus is virtually independent from flood patterns to reproduce and possibly benefits from flood absence and dry years (Agostinho et al. 2004a, b).
Water quality changes below dams are linked to water storage upstream, discharge regime, and their effects on sediment and nutrient loads (Brandt 2000; Wohl et al. 2015; Agostinho et al. 2016). Reservoir cascades, such as the one in the Paraná River, are common in many large river systems and can have cumulative and synergic effects on the trophic status of downstream reaches (Barbosa et al. 1999; Miranda et al. 2008), because these cascades exacerbate the sediment and nutrient trapping capacities of single reservoirs (Barbosa et al. 1999; Kummu and Varis 2007). The increase in water transparency caused by sediment trapping can increase the abundance of submerged macrophytes (Sousa et al. 2010), which in turn may maintain the clear water state of the environments in the absence of benthic fish (Mormul et al. 2012). Clear water also increases predation pressure on eggs, juveniles, and small-size species (Agostinho et al. 2004a; Turesson and Brönmark 2007; Dodrill et al. 2016). In fact, larval densities have decreased in our study area post closure of Porto Primavera, including those of some invertivorous fish (Sanches et al. 2006), thus decreasing recruitment success and population sizes, especially of species that do not provide parental care (such as T. paraguayensis, I. labrosus and P. maculatus, the invertivorous fish that decreased in our study). The two invertivorous species whose densities were apparently not affected by construction of the Porto Primavera Reservoir (H. littorale and C. callichtys) also depend on flooding to reproduce, but take care of their offspring in nests (Mol 1996; Hostache and Mol 1998), possibly offsetting the effects of increased predation induced by water transparency on reproduction success.
Besides the observed changes in water regime and water quality, damming reportedly causes modifications in channel substrate downstream (Brandt 2000), such as the coarsening of bed material, decreased sediment heterogeneity, and increased bank erosion (Stevaux et al. 2009). These changes directly affect the benthic macroinvertebrate community (Stevaux and Takeda 2002; Bredenhand and Samways 2009), whose composition and abundance are influenced by sediment type and grain-size (Quinn and Hickey 1990; Takeda and Fujita 2004). In turn, these substrate changes affect invertivorous fish populations through food availability and diversity (e.g. Paragamian 2002). Nevertheless, invertivorous fish assemblage structure, as well as that of other fish guilds, is probably indirectly influenced by factors and interactions that we were unable to assess in this study. For example, substrates can be altered by (1) degradation of the littoral zone caused by increased daily variability in water level required by energy production demands (Souza Filho 2009; Poff and Schmidt 2016); and (2) alterations in riparian vegetation caused by longer dry periods and subsequent changes in land use (e.g. expansion of cattle grazing and agriculture in the floodplain). These changes rework the input of allochthonous resources into substrates (Agostinho and Zalewski 1995; Thomaz et al. 2004), changing bottom up regulation of populations and aquatic community production.
Freshwater ecosystems provide many services including biomass production and nutrient cycling (Dobson et al. 2006), which are directly related to energy flow. The relative abundance of organisms in various trophic levels is an important factor in energy flow through the ecosystem (Depczynski et al. 2007). For example, decreased abundance of intermediate predators (such as invertivorous fish) could shorten food chain length (Diehl and Feissel 2001; Post 2002; Hoeinghaus et al. 2008). In fact, the Paraná River was found to have a shorter food chain when compared to the Ivinhema River in our study area (Hoeinghaus et al. 2008), which was attributed to changes in relative importance of different trophic pathways (i.e. based on plant detritus, phytoplankton or periphyton) in river ecosystems after impoundment (Hoeinghaus et al. 2007, 2008). Because invertivorous fish are directly linked to both the detritus and periphyton pathways (Hahn et al. 2004; Lopes et al. 2009), changing their assemblage structure or representation in the fish community could decrease the diversity of mechanisms for energy flow in the ecosystem. These ecosystem changes are expected to influence other fish guilds as well (e.g. Mérona et al. 2005), through either direct impacts on populations or changing biological interactions. For example, in the Upper Paraná River floodplain, detritus processing might have declined due to diminishing populations of the detrivore Prochilodus lineatus caused by decreased access to backwaters (Gubiani et al. 2007), and diet shifts of piscivores have been reported following changing prey assemblages, including decreased consumption of T. paraguayensis (Luz-Agostinho et al. 2008).
The damming of the Upper Paraná River Basin is a big-scale disturbance that has produced big changes in hydrology and fish assemblages, and may require big solutions if fish assemblages are to be preserved until the societal services provided by dams are no longer needed. Various climate change models and scenarios predict moderate increases in precipitation and runoff in the Paraná Basin (Nohara et al. 2006; Marengo et al. 2009, 2012), but likely insufficient to ease the impacts of impoundments on the water regime. Thus, to preserve native fish assemblages further intervention may be needed. A number of big-scale management actions may be applied separately or jointly to address the conservation objective, although practicality and effectiveness may vary. First, keeping existing unimpounded tributaries free of dams can prevent further changes in the natural water regime and biotic assemblages (Affonso et al. 2015; Braghin et al. 2015). Second, engineering artificial floods (e.g. Acreman et al. 2000; Patten et al. 2001) by coordinated water releases from reservoirs throughout the upper Paraná Basin may preserve connectivity in the floodplain (Agostinho et al. 2009; Oliveira et al. 2014). This action may not be feasible if discharge volumes required to engineer sufficiently large and lengthy floods represent a sizeable fraction of the water impounded in the basin, even if the floods are not created annually. Nevertheless, this is an alternative worth exploring with modelling tools (e.g. Rauschenbach 2016) and in collaboration with water resource engineers. Third, intensive management of fish habitat within the floodplain (e.g. Buijse et al. 2002; Gregory et al. 2008; Ollero 2010) to maintain flow and suspended sediment, to retain lateral connectivity between channels and lakes, and to continue access to other habitat needs, even at the reduced water levels dictated by the new water regime, may preserve functionality of the floodplain. Coordinated action among public and private sectors is essential to successfully implement these and other conservation activities.
References
Abujanra F, Agostinho AA, Hahn NS (2009) Effects of the flood regime on the body condition of fish of different trophic guilds in the Upper Paraná River floodplain, Brazil. Brazil J Biol 69:469–479. https://doi.org/10.1590/S1519-69842009000300003
Acreman MC, Farquharson FAK, McCartney MP et al. (2000) Managed flood releases from reservoirs: issues and guidance. Report to DFID and the World Commisson on Dams, Centre for Ecology and Hydrology, Wallingford
Affonso IP, Azevedo RF, Santos NCL dos, Dias RM, Agostinho AA, Gomes LC (2015) Pulling the plug: replicable strategies to preclude dam expansion in Brazilian rivers of high-priority for conservation. Natureza Conservação 13:199–203
Agostinho AA, Zalewski M (1995) The dependence of fish community structure and dynamics on floodplain and riparian ecotone zone in Paraná River, Brazil. Hydrobiologia 303:141–148
Agostinho AA, Thomaz SM, Minte-Vera CV, Winemiller KO (2000) Biodiversity in the High Paraná River Floodplain. In: Gopal B, Junk WJ, Davis JA (eds) Biodiversity in wetlands: assesment, function and conservation, vol 1. Backhuys Publishers, Leiden, pp 89–118
Agostinho AA, Gomes LC, Zalewski M (2001) The importance of floodplains for the dynamics of fish communities of the upper river Paraná. Int J Ecohydrol Hydrobiol 1:209–217
Agostinho AA, Gomes LC, Thomaz SM, Hahn NS (2004a) The Upper Paraná River and its floodplain: main characteristics and perspectives for management and conservation. In: Thomaz SM, Agostinho AA, Hahn NS (eds) The Upper Parana River and its Floodplain: physical aspects, ecology and conservation. Backhuys Publishers, Leiden, pp 381–393
Agostinho AA, Gomes LC, Veríssimo S, Okada EK (2004b) Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Rev Fish Biol Fish 14:11–19. https://doi.org/10.1007/s11160-004-3551-y
Agostinho AA, Thomaz SM, Gomes LC (2004c) Threats for biodiversity in the floodplain of the Upper Paraná River: effects of hydrological regulation by dams. Ecohydrol Hydrobiol 4:255–268
Agostinho A, Pelicice F, Gomes L (2008) Dams and the fish fauna of the Neotropical region: impacts and management related to diversity and fisheries. Brazilian J Biol 68:1119–1132. https://doi.org/10.1590/S1519-69842008000500019
Agostinho AA, Bonecker CC, Gomes LC (2009) Effects of water quantity on connectivity: the case of the upper Paraná River floodplain. Ecohydrol Hydrobiol 9:99–113. https://doi.org/10.2478/v10104-009-0040-x
Agostinho AA, Gomes LC, dos Santos NCL, Ortega JCG, Pelicice FM (2016) Fish assemblages in Neotropical reservoirs: colonization patterns, impacts and management. Fish Res 173:26–36. https://doi.org/10.1016/j.fishres.2015.04.006
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecolo 26:32–46
Angulo-Valencia MA, Agostinho AA, Suzuki HI, da Luz-Agostinho KDG, Agostinho CS (2016) Impoundments affect fish reproduction regardless of reproductive strategy. Lakes Reserv Res Manag 21:362–374. https://doi.org/10.1111/lre.12151
Arai FK, Pereira SB, Gonçalves GGG (2012) Characterization of water availability in a hydrographic basin. Eng Agrícola 32:591–601. https://doi.org/10.1590/S0100-69162012000300018
Bailly D, Agostinho AA, Suzuki HI (2008) Influence of the flood regime on the reproduction of fish species with different reproductive strategies in the Cuiabá River, Upper Pantanal, Brazil. River Res Appl 24:1218–1229. https://doi.org/10.1002/rra
Barbosa FAR, Padisak J, Espíndola ELG et al (1999) The Cascading reservoir continuum concept (CRCC) and its application to the River Tietê basin, São Paulo state, Brazil. In: Tundisi JG, Straskraba M (eds) Theoretical reservoir ecology and its applications. Backhyus Publishers, Leiden, pp 425–437
BCD: Brazilian Committee on Dams (2009) Main Brazilian dams III: design, construction, and performance. BCD, Botafogo, Rio de Janeiro, Brasil. http://www.cbdb.org.br/documentos/mbdiii/porto%20Primavera.pdf. Accessed 16 May 2017
Braghin LSM, Figueiredo BRS, Meurer T et al (2015) Zooplankton diversity in a dammed river basin is maintained by preserved tributaries in a tropical floodplain. Aquat Ecol 49:175–187. https://doi.org/10.1007/s10452-015-9514-7
Brandt SA (2000) Classification of geomorphological effects downstream of dams. Catena 40:375–401. https://doi.org/10.1016/S0341-8162(00)00093-X
Bredenhand E, Samways MJ (2009) Impact of a dam on benthic macroinvertebrates in a small river in a biodiversity hotspot: Cape Floristic Region, South Africa. J Insect Conserv 13:297–307. https://doi.org/10.1007/s10841-008-9173-2
Buijse AD, Coops H, Staras M et al (2002) Restoration strategies for river floodplains along large lowland rivers in Europe. Freshw Biol 47:889–907. https://doi.org/10.1046/j.1365-2427.2002.00915.x
Bunn SE, Arthington AH (2002) Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ Manage 30:492–507. https://doi.org/10.1007/s00267-002-2737-0
Clarke KR, Somerfield PJ, Chapman MG (2006) On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. J Exp Mar Bio Ecol 330:55–80. https://doi.org/10.1016/j.jembe.2005.12.017
Comunello E, Souza Filho EE, Rocha PC, Nanni MR (2003) Dinâmica de inundação de áreas sazonalmente alagáveis na planície aluvial do Alto Rio Paraná: estudo preliminar. In: Anais do 11° Simpósio brasileiro de sensoriamento remoto. São José Dos Campos, Inpe, pp 2459–2466
Depczynski M, Fulton CJ, Marnane MJ, Bellwood DR (2007) Life history patterns shape energy allocation among fishes on coral reefs. Oecologia 153:111–120. https://doi.org/10.1007/s00442-007-0714-2
Diehl S, Feissel M (2001) Intraguild prey suffer from enrichment of their resources: a microcosm experiment with ciliates. Ecology 82:2977–2983. https://doi.org/10.1890/0012-9658(2001)082
Dobson A, Lodge D, Alder J et al (2006) Habitat loss, trophic collapse, and the decline of ecosystem services. Ecology 87:1915–1924
Dodrill MJ, Yard MD, Pine WE (2016) Assessing predation risks for small fish in a large river ecosystem between contrasting habitats and turbidity conditions. Am Midl Nat J 175:206–221
Friedl G, Wüest A (2002) Disrupting biogeochemical cycles: consequences of damming. Aquat Sci 64:55–65. https://doi.org/10.1007/s00027-002-8054-0
Fugi R, Agostinho AA, Hahn NS (2001) Trophic morphology of five benthic-feeding fish species of a tropical floodplain. Braz J Biol 61:27–33. https://doi.org/10.1590/S0034-71082001000100005
Graça WJ, Pavanelli CS (2007) Peixes da planície de inundação do alto rio Paraná e áreas adjacentes. EDUEM, Maringá
Gregory KJ, Benito G, Downs PW (2008) Applying fluvial geomorphology to river channel management: background for progress towards a palaeohydrology protocol. Geomorphology 98:153–172. https://doi.org/10.1016/j.geomorph.2007.02.031
Gubiani ÉA, Gomes LC, Agostinho AA, Okada EK (2007) Persistence of fish populations in the upper Paraná River: effects of water regulation by dams. Ecol Freshw Fish 16:191–197. https://doi.org/10.1111/j.1600-0633.2007.00211.x
Hahn NS, Cunha F (2005) Feeding and trophic ecomorphology of Satanoperca pappaterra (Pisces, Cichlidae) in the Manso Reservoir, Mato Grosso State, Brazil. Brazilian Arch Biol Technol 48:1007–1012. https://doi.org/10.1590/S1516-89132005000800017
Hahn NS, Fugi R, Andrian IF (2004) Trophic ecology of the fish assemblages. In: Thomaz SM, Agostinho AA, Hahn NS (eds) The Upper Parana River and its Floodplain: physical aspects, ecology and conservation. Backhuys Publishers, Leiden, pp 247–270
Hoeinghaus DJ, Winemiller KO, Agostinho AA (2007) Landscape-scale hydrologic characteristics differentiate patterns of carbon flow in large-river food webs. Ecosystems 10:1019–1033. https://doi.org/10.1007/s10021-007-9075-2
Hoeinghaus DJ, Winemiller KO, Agostinho AA (2008) Hydrogeomorphology and river impoundment affect food-chain length of diverse Neotropical food webs. Oikos 117:984–995. https://doi.org/10.1111/j.0030-1299.2008.16459.x
Holzbach AJ, Gubiani ÉA, Baumgartner G (2009) Iheringichthys labrosus (Siluriformes: Pimelodidae) in the Piquiri River, Paraná, Brazil: population structure and some aspects of its reproductive biology. Neotrop Ichthyol 7:55–64. https://doi.org/10.1590/S1679-62252009000100008
Hostache G, Mol JH (1998) Reproductive biology of the neotropical armoured catfish Hoplosternum littorale (Siluriformes—Callichthyidae): a synthesis stressing the role of the floating bubble nest. Aquat Living Resour 11:173–185
Hudon C, Cattaneo A, Poirier AMT et al (2012) Oligotrophication from wetland epuration alters the riverine trophic network and carrying capacity for fish. Aquat Sci 74:495–511. https://doi.org/10.1007/s00027-011-0243-2
Junk WJ, Wantzen KM (2004) The flood pulse concept: new aspects, approaches and applications—an update. In: Welcomme RL, Petr T (eds) Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries. Food and Agriculture Organization and Mekong River Commission, FAO Regional Office for Asia and the Pacific, Bangkok, pp 117–149
Junk WJ, Bayley PB, Sparks RE (1989) The flood pulse concept in river-floodplain systems. In: Dodge PD (ed) Proceedings of the international large river symposium, Canadian special publication of fisheries and aquatic sciences, pp 110–127
Kramer MJ, Bellwood O, Bellwood DR (2013) The trophic importance of algal turfs for coral reef fishes: the crustacean link. Coral Reefs 32:575–583. https://doi.org/10.1007/s00338-013-1009-1
Kramer MJ, Bellwood O, Fulton CJ, Bellwood DR (2015) Refining the invertivore: diversity and specialisation in fish predation on coral reef crustaceans. Mar Biol 162:1779–1786. https://doi.org/10.1007/s00227-015-2710-0
Kummu M, Varis O (2007) Sediment-related impacts due to upstream reservoir trapping, the Lower Mekong River. Geomorphology 85:275–293. https://doi.org/10.1016/j.geomorph.2006.03.024
Langeani F, Castro RMC, Oyakawa OT et al (2007) Diversidade da ictiofauna do Alto Rio Paraná: composição atual e perspectivas futuras. Biota Neotrop 7:181–197. https://doi.org/10.1590/S1676-06032007000300020
Lima AC, Agostinho CS, Soares AMVM., Monaghan KA (2016) Alternative ways to measure impacts of dam closure to the structure of fish communities of a neotropical river. Ecohydrology 9:860–870. https://doi.org/10.1002/eco.1688
Lima-Junior SE, Goitein R (2003) Ontogenetic diet shifts of a Neotropical catfish, Pimelodus maculatus (Siluriformes, Pimelodidae): an ecomorphological approach. Environ Biol Fishes 68:73–79. https://doi.org/10.1023/A:1026079011647
Lobon-Cervia J, Bennemann S (2000) Temporal trophic shifts and feeding diversity in two sympatric, neotropical, omnivorous fishes: Astyanax bimaculatus and Pimelodus maculatus in Tibagi Rio (Parana, Southern Brazil). Arch für Hydrobiol 149:285–306. https://doi.org/10.1127/archiv-hydrobiol/149/2000/285
Lopes CA, Benedito E, Martinelli LA (2009) Trophic position of bottom-feeding fish in the Upper Paraná River floodplain. Brazi J Biol 69:573–581
Luz-Agostinho KDG, Agostinho AA, Gomes LC, Júlio HF Jr (2008) Influence of flood pulses on diet composition and trophic relationships among piscivorous fish in the upper Parana River floodplain. Hydrobiologia 607:187–198. https://doi.org/10.1007/s10750-008-9390-4
Mackereth FJH, Heron J, Talling JF (1978) Water analysis: some revised methods for limnologists, vol 36. Freshwater Biological Association, Scientific Publication, Cumbria
Makrakis MC, Miranda LE, Makrakis S et al (2012) Diversity in migratory patterns among Neotropical fishes in a highly regulated river basin. J Fish Biol 81:866–881. https://doi.org/10.1111/j.1095-8649.2012.03346.x
Manyari WV, de Carvalho OA (2007) Environmental considerations in energy planning for the Amazon region: downstream effects of dams. Energy Policy 35:6526–6534. https://doi.org/10.1016/j.enpol.2007.07.031
Marengo JA, Jones R, Alves LM, Valverde MC (2009) Future change of temperature and precipitation extremes in South America as derived from the PRECIS regional climate modeling system. Int J Climatol 29:2241–2255. https://doi.org/10.1002/joc.1863
Marengo JA, Chou SC, Kay G et al (2012) Development of regional future climate change scenarios in South America using the Eta CPTEC/HadCM3 climate change projections: climatology and regional analyses for the Amazon, São Francisco and the Paraná River basins. Clim Dyn 38:1829–1848. https://doi.org/10.1007/s00382-011-1155-5
Mérona B de, dos Santos GM, de Almeida RG (2001) Short term effects of Tucurui Dam (Amazonia, Brazil) on the trophic organization of fish communities. Environ Biol Fishes 60:375–392. https://doi.org/10.1023/A:1011033025706
Mérona B de, Vigouroux R, Tejerina-Garro FL (2005) Alteration of fish diversity downstream from Petit-Saut Dam in French Guiana: implication of ecological strategies of fish species. Hydrobiologia 551:33–47. https://doi.org/10.1007/s10750-005-4448-z
Miranda LE, Habrat MD, Miyazono S (2008) Longitudinal gradients along a reservoir cascade. Trans Am Fish Soc 137:1851–1865. https://doi.org/10.1577/T07-262.1
Mol JH (1996) Reproductive seasonality and nest-site differentiation in three closely related armoured catfishes (Siluriformes: Callichthyidae). Environ Biol Fishes 45:363–381. https://doi.org/10.1007/BF00002529
Mormul RP, Thomaz SM, Agostinho AA et al (2012) Migratory benthic fishes may induce regime shifts in a tropical floodplain pond. Freshw Biol 57:1592–1602. https://doi.org/10.1111/j.1365-2427.2012.02820.x
Ney JJ (1996) Oligotrophication and its discontents: effects of reduced nutrient loading on reservoir fisheries. Am Fish Soc Symp 16:285–295
Nilsson C, Reidy CA, Dynesius M, Revenga C (2005) Fragmentation and flow regulation of the world’s large river systems. Science 308:405–408. https://doi.org/10.1126/science.1107887
Nohara D, Kitoh A, Hosaka M, Oki T (2006) Impact of climate change on river discharge projected by multimodel ensemble. J Hydrometeorol 7:1076–1089. https://doi.org/10.1175/JHM531.1
Novakowski GC, Cassemiro FAS, Hahn NS (2016) Diet and ecomorphological relationships of four cichlid species from the Cuiabá River basin. Neotrop Ichthyol 14:e150151. https://doi.org/10.1590/1982-0224-20150151
Oliveira AG, Suzuki HI, Gomes LC, Agostinho AA (2014) Interspecific variation in migratory fish recruitment in the Upper Paraná River: effects of the duration and timing of floods. Environ Biol Fishes 98:1327–1337. https://doi.org/10.1007/s10641-014-0361-5
Ollero A (2010) Channel changes and floodplain management in the meandering middle Ebro River, Spain. Geomorphology 117:247–260. https://doi.org/10.1016/j.geomorph.2009.01.015
Ortega JCG, Júlio HF, Gomes LC, Agostinho AA (2015) Fish farming as the main driver of fish introductions in Neotropical reservoirs. Hydrobiologia 746:147–158. https://doi.org/10.1007/s10750-014-2025-z
Paragamian VL (2002) Changes in the species composition of the fish community in a reach of the Kootenai River, Idaho, after construction of Libby Dam. J Freshw Ecol 17:375–383. https://doi.org/10.1080/02705060.2002.9663911
Patten DT, Harpman DA, Voita MI, Randle TJ (2001) A managed flood on the Colorado River: background, objectives, design, and implementation. Ecol Appl 11:635–643
Poff NL, Schmidt JC (2016) How dams can go with the flow. Science 353:1099–1100
Post DM (2002) The long and short of food-chain length. Trends Ecol Evol 17:269–277. https://doi.org/10.1016/j.cell.2013.04.003
Quinn JM, Hickey CW (1990) Magnitude of effects of substrate particle size, recent flooding and catchment development on benthic invertebrates in 88 New Zealand rivers. New Zeal J of Mar Freshw Res 24:411–427. https://doi.org/10.1080/00288330.1990.9516433
Rauschenbach T (2016) Modeling, control, and optimization of water systems. Springer, New York
Richter BD, Baumgartner JV, Braun DP, Powell J (1998) A spatial assessment of hydrologic alteration within a river network. Regul Rivers Res Manag 14:329–340. https://doi.org/10.1002/(SICI)1099-1646(199807/08)14:4<329::AID-RRR505>3.0.CO;2-E
Roberto MC, Santana NN, Thomaz SM (2009) Limnology in the Upper Paraná River floodplain: large-scale spatial and temporal patterns, and the influence of reservoirs. Braz J Biol 69:717–725. https://doi.org/10.1590/S1519-69842009000300025
Rocha PC (2010) Indicadores de alteração hidrológica no Alto Rio Paraná: intervenções humanas e implicações na dinâmica do ambiente fluvial. Soc Nat 22:191–211. https://doi.org/10.1590/S1982-45132010000100014
Rocha RRA, Thomaz SM (2004) Variação temporal de fatores limnológicos em ambientes da planície de inundação do alto rio Paraná (PR/MS—Brasil). Acta Sci Biol Sci 26:261–271. https://doi.org/10.4025/actascibiolsci.v26i3.1538
Sampaio EV, Rocha O, Matsumura-Tundisi T, Tundisi JG (2002) Composition and abundance of zooplankton in the limnetic zone of seven reservoirs of the Paranapanema River, Brazil. Braz J Biol 62:525–545. https://doi.org/10.1590/S1519-69842002000300018
Sanches PV, Nakatani K, Bialetzki A et al (2006) Flow regulation by dams affecting ichthyoplankton: the case of the Porto Primavera Dam, Paraná River, Brazil. River Res Appl 22:555–565. https://doi.org/10.1002/rra.922
Santos NCLdos, de Santana HS, Dias RM et al (2016) Distribution of benthic macroinvertebrates in a tropical reservoir cascade. Hydrobiologia 765:265–275. https://doi.org/10.1007/s10750-015-2419-6
Sousa WTZ, Thomaz SM, Murphy KJ (2010) Response of native Egeria najas Planch. and invasive Hydrilla verticillata (L.f.) Royle to altered hydroecological regime in a subtropical river. Aquat Bot 92:40–48. https://doi.org/10.1016/j.aquabot.2009.10.002
Souza Filho EE (2009) Evaluation of the Upper Paraná River discharge controlled by reservoirs. Braz J Biol 69:707–716
Souza Filho EE, Rocha PC, Comunello E, Stevaux JC (2004) Effects of the Porto Primavera Dam on physical environment of the downstream floodplain. In: Thomaz SM, Agostinho AA, Hahn NS (eds) The Upper Parana River and its Floodplain: physical aspects, ecology and conservation. Backhuys Publishers, Leiden, pp 55–74
Stevaux JC, Takeda AM (2002) Geomorphological processes related to density and variety of zoobenthic community of the upper Paraná River, Brazil. Zeitschrift Fuer Geomorphol 129:143–158
Stevaux JC, Martins DP, Meurer M (2009) Changes in a large regulated tropical river: the Paraná River downstream from the Porto Primavera Dam, Brazil. Geomorphology 113:230–238. https://doi.org/10.1016/j.geomorph.2009.03.015
Stockner JG, Rydin E, Hyenstrand P (2000) Cultural oligotrophication: causes and consequences for fisheries resources. Fisheries 25:7–14. https://doi.org/10.1577/1548-8446(2000)025<0007
Takeda AM, Fujita DS (2004) Benthic invertebrates. In: Thomaz SM, Agostinho AA, Hahn NS (eds) The Upper Parana River and its Floodplain: physical aspects, ecology and conservation. Backhuys Publishers, Leiden, pp 191–208
Thomaz SM, Pagioro TA, Bini LM et al (2004) Limnological characterization of the aquatic environments and the influence of hydrometric levels. In: Thomaz SM, Agostinho AA, Hahn NS (eds) The Upper Parana River and its Floodplain: physical aspects, ecology and conservation. Backhuys Publishers, Leiden, pp 75–102
Thomaz SM, Bini LM, Bozelli RL (2007) Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 579:1–13. https://doi.org/10.1007/s10750-006-0285-y
TNC: The Nature Conservancy (2009) Indicators of Hydrologic Alteration Version 7.1 User’s Manual. https://www.conservationgateway.org/ConservationPractices/Freshwater/EnvironmentalFlows/MethodsandTools/IndicatorsofHydrologicAlteration/Documents/IHAV7.pdf. Accessed 11 May 2017
Turesson H, Brönmark C (2007) Predator-prey encounter rates in freshwater piscivores: effects of prey density and water transparency. Oecologia 153:281–290. https://doi.org/10.1007/s00442-007-0728-9
Vannote RL, Minshall GW, Cummins KW et al (1980) The river continuum concept. Can J Fish Aquat Sci 37:130–137. https://doi.org/10.1139/f80-017
Vazzoler AEA de M (1996) Biologia da reprodução de peixes teleósteos: teoria e prática. EDUEM, Maringá
Verba JT, Lampert VR, Azevedo M (2011) Reproductive traits of Gymnogeophagus labiatus (Teleostei, Cichlidae), in an upper stretch of Sinos river, Caraá, Brazil. Iheringia Ser Zool 101:200–206. https://doi.org/10.1590/S0073-47212011000200007
Ward JV, Stanford JA (1983) Serial Discontinuity Concept of lotic ecosystems. In: Fontaine D, Bartell S (eds) Dynamics of Lotic Systems. Science Publishers, Ann Arbor, pp 29–42
Ward JV, Stanford JA (1995a) Ecological connectivity in alluvial river ecosystems and its disruption by flow regulation. Regul Rivers Res Manag II:105–119
Ward JV, Stanford J (1995b) The serial discontinuity concept: extending the model to floodplain rivers. Regul Rivers Res Manag 10:159–168
Winemiller KO (1989) Patterns of variation in life history among South American fishes in seasonal environments. Oecologia 81:225–241. https://doi.org/10.1007/BF00379810
Winemiller KO, McIntyre PB, Castello L et al (2016) Balancing hydropower and biodiversity in the Amazon, Congo, and Mekong. Science 351:128–129. https://doi.org/10.1126/science.aac7082
Wohl E, Bledsoe BP, Jacobson RB et al (2015) The natural sediment regime in rivers: broadening the foundation for ecosystem management. Bioscience 65:358–371. https://doi.org/10.1093/biosci/biv002
Acknowledgements
We are grateful to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the financial support through the Long Term Ecological Research Program (PELD—site 6). We thank the Programa de Excelência Acadêmica (PROEX) and the Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (NUPELIA) for providing the infrastructure for research. David Schumann, Igor de Paiva Affonso and Norma Segatti Hahn provided a constructive review to the initial manuscript. We also thank João D. Latini for helping with datasets and Jaime L. L. Pereira for helping with figures. RVG is thankful to CNPq for the graduate student scholarship and to the Department of Wildlife, Fisheries and Aquaculture at Mississippi State University for additional financial support. The Mississippi Cooperative Fish and Wildlife Research Unit is a cooperative effort of the Mississippi Department of Wildlife, Fisheries and Parks; Mississippi State University; and US Geological Survey. Any use of trade, firm or product names is for descriptive purposes only and does not imply endorsement by the US Government.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Granzotti, R.V., Miranda, L.E., Agostinho, A.A. et al. Downstream impacts of dams: shifts in benthic invertivorous fish assemblages. Aquat Sci 80, 28 (2018). https://doi.org/10.1007/s00027-018-0579-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00027-018-0579-y