
Abstract
Insect–plant interactions are complex and dynamic relationships that have evolved over millions of years. Plants have developed various adaptations to deter insect herbivores, including physical, chemical, and induced defences. In response, insects have evolved detoxification mechanisms, behavioural adaptations, and physiological adaptations to overcome these defences. This coevolutionary arms race has shaped the interactions between plants and insects, leading to a diverse array of strategies and counter-strategies. Additionally, other associated organisms such as endosymbionts and rhizosphere microbes have been shown to play a critical role in these interactions. Endosymbionts can alter the nutritional quality of plant tissue and confer resistance to environmental stressors, while rhizosphere microbes can influence plant growth and nutrient uptake. Understanding the coevolutionary arms race and the role of associated organisms in insect–plant interactions has important implications for plant protection and management. By leveraging these relationships, we can develop sustainable and eco-friendly approaches to crop protection and pest management.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
2.1 Introduction
2.1.1 Overview
The term ‘Coevolution’ was introduced by Ehrlich and Raven (1964) to describe the close interaction between two or more species in an ecosystem, resulting in genetic adaptations in one or both species due to natural selection imposed by the other interacting species. This process is a product of evolution and can have reciprocal impacts (Janzen 1980; Berenbaum and Zangerl 1998; Woolhouse et al. 2002). The interaction between any groups of interacting species, including mutualists, pathogens, competitors, and predators and prey, can lead to coevolution. The nature and intensity of their interaction determine the selection pressures exerted by each species on the other. Previous research has demonstrated the significance of the relationship between two species in determining the selection pressures on each other (Hochberg et al. 2000; Thompson and Cunningham 2002).
Interaction between plants and insects is a constantly changing system. These interactions have a long history of coevolution, with each group engaged in an ongoing arms race, thus influencing the other’s evolutionary trajectory (Labandeira 1998). The theory of coevolution developed by Ehrlich and Raven (1964) forms the foundation for the current understanding of the dynamic interplay between plants and insects. The relationship between insects and plants has been dynamically occurring for over 400 million years, resulting in a complex system of interactions that includes herbivory, pollination, and other mutualistic relationships (Labandeira 2013). Insects perform essential functions for plants, such as defence and pollination, while plants provide critical resources such as shelter, oviposition sites, and food for insect growth and reproduction. Nonetheless, herbivores can pose a significant threat to plants and exert strong selection pressure to evolve strategies to tolerate or resist them (Panda and Khush 1995).
The coevolution of insect–plant interactions has resulted in a complex system of adaptations and counter-adaptations, with each group influencing the other’s evolutionary trajectory. This ongoing process has led to the diversity and complexity of the natural world we see today, with insects and plants playing critical roles in ecosystem function and biodiversity.
2.1.2 Coevolution in Shaping Various Insect–Plant Interactions
One of the most prominent examples of coevolution between insects and plants is the evolution of morphological adaptations and specialized feeding strategies. Many insects have evolved to feed on specific plants. For example, some insects have evolved specialized mouthparts to pierce and suck plant tissue, while others have developed chewing mouthparts to consume leaves (Krenn 2019). However, excessive herbivory on leaves, stems, flowers, and other plant parts can lead to reduced growth and reproduction, and in some cases, can even kill the plant. Accordingly, plants have evolved specific mechanisms to deter or tolerate herbivory. They have evolved various structures such as thorns, spines, tough leaves, and other physical barriers to deter herbivores (War et al. 2012). Some plants also produce/increase toxic chemicals that are harmful to herbivores, in response to which insects have developed mechanisms to detoxify these chemicals (Gatehouse 2002). This arms race between plants and insects has resulted in evolution of a vast array of chemical compounds and biochemical pathways (Schoonhoven et al. 2005; Howe and Jander 2008). Plants also recruit other organisms such as predators or parasitoids to attack the herbivores.
The coevolution of insect–plant interactions has also resulted in various mutualistic relationships (Bronstein and Huxman 1997). Many plants rely on insects for pollination, and in return, provide them with a source of nectar or pollen. This mutualistic relationship has led to the coevolution of floral morphology and insect behaviour. Plants have evolved to form floral structures, shapes and colours (Willmer 2011), and chemical signals (Raguso 2008) to attract specific pollinators. Accordingly, insect pollinators perceive and interpret these signals by using their visual and olfactory sense organs (Chittka and Raine 2006). These insects feed on the nectar produced by plants (Heil 2011) and aid in pollination by developing specialized adaptations for collecting and transferring pollen, such as long tongues or hairy bodies (Johnson and Steiner 2000).
Coevolutionary mutualistic relationships between insects and plants can extend beyond just pollination. In some cases, insects live in close association with the plant and providing the plant with protection from predators in exchange for nutrients (Bronstein et al. 2006). This type of relationship has led to the evolution of specialized structures in plants to accommodate the insects.
One such structure is called a gall, which is a growth on the plant caused by the interaction between the plant and an insect, mite, or other arthropod. Gall-forming insects, such as wasps and flies, lay their eggs in plant tissues, which induces the plant to form a protective structure called a gall around the developing insect. The gall provides a safe environment for the insect to develop and feed, and in exchange, the insect secretes chemicals that alter the physiology of the plant to make it more suitable for its needs (Stone and Schönrogge 2003; Jansen-González et al. 2012). Another specialized structure is the domatia, which are small cavities or pouches that are formed in the leaves or stems of some plants, and are inhabited by mutualistic arthropods such as mites, spiders or insects, which provide plant with protection (Agrawal and Karban 1997). Similarly, many ant species depend on plants for food and housing in exchange for protection against predators (Heil and McKey 2003; Nelson et al. 2019).
Furthermore, the coevolution of insect–plant interactions has also driven the evolution of mimicry. Some insects have evolved to mimic the physical appearance or chemical signals of other insects to avoid being detected by predators or to attract prey. In turn, some plants have evolved to mimic the physical appearance or scent of other plants to deceive herbivores or attract specific pollinators (Schaefer and Ruxton 2009).
These types of plant–insect relationships have likely evolved through a process of coevolution, where the plants and insects have adapted to each other’s presence over time. As with pollination, the specific details of these relationships can vary widely depending on the species involved. However, in this review, we will focus on the coevolutionary dynamics of herbivore insect–plant relationship.
2.2 Coevolutionary Arms Race during Insect Herbivory
The concept of coevolutionary arms race describes the evolutionary adaptations that occur between two species, where each species is under selective pressure to evolve in response to the adaptations of the other. In the context of plant–insect herbivore interactions, this refers to the adaptations that occur between plants and the insect herbivores that feed on them (Ehrlich and Raven 1964).
Insect herbivory has been one of the major driving forces in the evolution of plant defence mechanisms. Plants have developed a variety of physical and chemical defences to deter herbivores, including thorns, spines, tough fibrous leaves and toxic secondary metabolites, such as alkaloids and terpenoids (Hanley et al. 2007; Wöll et al. 2013). Moreover, depending on the intensity and kind of an insect damage, plants modify the synthesis and distribution of these defence chemicals. Insects, in turn, have evolved mechanisms to overcome these defences, such as specialized mouthparts for feeding, specific digestive enzymes, detoxification mechanisms, and behavioural adaptations (War et al. 2018). This dynamic relationship between insects and plants has driven the coevolution of traits in both groups, resulting in a diverse array of interactions (Sharma et al. 2021).
The interaction between Asclepias syriaca (milkweed) and Danaus plexippus (monarch butterfly) is a typical example involving coevolution through toxic cardenolides. Milkweed plants produce toxic cardenolides that deter most insect herbivores, while monarch butterflies have evolved to store these toxins in their bodies, making them unpalatable to predators. As a response, milkweed plants have evolved to produce more complex mixtures of cardenolides, making it harder for monarch butterflies to sequester them (Agrawal et al. 2012).
Thus, many herbivores have evolved the ability to break down and detoxify plant toxins. In response, plants have evolved more complex and diverse chemical defences to counteract these adaptations. In some cases, plants have even evolved mutualistic relationships with other organisms, such as parasitoids, ants or fungi, which provide additional protection against herbivores (Ali and Agrawal 2012; Mathur et al. 2013a).
2.3 Plant Adaptations to Insect Herbivory
The coevolution of insects and plants results in plants acquiring adaptations that improve their own chances of surviving and reproducing in an environment with multiple kinds of herbivory. Plant defence against insect herbivory can be broadly classified into two genetic strategies, namely resistance and tolerance mechanisms. These strategies enable plants to protect themselves from damage caused by herbivorous insects through the production of chemical or physical barriers, as well as by regenerating lost or damaged tissues (Agrawal 2000; Stowe et al. 2001).
Resistance mechanisms involve the production of compounds that deter or harm herbivores, such as toxic chemicals, physical barriers or structural defences. These compounds may be constitutive, meaning they are present in the plant at all times, or induced, meaning they are produced in response to herbivory (Karban and Myers 1989). Plants have evolved various physical features, such as resins, wax, silica and lignins, which serve as direct defence mechanisms against herbivorous insects. These morphological traits, together with secondary metabolites, constitute a crucial aspect of the plant–insect interaction (Hanley et al. 2007; Belete 2018).
Tolerance mechanisms involve the ability of plants to recover from herbivory by regenerating damaged tissues, reallocating resources to compensate for lost tissues, or by increasing photosynthesis to produce more energy. Tolerant plants can often withstand higher levels of herbivory without experiencing significant reductions in growth or reproduction (Strauss and Agrawal 1999). For example, some plants may increase the growth rate of undamaged leaves after herbivory to compensate for the loss of damaged leaves (Garcia and Eubanks 2019).
Plants may use a combination of resistance and tolerance mechanisms to defend themselves against insect herbivores. The specific mechanisms used by a plant depend on factors such as the type of herbivore, the intensity of herbivory, and the availability of resources for growth and repair.
In addition, plants employ indirect defence systems such as the production of extrafloral nectaries and volatile chemicals to attract predators and other enemies of herbivores to assist in their defence (Heil and Karban 2010). Over time, the diversity and complexity of plant secondary metabolites have increased, placing increased adaptive pressure on herbivores.
2.3.1 Morphological Features
Plants possess a cuticle covered by epicuticular waxes which form films and crystals (Koch et al. 2004). These waxes provide protection against desiccation and pathogens, as well as increasing the slipperiness of the cuticle, discouraging non-specialized insects from populating leaf surfaces (Muller et al. 2007). The biosynthesis and composition of these waxes vary during plant development, and their physical–chemical properties respond to changes in temperature and season (Howe and Schaller 2008). Even changes in the wax composition due to egg deposition were found to increase fatty acid tetratriacontanoic acid (C34) and a decrease tetracosanoic acid (C24), thereby attracting the egg parasitoid (Blenn et al. 2012).
Moreover, plants have various structures such as thorns and spines that protect them mainly from mammals, and hairs called trichomes that protect against insects (Karban and Myers 1989). When trichomes are removed, herbivorous insects feed and grow more easily. Insect feeding has been observed to cause an increase in trichome density (Fordyce and Agrawal 2001; Mathur et al. 2011).
Scientific evidence suggests that the toughness of leaves can prevent the penetration by insects with piercing-sucking mouthparts, while increasing the wear and tear on the mandibles of herbivores with biting-chewing mouthparts (Raupp et al. 2008). While younger leaves may contain higher levels of chemical defences, mature leaves can be strengthened with various macromolecules such as lignin, cellulose, suberin, and callose, as well as small organic molecules like phenolics and even inorganic silica particles (Schoonhoven et al. 2005). When roots are consumed by insect herbivores, they exhibit significant regrowth in both density and quantity. Furthermore, genotypes with long and fine roots are less susceptible to herbivory than those with short and thick roots (Belete 2018).
Many plant species have laticifers and resin ducts in their vascular tissues that store latex and resins under internal pressure. When the channels are broken, the substances are secreted and can entrap or intoxicate the herbivore (Pickard 2008). Laticifers are found in over 10% of angiosperms and are most common in tropical regions (Agrawal and Konno 2009). Among more than 50 plant families that are known to produce latex, Asclepias, commonly referred to as milkweed, stands out as one of the most extensively studied plant groups (Dussourd and Hoyle 2000; Buttery and Boatman 1976). Specialist herbivores that feed on latex-producing plants can block the flow of latex by cutting veins or trenches in the leaves (Carroll and Hoffman 1980). Conifers produce oleoresins which are mixture of terpenoids and phenolics stored in high pressurized intercellular spaces called ducts (Phillips and Croteau 1999). According to scientific studies, when herbivores cause damage to a tree, the flow of resin can remove stem-boring bark beetles from the borehole (Mumm and Hilker 2006). The resin acids contain highly volatile monoterpenes and sesquiterpenes, which can deter insects while the wound is healing. However, specialist insects have evolved to overcome this defence mechanism by cutting across resin ducts, and some even use the resin as an olfactory cue to locate and select their preferred host plant (Raffa et al. 2016).
2.3.2 Chemical Defences
Plants synthesize a variety of chemical compounds that are categorized into primary and secondary metabolites based on their biological roles. Primary metabolites are essential for basic life processes such as growth, development, and reproduction. Conversely, secondary metabolites, also known as bioactive compounds, serve several ecological functions such as defence against herbivores and microbial pathogens, attraction of pollinators and seed-dispersing animals, and facilitation of competitive and symbiotic interactions with other plants and microbes (Jain et al. 2019; Salam et al. 2023). These chemical compounds have evolved to specifically target the unique biological systems of herbivores, such as their nervous, digestive, and endocrine organs, and can be produced either constitutively or upon induction (Senthil-Nathan 2013; Karban and Myers 1989). Additionally, they contribute significantly to the sensory properties of plants, including their odours, tastes, and colours. Generally, bioactive specialized compounds act as repellents for generalist insects while attracting specialist insects. Toxic compounds are more likely to intoxicate generalist herbivores, while specialists will need to allocate resources to detoxify them, thereby slowing down their growth and development (Kessler and Baldwin 2002; Macel 2011).
More than 100,000 plant secondary compounds, such as phenolics, terpenoids, alkaloids, cyanogenic glucosides, and glucosinolates have been identified (Hadacek 2002; Howe and Schaller 2008), with each plant species produces only a small, but unique combination of these compounds. While many secondary metabolites are found in multiple plant families, some compounds are specific to certain plant families or genera. A good example of this is the Solanaceae family, which contains alkaloids such as nicotine and solanine that are not commonly found in other plant families (Fiesel et al. 2022). Similarly, the Brassicaceae family contains glucosinolates such as sinigrin and brassicanapin that are not typically found in other plant families (Nguyen et al. 2020). The presence of unique secondary metabolites in different plant families can be attributed to their evolution and adaptation to specific environmental pressures and ecological niches. For instance, certain plant families may have evolved unique secondary metabolites to defend themselves against specific herbivores or pathogens that are prevalent in their native habitats.
2.3.2.1 Alkaloids
Alkaloids are bioactive natural products that are widely distributed and can be found in over 15,000 different plants. They are primarily found in Leguminosae, Liliaceae, Solanaceae, and Amaryllidaceae and have evolved as a defence against insect herbivory (Wink 2020). They are characterized by their alkaline properties and nitrogen-containing heterocyclic rings, which are synthesized from amino acids in the roots and then accumulated above ground. The heterocyclic ring structure includes pyridines, pyrroles, indoles, pyrrolidines, isoquinolines, and piperidines. Some alkaloids, such as caffeine and solanidine, are alkaline but not derived from amino acids, while others like mescaline are alkaline and derived from amino acids but do not contain nitrogen in a heterocycle (Thawabteh et al. 2019; Phukan et al. 2023). Plants containing demissine and solanine alkaloids have been found to be resistant to certain herbivores, although some can detoxify them. Pyrrolizidine alkaloids (PAs) occur naturally in many plants as non-toxic N-oxides but become toxic when reduced in the digestive tracts of certain insect herbivores. Some herbivores, such as Utetheisa ornatrix, can detoxify PAs and use them as a defence against their own predators (Wink 2019; Bezzerides et al. 2004).
2.3.2.2 Cyanogenic Glucosides
Cyanogenic glucosides (CNglcs) are present in various plant species, including angiosperms, monocotyledons, dicotyledons, gymnosperms, and pteridophytes, with more than 2600 species from over 550 genera and 150 families (Yulvianti and Zidorn 2021). These amino acid derived glucosides come from tyrosine, valine, isoleucine, and phenylalanine and are stored in vacuoles (Gleadow and Møller 2014). Damage of plant tissue due to insect herbivory causes exposure to β-glucosidases leading to hydrolysis and the formation of toxic hydrogen cyanide (HCN) and a ketone or aldehyde (Boter and Diaz 2023). HCN is a potent toxin that can disrupt cellular respiration by inhibiting cytochrome c oxidase in the mitochondrial respiratory pathway. This can result in severe health complications and even death in high enough concentrations (Manoj et al. 2020). In addition to their toxic properties, CNglcs can also serve as nitrogen storage compounds or osmoprotectants, and their presence may increase a plant’s resistance against certain herbivores while acting as phagostimulants or oviposition cues for others. However, the production of CNglcs comes at a cost, as it can be energetically expensive and may hinder plant growth and development. Additionally, the release of HCN during CNglc production can harm the plant itself by inhibiting the production of phytoalexins, which are essential for defending against microbial pathogens (Vetter 2000).
2.3.2.3 Glucosinolates
Glucosinolates (GLS) are compounds containing sulphur and nitrogen that are found in Brassicaceae and Capparales. They are derived from amino acids, and there are over 200 different known structures (Ishida et al. 2014; Rosa-Téllez 2023). They are categorized into four groups based on their amino acid precursor of the side chain: aliphatic glucosinolates (50%) derived from methionine, indole glucosinolates (10%) synthesized from tryptophan, aromatic glucosinolates (10%) derived from phenylalanine or tyrosine, and structures synthesized from several different amino acids (30%) or with unknown biosynthetic origin (Hopkins et al. 2009). These compounds are more abundant in roots than in shoots, and different tissues have different dominant glucosinolates (Van Dam et al. 2009). When they are hydrolysed by myrosinases upon tissue disruption, they break down into toxic breakdown products such as isothiocyanates, nitriles, and thiocyanates, which are responsible for the flavours of several vegetable foods (Eisenschmidt-Bönn et al. 2019; Wieczorek et al. 2018). These breakdown products act as both herbivore toxins and feeding repellents (Jeschke et al. 2016). Different insects respond to glucosinolates differently (Mathur et al. 2013b). The metabolic diversity in toxin production by individual plants can also provide defence against herbivores with different feeding strategies or resistance mechanisms (Speed et al. 2015; Wittstock et al. 2003).
2.3.2.4 Phenolics
Phenolics of several classes are synthesized by a wide variety of plants. This group comprises approximately 10,000 distinct chemical entities such as tannins, vanillin, ferulic acid, and caffeic acid (Alamgir and Alamgir 2018; de la Rosa et al. 2019). They can be synthesized using either the malonic acid pathway or the Shikimic acid pathway (Saltveit 2017). Phenols serve numerous functions for plants, such as herbivore defence, pollinator attraction, mechanical support, systemic acquired resistance (SAR), and allelopathy (Divekar et al. 2022). There are multiple examples of phenolic compounds acting as defence mechanisms against insects. They can act as feeding deterrents by reducing the palatability and digestibility of plant tissues or directly inhibit insect growth and development or even cause mortality. Additionally, some phenolics attract natural enemies of insect herbivores, such as parasitoids or predators, which can help to reduce insect populations (Rehman et al. 2012).
2.3.2.5 Terpenoids
Terpenoids are a class of organic compounds synthesized in plants from either acetyl-CoA or glycolytic intermediates. They are classified based on the number of isoprene units they contain, ranging from monoterpenes with ten carbons to polyterpenes with more than 40 carbons (Ninkuu et al. 2021). Terpenes play various roles in plants, including defence against herbivores and pathogens, attraction of pollinators, response to abiotic stress, synthesis of plant hormones, and release of volatile organic compounds (Abbas et al. 2017). These functions contribute to the adaptation and survival of plants in different environments (Aharoni et al. 2005). Essential oils, a blend of volatile monoterpenes and sesquiterpenes with well-known insecticidal properties, are found in many plants. Leafcutter ants are repelled by the terpenoid limonene, which citrus trees generate (Fürstenberg-Hägg et al. 2013). Coniferous plants generate monoterpenes, which are poisonous to many insects, including bark beetles. While certain terpenoid amide derivatives can function as insect juvenile hormone analogs, phytoecdysones, which are steroids found in common fern, prevent insect moulting by imitating moulting hormones (Canals et al. 2005). Additionally, when combined, several terpenoids have synergistic effects that have a higher impact on insect fatality rates.
Many secondary metabolites are constitutively present in a certain amount irrespective of presence of herbivorous insects or other stressors. However, upon herbivore attack, plants may recognize elicitor molecules, which are then transduced into the cell to activate specific genes for the biosynthesis of diverse defensive chemicals with unique chemical activities. This process is mediated by complex cellular transduction mechanisms that regulate the genetic expression of biochemical pathways (Bonaventure et al. 2011; Maffei 2010; Maffei et al. 2012; Mithoefer and Boland 2012). These routes can diversify since they are not required for growth and development (Hartmann 2007). The diversity of metabolic products in plants can be attributed to several molecular processes such as gene and genome duplications, accumulation of point mutations, and multi-loci control. These mechanisms result in variations in gene expression, enzyme activity, and substrate specificity, leading to the production of diverse secondary metabolites (Kroymann 2011; Weng et al. 2012; Moore et al. 2014).
2.4 Insect Adaptations to Plant Defences
Insect herbivores have evolved various adaptations to cope with the complex challenges posed by their plant hosts, including the timing of plant growth and reproduction, the specific nutrient composition, and the chemical and physical defences of the plant. As a consequence of this coevolution, herbivorous insects have developed specialized feeding behaviours and are often restricted to a narrow range of host plants that are related phylogenetically and/or share similar biochemical compositions. This adaptation enables herbivorous insects to efficiently feed on their host plants, while minimizing the risks of consuming toxic or unsuitable plant material (Simon et al. 2015; War et al. 2018).
In response to selective pressure from plants that produce direct defence compounds, specialist herbivores have evolved various defence mechanisms to mitigate the toxicity of these compounds. Unlike generalist herbivores, they have developed the ability to detoxify the hazardous chemicals through enzymatic inactivation or sequestration. This adaptation allows them to tolerate the ingestion of plant material that would otherwise be toxic to them (Nishida 2002; Peng et al. 2007; Petschenka and Agrawal 2016). Additionally, specialized herbivores can use certain compounds, such as phagostimulants, to locate suitable host plants (del Campo et al. 2001; Picaud et al. 2003). Some herbivores can even store these protective compounds and use them for defence against predators and parasites or to attract mates (Cogni et al. 2012). This pattern of specialization highlights the strong evolutionary relationship between host plants and specialized herbivores and has been observed in numerous studies (Becerra 2007; Bandeili and Müller 2010; Richards et al. 2010; Agrawal et al. 2012).
2.4.1 Behavioural Adaptations
Insects have the ability to avoid consuming poisonous plants by utilizing either genetically predetermined or learned avoidance mechanisms that are triggered through visual, olfactory, or tactile exposure (Chapman 2003). Female insects possess genetically programmed oviposition behaviour that can prevent them from laying eggs on unsuitable plants (Fox et al. 2004). However, in some cases, larvae may need to migrate to locate a suitable host plant (Cotter and Edwards 2006). Furthermore, insects may consume non-toxic parts of plants or seek out environments that are free from toxins, as well as utilizing plants that are not currently producing toxins (Nealis and Nault 2005). Insects can exhibit diverse responses to toxins depending on their surrounding environmental conditions. For instance, solitary phase locusts are deterred by a bitter-tasting plant alkaloid, whereas gregarious-phase locusts are attracted to it. This difference in feeding behaviour is reflective of various anti-predator strategies, such as changes in colouring and grouping tendencies (Glendinning et al. 2002; Despland 2021; Després et al. 2007).
Insects utilize chemosensation as a means of detecting and avoiding potentially harmful secondary compounds present in their environment. Gustatory and olfactory receptor neurons allow insects to detect these poisons through their senses of taste, smell, and touch. Gustatory receptors are categorized into sweet, bitter, umami, salt, and carbon dioxide receptors, and bitter receptors are involved in sensing secondary compounds that inhibit insect intake by activating downstream signaling pathways. This aversive mechanism may be inherited or acquired (Zunjarrao et al. 2020). Some insects limit their diet to organs of non-toxic plants or eat only on plants with low levels of toxins to avoid hazardous substances (Fox et al. 2004). However, insects also avoid bitter substances even though they are not always harmful, which limits their host plant options and increases the cost of avoidance (Pelden and Meesawat 2019). To circumvent plant defences, insects have developed various strategies, such as recognizing previously induced reactions, feeding on diverse plant tissues, weaving silk over spines, making trenches across leaves before eating, and puncturing leaf veins to block the passage of poisons (Perkins et al. 2013).
2.4.2 Manipulation of Plant Chemical Defences
While avoiding plant allelochemicals is generally the most effective way for insects to minimize their exposure to harmful compounds, they have evolved additional mechanisms to cope with occasional consumption of toxic substances.
Many insects can affect a plant’s chemical defences by making tunnels in leaves to release pressure in secretory canals and remove poisonous exudates near their feeding spot (Helmus and Dussourd 2005; Becerra 2003). Insects have developed the ability to inhibit the plant’s defence mechanisms by secreting elicitors that can lower the expression of regulatory genes that are crucial for initiating the plant’s defences. This allows the insects to better exploit the plant and continue feeding on it without triggering a defensive response (Bede et al. 2006; Divekar et al. 2022). Gall-inducing insects, such as sawflies, have developed the ability to reduce the levels of toxic phenolic chemicals found in the galls where their larvae grow. This adaptation is thought to facilitate frequent switching between host plants, which can increase the survival and reproductive success of these insects (Nyman and Julkunen-Tiitto 2000). By reducing the levels of toxic chemicals in their feeding sites, gall-inducing insects can better tolerate a wider range of plant species and exploit new host plants as they become available, ultimately enhancing their ecological flexibility and evolutionary potential.
2.4.3 Sequestration of Plant Defence Compounds by Insects
Sequestration is a common defence strategy employed by insects that can have significant impacts on their interactions with plants. This strategy involves the selective absorption, transport, endogenous metabolism, and excretion of plant chemicals into different organs of the insect. Through this process, insects can accumulate and store large quantities of plant compounds, often in specialized tissues, for use in a variety of contexts, such as chemical defence against predators or as a source of nutrients. The sequestration of plant chemicals can also lead to coevolutionary dynamics between insects and plants, as insects may evolve mechanisms to overcome or exploit plant defences, while plants may evolve counter-strategies to defend against insect herbivory (Pentzold et al. 2014; Nishida 2002). Insects are known to sequester a diverse array of compounds from their host plants, including aromatic compounds, nitrogen-containing secondary compounds such as alkaloids, cyanogenic glycosides, glucosinolates, and isoprenoids such as cardiac glycosides, cucurbitacins and iridoid glycosides (Opitz and Müller 2009). These sequestered compounds can serve various functions, including as defensive agents against predators or parasites, as feeding stimulants or repellents, and as precursors for the biosynthesis of sex pheromones or other signaling molecules (Dobler 2001). The specific suite of sequestered compounds can vary widely among insect taxa, reflecting differences in the evolutionary history, feeding behaviour, and ecological pressures faced by each group.
Studies have revealed numerous mechanisms by which insects can acquire and store plant chemicals, including a shift from de novo synthesis of detoxifying compounds to sequestration of host phytochemicals. Sequestration is widespread in several insect groups because it is thought to have a lower metabolic cost than de novo production (Beran and Petschenka 2022). In spite of this, there has been limited experimental research on it since the evolutionary background and natural history of the study system aren’t well known.
Insects that sequester defence compounds from plants have evolved diverse mechanisms to accumulate these compounds in various parts of their bodies, such as the haemolymph, defence glands, or integument. Insects absorb plant defence chemicals from the gut lumen into the hemocoel through the peritrophic matrix and gut epithelium. The type of absorption mechanism depends on the chemical properties of the substances, with absorption occurring either through passive diffusion or carrier-mediated transport. The permeability of the epithelial layer also plays a role in absorption and may be modulated by efflux pumps that restrict the uptake of certain substances (Dobler et al. 2015; Sorensen and Dearing 2006). This selective accumulation is achieved through processes such as selective absorption through the gut, transport within the body, endogenous metabolism, and excretion via the malpighian tubules (Petschenka and Agrawal 2016). The cardiac glycoside digitoxin is one example. It was discovered to passively diffuse over the midgut of the milkweed insect Oncopeltus fasciatus, but not across the midguts of the desert locust Schistocerca gregaria and the American cockroach, Periplaneta americana (Scudder and Meredith 1982). This suggests that mechanisms that either aid or hinder the absorption of plant defence compounds across the gut are a key factor in the development of sequestration. It was demonstrated that the cytochrome P450 monooxygenases, a different group of membrane proteins, are implicated in nicotine transportation from the gut into the haemolymph of the tobacco hornworm, Manduca sexta (Kumar et al. 2014).
In addition, insects may modify the amounts and composition of sequestered defence chemicals based on the chemical composition of their food plant. For example, some insects can selectively sequester certain types of glucosinolates or iridoid glycosides depending on the plant species they consume (Baden et al. 2013; Beran et al. 2014; Yang et al. 2020). Nevertheless, because the processes of sequestration have only been studied in a small number of insect species, the significance of each of these aspects is not entirely known.
2.4.4 Detoxification
Insects have developed sophisticated detoxification mechanisms to biotransform and excrete secondary metabolites, which have the potential to cause toxicological harm (Erb and Robert 2016). This enables insects to minimize the deleterious effects of these compounds and maintain their ability to feed on plants that contain secondary metabolites. Some insects have specialized gut enzymes or gut microorganisms that detoxify ingested plant chemicals, allowing them to consume otherwise toxic plant material (van den Bosch and Welte 2017).
Insects possess a variety of enzymes that aid in detoxifying plant toxins through oxidation, reduction, hydrolysis, or conjugation of molecules (Birnbaum and Abbot 2018). The three primary types of enzymes involved in detoxification are cytochrome P450 monooxygenases (also known as CYPs or P450s), glutathione S-transferases (GSTs), and carboxylesterases (COEs) (Feyereisen 2005). Berenbaum and Zangerl (1998) demonstrated that the ability of insects to metabolize furanocoumarins, a class of compounds produced by plants, was found to be associated with the varying levels of furanocoumarin production among different plant species. This suggests a tight genetic linkage between the genes responsible for the insect’s capacity to utilize host plants and those involved in metabolizing these chemical compounds. The cytochrome P450-dependent monooxygenase gene is a notable example of a gene implicated in both plant–insect interactions and insect physiology. Insects, such as Papilio polyxenes, have developed adaptations to thrive on host plants that contain toxins by undergoing diversification of P450 enzymes, which play a key role in the detoxification process (Scott and Wen 2001).
2.4.5 Further Processing of Plant Toxins
A multitude of insects have developed the ability to eliminate or excrete detrimental plant compounds, and in some instances, these compounds are sequestered and utilized for diverse purposes, such as synthesizing pheromones, defending against infections or predators, and generating pigments for adult coloration (Beran and Petschenka 2022; Robinson et al. 2023). Selective transport and storage mechanisms that keep the poison from interfering with the insect’s physiological functions are necessary for sequestration (Kuhn et al. 2004). Molecular studies on leaf beetles have shown that a minor modification in ancestral defence mechanism has facilitated the utilization of plant components for predator defence. This adaptation has emerged convergently in multiple leaf beetle lineages and is metabolically advantageous due to its low energy cost (Kuhn et al. 2004; Termonia et al. 2001).
2.5 Involvement of Higher Trophic Level
Insects that feed on plants release volatile organic compounds (VOCs) and extrafloral nectar as a signal to be found and attacked by parasitoids and other natural enemies of the insect (Heil 2008, 2015; Shah et al. 2015). Insect-caused plant damage results in the induction of volatile and nectar and extrafloral nectar secretion to entice the herbivore’s natural enemies, such as ants, wasps, and parasitoids (Heil 2011, 2015; Mathur et al. 2013a, b). These natural enemies may exert selective pressure on insect herbivores, resulting in the evolution of various traits, such as behavioural changes or morphological adaptations that enable herbivores to avoid predation (Dicke and Baldwin 2010). These changes, in turn, can influence herbivore feeding behaviour and plant defence strategies, potentially leading to coevolutionary interactions between herbivores and their host plants (Vale et al. 2018). In contrast, when predators consume herbivores that are not well adapted to the plant’s chemical defences, it can lead to the selection for herbivores that can more efficiently detoxify or sequester plant compounds. This can result in an arms race between herbivores and plants, where plants evolve more potent chemical defences, and herbivores continue to adapt their detoxification and sequestration mechanisms to overcome them. The pressure from predators can, therefore, indirectly impact the coevolutionary dynamics between herbivores and their host plants, driving the evolution of novel traits that can enhance herbivore fitness and promote coevolutionary interactions between plants and insects.
2.6 Role of Symbiotic Microorganisms
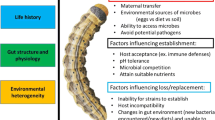
Insects and plants have a mutually beneficial relationship with their symbiotic microorganisms. This relationship creates a complex web of interactions that have both ecological and evolutionary implications. These microbes are critical components of the intricate relationships among plants, insects, and their environments. The interaction between insects and plants involves an indirect interaction with the microorganisms that inhabit each other. In other words, insects and plants have a shared relationship with the microorganisms living within them, which plays a critical role in shaping their interactions and the broader ecological and evolutionary consequences (Sharma et al. 2021; Sugio et al. 2015). The microorganisms that inhabit both plants and insects are highly diverse and can be found in various parts of their respective hosts. Endophytes, microorganisms that live within the tissues of plants, can be found in the roots, stem, leaves, seeds, and fruits of plants, while insects harbour the symbiotic microorganisms throughout their bodies (Compant et al. 2021; Provorov and Onishchuk 2018).
Endophytes are known to play a crucial role in insect–plant interactions by influencing the behaviour, fitness, and survival of their insect hosts (Shikano et al. 2017; Grunseich et al. 2019). Endophytes can produce various secondary metabolites that have insecticidal properties, which can help to deter herbivorous insects from feeding on the plant. The interdependent relationship between plants and endophytes has been demonstrated to promote plant growth and improve their physiological condition, as well as providing a vital defence mechanism against harsh environmental conditions (Malinowski et al. 2000; Peschiutta et al. 2018; Mathur and Ulanova 2022). They contribute to not only growth-promoting phytohormones such as indole acetic acid and gibberellic acids, but also various metabolic compounds such as alkaloids, terpenoids, and flavonoids (Mukherjee et al. 2022; Sharma et al. 2023; Tan and Zou 2001). Endophytes can also influence the nutritional quality of the plant for insect herbivores, by altering the levels of carbohydrates, proteins, and other nutrients in the plant tissue. This can affect the performance and development of herbivorous insects that feed on the plant. Furthermore, endophytes can also confer resistance to environmental stressors such as drought, salinity, and temperature fluctuations, which can improve the overall health and survival altering the levels of carbohydrates, proteins, and other nutrients in the plant tissue and its associated insect community (White et al. 2019). Root-associated microorganisms also contribute to the increased emission of volatiles. The rhizospheric microbiome is susceptible to changes caused by herbivory, which, in turn, can affect the relationship between the plant microbiome and the herbivore (Venturi and Keel 2016).
Insects harbour microorganisms both internally and externally that may be divided into two categories: primary and secondary symbionts. Primary symbionts are crucial for the survival and reproduction of insects and are typically obligatory and vertically transferred from mother to offspring (Bright and Bulgheresi 2010; Szklarzewicz and Michalik 2017). Secondary symbionts, on the other hand, are not essential for host survival and can reside in various insect tissues. They provide a range of benefits to their hosts, such as protection against environmental stresses, natural enemies, and plant toxins (Brownlie and Johnson 2009; van den Bosch and Welte 2017). Secondary symbionts reproduce only in association with their hosts, but they can undergo horizontal transfer. Many studies have highlighted the importance of secondary endosymbionts in insect–plant interactions, including recognition, defence, digestion, and fitness of the insect (Frago et al. 2012; Paniagua Voirol et al. 2018).
Microbial symbionts play a crucial role in insect–plant interactions in several ways. They are known to aid in the recognition of host plants by altering the sensory abilities of insects, such as their ability to detect plant volatiles. Moreover, they can provide protection against natural enemies of insects, such as predators, parasites, and pathogens, which can increase the survival and fitness of the insect. They also help insects to digest plant material and extract nutrients from it, which can be difficult to break down without the aid of microorganisms. Additionally, they confer resistance against plant toxins, allowing the insect to consume a wider range of plant species. These symbiotic microorganisms can also contribute to the overall health and fitness of the insect, which can affect its reproductive success and ability to survive in its environment (Frago et al. 2012; Frago and Zytynska 2023; Hansen and Moran 2014). Overall, microbial symbionts have a significant impact on the interactions between insects and plants, and understanding these relationships can provide insights into the ecology and evolution of both groups of organisms.
2.7 Role of Rhizosphere in Shaping Insect–Plant Interactions
Rhizosphere is a complex and dynamic system that plays a crucial role in shaping insect–plant interactions. One way in which the rhizosphere influences these interactions is by providing a habitat for beneficial microorganisms, such as bacteria and fungi, that can promote plant growth and health. These microorganisms can produce plant growth-promoting substances, such as phytohormones and enzymes, that stimulate plant growth and increase the nutritional quality of the plant for herbivorous insects (Berendsen et al. 2012). As a result, plants growing in healthy and diverse rhizospheres can be more attractive to herbivorous insects, which can lead to increased herbivory and pest pressure. However, these rhizosphere microbes can modulate insect herbivory in different manners and are powerful drivers of insect–plant coevolution (Rasmann et al. 2017; Van der Putten et al. 2001).
In contrast, studies have reported that by creating volatiles, the microbial populations found in the roots help plants survive a variety of biotic and abiotic stressors (Garbeva and Weisskopf 2020). Depending on the insect’s feeding guild and specialization, these microorganisms have varying effects on insect herbivory (Pineda et al. 2010). By boosting sugar and protein production, causing metabolic changes, and lowering root pathogen infestation, these bacteria give the plant a selective advantage during insect attacks (Westman et al. 2019). Studies have demonstrated that in order to survive without their insect prey, soil-dwelling entomopathogenic fungi like Metarhizium and Beauveria have evolved a symbiotic association with plants (Hu and St. Leger 2002). It is hypothesized that other fungi connected to plants may have provided these microorganisms with the genes for insect disease. Rhizospheric microbial communities, such as PGPR, PSB, and VAM, have been reported to boost the antioxidant activity of plants after insect damage (Song et al. 2014; Kousar et al. 2020; Sharma and Mathur 2020).
Thus, interactions between plants, insects, and the rhizosphere can provide insights into the ecology and evolution of these organisms and can help to develop sustainable strategies for managing insect pests in agricultural and natural ecosystems.
2.8 Conclusion
Over the period of 400 million years, several strategies for dealing with chemical defences of their hosts have been developed by insects that feed on plants. The response of plants to insect herbivores has significant implications not only for the interacting players themselves but also for the interactions between responding plants and other organisms associated with plants. This coevolutionary arms race between plants and insect herbivores can have significant ecological consequences. It can influence the structure and diversity of plant communities, as well as the evolution of insect herbivore species. The interactions between plants, insects, and their biotic and abiotic factors such as parasitoids and predators, endosymbionts, and the rhizosphere associated with them are shaped by coevolutionary processes, which have driven the adaptation of these organisms to each other over time. Plants and insects have co-evolved complex mechanisms of defence and counter-defence that allow them to interact in a dynamic and constantly evolving way. It can also have economic impacts, as some insect herbivores are pests that can cause significant damage to crops.
In summary, the coevolutionary arms race between plants and insect herbivores is an ongoing process of adaptation and counter-adaptation, where each species evolves in response to the adaptations of the other. This process can have significant ecological and economic implications and provides a fascinating example of the complexity of evolutionary interactions between species. Understanding the coevolutionary dynamics between plants, insects, and the rhizosphere can provide valuable insights into the ecology and evolution of these organisms and can help to develop sustainable strategies for managing insect pests in agricultural and natural ecosystems.
References
Abbas F, Ke Y, Yu R, Yue Y, Amanullah S, Jahangir MM, Fan Y (2017) Volatile terpenoids: multiple functions, biosynthesis, modulation and manipulation by genetic engineering. Planta 246:803–816
Agrawal AA (2000) Benefits and costs of induced plant defense for Lepidium virginicum (Brassicaceae). Ecology 81:1804–1813
Agrawal AA, Karban R (1997) Domatia mediate plant arthropod mutualism. Nature 387:562–563
Agrawal AA, Konno K (2009) Latex: a model for understanding mechanisms ecology and evolution of plant defense against herbivory. Annu Rev Ecol Evol Syst 40:311–331
Agrawal AA, Petschenka G, Bingham RA, Weber MG, Rasmann S (2012) Toxic cardenolides: chemical ecology and coevolution of specialized plant–herbivore interactions. New Phytol 194:28–45
Aharoni A, Jongsma MA, Bouwmeester HJ (2005) Volatile science? Metabolic engineering of terpenoids in plants. Trends Plant Sci 10(12):594–602
Alamgir ANM, Alamgir ANM (2018) Secondary metabolites: secondary metabolic products consisting of C and H; C, H, and O; N, S, and P elements; and O/N heterocycles. In: Therapeutic use of medicinal plants and their extracts: volume 2: phytochemistry and bioactive compounds, pp 165–309
Ali JG, Agrawal AA (2012) Specialist versus generalist insect herbivores and plant defense. Trends Plant Sci 17:293–302
Baden CU, Franke S, Dobler S (2013) Host dependent iridoid glycoside sequestration patterns in Cionus hortulanus. J Chem Ecol 39:1112–1114
Bandeili B, Müller C (2010) Folivory versus florivory—adaptiveness of flower feeding. Naturwissenschaften 97:79–88
Becerra JX (2003) Synchronous coadaptation in an ancient case of herbivory. Proc Natl Acad Sci 100:12804–12807
Becerra JX (2007) The impact of herbivore–plant coevolution on plant community structure. Proc Natl Acad Sci 104:7483–7488
Bede JC, Musser RO, Felton GW, Korth KL (2006) Caterpillar herbivory and salivary enzymes decrease transcript levels of Medicago truncatula genes encoding early enzymes in terpenoid biosynthesis. Plant Mol Biol 60:519–531
Belete T (2018) Defense mechanisms of plants to insect pests: from morphological to biochemical approach. Trends Tech Sci Res 2:555–584
Beran F, Petschenka G (2022) Sequestration of plant defense compounds by insects: from mechanisms to insect–plant coevolution. Annu Rev Entomol 67:163–180
Beran F, Pauchet Y, Kunert G, Reichelt M, Wielsch N, Vogel H, Heckel DG (2014) Phyllotreta striolata flea beetles use host plant defense compounds to create their own glucosinolate-myrosinase system. Proc Natl Acad Sci 111:7349–7354
Berenbaum MR, Zangerl AR (1998) Chemical phenotype matching between a plant and its insect herbivore. Proc Natl Acad Sci 95:13743–13748
Berendsen RL, Pieterse CM, Bakker PA (2012) The rhizosphere microbiome and plant health. Trends Plant Sci 17(8):478–486
Bezzerides A, Yong TH, Bezzerides J, Husseini J, Ladau J, Eisner M, Eisner T (2004) Plant-derived pyrrolizidine alkaloid protects eggs of a moth (Utetheisa ornatrix) against a parasitoid wasp (Trichogramma ostriniae). Proc Natl Acad Sci 101(24):9029–9032
Birnbaum SSL, Abbot P (2018) Insect adaptations toward plant toxins in milkweed–herbivores systems—a review. Entomol Exp Appl 166:357–366
Blenn B, Bandoly M, Küffner A, Otte T, Geiselhardt S, Fatouros NE, Hilker M (2012) Insect egg deposition induces indirect defense and epicuticular wax changes in Arabidopsis thaliana. J Chem Ecol 38:882–892
Bonaventure G, VanDoorn A, Baldwin IT (2011) Herbivore-associated elicitors: FAC signaling and metabolism. Trends Plant Sci 16(6):294–299
Boter M, Diaz I (2023) Cyanogenesis, a plant defence strategy against herbivores. Int J Mol Sci 24(8):6982
Bright M, Bulgheresi S (2010) A complex journey: transmission of microbial symbionts. Nat Rev Microbiol 8(3):218–230
Bronstein JL, Huxman TE (1997) The evolution of plant–insect mutualisms. New Phytol 136:373–384
Bronstein JL, Alarcón R, Geber M (2006) The evolution of plant–insect mutualisms. New Phytol 172:412–428
Brownlie JC, Johnson KN (2009) Symbiont-mediated protection in insect hosts. Trends Microbiol 17(8):348–354
Buttery BR, Boatman SG (1976) Water deficits and flow of latex in soil water measurement plant responses and breeding for drought resistance. Academic Press, pp 233–289
Canals D, Irurre-Santilari J, Casas J (2005) The first cytochrome P450 in ferns: evidence for its involvement in phytoecdysteroid biosynthesis in Polypodium vulgare. FEBS J 272(18):4817–4825
Carroll CR, Hoffman CA (1980) Chemical feeding deterrent mobilized in response to insect herbivory and counter adaptation by Epilachna tredecimnotata. Science 209:414–416
Chapman RF (2003) Contact chemoreception in feeding by phytophagous insects. Annu Rev Entomol 48:455–484
Chittka L, Raine NE (2006) Recognition of flowers by pollinators. Curr Opin Plant Biol 9:428–435
Cogni R, Trigo JR, Futuyma DJ (2012) A free lunch? No cost for acquiring defensive plant pyrrolizidine alkaloids in a specialist arctiid moth (Utetheisa ornatrix). Mol Ecol 21:6152–6162
Compant S, Cambon MC, Vacher C, Mitter B, Samad A, Sessitsch A (2021) The plant endosphere world—bacterial life within plants. Environ Microbiol 23(4):1812–1829
Cotter SC, Edwards OR (2006) Quantitative genetics of preference and performance on chickpeas in the noctuid moth, Helicoverpa armigera. Heredity 96:396–402
de la Rosa LA, Moreno-Escamilla JO, Rodrigo-García J, Alvarez-Parrilla E (2019) Phenolic compounds. In: Postharvest physiology and biochemistry of fruits and vegetables. Woodhead Publishing, pp 253–271
del Campo ML, Miles CI, Schroeder FC, Mueller C, Booker R, Renwick JA (2001) Host recognition by the tobacco hornworm is mediated by a host plant compound. Nature 411:186–189
Despland E (2021) Selection forces driving herding of herbivorous insect larvae. Front Ecol Evol 9:760–806
Després L, David JP, Gallet C (2007) The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol Evol 22:298–307
Dicke M, Baldwin IT (2010) The evolutionary context for herbivore-induced plant volatiles: beyond the ‘cry for help’. Trends Plant Sci 15:167–175
Divekar PA, Narayana S, Divekar BA, Kumar R, Gadratagi BG, Ray A, Behera TK (2022) Plant secondary metabolites as defense tools against herbivores for sustainable crop protection. Int J Mol Sci 23:2690
Dobler S (2001) Evolutionary aspects of defense by recycled plant compounds in herbivorous insects. Basic Appl Ecol 2:15–26
Dobler S, Petschenka G, Wagschal V, Flacht L (2015) Convergent adaptive evolution—how insects master the challenge of cardiac glycoside-containing host plants. Entomol Exp Appl 157:30–39
Dussourd DE, Hoyle AM (2000) Poisoned plusiines: toxicity of milkweed latex and cardenolides to some generalist caterpillars. Chemoecology 10:11–16
Ehrlich PR, Raven PH (1964) Butterflies and plants: a study in coevolution. Evolution 18:586–608
Eisenschmidt-Bönn D, Schneegans N, Backenköhler A, Wittstock U, Brandt W (2019) Structural diversification during glucosinolate breakdown: mechanisms of thiocyanate, epithionitrile and simple nitrile formation. Plant J 99(2):329–343
Erb M, Robert CA (2016) Sequestration of plant secondary metabolites by insect herbivores: molecular mechanisms and ecological consequences. Curr Opin Insect Sci 14:8–11
Feyereisen R (2005) Insect P450. In: Comprehensive molecule insect science. Elsevier, Oxford
Fiesel PD, Parks HM, Last RL, Barry CS (2022) Fruity, sticky, stinky, spicy, bitter, addictive, and deadly: evolutionary signatures of metabolic complexity in the Solanaceae. Nat Prod Rep 39(7):1438–1464
Fordyce JA, Agrawal AA (2001) The role of plant trichomes and caterpillar group size on growth and defence of the pipevine swallowtail Battus philenor. J Anim Ecol 70:997–1005
Fox CW, Stillwell RC, Amarillo-s AR, Czesak ME, Messina FJ (2004) Genetic architecture of population differences in oviposition behaviour of the seed beetle Callosobruchus maculatus. J Evol Biol 17:1141–1151
Frago E, Zytynska S (2023) Impact of herbivore symbionts on parasitoid foraging behaviour. Curr Opin Insect Sci 57:101027
Frago E, Dicke M, Godfray HCJ (2012) Insect symbionts as hidden players in insect–plant interactions. Trends Ecol Evol 27(12):705–711
Fürstenberg-Hägg J, Zagrobelny M, Bak S (2013) Plant defense against insect herbivores. Int J Mol Sci 14(5):10242–10297
Garbeva P, Weisskopf L (2020) Airborne medicine: bacterial volatiles and their influence on plant health. New Phytol 226(1):32–43
Garcia LC, Eubanks MD (2019) Overcompensation for insect herbivory: a review and meta-analysis of the evidence. Ecology 100:e02585
Gatehouse JA (2002) Plant resistance towards insect herbivores: a dynamic interaction. New Phytol 156:145–169
Gleadow RM, Møller BL (2014) Cyanogenic glycosides: synthesis, physiology, and phenotypic plasticity. Annu Rev Plant Biol 65:155–185
Glendinning JI, Davis A, Ramaswamy S (2002) Contribution of different taste cells and signaling pathways to the discrimination of “bitter” taste stimuli by an insect. J Neurosci 22:7281–7287
Grunseich JM, Thompson MN, Aguirre NM, Helms AM (2019) The role of plant-associated microbes in mediating host-plant selection by insect herbivores. Plants (Basel) 9(1):6
Hadacek F (2002) Secondary metabolites as plant traits: current assessment and future perspectives. Crit Rev Plant Sci 21:273–322
Hanley ME, Lamont BB, Fairbanks MM, Rafferty CM (2007) Plant structural traits and their role in anti-herbivore defence. Perspect Plant Ecol Evol Syst 8:157–178
Hansen AK, Moran NA (2014) The impact of microbial symbionts on host plant utilization by herbivorous insects. Mol Ecol 23(6):1473–1496
Hartmann T (2007) From waste products to ecochemicals: fifty years research of plant secondary metabolism. Phytochemistry 68(22–24):2831–2846
Heil M (2011) Nectar: generation regulation and ecological functions. Trends Plant Sci 16:191–200
Heil M (2015) Extrafloral nectar at the plant-insect interface: a spotlight on chemical ecology, phenotypic plasticity, and food webs. Annu Rev Entomol 60:213–232
Heil M, Karban R (2010) Explaining evolution of plant communication by airborne signals. Trends Ecol Evol 25:137–144
Heil M, McKey D (2003) Protective ant-plant interactions as model systems in ecological and evolutionary research. Annu Rev Ecol Evol Syst 34:425–553
Helmus MR, Dussourd DE (2005) Glues or poisons: which triggers vein cutting by monarch caterpillars? Chemoecology 15:45–49
Hochberg ME, Gomulkiewicz R, Holt RD, Thompson JN (2000) Weak sinks could cradle mutualistic symbioses-strong sources should harbour parasitic symbioses. J Evol Biol 13:213–222
Hopkins RJ, van Dam NM, van Loon JJ (2009) Role of glucosinolates in insect-plant relationships and multitrophic interactions. Annu Rev Entomol 54:57–83
Howe GA, Jander G (2008) Plant immunity to insect herbivores. Annu Rev Plant Biol 59:41–66
Howe GA, Schaller A (2008) Direct defenses in plants and their induction by wounding and insect herbivores. In: Induced plant resistance to herbivory. Springer, Dordrecht, pp 7–29
Hu G, St. Leger RJ (2002) Field studies using a recombinant mycoinsecticide (Metarhizium anisopliae) reveal that it is rhizosphere competent. Appl Environ Microbiol 68(12):6383–6387
Kroymann J (2011) Natural diversity and adaptation in plant secondary metabolism. Curr Opin Plant Biol 14(3):246–251
Ishida M, Hara M, Fukino N, Kakizaki T, Morimitsu Y (2014) Glucosinolate metabolism, functionality and breeding for the improvement of Brassicaceae vegetables. Breed Sci 64(1):48–59
Jain C, Khatana S, Vijayvergia R (2019) Bioactivity of secondary metabolites of various plants: a review. Int J Pharm Sci Res 10(2):494–504
Jansen-González S, Teixeira SDP, Pereira RAS (2012) Mutualism from the inside: coordinated development of plant and insect in an active pollinating fig wasp. Arthropod Plant Interact 6:601–609
Janzen DH (1980) When is it coevolution? Evolution 34:611–612
Jeschke V, Gershenzon J, Vassão DG (2016) Insect detoxification of glucosinolates and their hydrolysis products. In: Advances in botanical research, vol 80. Academic Press, pp 199–245
Johnson SD, Steiner KE (2000) Long-tongued fly pollination and evolution of floral spur length in the Disadraconis complex (Orchidaceae). Evolution 54:1713–1722
Karban R, Myers JH (1989) Induced plant responses to herbivory. Annu Rev Ecol Syst 20(1):331–348
Kessler A, Baldwin IT (2002) Plant responses to insect herbivory: the emerging molecular analysis. Annu Rev Plant Biol 53(1):299–328
Koch K, Neinhuis C, Ensikat HJ, Barthlott W (2004) Self-assembly of epicuticular waxes on living plant surfaces imaged by atomic force microscopy (AFM). J Exp Bot 55:711–718
Kousar B, Bano A, Khan N (2020) PGPR modulation of secondary metabolites in tomato infested with Spodoptera litura. Agronomy 10(6):778
Krenn HW (2019) Insect mouthparts: form, function, development and performance, vol 5. Springer Nature, Cham
Kuhn J, Pettersson EM, Feld BK, Burse A, Termonia A, Pasteels JM, Boland W (2004) Selective transport systems mediate sequestration of plant glucosides in leaf beetles: a molecular basis for adaptation and evolution. Proc Natl Acad Sci 101:13808–13813
Kumar P, Rathi P, Schöttner M, Baldwin IT, Pandit S (2014) Differences in nicotine metabolism of two Nicotiana attenuata herbivores render them differentially susceptible to a common native predator. PLoS One 9(4):e95982
Labandeira CC (1998) Early history of arthropod and vascular plant associations. Annu Rev Earth Plants Sci 26(1):329–377
Labandeira CC (2013) A paleobiologic perspective on plant–insect interactions. Curr Opin Plant Biol 16:414–421
Macel M (2011) Attract and deter: a dual role for pyrrolizidine alkaloids in plant–insect interactions. Phytochem Rev 10:75–82
Maffei ME (2010) Sites of synthesis, biochemistry and functional role of plant volatiles. S Afr J Bot 76(4):612–631
Maffei ME, Arimura GI, Mithöfer A (2012) Natural elicitors, effectors and modulators of plant responses. Nat Prod Rep 29(11):1288–1303
Malinowski DP, Alloush GA, Belesky DP (2000) Leaf endophyte Neotyphodium coenophialum modifies mineral uptake in tall fescue. Plant Soil 227(1–2):115–126
Manoj SR, Karthik C, Kadirvelu K, Arulselvi PI, Shanmugasundaram T, Bruno B, Rajkumar M (2020) Understanding the molecular mechanisms for the enhanced phytoremediation of heavy metals through plant growth promoting rhizobacteria: a review. J Environ Manag 254:109779
Mathur V, Ulanova D (2022) Microbial metabolites beneficial to plant hosts across ecosystems. Microb Ecol 86:25–48
Mathur V, Ganta S, Raaijmakers CE, Reddy AS, Vet LE, van Dam NM (2011) Temporal dynamics of herbivore-induced responses in Brassica juncea and their effect on generalist and specialist herbivores. Entomol Exp Appl 139:215–225
Mathur V, Tytgat TO, Hordijk CA, Harhangi HR, Jansen JJ, Reddy AS, Van Dam NM (2013a) An ecogenomic analysis of herbivore-induced plant volatiles in Brassica juncea. Mol Ecol 22(24):6179–6196
Mathur V, Wagenaar R, Caissard JC, Reddy AS, Vet LE, Cortesero AM, Van Dam NM (2013b) A novel indirect defence in Brassicaceae: structure and function of extrafloral nectaries in Brassica juncea. Plant Cell Environ 36:528–541
Mithöfer A, Boland W (2012) Plant defense against herbivores: chemical aspects. Annu Rev Plant Biol 63:431–450
Moore BD, Andrew RL, Külheim C, Foley WJ (2014) Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol 201(3):733–750
Mukherjee A, Gaurav AK, Singh S, Yadav S, Bhowmick S, Abeysinghe S, Verma JP (2022) The bioactive potential of phytohormones: a review. Biotechnol Rep 35:e00748
Muller B, Bourdais G, Reidy B, Bencivenni C, Massonneau A, Condamine P, Rolland G, Conéjéro G, Rogowsky P, Tardieu F (2007) Association of specific expansins with growth in maize leaves is maintained under environmental, genetic, and developmental sources of variation. Plant Physiol 143:278–290
Mumm R, Hilker M (2006) Direct and indirect chemical defence of pine against folivorous insects. Trends Plant Sci 11:351–358
Nealis VG, Nault JR (2005) Seasonal changes in foliar terpenes indicate suitability of Douglas-fir buds for western spruce budworm. J Chem Ecol 31:683–696
Nelson AS, Acosta NC, Mooney KA (2019) Plant chemical mediation of ant behavior. Curr Opin Insect Sci 32:98–103
Nguyen VT, Stewart J, Lopez M, Ioannou I, Allais F (2020) Glucosinolates: natural occurrence, biosynthesis, accessibility, isolation, structures, and biological activities. Molecules 25(19):4537
Ninkuu V, Zhang L, Yan J, Fu Z, Yang T, Zeng H (2021) Biochemistry of terpenes and recent advances in plant protection. Int J Mol Sci 22(11):5710
Nishida R (2002) Sequestration of defensive substances from plants by Lepidoptera. Annu Rev Entomol 47:57–92
Nyman T, Julkunen-Tiitto R (2000) Manipulation of the phenolic chemistry of willows by gall-inducing sawflies. Proc Natl Acad Sci 97:13184–13187
Opitz SE, Müller C (2009) Plant chemistry and insect sequestration. Chemoecology 19:117–154
Panda N, Khush GS (1995) Host plant resistance to insects. CAB International, Wallingford
Paniagua Voirol LR, Frago E, Kaltenpoth M, Hilker M, Fatouros NE (2018) Bacterial symbionts in Lepidoptera: their diversity, transmission, and impact on the host. Front Microbiol 9:556
Pelden D, Meesawat U (2019) Foliar idioblasts in different-aged leaves of a medicinal plant (Annona muricata L.). Songklanakarin J Sci Technol 41:414–420
Peng J, Li Z, Huang Y (2007) Plant species specific defense signal communication differentially regulates glutathione S-transferase activity and gene expression in the Helicoverpa armigera (Hubner). J Plant Interact 2:93–99
Pentzold S, Zagrobelny M, Rook F, Bak S (2014) How insects overcome two-component plant chemical defence: plant β-glucosidases as the main target for herbivore adaptation. Biol Rev 89:531–551
Perkins LE, Cribb BW, Brewer PB, Hanan J, Grant M, de Torres M, Zalucki MP (2013) Generalist insects behave in a jasmonate-dependent manner on their host plants, leaving induced areas quickly and staying longer on distant parts. Proc R Soc B Biol Sci 280:20122646
Peschiutta ML, Scholz FG, Goldstein G, Bucci SJ (2018) Herbivory alters plant carbon assimilation, patterns of biomass allocation and nitrogen use efficiency. Acta Oecol 86:9–16
Petschenka G, Agrawal AA (2016) How herbivores coopt plant defenses: natural selection, specialization, and sequestration. Curr Opin Insect Sci 14:17–24
Phillips MA, Croteau RB (1999) Resin-based defenses in conifers. Trends Plant Sci 4(5):184–190
Phukan MM, Sangma SR, Kalita D, Bora P, Das PP, Manoj K et al (2023) Alkaloids and terpenoids: synthesis, classification, isolation and purification, reactions, and applications. In: Handbook of biomolecules, pp 177–213
Picaud F, Bonnet E, Gloaguen V, Petit D (2003) Decision making for food choice by grasshoppers (Orthoptera: Acrididae): comparison between a specialist species on a shrubby legume and three graminivorous species. Environ Entomol 32:680–688
Pickard WF (2008) Laticifers and secretory ducts: two other tube systems in plants. New Phytol 177:877–888
Pineda A, Zheng SJ, van Loon JJ, Pieterse CM, Dicke M (2010) Helping plants to deal with insects: the role of beneficial soil-borne microbes. Trends Plant Sci 15(9):507–514
Provorov NA, Onishchuk OP (2018) Microbial symbionts of insects: genetic organization, adaptive role, and evolution. Microbiology 87(2):151–163
Raffa KF, Andersson MN, Schlyter F (2016) Host selection by bark beetles: playing the odds in a high-stakes game. Adv Insect Physiol 50:1–74
Raguso RA (2008) Wake up and smell the roses: the ecology and evolution of floral scent. Annu Rev Ecol Evol Syst 39:549–569
Rasmann S, Bennett A, Biere A, Karley A, Guerrieri E (2017) Root symbionts: powerful drivers of plant above-and belowground indirect defenses. Insect Sci 24(6):947–960
Raupp MJ, Ahern R, Onken B, Reardon R, Bealmear S, Doccola J, Webb R (2008) Comparing hemlock woolly adelgid and elongate hemlock scale control in ornamental and forest hemlocks. In: Onken B, Reardon R [compilers] FourthSympos. Hemlock Woolly Adelgid Eastern United States FHTET-2008-01
Rehman F, Khan FA, Badruddin SMA (2012) Role of phenolics in plant defense against insect herbivory. In: Chemistry of phytopotentials: health, energy and environmental perspectives, pp 309–313
Richards LA, Dyer LA, Smilanich AM, Dodson CD (2010) Synergistic effects of amides from two piper species on generalist and specialist herbivores. J Chem Ecol 36:1105–1113
Robinson ML, Weber MG, Freedman MG, Jordan E, Ashlock SR, Yonenaga J, Strauss SY (2023) Macroevolution of protective coloration across caterpillars reflects relationships with host plants. Proc R Soc B 290:20222293
Rosa-Téllez S, Casatejada-Anchel R, Alcántara-Enguídanos A, Torres-Moncho A, Dohgri M, Martínez-Serra C et al (2023) Serine metabolic networks in plants
Salam U, Ullah S, Tang ZH, Elateeq AA, Khan Y, Khan J et al (2023) Plant metabolomics: an overview of the role of primary and secondary metabolites against different environmental stress factors. Life 13(3):706
Saltveit ME (2017) Synthesis and metabolism of phenolic compounds. In: Fruit and vegetable phytochemicals: chemistry and human health, 2nd edn, pp 115–124
Senthil-Nathan S (2013) Physiological and biochemical effect of neem and other Meliaceae plants secondary metabolites against Lepidopteran insects. Front Physiol 4:359
Schaefer HM, Ruxton GD (2009) Deception in plants: mimicry or perceptual exploitation? Trends Ecol Evol 24:676–685
Schoonhoven LM, van Loon JJA, Dicke M (2005) Insect–plant biology, 2nd edn. Oxford University Press, Oxford
Scott JG, Wen Z (2001) Cytochromes P450 of insects: the tip of the iceberg. Pest Manag Sci 57:958–967
Scudder GGE, Meredith J (1982) The permeability of the midgut of three insects to cardiac glycosides. J Insect Physiol 28:689–694
Shah MMR, Zhang S, Liu T (2015) Whitefly, host plant and parasitoid: a review on their interactions. Asian J Appl Sci Eng 4:48–61
Sharma G, Mathur V (2020) Modulation of insect-induced oxidative stress responses by microbial fertilizers in Brassica juncea. FEMS Microbiol Ecol 96(4):fiaa040
Sharma G, Malthankar PA, Mathur V (2021) Insect–plant interactions: a multilayered relationship. Ann Entomol Soc Am 114:1–16
Sharma G, Agarwal S, Verma K, Bhardwaj R, Mathur V (2023) Therapeutic compounds from medicinal plant endophytes: molecular and metabolic adaptations. J Appl Microbiol 134:lxad074
Shikano I, Rosa C, Tan CW, Felton GW (2017) Tritrophic interactions: microbe-mediated plant effects on insect herbivores. Annu Rev Phytopathol 55:313–331
Simon JC, d’Alencon E, Guy E, Jacquin-Joly E, Jaquiery J, Nouhaud P et al (2015) Genomics of adaptation to host-plants in herbivorous insects. Brief Funct Genomics 14(6):413–423
Song YY, Ye M, Li C, He X, Zhu-Salzman K, Wang RL, Zeng RS (2014) Hijacking common mycorrhizal networks for herbivore-induced defence signal transfer between tomato plants. Sci Rep 4(1):3915
Sorensen JS, Dearing MD (2006) Efflux transporters as a novel herbivore counter mechanism to plant chemical defenses. J Chem Ecol 32:1181–1196
Speed MP, Fenton A, Jones MG, Ruxton GD, Brockhurst MA (2015) Coevolution can explain defensive secondary metabolite diversity in plants. New Phytol 208(4):1251–1263
Stone GN, Schönrogge K (2003) The adaptive significance of insect gall morphology. Trends Ecol Evol 18:512–522
Stowe KA, Marquis RJ, Hochwender CG, Simms EL (2001) The evolutionary ecology of tolerance to consumer damage. Annu Rev Ecol Syst 31:565–595
Strauss SY, Agrawal AA (1999) The ecology and evolution of plant tolerance to herbivory. Trends Ecol Evol 14:179–185
Sugio A, Dubreuil G, Giron D, Simon JC (2015) Plant–insect interactions under bacterial influence: ecological implications and underlying mechanisms. J Exp Bot 66:467–478
Szklarzewicz T, Michalik A (2017) Transovarial transmission of symbionts in insects. In: Oocytes: maternal information and functions. Springer, Cham, pp 43–67
Tan RX, Zou WX (2001) Endophytes: a rich source of functional metabolites. Nat Prod Rep 18(4):448–459
Termonia A, Hsiao TH, Pasteels JM, Milinkovitch MC (2001) Feeding specialization and host-derived chemical defense in Chrysomeline leaf beetles did not lead to an evolutionary dead end. Proc Natl Acad Sci 98:3909–3914
Thawabteh A, Juma S, Bader M, Karaman D, Scrano L, Bufo SA, Karaman R (2019) The biological activity of natural alkaloids against herbivores, cancerous cells and pathogens. Toxins 11(11):656
Thompson JN, Cunningham BM (2002) Geographic structure and dynamics of coevolutionary selection. Nature 417:735–738
Vale PF, Siva-Jothy J, Morrill A, Forbes M (2018) The influence of parasites on insect behavior. In: Insect behavior: from mechanisms to ecological and evolutionary consequences. Oxford University Press, Oxford, pp 274–291
Van Dam NM, Tytgat TO, Kirkegaard JA (2009) Root and shoot glucosinolates: a comparison of their diversity, function and interactions in natural and managed ecosystems. Phytochem Rev 8:171–186
van den Bosch TJ, Welte CU (2017) Detoxifying symbionts in agriculturally important pest insects. Microb Biotechnol 10:531–540
Van der Putten WH, Vet LE, Harvey JA, Wäckers FL (2001) Linking above-and belowground multitrophic interactions of plants, herbivores, pathogens, and their antagonists. Trends Ecol Evol 16(10):547–554
Venturi V, Keel C (2016) Signaling in the rhizosphere. Trends Plant Sci 21(3):187–198
Vetter J (2000) Plant cyanogenic glycosides. Toxicon 38(1):11–36
War AR, Paulraj MG, Ahmad T, Buhroo AA, Hussain B, Ignacimuthu S, Sharma HC (2012) Mechanisms of plant defense against insect herbivores. Plant Signal Behav 7:1306–1320
War AR, Taggar GK, Hussain B, Taggar MS, Nair RM, Sharma HC (2018) Plant defence against herbivory and insect adaptations. AoB Plants 10(4):ply037
Weng JK, Philippe RN, Noel JP (2012) The rise of chemodiversity in plants. Science 336(6089):1667–1670
Westman SM, Kloth KJ, Hanson J, Ohlsson AB, Albrectsen BR (2019) Defence priming in Arabidopsis—a meta-analysis. Sci Rep 9(1):13309
White JF, Kingsley KL, Zhang Q, Verma R, Obi N, Dvinskikh S, Kowalski KP (2019) Endophytic microbes and their potential applications in crop management. Pest Manag Sci 75(10):2558–2565
Wieczorek MN, Walczak M, Skrzypczak-Zielińska M, Jeleń HH (2018) Bitter taste of Brassica vegetables: the role of genetic factors, receptors, isothiocyanates, glucosinolates, and flavor context. Crit Rev Food Sci Nutr 58(18):3130–3140
Willmer P (2011) Pollination and floral ecology. Princeton University Press, Princeton
Wink M (2019) Quinolizidine and pyrrolizidine alkaloid chemical ecology–a mini-review on their similarities and differences. J Chem Ecol 45(2):109–115
Wink M (2020) Evolution of the angiosperms and co-evolution of secondary metabolites, especially of alkaloids. In: Coevolution of secondary metabolites, pp 151–174
Wöll S, Kim SH, Greten HJ, Efferth T (2013) Animal plant warfare and secondary metabolite evolution. Nat Prod Bioprospect 3:1–7
Woolhouse ME, Webster JP, Domingo E, Charlesworth B, Levin BR (2002) Biological and biomedical implications of the co-evolution of pathogens and their hosts. Nat Genet 32:569–577
Wittstock U, Kliebenstein DJ, Lambrix V, Reichelt M, Gershenzon J (2003) Chapter five Glucosinolate hydrolysis and its impact on generalist and specialist insect herbivores. Recent Adv Phytochem 37:101–125
Yang ZL, Kunert G, Sporer T, Körnig J, Beran F (2020) Glucosinolate abundance and composition in Brassicaceae influence sequestration in a specialist flea beetle. J Chem Ecol 46:186–197
Yulvianti M, Zidorn C (2021) Chemical diversity of plant cyanogenic glycosides: an overview of reported natural products. Molecules 26(3):719
Zunjarrao SS, Tellis MB, Joshi SN, Joshi RS (2020) Plant–insect interactions: the saga of molecular coevolution. In: Co-evolution of secondary metabolites. Springer, Cham, pp 19–45
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2024 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Mathur, V., Sinha, P.G., Noor, S.A. (2024). Unraveling the Coevolutionary Arms Race: Insights into the Dynamic Interplay of Plants, Insects and Associated Organisms. In: Kumar, S., Furlong, M. (eds) Plant Resistance to Insects in Major Field Crops. Springer, Singapore. https://doi.org/10.1007/978-981-99-7520-4_2
Download citation
DOI: https://doi.org/10.1007/978-981-99-7520-4_2
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-99-7519-8
Online ISBN: 978-981-99-7520-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)