
Abstract
Rice is stable and principle crop that feeds the maximum world population. Under the present climatic change scenario, there is a rise in demand for rice production for sustaining an increasing population as the climatic changes leads to yield reduction and quality deterioration of rice. In rice, anthesis and grain filling are considered as very crucial stages, and any changes in the temperature and water availability cause significant reduction in yield. In this climatic change scenario, the rise in temperature is the most potent factor, which is responsible for the reduction of plant growth and yield. It does not only reduce the yield, but also deteriorates the quality of grains in all agricultural crops. Most of the rice is currently cultivated in regions where temperatures are above the optimal for growth (28/22 °C) if there is an increasing temperature above optimal will result in a reduction in productivity and quality. As the temperature rise caused a reduction in precipitation, this will lead to drought stress. Because of these concerning reasons, the demand for tolerant varieties is increasing nowadays. Physiological and biochemical approaches are proving to be the most important screening tools for the identification/development of tolerant genotypes in any crops under high temperature and drought stresses. The chapter was to focus on identifying the major traits that help in developing the significant adaptive and resilient rice varieties which can sustain high temperature and drought stress. This can act as a potential donor or feeding variety to the current population.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
4.1 Introduction
The present climatic change scenario is a challenging act in agriculture that causes a catastrophic impact on food security to feed the rapidly increasing world population. The effects of these climatic changes have started showing their impact on the crops’ productivity and nutritional quality of all the produce (Kadam et al. 2014). Cereals, mainly rice, wheat, and maize, are essential for present food security demands. In all of these, rice is an important staple crop and a primary source of nutrition to more than 3.7 billion people across the globe (International Rice Research Institute, 2016 http://ricestat.irri.org:8080/wrsv3/entrypoint.htm, accessed on April 10, 2021). Demand for food will increase, as the growing population may be reaching nine billion by 2050 (Godfray et al. 2010), mainly in rice-feeding people living in Africa and Asia (FAO 2014). Even with high production demand, less available freshwater and climate change are posing a severe risk to global rice production (Emerick and Ronald 2019). The projected global temperature will rise by 2.8–4.8 °C by 2100 (IPCC, 2018-19). Repeatedly episode of warmth waves has been reported at the regional scale in the last ten years and had a disastrous impact on agricultural crop production across the globe (Kadam et al. 2014). According to the A1B climate change scenario, approximately 121 million ha of irrigated global rice area is projected to be vulnerable to a high temperature by 2100 (Teixeira et al. 2013). Another critical factor that affects the production was the availability of freshwater crops development and growth. Under these high-temperature scenarios, precipitation levels are also affected, which will ultimately lead to low water. This climate change impacts the earth’s crust resulting in infrequent and erratic precipitations, elevated temperatures, and expansion of affected land areas under flood or water deficit. These adverse conditions contribute to the development of the drought-prone regions and, consequently, plant growth and crop productivity.
This stress showed significant detrimental effects on the rice reproductive stage. Rice reproductive stage is considered the most sensitive stage to high stress (HS) (Wassmann et al. 2009; Jagadish et al. 2008; Yoshida 1981). High temperature coinciding at the reproductive stage could result in impaired and poor pollen development, and another dehiscence resulted in high spikelet sterility in rice (Powell et al. 2012; Jagadish et al. 2010a, b). Bahuguna and Krishna (2015) suggested that a high temperature beyond 33 °C (physiological optimum) could affect plant development via alternating molecular and physiological events. Generation of reactive oxygen species (ROS) content is one of the primary events under HS, resulting in the peroxidation of lipids and lower plasma membrane integrity. On the other hand, plants have a robust defense mechanism (antioxidant defense) in maintaining ROS levels under detrimental limits (Szymańska et al. 2017). High-temperature stress also affects the photosynthetic machinery by affecting photochemical reactions in thylakoid lamellae and carbon metabolism within the stroma of chloroplast (Wise et al. 2004; Wahid 2007). Bahuguna and Krishna (2015) suggested that the HS at the later stage could affect the reproductive stage, seed set, and grain filling in rice plants by changing carbon metabolism enzymes and hormonal regulation. Starch synthesis and accumulation during early to mid-grain filling stage result in poor grain filling in HS (Bahuguna et al. 2015).
During flowering, drought stress (DS) on rice has detrimental effects on rice, which resulted in a loss in viability of pollen, pollination failure, and increased flower abortion (Westgate and Ladisch 1993). The key reason for these failures was a reduction in the water potential of floral tissue, i.e., pollen and ovary. Under drought stress, higher production of ROS at the cellular level resulted in significant oxidative damage (Mittler et al. 2004). Short-term and prolonged drought exposure resulted in decreased photosynthesis, leaf development, transpiration, loss of tissue turgor pressure, and various other physiological processes (Tardieu et al. 2018; Jongdee et al. 2002). These changes ultimately affected the anthesis and grain filling stage and resulted in spikelet sterility, and poor grain yield under soil water deficit environment in rice (Jongdee et al. 2002).
Various researchers have studied the impact of these stresses individually, but no reports are available for combined stress effects considered necessary in climatic change. Under natural field conditions, drought and high-temperature stresses can occur concurrently and have more detrimental effects (Rang et al. 2011) than individual stress effects. In Asia, in rice-growing areas, the episode of drought and heat stress at two crucial stages, i.e., flowering and early grain filling, are much every day and responsible for the significant detrimental effect on rice growth and productivity (Wassmann et al. 2009). Other studies documented on various other cereals (rice, maize, wheat, and barley) suggested that combined stress of drought and high temperature showed more detrimental effects than individual ones (Zhang et al. 2013). Heat and drought stresses combined to cause more oxidative stress at the tissue level through overproduction of reactive oxygen species, which damaged protein functions and membrane integrity.
Agricultural crop production faces significant challenges and is considered vulnerable to climatic change like heat spikes, drought spells, increased CO2, and changing rainfall patterns. Crops productivity was dwindled because of both abiotic and biotic factors. In the above stresses, high temperature and drought are the significant constraints that deteriorated productivity and affect the quality of agricultural food crops worldwide (Schleussner et al. 2018). The rising world population and more demands on agricultural productivity to sustain the world population is a challenging task. Therefore rise in demand for cultivated stress-tolerant crop production to uphold the world population is needed (FAO 2017; Godfray et al. 2010). Crop plants can sense stress responses; adapting and sustaining these responses was carried out by alteration in physiological and metabolisms processes. These advances facilitate us to understate the adaptive strategies of plants in these stress conditions (Bita and Gerats 2013; Basu et al. 2016). Crop productive and metabolite process under these stresses was regulated by using certain growth regulators. Due to these climatic changes variability, plants need to develop adaptive mechanisms, modify their metabolism, or alter specific metabolites to cope with these stresses.
4.2 Rice and Its Importance
Cereals play a significant role in human nourishment, contributing to many calories on a per-day basis. Maize, wheat, and rice contribute 10%, 17%, and 23% calories worldwide (Khush et al. 2001). Among all cereals, rice (Oryza sativa L.) is a significant and second most cultivated cereals crop. Rice is grown in flooded conditions and averagely consuming 24.0–30.0% of total available freshwater (Bouman et al. 2007) and requires an optimum air temperature of 32/28 °C day and night temperature. Rice is sensitive to high temperature and drought stress conditions, and the reproductive stage is considered as the most sensitive stage to these stresses (Yoshida 1981; Jagadish et al. 2007, 2008; Sarsu et al. 2018; Basu et al. 2016; Bahuguna et al. 2018). However, the present climate change prediction model suggests a rise of 2.5–4.8 °C temperature by 2100 (Alexander 2016; Masson-Delmotte et al. 2018). As the population demand for freshwater consumption increases and freshwater availability can limit and threaten global rice production (Kadam et al. 2015). In the rice-growing season, the primary concern of drought and HS was conceded at the flowering time, which resulted in S.F. and yield penalty under natural conditions (Matsui and Omasa 2002; Serraj et al. 2011; Liu et al. 2004; Teixeira et al. 2013; Kadam et al. 2015; Pandey and Shukla 2015). Moreover, accurate phenotyping and characterizing a diverse set of germplasms in ambient conditions (field) might give potential tolerant donors which can be utilized in the breeding programs. Although, very few genotypes in rice have been identified and characterized for HS-tolerance.
4.3 Effect of High-Temperature Stress on Rice
Rice’s most susceptible stage to high temperature is flowering (anthesis and fertilization) and also some level at micROS porogenesis stage at booting (Yoshida 1981; Prasad et al. 2006; Jagadish et al. 2008; Farrell et al. 2006). The global climatic change in air surface temperature rises significantly by “0.15 ± 0.05 °C” in 10 years (Jones et al. 2016). The present climatic change model predicate that the average rise in air surface temperature will be of “1.4–5.8 °C by 2100” (Corbera et al. 2016).
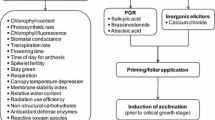
The optimum temperature that’s required for proper rice growth is 32/28 °C day/night temperature. It has been suggested that high temperature at the flowering and grain filling stage was more detrimental in rice (Yang et al. 2007). Peng et al. (2004) revealed the rise in average nighttime temperature by 1.13 °C over 25 years in these stages caused severe damage to growth and yield in rice. It resulted in the sterility of spikelet and decline in yield per plant (Nakagawa et al. 2003). The negative effect of short HS spells has been demonstrated at the flowering stage, resulting in a significantly reduced yield in past decades in China (Welch et al. 2010; Yang et al. 2004; Zou et al. 2009). Hasegawa et al. (2009) suggested that high temperatures at the flowering stage result in the reduction of rice productivity. Shi and Chan (2014) investigated popular rice cultivars grown in the high-temperature sensitive region and found much more prone to HS at the flowering stage. For this problem, there is a need to identify true tolerant and high-yielding varieties that did not only sustain but also get yield under HS conditions (Battisti and Naylor 2009; Lobell and Burke 2008). It was reported earlier that N-22 and NL-44 are high-temperature stress tolerance donors at the vegetative and flowering stages by maintaining higher biomass (Bahuguna et al. 2015; Jagadish et al. 2007). Moreover, precise high-temperature phenotyping is needed to characterize germplasms set under natural and field conditions to identify potential donors used in the future breeding program (Fig. 4.1).

Physiochemical markers to illustrate the effect of high-temperature stress in rice
4.4 Effect of Drought Stress on Rice
Another major constraint responsible for decreased agricultural productivity is drought or water deficit stress (Sharp et al. 2004; Saini and Westgate 2000). Rice is an irrigated crop and is highly vulnerable to drought (Cabuslay et al. 2002). Around ~23 million ha Oryza Sativa L. production is affected by water deficit across the globe (Serraj et al. 2011; Pandey and Shukla 2015). Moreover, its impact is more pronounced because of increased unnatural climatic conditions, which promote drought stress events (Pachauri et al. 2014). As per the climatic model, the frequency of drought spells shortly can be more coinciding with acute stages and results in losses to rice productivity (Kumar et al. 2014). O’toole (1982) reported that drought stress at the vegetative stage result in a lesser reduction in productivity, while the same treatment at meiosis and fertilization results in more considerable yield reduction. At flowering, water deficit resulted in the decline of the grain yield by 30–69% under natural conditions (Cabuslay et al. 2002; Garrity and O’Toole 1994; Sheoran and Saini 1996). Water deficit resulted in stomatal closure and limited carbon accumulation in plants. Leaf water potential was also affected, which diminished turgor pressure in leaf and inhibited stomatal activity and another metabolism (Jaleel et al. 2008). Under drought stress, various plant physiological and biochemical processes were hindered such as membrane integrity, plant water relation, gas exchange, osmolytes content, nutrient management, source-sink association, and growth (Farooq et al. 2008; Benjamin and Nielsen 2006; Jaleel et al. 2008; Razmjoo et al. 2008). Breeding efforts for developing drought resistance rice cultivar that will perform under these predicated climatic change scenarios, but the drought resistance mechanism is poorly understood at the flowering stage (Cabuslay et al. 2002; Condon et al. 2004).
4.5 Effect of High Temperature on Rice Grain Yield, Pollen Viability, and Spikelet Fertility
The flowering stage in rice is most prone to HS and D.S. concerning the vegetative stage (Yoshida 1981). At flowering or anthesis, pollen containing anthers further followed to micro-gametogenesis is the most sensitive tissue to HS and drought stress (Satake and Yoshida 1978; Sheoran and Saini 1996; Nakagawa et al. 2003). The previous report from Yoshida (1981) suggested that manual pollen shaded from non-stress plants to stress stigma showed an ability to fertilize at 41 °C in rice. Therefore pollen grains in the reproductive organ are responsible for lesser S.F. and grain yield in Oryza Sativa L. (Wassmann et al. 2009). A study performed in rice at 35 °C for 5 days during panicle development to anthesis suggested a 35% decline of spikelet fertility (Yoshida 1981). A similar response has been conformed in wheat under high temperatures (Saini and Aspinall 1982).
High-temperature tolerance is mainly associated with three primary mechanisms, i.e., avoiding, escaping, and withstand adverse weather conditions. In avoiding high temperature, stress plant adjusts its tissue temperature by maintaining transpiration rate (Weerakoon et al. 2008). Escaping was associated with early morning flowering and avoiding during peak hours (Ishimaru et al. 2010). Another process is to withstand the adverse climatic conditions with resilient developing reproductive physiology (Jagadish et al. 2010a, b). Based on a field study, Prasad et al. (2006) demonstrated that a rise in 5 °C above ambient temperature significantly reduces spikelet fertility, grain yield, and reduction in harvest index, in various rice cultivars was mainly genotype-dependent. They have also shown a strong positive correlation between S.F. and pollen receptivity and pollen production. A similar response is reported in Japanese cultivars by Matsui et al. (2001) and attributed mainly because of the lower pollen count and poor anther dehiscence on the stigma surface. Jagadish et al. (2007) conducted a controlled environment study in rice and reported that at flowering <1 h, high-temperature exposure was sufficient to decline spikelet fertility. It is related to the fertilization and anther dehiscence in a genotypically dependent manner. Other reports suggested that flowering patterns or early flowering attributes under high temperature can be an essential adaptive trait associated with HS (Jones et al. 1997; Ishimaru et al. 2010).
A comparative study by Fu et al. (2008) reported that HS under field and greenhouse recommended that rise in daily mean temperature was responsible for the reduction of spikelet fertility (6–16%) and grain yield and one thousand grain weight in sensitive genotype more as compared to a tolerant hybrid. Jagadish et al. (2008) reported that 6-h exposure to high temperatures at 33.7 °C and 36.2 °C showed a reduction in spikelet fertility and affected the flowering pattern of rice genotypes. Similarly, Cao et al. (2009) suggested that high temperature from the heading resulted in a more considerable yield reduction in temperature-sensitive genotypes due to poor fertilization and low-speed setting. In contrast, the minimal effect was observed on female reproductive organs in rice. Spikelet fertility reduced significantly in sensitive genotypes, and this reduction was closely associated with germinating pollen on stigma and pollen tube length to the ovary (Jagadish et al. 2010a, b). A study performed by Nagarajan et al. (2010) in rice (basmati and non-basmati genotypes) under high temperature recorded a reduction of S.F. and yield/plant. Bahuguna et al. (2015) identified Nerica L44 (NL-44) for reproductive resilience, with better ROS managing ability, robust antioxidant enzyme system, and high spikelet fertility and grain yield in HS at flowering and vegetative stages.
4.6 Effect of Stress on Rice Grain Yield, Spikelet Fertility, and Pollen Viability
The HS stress is not the only factor responsible for reducing S.F., P.V., and grain yield; drought stress may be responsible for the decline. Mild drought at flowering resulted in a 70% reduction in secondary branching and decreased by 40% in spikelet fertility and panicle number (Kato et al. 2008). In maize, prolonged drought results in reduced sink size and grains number (Bahuguna et al. 2018; Barnabás et al. 2008; Aslam et al. 2013). Drought stress at anthesis results in increased improper anther dehiscence, ovary and pollen abortion, panicle exertion, lower spikelet fertility in rice maize and wheat (Rang et al. 2011; Powell et al. 2012; Praba et al. 2009; Aslam et al. 2013). Drought at pre-anthesis results in asynchronous flowering, or delayed flowering was recorded in wheat (Cattivelli et al. 2008; Foulkes et al. 2007).
The drought stress can affect panicle exertion and cause a reduction in spikelet fertility and grain yield in rice. O’Toole and Namuco (1983) reported that decline in spikelet fertility related to failure in panicle exertion from leaf sheath and matrix water potential of the flag leaf. Selote and Khanna-Chopra (2004) suggested that water stress can decrease spikelet fertility in sensitive rice high-yielding genotypes compared to N22 (tolerant). Various studies indicate that the reproductive stage comprises the microsporangium process for developing functional and viable pollen grains for fertilization and is most critical to stress. Under drought stress, structural and functional disparities create and lead to improper fertilization and seed setting (Saini and Westgate 2000). Pollen development processes like anther dehiscence and pollen shedding are also affected by drought stress (Ekanayake et al. 1990; Satake and Yoshida 1978). It suggested that in DS, sucrose transport to pollen represses, which results in lower P.V. in rice and other cereals (Powell et al. 2012). A similar drought result suggested that drought affects pollen viability during the pollen development stage (Kato et al. 2008; Dolferus et al. 2011). Lilley et al. (1996) and Ekanayake et al. (1993) also suggested that upland rice was more adapted to the water conserved mechanisms than others during drought spells. A greenhouse study performed by Rang et al. (2011) tested five genotypes to drought, found a significant decline in pollen count (52 to 70%), pollen on stigma (59%), and spikelet fertility by 81% in sensitivity genotypes, whereas tolerant (N22) have a lesser reduction in all above traits.
Sharma et al. (2018) performed a pot study in rice and phenotyped 15 rice cultivars by withholding irrigation at anthesis and drought were maintained at ~70 kPa and reported that P.V. (50%), S.F. (48%), and grain yield per plant and 1000 grain weight significantly affected under drought as compared to well-watered conditions. Bahuguna et al. (2018) reported that drought at flowering reduces spikelet fertility and grain yield/plant. They also recommended that priming at the critical stage helps in kick-start stress signaling and helps in developing acclimatize response against stress. The genetic diversity analysis performed on 296 Indica rice panels under drought stress also significantly reduced grain yield, spikelet fertility, 1000 grain weight, and spikelet number per panicle at 64 kPa and 45 kPa, respectively (Kadam et al. 2018).
4.7 Effect of High Temperature on Rice Grain Yield, Spikelet Fertility, and Pollen Viability
Some studies have reported combined effects of drought and high-temperatures stress and suggested that combined stress was much more detrimental than individual ones. Rang et al. (2011) indicated a decline in S.F. concerning particular stress and normal conditions. Lawas et al. (2019) reported a reduction of 73% yield due to combined high temperature and drought stress in sensitive genotypes compared to control ones.
4.8 Heat Susceptibility Index and Cumulative Stress Response Index
Fischer and Maurer (1978) categorized wheat genotypes in water deficit based on yield; they performed under drought stress and identified them according to their susceptibility index. In brief, the rate of change in yield between two environments relatively means a shift in overall genotypes. Later, Dai et al. (1994) and Koti et al. (2007) suggested the cumulative stress response index, all the sum of individual relative components response of treatment was the best way to identify the actual response of cultivars in any stress conditions.
4.9 Effect of High Temperature on Rice Seed Quality
Studying the HS effect on rice quality is essential for rice productivity and marketability. Rice’s appearance, cooking, and texture are the main quality traits that are major concerns of consumers (Okabe 1979; Rousset et al. 1999). Various studies have suggested rice grain quality can be a more concerning issue in HS (Madan et al. 2012; Lyman et al. 2013). Most reports are available on control conditions studies (Fitzgerald and Resurreccion 2009; Lisle et al. 2000), and very few studies carried in field conditions. Heat stress at the grain filling and flowering stage can make an abnormal change in color and grain texture due to insufficient starch accumulation (Tsukaguchi and Iida 2008). The grain chalkiness was the major component in determining the rice price and quality.
Yoshioka et al. (2007) characterized conventionally chalky grains using image-based techniques in the white core, milky white, white back, and white belly rice. High temperature deteriorated quality traits like grain appearance hulling, milling yield, high chalkiness percentage, head rice recovery, amylose content which can affect market values of rice (Inatsu 1979; Cock et al. 1976). It may result from changes in the activity starch biosynthesis pathway and alteration in granules packing in rice grains (Tetlow and Emes 2014; Dai et al. 2005).
Head rice recovery is an essential trait for rice selection in West Africa (Sakurai et al. 2006). The study suggested that head recovery is affected by high air temperature and relative humidity (Cnossen et al. 2003; Schluterman and Siebenmorgen 2007). Cooper et al. (2008) suggested that high night temperature was also responsible for significantly reducing head rice yields. Nagata et al. (2004) indicated under high temperature at early grain filling and post-flowering was primarily accountable for rice grain fissuring, which resulted in the reduction of head rice yield. Under high-temperature stress, sink capacity is altered, resulting in lower starch synthesis activity and 1000 grain weight (Jeng et al. 2003; Oh-e et al. 2007). Grain chalkiness is another essential trait that determines rice quality and price in the international market. The high temperature at early grain filling and post-flowering affects the packing of starch granules and created air spaces. As a result, in grains chalky regions developed at maturity (Tashiro and Wardlaw 1991; Zakaria et al. 2002). These loose packing of amyloplasts in kernels results in the formation of chalky grain and differs from translucent grains (Lisle et al. 2000).
Rice grain mainly consists of two types of starch, i.e., amylopectin and amylose. Under normal conditions, amylose concerning amylopectin is responsible for the firmness of cooking grain (Juliano 1992; Blakeney et al. 1994). Under high-temperature reduction of amylose content increases amylopectin in rice grains (Umemoto et al. 2002; Asaoka et al. 1989; Umemoto and Terashima 2002). These changes in amylose content led to sticky rice after cooking (Sakurai et al. 2006) and are not favorable for consumption (Sakurai et al. 2006).
4.10 High Temperature Effect on Gaseous Exchange and Tissue Temperature
Photosynthesis rate (i.e., CO2 uptake) and transpiration rate (i.e., H2O loss) are regulated through the stomatal behavior of the plants. This stomatal pore opening and closing maintain the tissue temperature and moment of metabolites or signaling compounds in plants (Brownlee 2001; Lake et al. 2001).
Pn “(rate of photosynthesis)” is affected under high temperature and drought (Hassan 2006; Yordanov et al. 1999). The limitation in the rate of photosynthesis in DS and HS can occur either due to nonstomatal or stomatal attributes (Shangguan et al. 1999; Ort et al. 1994). Whichever limitation in gas exchange might limit plant growth and development in various stresses. Various researchers recommended that in high temperature and drought individually or in combination rate of photosynthesis and transpiration decline (Zandalinas et al. 2016; Arbona et al. 2013). In high temperatures, the reduction in yield in cereals is associated with carbon assimilation (photosynthesis and transpiration) (Stone 2001). High temperature resulted in a lower rate of photosynthesis because of (1) inactivation of Rubisco enzyme activity (Prasad et al. 2004) and (2) destruction of Photosystem II (PSII) (3) by the decreased rate of consumption of CO2/O2 and/or increased photorespiration in mesophyll cells (Leegood and Lea 1999). PSII is the most prominent high-temperature sensitive component (Gombos et al. 1994). Wise et al. (2004) recorded that HS stress carbon metabolism in the stroma and photochemical reaction in thylakoid lamella of the chloroplast are affected.
Various studies have suggested that high-temperature stress can cause degradation of chlorophyll a and b in the leaf of different crop species (Karim et al. 1999). In Solanum Lycopersicum and Saccharum officinarum, HS increased the chlorophyll a:b ratio and decreased chlorophyll:carotenoids ratio in the least susceptible genotypes (Camejo et al. 2005; Wahid and Ghazanfar 2006). Such chlorophyll degradation or photosynthesis apparatus has been associated with ROS production (Guo et al. 2006). The rate of photosynthesis is affected under high temperatures due to disruption of the thylakoid membrane or inhibiting membrane-associated electron carriers and enzymes (Rexroth et al. 2011). More than 35 °C decreased the activity of Rubisco and can limit the photosynthesis system in various plants (Griffin et al. 2004). Zhang et al. (2018) reported lower photosynthetic rate (P.N.) insensitive rice genotypes as compared to N22 under heat stress. Similarly, reduction in Rubisco activity in rice and wheat study has been suggested as a cause of lower P.N. in HS (Perdomo et al. 2017)
In drought stress, stomatal conductance decreases and ultimately reduces CO2 flow in mesophyll cells. Similarly, reduction in the activity of RuBP (Ribulose biphosphate) (Parry et al. 2002) or Rubisco content (Bota et al. 2004) can affect the rate of photosynthesis under drought stress. Speer et al. (1988) reported that drought stress caused instability to the photosynthesis membrane via dehydration in later stages on the plasma membrane. Chaves et al. (2011) suggested that under severe DS, de novo synthesis of photosynthesis proteins slows down, which adversely affects the photosynthesis capacity and enhanced leaf senescence.
4.11 Effect of High Temperature on Relative Water Content (R.W.C.)
Relative water content (R.W.C.) is a critical trait that showed the degree of hydration in tissue and cell and is an essential feature in better growth and physiological functioning (Silva et al. 2007). Blokhina et al. (2003) suggested that plasma membrane permeability and suitability decreased due to dehydration under drought stress. Vendruscolo et al. (2007) indicated that tolerant varieties maintain R.W.C. concerning sensitive ones under water deficit environments. Those varieties keep relative water content-maintained protoplasm under water stress and ensure yield in rice (Sikuku et al. 2012).
4.12 Effect of High Temperature on Membrane Stability Index (MSI)
The membrane integrity defines the permeability or integrity of the plasma membrane (Blokhina et al. 2003). Under stress conditions, plasma membrane permeability increases and increases “electrolytes leakage” from the stress-imposed tissue (Chaturvedi et al. 2012). This electrolytic leakage can be addressed as electrolytic conductance (Agarie et al. 1995). Swapna and Shylaraj (2017) suggested that tolerant varieties have better MSI under drought stress conditions as compared to sensitive ones.
4.13 Effect of High Temperature on Reactive Oxygen Species and Antioxidant System
Under adverse environmental stress conditions, plants activate various types of defense mechanisms in genes, proteins, and signaling cascades to confer tolerance. These stress conditions are liable for the generation of ROS in the organism. ROS act as double-edged sword nature molecules in lower concentration by acting as signaling compounds, whereas in higher it was malevolent nature (Noctor et al. 2014; Baxter et al. 2014). A report summarized ROS as harmful molecules which were highly reactive to nucleic acid, proteins, lipids, and other metabolites (Gill and Tuteja 2010a, b). These ROS molecules comprise various molecules, namely H2O2 oxide radicals (O•2−, OH•, singlet oxygen, and many others). Many reports have suggested that ROS overproduction caused toxicity to nucleic acid, protein, and other metabolites, resulting in cell death and damage (Gill and Tuteja 2010a, b). Under natural conditions balance of ROS and the scavenging mechanism is there, but under stress conditions, this balance gets disturbed and caused cellular damage (Das and Roychoudhury 2014). Szymańska et al. (2017) proposed a robust defense mechanism (antioxidant defense) in maintaining ROS levels under detrimental limits. As ROS activity increased, during HS and D.S., which will possibly affect in peroxidation of lipids and disturbing the integrity of the plasma membrane.
The ROS scavenging components are presented in both enzymatic and non-enzymatic forms. Researchers reported that under drought stress conditions, the expression of enzymatic antioxidants increased in rice (Fahramand et al. 2014) which include S.O.D. (superoxide dismutase), C.A.T. (catalase), A.P.X. (ascorbate peroxidase), G.P.X. (guaiacol peroxidase), G.R. (glutathione reductase), and others (Sahebi et al. 2018), while non-enzymatic are A.A. (ascorbic Acid), G.S.H. (reduced glutathione), α-tocopherol, carotenoids, flavonoids, and proline (Boukar et al. 2019). Abiko et al. (2005) suggested that tolerant plants protect themselves from the harmful effect of ROS by synthesizing antioxidants components. These antioxidants are located at various organs of cells and collaborate to detoxify reactive oxygen species. You and Chan (2015) suggested that in all the enzymatic antioxidants, S.O.D. are the primary defense line of defense which convert O2•- into H2O2 then C.A.T., G.P.X., and A.P.X. play key role in detoxification of H2O2 (You and Chan 2015). It has been shown that ascorbic acid is required for A.P.X. activity while glycol for G.P.X. (Noctor et al. 2014; Dietz et al. 2006; Meyer et al. 2012). Several studies reported that in the rice genome, eight S.O.D. genes encoded viz. one Mn-SOD (manganese S.O.D.), CuZn-SOD (plastidial S.O.D.), CuZn-SOD-L (putative CuZnSOD-like) and 2 copper-zinc S.O.D, cytosolic (cCuZn-SOD1 and cCuZn-SOD2) and iron S.O.D.s (Fe-SOD2 and Fe-SOD3) (Nath et al. 2014). Sharma and Dubey (2005) demonstrated an upregulated activity of S.O.D. in Oryza Sativa L. in water deficit stress. Bahuguna et al. (2015) demonstrated a rise in activity S.O.D., A.P.X., and G.P.X. of tolerant genotypes under high temperature in rice in both vegetative and spikelets and helped minimize high-temperature stress levels. The various transgenic approaches also justified that under-stress overexpression of Mn-SOD1 reduces stress induction (Li et al. 2013). Teixeira et al. (2004, 2006) reported 8 APX genes in rice. Two mitochondrial A.P.X.s (OsAPX5 and OsAPX6), two peroxisomal A.P.X.s (OsAPX3 and OsAPX4), two cytosolic A.P.X.s (OsAPX1 and OsAPX2), and two chloroplastic A.P.X.s (OsAPX7 and OsAPX8) and two cytosolic A.P.X. genes have a crucial role in abiotic stress tolerance (Zhang et al. 2013; Sato et al. 2011) rice. Several researchers proposed under high-temperature stress condition ROS like H2O2, hydroxyl radicals produces at PSII system was scavenge by antioxidant including S.O.D. In signal transduction, the involvement of ROS implies the intensity of stress and to maintain ROS at nontoxic levels, there must be a synchronized function of the regulatory network in balancing ROS production.
Hussain et al. (2019) suggested that in combined HS and DS conditions, metabolites accumulation and enzymatic and non-enzymatic antioxidants increased to prevent oxidative damage in maize. Zhang et al. (2018) suggested similar in rice anthers under high temperature.
4.14 Osmolytes Accumulations in High Temperature and Drought
The critical adaptation for many kinds of cereal under abiotic stress, i.e., heat and drought, is the accumulation of specific low molecular mass organic molecules called osmolytes. Under stress conditions, these osmolytes are accumulated in plants to regulate stress tolerance. Common examples of these osmolytes are sugar, proline, glycine betaine, tertiary and quaternary ammonium compounds, and tertiary sulfonium compounds.
Amphoteric quaternary amine, glycine betaine (G.B.), has a vital role as compatible solutes in various plant abiotic stresses, especially in high temperatures and drought. The glycine betaine content varied from species to species under stress conditions. In maize, it has been reported that more accumulation of G.B. enhances the stress levels under high temperature and drought stress conditions. It was previously reported that it helps improve photosynthesis rate in individual heat and drought and combine stress in wheat. Under water scarcity, it has been reported rise in the level of G.B. in maize and sugarcane (Wahid 2007). It has also been suggested that a rise in temperature by 4–5 °C PSII efficiency inhibited and leads to the accumulation of G.B., which was reported to protect the PSII system from photoinhibition in high-temperature stress. In high-temperature stress, CO2 fixation was affected, resulting in the generation of ROS, G.B. stabilized the Rubisco, resulting in a reduction in ROS generation in crops.
Like glycine, proline is also a critical osmolyte, which accumulates in large quantities under environmental stress conditions. Proline is synthesized from glutamate by the action of two enzymes, Δ1-pyrroline-5-carboxylate synthetase and Δ1-pyrroline-5-carboxylate reductase. It was known that proline also regulates the cellular redox potential under stress environments, exceptionally high temperature, and drought stresses. It also has various functions, such as signaling molecule, a metal chelator, and antioxidant defense. High temperature showed its first presence at the cellular level, especially at the plasma membrane, affecting biochemical events. Kumar et al. (2016) suggested that under high temperature rise in proline content was noted. Under high temperatures, fruit set in tomato plants failed due to disrupting sugar metabolism and proline transport during the narrow window of male reproductive development. Hexose sensing in transgenic plants engineered to produce trehalose, fructans, or mannitol may significantly contribute to the stress-tolerant phenotypes.
Trehalose is a non-reducing disaccharide of glucose that functions as a compatible solute in stabilizing biological structures under abiotic stress in bacteria, fungi, and invertebrates. Trehalose also acts as a storage carbohydrate, and it possesses the unique feature of reversible water absorption capacity to protect biological structures from damage during drought and high temperatures. When water dissipates from the shell of macromolecules (such as protein) during severe dehydration, trehalose can act as a water substitute on the surface of the dried protein. Thus, proteins’ native folding and biological activity are maintained, and denaturation and aggregation are prevented. A study revealed that upregulation of trehalose biosynthetic genes in rice has considerable potential for improving abiotic stress tolerance and, at the same time, augmenting productivity under both stress and non-stress conditions. It suggested that from overexpression of trehalose in a higher capacity for photosynthesis and a concomitant decrease in the extent of photo-oxidative damage during stress. In addition, trehalose must be interacting with other physiological processes to account for changes in ion uptake and partitioning during salt stress. Because other cereal crops, like rice, are also sensitive to abiotic stresses, likely, overexpression of trehalose biosynthetic genes in maize and wheat may also confer high levels of abiotic stress tolerance.
4.15 Hormone Metabolism in High-Temperature Stress
Crops are very susceptible to these changing environmental conditions. To overcome these harmful and adverse conditions, phytohormone, plays a key role, whose signaling helps the plants to adapt and survive under these stress conditions. These are part of key metabolic pathways that help sustain plants and control the yield components (Peleg and Blumwald 2011). Some basic phytohormones are classified into five standard groups viz. ethylene (E.T.), Abscisic acid (A.B.A.), auxin (I.A.A.), gibberellins (G.A.), and cytokinins (C.K.) were mainly present.
Under abiotic stress, crosstalk between hormones develops synergetic or antagonistic relations, which plays a pivotal role in plant development processes. Previous research suggested that auxin (I.A.A.), cytokinin (C.K.), and gibberellic acid (G.A.) are involved in plant development and regulate the response to stress environments (Eyidogan et al. 2012). Zhang et al. (2009) suggested that auxin regulation in modulating the drought response. Likewise, A.B.A. is involved in regulating stomatal closure to prevent water loss to minimize transpiration and decrease growth under drought stress (Schroeder et al. 2001). Xue-Xuan et al. (2010) reported that A.B.A. signaling plays an essential role in developing plants’ drought and cold stress tolerance. E.T. is a gaseous hormone regulating process involving various stages of growth phases, viz. leaf, petals abscission, flower senescence, and fruit ripening and developed in abiotic stress response (Abeles et al. 2012).
Another newly identified group of chemical groups are nitric oxide (NO), salicylic acid (S.A.), strigolactone (S.L.), brassinosteroids (B.R.), jasmonate (J.A.), and polyamines (PAs). In all above, PAs (polyamine) are newly class molecules that play a crucial role in plant development and response under abiotic stresses conditions (Minocha et al. 2014). In various observations, PAs different concentrations were noted during “plant growth and development” and were correlated to “different stresses at cellular levels.” The polyamines are reported for various adaptation and acclimation mechanisms to cope with different stress conditions (Pál et al. 2015; Karwa et al. 2020). Liu et al. (2007) suggested that polyamines act as a secondary messenger in a common signal transduction pathway in Arabidopsis thaliana. Various early and recent work told that PAs have crosstalk with various other hormones viz. gibberellins, auxin, and ethylene for maintaining different processes, but their pronounced relation is not establishing the interaction and mechanism of such crosstalks (Kusano et al. 2008; Alcázar et al. 2010; Bitrián et al. 2012; Xu et al. 2014).
4.16 Conclusion
As the rice is the source of nutrition to the large population of the world, the increasing high-temperature spikes and drought spell the demand for resilient rice varieties rises. Lack of knowledge, how plants can sense and cope up these harsh environments, physiological and biochemical traits which at par correlate with yield and quality traits that can also help us to identify the most important one under these stresses. Under the stress condition, plant either showed tolerant, escape, or avoiding the mechanism for which either it regulates the tissue temperature or by early flowering traits it avoids the high temperature and drought spells. There are various noticeable effects of high temperature and drought is observed on pollen viability, affected grain filling, structural changes of cell organelles, oxidative stress, lipid peroxidation of cell membranes, disruption of leaf water relations, and reduction in photosynthesis. All these effects regulate the water potential, membrane permeability, and osmolytes regulation, which can help in restricting the water losses under stress conditions. Hormone regulation helps in generating the systemic signaling pathway which regulates the H2O2 content and HSPs that help in proper regulation of photosynthesis and transpiration process under stress conditions.
In the future, efforts are needed to phenotype large genotypic variability on the basis of early flowers opening traits and by maintaining spikelet fertility, cell membrane thermostability, photosynthesis, transpiration, HSPs, and maintaining a greater non-structural carbohydrate pool under high temperature and drought stress. A challenge for the future is to achieve the robust tolerant mechanism that can help the plant to sustain high temperature and drought stress environments in climatic change scenarios.
References
Abeles FB, Morgan PW, Saltveit ME (2012) Ethylene in plant biology. Academic
Abiko M, Akibayashi K, Sakata T, Kimura M, Kihara M, Itoh K, Asamizu E, Sato S, Takahashi H, Higashitani A (2005) High-temperature induction of male sterility during barley (Hordeum vulgare L.) anther development is mediated by transcriptional inhibition. Sex Plant Reprod 18:91–100
Agarie S, Hanaoka N, Kubota F, Agata W, Kaufman PB (1995) Measurement of cell membrane stability evaluated by electrolyte leakage as a drought and heat tolerance test in rice (Oryza sativa L.). J Fac Agric Kyushu Univ 40:233–240
Alcázar R, Altabella T, Marco F, Bortolotti C, Reymond M, Koncz C, Carrasco P, Tiburcio AF (2010) Polyamines: molecules with regulatory functions in plant abiotic stress tolerance. Planta 231:1237–1249
Alexander LV (2016) Global observed long-term changes in temperature and precipitation extremes: a review of progress and limitations in IPCC assessments and beyond. Weather Clim Extremes 11:4–16
Arbona V, Manzi M, Ollas CD, Gómez-Cadenas A (2013) Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int J Mol Sci 14:4885–4911
Asaoka M, Okuno K, Hara K, Oba M, Fuwa H (1989) Effects of environmental temperature at the early developmental stage of seeds on the characteristics of endosperm starches of rice (Oryza sativa L.). J Jpn Soc Starch Sci 36:1–8
Aslam M, Zamir MSI, Afzal I, Yaseen M, Mubeen M, Shoaib A (2013) Drought stress, its effect on maize production and development of drought tolerance through potassium application. Cercetari agronomice în Moldova 46(2):154
Bahuguna RN, Krishna SV (2015) Temperature regulation of plant phenological development. Environ Exp Bot 111:83–90
Bahuguna RN, Jha J, Pal M, Shah D, Lawas LM, Khetarpal S, Jagadish KSV (2015) Physiological and biochemical characterization of NERICA-L-44: a novel source of heat tolerance at the vegetative and reproductive stages in rice. Physiol Plant 154:543–559
Bahuguna RN, Tamilselvan A, Muthurajan R, Solis CA, Jagadish SVK (2018) Mild preflowering drought priming improves stress defences, assimilation and sink strength in rice under severe terminal drought. Funct Plant Biol 45:827–839
Barnabás B, Jäger K, Fehér A (2008) The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ 31:11–38
Basu S, Ramegowda V, Kumar A, Pereira A (2016) Plant adaptation to drought stress, vol F1000Research, p 5
Battisti DS, Naylor RL (2009) Historical warnings of future food insecurity with unprecedented seasonal heat. Science 323:240–244
Baxter A, Mittler R, Suzuki N (2014) ROS as key players in plant stress signalling. J Exp Bot 65:1229–1240
Benjamin J, Nielsen DC (2006) Water deficit effects on root distribution of soybean, field pea and chickpea. Field Crop Res 97:248–253
Bita C, Gerats T (2013) Plant tolerance to high temperature in a changing environment: scientific fundamentals and production of heat stress-tolerant crops. Front Plant Sci 4:273
Bitrián M, Zarza X, Altabella T, Tiburcio AF, Alcázar R (2012) Polyamines under abiotic stress: metabolic crossroads and hormonal crosstalks in plants. Meta 2:516–528
Blakeney A, Welsh L, Martin M (1994) Analytical methods for wheat starch amylose. [Conference abstract]. Chemistry in Australia (Australia)
Blokhina O, Virolainen E, Fagerstedt KV (2003) Antioxidants, oxidative damage and oxygen deprivation stress: a review. Ann Bot 91:179–194
Bota J, Medrano H, Flexas J (2004) Is photosynthesis limited by decreased rubisco activity and RuBP content under progressive water stress? New Phytol 162:671–681
Boukar O, Togola A, Chamarthi S, Belko N, Ishikawa H, Suzuki K, Fatokun C (2019) Cowpea [Vigna unguiculata (L.) Walp.] breeding. In: Advances in plant breeding strategies: legumes. Springer, Cham, pp 201–243
Bouman B, Humphreys E, Tuong T, Barker R (2007) Rice and water. Adv Agronomy 92:187–237
Brownlee C (2001) The long and the short of stomatal density signals. Trends Plant Sci 6:441–442
Cabuslay GS, Ito O, Alejar AA (2002) Physiological evaluation of responses of rice (Oryza sativa L.) to water deficit. Plant Sci 163:815–827
Camejo D, Rodríguez P, Morales MA, Dell’Amico JM, Torrecillas A, Alarcón JJ (2005) High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J Plant Physiol 162:281–289
Cao et al (2009) Effect of high temperature during heading and early filling on grain yield and physiological characteristics in indica rice. Acta Agron Sin 35:512–521
Cattivelli L, Rizza F, Badeck F-W, Mazzucotelli E, Mastrangelo AM, Francia E, Marè C, Tondelli A, Stanca AM (2008) Drought tolerance improvement in crop plants: an integrated view from breeding to genomics. Field Crop Res 105:1–14
Chaturvedi GS, Singh A, Bahadur R (2012) Screening techniques for evaluating crop germplasm for drought tolerance. Plant Archives 12:11–18
Chaves MM, Costa JM, Saibo NJM (2011) Recent advances in photosynthesis under drought and salinity. In: Advances in botanical research, vol 57. Academic, pp 49–104
Cnossen AG, Jimenez MJ, Siebenmorgen TJ (2003) Rice fissuring response to high drying and tempering temperatures. J Food Eng 59(1):61–69
Cock J, Yoshida S, Forno DA (1976) Laboratory manual for physiological studies of rice. Int. Rice Res, Inst
Condon AG, Richards RA, Rebetzke GJ, Farquhar GD (2004) Breeding for high water-use efficiency. J Exp Bot 55(407):2447–2460
Cooper NTW, Siebenmorgen TJ, Counce PA (2008) Effects of nighttime temperature during kernel development on rice physicochemical properties. Cereal Chem 85(3):276–282
Corbera E, Calvet-Mir L, Hughes H, Paterson M (2016) Patterns of authorship in the IPCC working group III report. Nat Clim Chang 6(1):94–99
Dai Q, Shaobing P, Chavez AQ, Vergara BS (1994) Intraspecific responses of 188 rice cultivars to enhanced UVB radiation. Environ Exp Bot 34(4):433–442
Dai T, Jing Q, Wang X, Jiang D, Cao W (2005) Ecological variations in yield and rice quality under two environments in different rice genotypes. J Nanjing Agric Univ 28(2):1–6
Das K, Roychoudhury A (2014) Reactive oxygen species (R.O.S.) and response of antioxidants as ROS -scavengers during environmental stress in plants. Front Environ Sci 2:53
Dietz KJ, Jacob S, Oelze ML, Laxa M, Tognetti V, de Miranda SMN, Finkemeier I (2006) The function of peroxiredoxins in plant organelle redox metabolism. J Exp Bot 57(8):1697–1709
Dolferus R, Ji X, Richards RA (2011) Abiotic stress and control of grain number in cereals. Plant Sci 181(4):331–341
Ekanayake IJ, De Datta SK, Steponkus PL (1993) Effect of water deficit stress on diffusive resistance, transpiration, and spikelet desiccation of rice (Oryza sativa L.). Ann Bot 72(1):73–80
Ekanayake IJ, Steponkus PL, De Datta SK (1990) Sensitivity of pollination to water deficits at anthesis in upland rice. Crop Sci 30(2):310–315
Emerick K, Ronald PC (2019) Sub1 rice: engineering rice for climate change. Cold Spring Harb Perspect Biol 11(12):a034637
Eyidogan F, Oz MT, Yucel M, Oktem HA (2012) Signal transduction of phytohormones under abiotic stresses. In: Phytohormones and abiotic stress tolerance in plants (pp 1–48). Springer, Berlin
Fahramand M, Mahmoody M, Keykha A, Noori M, Rigi K (2014) Influence of abiotic stress on proline, photosynthetic enzymes and growth. Int Res J Appl Basic Sci 8(3):257–265
FAO (2014) (Food and Agriculture Organization) Accessed January 05 2018. http://www.fao.org/faostat/en/?#home
FAO I, UNICEF W (2017) The state of food security and nutrition in the world. Food and Agriculture Organization of the United Nations, Rome
Farooq M, Basra SMA, Wahid A, Cheema ZA, Cheema MA, Khaliq A (2008) Physiological role of exogenously applied glycinebetaine to improve drought tolerance in fine grain aromatic rice (Oryza sativa L.). J Agron Crop Sci 194(5):325–333
Farrell TC, Fox KM, Williams RL, Fukai S (2006) Genotypic variation for cold tolerance during reproductive development in rice: screening with cold air and cold water. Field Crop Res 98(2-3):178–194
Fischer RA, Maurer R (1978) Drought resistance in spring wheat cultivars. I. Grain yield responses. Aust J Agric Res 29(5):897–912
Fitzgerald MA, Resurreccion AP (2009) Maintaining the yield of edible rice in a warming world. Funct Plant Biol 36(12):1037–1045
Foulkes MJ, Sylvester-Bradley R, Weightman R, Snape JW (2007) Identifying physiological traits associated with improved drought resistance in winter wheat. Field Crop Res 103(1):11–24
Fu GF, Tao LX, Jian SONG, Xi WANG, Cao LY, Cheng SH (2008) Responses of yield characteristics to high temperature during flowering stage in hybrid rice Guodao 6. Rice Sci 15(3):215–222
Garrity DP, O’Toole JC (1994) Screening rice for drought resistance at the reproductive phase. Field Crop Res 39(2-3):99–110
Gill SS, Tuteja N (2010a) Polyamines and abiotic stress tolerance in plants. Plant Signal Behav 5(1):26–33
Gill SS, Tuteja N (2010b) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48(12):909–930
Godfray HCJ, Beddington JR, Crute IR, Haddad L, Lawrence D, Muir JF, Pretty J, Robinson S, Thomas SM, Toulmin C (2010) Food security: the challenge of feeding 9 billion people. Science 327(5967):812–818
Gombos Z, Wada H, Hideg E, Murata N (1994) The unsaturation of membrane lipids stabilizes photosynthesis against heat stress. Plant Physiol 104(2):563–567
Griffin JJ, Ranney TG, Pharr DM (2004) Photosynthesis, chlorophyll fluorescence, and carbohydrate content of illicium taxa grown under varied irradiance. J Am Soc Hortic Sci 129(1):46–53
Guo YP, Zhou HF, Zhang LC (2006) Photosynthetic characteristics and protective mechanisms against photooxidation during high temperature stress in two citrus species. Sci Hortic 108(3):260–267
Hasegawa T, Kuwagata T, Nishimori M, Ishigooka Y, Murakami M, Yoshimoto M, Kondo M, Ishimaru T, Sawano S, Masaki Y, Matsuzaki H (2009) Recent warming trends and rice growth and yield in Japan. In: MARCO symposium on crop production under heat stress: monitoring, impact assessment and adaptation. National Institute for Agro-Environmental Studies, Tsukuba.
Hassan IA (2006) Effects of water stress and high temperature on gas exchange and chlorophyll fluorescence in Triticum aestivum L. Photosynthetica 44(2):312–315
Hussain HA, Men S, Hussain S, Chen Y, Ali S, Zhang S, Zhang K, Li Y, Xu Q, Liao C, Wang L (2019) Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci Rep 9(1):1–12
Inatsu O (1979) Improvement of the quality of rice grown in Hokkaido. J Jpn Soc Starch Sci 26(3):191–197
Ishimaru T, Hirabayashi H, Ida M, Takai T, San-Oh YA, Yoshinaga S, Ando I, Ogawa T, Kondo M (2010) A genetic resource for early-morning flowering trait of wild rice Oryza officinalis to mitigate high temperature-induced spikelet sterility at anthesis. Ann Bot 106(3):515–520
Jagadish SVK, Cairns J, Lafitte R, Wheeler TR, Price AH, Craufurd PQ (2010a) Genetic analysis of heat tolerance at anthesis in rice. Crop Sci 50(5):1633–1641
Jagadish SVK, Craufurd PQ, Wheeler TR (2007) High temperature stress and spikelet fertility in rice (Oryza sativa L.). J Exp Bot 58(7):1627–1635
Jagadish SVK, Craufurd PQ, Wheeler TR (2008) Phenotyping parents of mapping populations of rice for heat tolerance during anthesis. Crop Sci 48(3):1140–1146
Jagadish SVK, Muthurajan R, Oane R, Wheeler TR, Heuer S, Bennett J, Craufurd PQ (2010b) Physiological and proteomic approaches to address heat tolerance during anthesis in rice (Oryza sativa L.). J Exp Bot 61(1):143–156
Jaleel CA, Manivannan P, Lakshmanan GMA, Gomathinayagam M, Panneerselvam R (2008) Alterations in morphological parameters and photosynthetic pigment responses of Catharanthus roseus under soil water deficits. Colloids Surf B: Biointerfaces 61(2):298–303
Jeng TL, Wang CS, Chen CL, Sung JM (2003) Effects of grain position on the panicle on starch biosynthetic enzyme activity in developing grains of rice cultivar Tainung 67 and its NaN 3-induced mutant. J Agric Sci 141(3-4):303–311
Jones MP, Dingkuhn M, Aluko GK, Semon M (1997) Interspecific Oryza sativa L. x O. glaberrima Steud. progenies in upland rice improvement. Euphytica 94(2):237–246
Jones PD, Parker DE, Osborn TJ, Briffa KR (2016) Global and hemispheric temperature anomalies: land and marine instrumental records (1850-2015). Environmental system science data infrastructure for a virtual ecosystem. Carbon Dioxide Information Analysis Center (CDIAC), Oak Ridge National Laboratory, Oak Ridge, TN (USA)
Jongdee B, Fukai S, Cooper M (2002) Leaf water potential and osmotic adjustment as physiological traits to improve drought tolerance in rice. Field Crop Res 76:153–163
Juliano BO (1992) Structure, chemistry, and function of the rice grain and its fractions. Cereal Foods World 37:772–772
Karwa S, Bahuguna RN, Chaturvedi AK et al (2020) Phenotyping and characterization of heat stress tolerance at reproductive stage in rice (Oryza sativa L.). Acta Physiol Plant 42:29. https://doi.org/10.1007/s11738-020-3016-5
Kadam NN, Struik PC, Rebolledo MC, Yin X, Jagadish SK (2018) Genome-wide association reveals novel genomic loci controlling rice grain yield and its component traits under water-deficit stress during the reproductive stage. J Exp Bot 69(16):4017–4032
Kadam NN, Yin X, Bindraban PS, Struik PC, Jagadish KS (2015) Does morphological and anatomical plasticity during the vegetative stage make wheat more tolerant of water deficit stress than rice? Plant Physiol 167(4):1389–1401
Kadam NN, Xiao G, Melgar RJ, Bahuguna RN, Quinones C, Tamilselvan A, Prasad PVV, Jagadish KS (2014) Agronomic and physiological responses to high temperature, drought, and elevated CO2 interactions in cereals. In: Advances in agronomy, vol 127. Academic, pp 111–156
Karim MA, Fracheboud Y, Stamp P (1999) Photosynthetic activity of developing leaves of Zea mays is less affected by heat stress than that of developed leaves. Physiol Plant 105(4):685–693
Kato Y, Kamoshita A, Yamagishi J (2008) Preflowering abortion reduces spikelet number in upland rice (Oryza sativa L.) under water stress. Crop Sci 48(6):2389–2395
Khush GS, Brar DS, Hardy B (eds) (2001) Rice genetics IV, vol 4. Rice Res. Inst, Int
Koti S, Reddy KR, Kakani VG, Zhao D, Gao W (2007) Effects of carbon dioxide, temperature and ultraviolet-B radiation and their interactions on soybean (Glycine max L.) growth and development. Environ Exp Bot 60(1):1–10
Kumar A, Sharma P, Ambrammal SK (2014) Climatic effects on food grain productivity in India. J Stud Dyn Change 1(1):38–48
Kumar RR, Goswami S, Singh K, Dubey K, Singh S, Sharma R, Verma N, Kala YK, Rai GK, Grover M, Mishra DC (2016) Identification of putative RuBisCo Activase (TaRca1)—the catalytic chaperone regulating carbon assimilatory pathway in wheat (Triticum aestivum) under the heat stress. Front Plant Sci 7:986
Kusano T, Berberich T, Tateda C, Takahashi Y (2008) Polyamines: essential factors for growth and survival. Planta 228(3):367–381
Lake JA, Quick WP, Beerling DJ, Woodward FI (2001) Signals from mature to new leaves. Nature 411(6834):154–154
Lawas LMF, Li X, Erban A, Kopka J, Jagadish SK, Zuther E, Hincha DK (2019) Metabolic responses of rice cultivars with different tolerance to combined drought and heat stress under field conditions. GigaScience 8(5):giz050
Leegood RC, Lea PJ (eds) (1999) Plant biochemistry and molecular biology, vol (No. QK861 P42). Wiley
Li CR, Liang DD, Li J, Duan YB, Li HAO, Yang YC, Qin RY, Li LI, Wei PC, Yang JB (2013) Unravelling mitochondrial retrograde regulation in the abiotic stress induction of rice ALTERNATIVE OXIDASE 1 genes. Plant Cell Environ 36(4):775–788
Lilley JM, Ludlow MM, McCouch SR, O’Toole JC (1996) Locating QTL for osmotic adjustment and dehydration tolerance in rice. J Exp Bot 47(9):1427–1436
Lisle AJ, Martin M, Fitzgerald MA (2000) Chalky and translucent rice grains differ in starch composition and structure and cooking properties. Cereal Chem 77(5):627–632
Liu HP, Dong BH, Zhang YY, Liu ZP, Liu YL (2004) Relationship between osmotic stress and the levels of free, conjugated and bound polyamines in leaves of wheat seedlings. Plant Sci 166(5):1261–1267
Liu JH, Kitashiba H, Wang J, Ban Y, Moriguchi T (2007) Polyamines and their ability to provide environmental stress tolerance to plants. Plant Biotechnol 24(1):117–126
Lobell DB, Burke MB (2008) Why are agricultural impacts of climate change so uncertain? The importance of temperature relative to precipitation. Environ Res Lett 3(3):034007
Lyman NB, Jagadish KS, Nalley LL, Dixon BL, Siebenmorgen T (2013) Neglecting rice milling yield and quality underestimates economic losses from high-temperature stress. PLoS One 8(8)
Madan P, Jagadish SVK, Craufurd PQ, Fitzgerald M, Lafarge T, Wheeler TR (2012) Effect of elevated CO2 and high temperature on seed-set and grain quality of rice. J Exp Bot 63(10):3843–3852
Masson-Delmotte V, Zhai P, Pörtner HO, Roberts D, Skea J, Shukla PR, Pirani A, Moufouma-Okia W, Péan C, Pidcock R, Connors S (2018) IPCC, 2018: summary for policymakers. Global warming of 1
Matsui T, Omasa K (2002) Rice (Oryza sativa L.) cultivars tolerant to high temperature at flowering: anther characteristics. Ann Bot 89(6):683–687
Matsui T, Omasa K, Horie T (2001) The difference in sterility due to high temperatures during the flowering period among japonica-rice varieties. Plant Prod Sci 4(2):90–93
Meyer Y, Belin C, Delorme-Hinoux V, Reichheld JP, Riondet C (2012) Thioredoxin and glutaredoxin systems in plants: molecular mechanisms, crosstalks, and functional significance. Antioxid Redox Signal 17(8):1124–1160
Minocha R, Majumdar R, Minocha SC (2014) Polyamines and abiotic stress in plants: a complex relationship1. Front Plant Sci 5:175
Mittler R, Vanderauwera S, Gollery M, Van Breusegem F (2004) Reactive oxygen gene network of plants. Trends Plant Sci 9(10):490–498
Nagarajan S, Jagadish SVK, Prasad AH, Thomar AK, Anand A, Pal M, Agarwal PK (2010) Local climate affects growth, yield and grain quality of aromatic and non-aromatic rice in northwestern India. Agric Ecosyst Environ 138(3-4):274–281
Nagata K, Takita T, Yoshinaga S, Terashima K, Fukuda A (2004) Effect of air temperature during the early grain-filling stage on grain fissuring in rice [Oryza sativa]. Jpn J Crop Sci (Jpn)
Nakagawa H, Horie T, Matsui T (2003) Effects of climate change on rice production and adaptive technologies. In: International rice research conference, Beijing, China, 16–19 September 2002. International Rice Research Institute.
Nath K, Kumar S, Poudyal RS, Yang YN, Timilsina R, Park YS, Nath J, Chauhan PS, Pant B, Lee CH (2014) Developmental stage-dependent differential gene expression of superoxide dismutase isoenzymes and their localization and physical interaction network in rice (Oryza sativa L.). Genes Genomics 36(1):45–55
Noctor G, Mhamdi A, Foyer CH (2014) The roles of reactive oxygen metabolism in drought: not so cut and dried. Plant Physiol 164(4):1636–1648
O’Toole JC, Namuco OS (1983) Role of panicle exsertion in water stress induced sterility 1. Crop Sci 23(6):1093–1097
Oh-e I, Saitoh K, Kuroda T (2007) Effects of high temperature on growth, yield and dry-matter production of rice grown in the paddy field. Plant Prod Sci 10(4):412–422
Okabe M (1979) Texture measurement of cooked rice and its relationship to the eating quality. J Texture Stud 10(2):131–152
Ort DR, Oxborough K, Wise RR (1994) Depressions of photosynthesis in crops with water deficits. Photoinhibition of photosynthesis from molecular mechanisms to the field, pp 315–329
O’toole JC (1982) Adaptation of rice environments. Drought resistance in crops with emphasis on rice, pp 195–213
Pachauri RK, Allen MR, Barros VR, Broome J, Cramer W, Christ R, Church JA, Clarke L, Dahe Q, Dasgupta P, Dubash NK (2014) Climate change 2014: synthesis report. contribution of working groups I. II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change, 151
Pál M, Szalai G, Janda T (2015) Speculation: polyamines are important in abiotic stress signaling. Plant Sci 237:16–23
Pandey V, Shukla A (2015) Acclimation and tolerance strategies of rice under drought stress. Rice Sci 22(4):147–161
Parry MA, Andralojc PJ, Khan S, Lea PJ, Keys AJ (2002) Rubisco activity: effects of drought stress. Ann Bot 89(7):833–839
Peleg Z, Blumwald E (2011) Hormone balance and abiotic stress tolerance in crop plants. Curr Opin Plant Biol 14(3):290–295
Peng S, Huang J, Sheehy JE, Laza RC, Visperas RM, Zhong X, Centeno GS, Khush GS, Cassman KG (2004) Rice yields decline with higher night temperature from global warming. Proc Natl Acad Sci 101(27):9971–9975
Perdomo JA, Capó-Bauçà S, Carmo-Silva E, Galmés J (2017) Rubisco and rubisco activase play an important role in the biochemical limitations of photosynthesis in rice, wheat, and maize under high temperature and water deficit. Front Plant Sci 8:490
Powell N, Ji X, Ravash R, Edlington J, Dolferus R (2012) Yield stability for cereals in a changing climate. Funct Plant Biol 39(7):539–552
Praba ML, Cairns JE, Babu RC, Lafitte HR (2009) Identification of physiological traits underlying cultivar differences in drought tolerance in rice and wheat. J Agron Crop Sci 195(1):30–46
Prasad PVV, Boote KJ, Allen LH Jr, Sheehy JE, Thomas JMG (2006) Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crop Res 95(2-3):398–411
Prasad PV, Boote KJ, Vu JC, Allen LH Jr (2004) The carbohydrate metabolism enzymes sucrose-P synthase and ADG-pyrophosphorylase in phaseolus bean leaves are upregulated at elevated growth carbon dioxide and temperature. Plant Sci 166(6):1565–1573
Rang ZW, Jagadish SVK, Zhou QM, Craufurd PQ, Heuer S (2011) Effect of high temperature and water stress on pollen germination and spikelet fertility in rice. Environ Exp Bot 70(1):58–65
Razmjoo K, Heydarizadeh P, Sabzalian MR (2008) Effect of salinity and drought stresses on growth parameters and essential oil content of Matricaria chamomile. Int J Agric Biol 10(4):451–454
Rexroth S, Mullineaux CW, Ellinger D, Sendtko E, Rögner M, Koenig F (2011) The plasma membrane of the cyanobacterium Gloeobacter violaceus contains segregated bioenergetic domains. Plant Cell 23(6):2379–2390
Rousset S, Pons B, Martin JF (1999) Identifying objective characteristics that predict clusters produced by sensory attributes in cooked rice. J Texture Stud 30(5):509–532
Sahebi M, Hanafi MM, Rafii MY, Mahmud TMM, Azizi P, Osman M, Abiri R, Taheri S, Kalhori N, Shabanimofrad M, Miah G (2018) Improvement of drought tolerance in rice (Oryza sativa L.): genetics, genomic tools, and the WRKY gene family. BioMed research international, 2018.
Saini HS, Westgate ME (2000) Advances in agronomy
Saini HS, Aspinall D (1982) Abnormal sporogenesis in wheat (Triticum aestivum L.) induced by short periods of high temperature. Ann Bot 49(6):835–846
Sakurai T, Furuya J, Futakuchi K (2006) Rice miller cluster in Ghana and its effects on efficiency and quality improvement (No:1004-2016-78639)
Sarsu F, Mukhtar AG, Forster BP, Ingelbrecht I, Pareek A, Das P, Bahuguna RN, Ashraf M, Kusolwa PM, Singla-Pareek SL (2018) Screening protocols for heat tolerant mutants in rice (No. IAEA-CN--263)
Satake T, Yoshida S (1978) High temperature-induced sterility in indica rice at flowering. Jpn J Crop Sci 47(1):6–17
Sato Y, Masuta Y, Saito K, Murayama S, Ozawa K (2011) Enhanced chilling tolerance at the booting stage in rice by transgenic overexpression of the ascorbate peroxidase gene. OsAPXa Plant Cell Rep 30(3):399–406
Schleussner CF, Deryng D, Müller C, Elliott J, Saeed F, Folberth C, Liu W, Wang X, Pugh TA, Thiery W, Seneviratne SI (2018) Crop productivity changes in 1.5 C and 2 C worlds under climate sensitivity uncertainty. Environ Res Lett 13(6):064007
Schluterman DA, Siebenmorgen TJ (2007) Relating rough rice moisture content reduction and tempering duration to head rice yield reduction. Trans ASABE 50(1):137–142
Selote DS, Khanna-Chopra R (2004) Drought-induced spikelet sterility is associated with an inefficient antioxidant defence in rice panicles. Physiol Plant 121(3):462–471
Schroeder JI, Allen GJ, Hugouvieux V, Kwak JM, Waner D (2001) Guard cell signal transduction. Annu Rev Plant Biol 52(1):627–658
Serraj R, McNally KL, Slamet-Loedin I, Kohli A, Haefele SM, Atlin G, Kumar A (2011) Drought resistance improvement in rice: an integrated genetic and resource management strategy. Plant Prod Sci 14(1):1–14
Shangguan Z, Shao M, Dyckmans J (1999) Interaction of osmotic adjustment and photosynthesis in winter wheat under soil drought. J Plant Physiol 154(5-6):753–758
Sharma L, Dalal M, Verma RK, Kumar SV, Yadav SK, Pushkar S, Kushwaha SR, Bhowmik A, Chinnusamy V (2018) Auxin protects spikelet fertility and grain yield under drought and heat stresses in rice. Environ Exp Bot 150:9–24
Sharma P, Dubey RS (2005) Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul 46(3):209–221
Sharp RE, Poroyko V, Hejlek LG, Spollen WG, Springer GK, Bohnert HJ, Nguyen HT (2004) Root growth maintenance during water deficits: physiology to functional genomics. J Exp Bot 55(407):2343–2351
Sheoran IS, Saini HS (1996) Drought-induced male sterility in rice: changes in carbohydrate levels and enzyme activities associated with the inhibition of starch accumulation in pollen. Sex Plant Reprod 9(3):161–169
Shi H, Chan Z (2014) Improvement of plant abiotic stress tolerance through modulation of the polyamine pathway. J Integr Plant Biol 56(2):114–121
Sikuku PA, Netondo GW, Onyango JC (2012) Physiological and biochemical responses of five nerica rice varieties (Oryza sativa L.) to water deficit at vegetative and reproductive stage.
Silva MDA, Jifon JL, Da Silva JA, Sharma V (2007) Use of physiological parameters as fast tools to screen for drought tolerance in sugarcane. Braz J Plant Physiol 19(3):193–201
Speer M, Schmidt JE, Kaiser WM (1988) Effects of water stress on photosynthesis and related processes. In: Plant membranes. Structure, assembly and function. The Biochemical Society, London, pp 209–221
Stone P (2001) The effects of heat stress on cereal yield and quality. In: Crop responses and adaptations to temperature stress, pp 243–291
Swapna S, Shylaraj KS (2017) Screening for osmotic stress responses in Rice varieties under drought condition. Rice Sci 24(5):253–263
Szymańska R, Ślesak I, Orzechowska A, Kruk J (2017) Physiological and biochemical responses to high light and temperature stress in plants. Environ Exp Bot 139:165–177
Teixeira EI, Fischer G, Van Velthuizen H, Walter C, Ewert F (2013) Global hotspots of heat stress on agricultural crops due to climate change. Agric For Meteorol 170:206–215
Teixeira FK, Menezes-Benavente L, Galvão VC, Margis R, Margis-Pinheiro M (2006) Rice ascorbate peroxidase gene family encodes functionally diverse isoforms localized in different subcellular compartments. Planta 224(2):300
Teixeira FK, Menezes-Benavente L, Margis R, Margis-Pinheiro M (2004) Analysis of the molecular evolutionary history of the ascorbate peroxidase gene family: inferences from the rice genome. J Mol Evol 59(6):761–770
Tashiro TWIF, Wardlaw IF (1991) The effect of high temperature on kernel dimensions and the type and occurrence of kernel damage in rice. Aust J Agric Res 42(3):485–496
Tardieu F, Simonneau T, Muller B (2018) The physiological basis of drought tolerance in crop plants: a scenario-dependent probabilistic approach. Annu Rev Plant Biol 69:733–759
Tetlow IJ, Emes MJ (2014) A review of starch-branching enzymes and their role in amylopectin biosynthesis. IUBMB Life 66(8):546–558
Tsukaguchi T, Iida Y (2008) Effects of assimilate supply and high temperature during grain-filling period on the occurrence of various types of chalky kernels in rice plants (Oryza sativa L.). Plant Prod Sci 11(2):203–210
Umemoto T, Terashima K (2002) Research note: activity of granule-bound starch synthase is an important determinant of amylose content in rice endosperm. Funct Plant Biol 29(9):1121–1124
Umemoto T, Yano M, Satoh H, Shomura A, Nakamura Y (2002) Mapping of a gene responsible for the difference in amylopectin structure between japonica-type and indica-type rice varieties. Theor Appl Genet 104(1):1–8
Vendruscolo ECG, Schuster I, Pileggi M, Scapim CA, Molinari HBC, Marur CJ, Vieira LGE (2007) Stress-induced synthesis of proline confers tolerance to water deficit in transgenic wheat. J Plant Physiol 164(10):1367–1376
Wahid A (2007) Physiological implications of metabolite biosynthesis for net assimilation and heat-stress tolerance of sugarcane (Saccharum officinarum) sprouts. J Plant Res 120(2):219–228
Wahid A, Ghazanfar A (2006) Possible involvement of some secondary metabolites in salt tolerance of sugarcane. J Plant Physiol 163(7):723–730
Wassmann R, Jagadish SVK, Sumfleth K, Pathak H, Howell G, Ismail A, Serraj R, Redona E, Singh RK, Heuer S (2009) Regional vulnerability of climate change impacts on Asian rice production and scope for adaptation. Adv Agron 102:91–133
Weerakoon WMW, Maruyama A, Ohba K (2008) Impact of humidity on temperature-induced grain sterility in rice (Oryza sativa L). J Agron Crop Sci 194(2):135–140
Welch JR, Vincent JR, Auffhammer M, Moya PF, Dobermann A, Dawe D (2010) Rice yields in tropical/subtropical Asia exhibit large but opposing sensitivities to minimum and maximum temperatures. Proc Natl Acad Sci 107(33):14562–14567
Westgate PJ, Ladisch MR (1993) Sorption of organics and water on starch. Ind Eng Chem Res 32(8):1676–1680
Wise RR, Olson AJ, Schrader SM, Sharkey TD (2004) Electron transport is the functional limitation of photosynthesis in field-grown Pima cotton plants at high temperature. Plant Cell Environ 27(6):717–724
Xu D, Guo J, Xu L, Sun X, Sun XZ (2014) The relationship between polyamine oxidase activity and lignin deposition and chrysanthemum flower bud differentiation. Acta Agric Boreali Sinica 29:164–169
Xue-Xuan X, Hong-Bo S, Yuan-Yuan M, Gang X, Jun-Na S, Dong-Gang G, Cheng-Jiang R (2010) Biotechnological implications from abscisic acid (A.B.A.) roles in cold stress and leaf senescence as an important signal for improving plant sustainable survival under abiotic-stressed conditions. Crit Rev Biotechnol 30(3):222–230
Yang HC, Huang ZQ, Jiang ZY, Wang XW (2004) High temperature damage and its protective technologies of early and middle season rice in Anhui province. J Anhui Agric Sci 32(1):3–4
Yang J, Zhang J, Liu K, Wang Z, Liu L (2007) Involvement of polyamines in the drought resistance of rice. J Exp Bot 58(6):1545–1555
Yordanov I, Velikova V, Tsonev T (1999) Influence of drought, high temperature, and carbamide cytokinin 4-PU-30 on photosynthetic activity of bean plants. 1. Changes in chlorophyll fluorescence quenching. Photosynthetica 37(3):447–457
Yoshida S (1981) Fundamentals of rice crop science. Int Rice Res, Inst
Yoshioka Y, Iwata H, Tabata M, Ninomiya S, Ohsawa R (2007) Chalkiness in rice: potential for evaluation with image analysis. Crop Sci 47(5):2113–2120
You J, Chan Z (2015) ROS regulation during abiotic stress responses in crop plants. Front Plant Sci 6:1092
Zakaria S, Matsuda T, Tajima S, Nitta Y (2002) Effect of high temperature at ripening stage on the reserve accumulation in seed in some rice cultivars. Plant Prod Sci 5(2):160–168
Zandalinas SI, Rivero RM, Martínez V, Gómez-Cadenas A, Arbona V (2016) Tolerance of citrus plants to the combination of high temperatures and drought is associated to the increase in transpiration modulated by a reduction in abscisic acid levels. BMC Plant Biol 16(1):105
Zhang C, Li G, Chen T, Feng B, Fu W, Yan J, Islam MR, Jin Q, Tao L, Fu G (2018) Heat stress induces spikelet sterility in rice at anthesis through inhibition of pollen tube elongation interfering with auxin homeostasis in pollinated pistils. Rice 11(1):1–12
Zhang SW, Li CH, Cao J, Zhang YC, Zhang SQ, Xia YF, Sun DY, Sun Y (2009) Altered architecture and enhanced drought tolerance in rice via the down-regulation of indole-3-acetic acid by TLD1/OsGH3. 13 activation. Plant Physiol 151(4):1889–1901
Zhang T, Huang Y, Yang X (2013) Climate warming over the past three decades has shortened rice growth duration in China and cultivar shifts have further accelerated the process for late rice. Glob Chang Biol 19(2):563–570
Zou J, Liu A, Chen X, Zhou X, Gao G, Wang W, Zhang X (2009) Expression analysis of nine rice heat shock protein genes under abiotic stresses and A.B.A. treatment. J Plant Physiol 166(8):851–861
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Karwa, S., Sakhare, A., Vishwakarma, C., Arya, S.S., Pal, M., Kumar, S. (2022). Physiological Traits for Improving Heat Stress Tolerance in Plants. In: Kumar, R.R., Praveen, S., Rai, G.K. (eds) Thermotolerance in Crop Plants. Springer, Singapore. https://doi.org/10.1007/978-981-19-3800-9_4
Download citation
DOI: https://doi.org/10.1007/978-981-19-3800-9_4
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-19-3799-6
Online ISBN: 978-981-19-3800-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)