
Abstract
The effects of UV-B radiation on photosynthesis and photosynthetic apparatus of higher plants are reviewed. In addition to the regulatory role, the UV-B represents an important stress agent. Results of numerous studies demonstrate the adverse effects of UV-B on different plant structures and components, including those essential for the photosynthetic processes. The plant species and genotypes differ in responses and susceptibility to UV-B stress, highlighting the role and importance of acclimation processes and protective mechanisms, including creating the efficient UV screen in plant epidermis and dynamic regulation of photosynthetic processes toward efficient photoprotection. Presented results demonstrate that the rapid, non-invasive, chlorophyll fluorescence-based methods may provide valuable information on the actual functional state of photosynthetic apparatus related to the processes of light energy conversion in the chloroplast, including monitoring of the UV-sensitive sites of the photosynthetic system. In addition to the damages, the acclimation processes to UV-B can be investigated. The prospects for future applications in crop breeding are proposed. In addition to UV-B crop resistance, the nutritional and health benefits of UV-B-induced accumulation of UV-absorbing compounds are discussed, with possible role of screening techniques in fresh vegetables and fruits improvement.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
11.1 Introduction
Plants are exposed to a very complex light environment characterized by large fluctuations of intensity and spectral characteristics of incident solar radiation. Even though ultraviolet (UV) radiation represents only a marginal fraction of solar radiation on the Earth’s surface, contributing by 6% in a case of UV-A (315–400 nm) and less than 0.5% in case of UV-B (280–315 nm) (Favory et al. 2009), the biological effects of the UV, especially of UV-B spectra on the biosphere, are significant, as this fraction of spectra represents an important environmental stressor for photosynthetic organisms (Caldwell et al. 2007).
In similar to visible light, UV intensity is very variable and fluctuating, depending enormously on geographic position, especially the latitude and altitude of the location. As a consequence of ozone depletion, UV levels have increased (Seckmeyer et al. 2008), with adverse effects for all kinds of living organisms, including plants (Vincent and Roy 1993; Ivanov et al. 2000; Herman 2010; Kataria et al. 2014).
Besides, the intensity of UV-B fluctuates over shorter time frames (diurnal or seasonal), following the natural rhythms in solar angles over a year or day, as well as the intra-seasonal fluctuations in the ozone layer (Madronich et al. 2011; Bais et al. 2015). The cycles of UV radiation are commonly disrupted by cloudy weather and plant cover, also generating erratic and unpredictable periodic sunflecks in the understory (Thiel et al. 1996; Heisler et al. 2003; Lopez et al. 2009; Aphalo 2017).
The leaves produced in low PAR environments, such as those in deep shade in canopies or understory environments or glasshouses, are exposed and acclimated to low UV levels (Pollastrini et al. 2011; Barnes et al. 2013). When these plants are exposed to high UV-B levels (e.g., vegetable crops propagated in glasshouses and transplanted to the field), the significant injury of photosynthetic apparatus by UV-B radiation may occur (Wargent 2017), which documents the importance of UV protection and acclimation for plants in a natural environment exposed to a significant intensity and fluctuations of UV radiation. In this chapter, we focused on the main biological effects of UV radiation, especially of UV-B fraction, on the photosynthetic apparatus of plant and crop species and the mechanisms by which plants acclimate to high UV-B levels.
11.2 General Effects of UV-B Radiation on Plants
11.2.1 UV-B as a Regulatory Factor
The effects of UV-B radiation are highly dependent on plant species, the doses of the radiation, and the acclimation level of the plants. Although the UV-B is studied mainly as a stress factor, UV-B plays a significant role in regulating the growth, development, and abiotic/biotic interactions of the plants (Llorens et al. 2020). UV-B radiation can act as a stress-inducing agent or a developmental cue, depending on its intensity and duration of exposure (Yin and Ulm 2017; Yadav et al. 2020). Besides being the source of energy in photosynthesis, light is an important signal regulating plant growth and development. Plants perceive light signals through several protein photoreceptors: phytochromes sensitive to red and far-red light (600–750 nm), cryptochromes, phototropins (and Zeitlupe proteins for blue and UV-A radiation (315–500 nm), and UV Resistance Locus 8 (UVR8) for sensing the UV-B radiation (Jiao et al. 2007; Heijde and Ulm 2012; Tossi et al. 2019). There is strong evidence that UV-B is an environmental regulator controlling gene expression, cellular and metabolic activities, and also the growth and development (Jenkins 2009), and multiple UV-B signaling pathways associated with UVR8 were observed in plants (Wu et al. 2012; Christie et al. 2012; Srivastava et al. 2014). Plants perceive the UV-B signal, leading to modulated growth, development, and metabolism in plant organs. Lower doses of UV-B support the plants by triggering the photomorphogenic responses, for example, downregulation of unnecessary stem elongation, cotyledon expansion, and opening of stomata (Kim et al. 1998; Ulm and Nagy 2005; Jenkins 2009). The most discussed UVR8-dependent responses, including are UV-B-induced photomorphogenesis and the accumulation of UV-B-absorbing flavonols (Tilbrook et al. 2013). The UVR8-mediated pathways are crucial for UV acclimation and plant tolerance (Ballaré et al. 2011; Mannuss et al. 2012; Wargent and Jordan 2013).
In addition to photoprotective responses, UV-B can mitigate the adverse effects of other stresses. The positive effects of UV-B pretreatment on drought tolerance were associated with a higher photosynthetic rate, biomass accumulation, and leaf water content (Manetas et al. 1997; Schmidt et al. 2000; Poulson et al. 2006), as well as enhanced antioxidative capacity of plant tissues under drought (Mátai et al. 2019).
11.2.2 UV-B as a Stress Agent
Besides the regulatory effects, the UV-B is most frequently reported as a stress agent negatively influencing wild and agricultural plants, with direct effects on food supply (Piri et al. 2011; Zuk-Golaszewska et al. 2003). The main reason for the harmful effects is the high energy of short wavelengths leading to initiations of photochemical reactions, including production of reactive oxygen species (ROS), such as superoxide (O2•−) and hydroxyl radicals (•OH), but also hydrogen peroxide (H2O2) and singlet oxygen. These ROS can cause oxidative damage to membrane lipids, nucleic acids, and proteins even at low fluence rates (Hideg and Vass 1996; Jansen et al. 1998; Hideg et al. 2002; Brosche and Strid 2003; Hideg et al. 2013; Hideg and Vass 1996; Foyer et al. 1997). In addition, UV radiation is photochemically absorbed by not only biologically significant molecules, such as nucleic acids, proteins, and lipids, but also carotenoids, porphyrins, and quinones, leading to disruption of the integrity and function of essential macromolecules (DNA, proteins, and lipids), and related deleterious effects at the sub-cellular level (Harm 1980; Zu et al. 2010; Czégény et al. 2016; Strid and Hideg 2017).
Ultraviolet irradiation leads to several biological effects, such as reduction in the cell mitosis and chromosome aberration, and, of course, cell death (Yannarelli et al. 2006; Liu et al. 2015). Crucial effects of UV-B are related to photosynthetic processes, which are summarized in the following sections.
11.3 The Effects of UV-B Radiation on Plant Photosynthesis
Although the UV-B radiation affects multiple physiological processes in plants, the photosynthetic apparatus use to be indicated as the main action target of UV-B (Lidon et al. 2012), and the inhibition of photosynthetic processes are predominantly responsible for UV-B-induced reductions in the biomass of crop plants (Agrawal et al. 2004; Kataria et al. 2012; Kataria et al. 2014).
The downregulation of photosynthesis due to UV-B was observed in Arabidopsis thaliana (Coffey and Jansen 2019; Khudyakova et al. 2019; Schultze and Bilger 2019), blueberry (González-Villagra et al. 2020), grapevine (Doupis et al. 2016), sugar beet (Karvansara and Razavi 2019), and soybean (Choudhary and Agrawal 2015), eggplant (Romanatti et al. 2019), pea (Nogues and Baker 1995), cotton (Zhao et al. 2004), and oilseed rape (Allen et al. 1997).
The effects are partly differing depending on the way of examination. UV-B can reduce the CO2 assimilation rate to one-third of the original in experiments with supplementing UV-B (Bornman and Teramura 1993; Kakani et al. 2003; Lu et al. 2009; Kotilainen et al. 2011; Ranjbarfordoei et al. 2011; Lidon et al. 2012). However, a significant decrease in photosynthesis was also observed in more realistic UV-B exclusion studies (Ruhland et al. 2005; Albert et al. 2011; Berli and Bottini 2013; Gitz et al. 2013). On the other hand, the adverse effects of the natural fluence rates of UV-B radiation on well-acclimated plants of some species were found to be relatively low (Searles et al. 2001; Valkama et al. 2003; Ballaré et al. 2011; Hideg et al. 2013; Comont et al. 2013; Müller et al. 2013a, b; Vidović et al. 2015). The reported variability of the reported results highlights the importance of adaptation and acclimation of plants to UV-B and provides evidence on efficient mechanisms by which some species can resist high doses of UV-B. In the next sub-chapters, we will review the partial effects on different levels of photosynthetic apparatus.
11.4 The Effects on Plant Leaf Area and Leaf Anatomy
The overall photosynthetic performance depends not only on the photosynthetic rate but also on the overall leaf area and canopy structure, which may significantly influence the biomass production and yield of crops. Thus, the regulatory and inhibitory effects leading to reduction of leaf area, plant height, and related growth traits are highly relevant for crop productivity (Gerhardt et al. 2005; Vyšniauskienė and Rančelienė 2014). For example, in cotton plants (Gossypium hirsutum L.) exposed to elevated UV-B radiation (up to 10% higher dose of UV-B), the reductions of height (−14%), leaf area (−29%), and total biomass (−34%) were observed when compared with the plants grown under natural conditions (Gao et al. 2003). Reduced growth of leaf area was also observed in Triticum aestivum (Kataria and Guruprasad 2015), Vigna mungo, V. radiata, and Glycine max (Mazza et al. 1999; Amudha et al. 2005; Guruprasad et al. 2007), Amaranthus tricolor (Kataria and Guruprasad 2015), and Oryza sativa (Teramura et al. 1991). That leads to alterations in the canopy morphology that were described by different authors (Barnes et al. 1990; Ryel et al. 1990).
Additional effects are related to UV-exposed leaf anatomy and morphology changes, which also influence the photosynthetic functions. The characteristic visual symptoms are the changes in the thickness of epidermal, palisade, and mesophyll layers of leaves (Kakani et al. 2003). Palisade parenchyma being the first barrier against UV-B radiation is getting thicker and more compressed, thus decreasing the adverse UV-B effects on cells of spongy parenchyma, which are crucial for CO2 assimilation processes (Romanatti et al. 2019).
In addition to UV-B-induced upregulation of leaf thickness (Bornman and Vogelman 1991; Nagel et al. 1998), multiple anatomical and morphological changes were observed, such as a higher density of trichomes on the leaf surface (Barnes et al. 1996) or a decrease in diameter and number of xylem tubes (Lingakumar and Kulandaivelu 1993). Whereas necrosis and reductions of growth may be considered as the direct negative effects of UV-B, the alterations in morphological and anatomical traits represent the first level of acclimation of the photosynthetic apparatus ensuring plant survival, but with possible adverse effects on the photosynthetic capacity of plants and canopies.
11.5 The Effects on Stomata Functions
The stomata closure is responsible for a substantial part of photosynthetic limitation in various stress conditions (Zhao et al. 2004; Zivcak et al. 2013). However, in the case of UV-B stress, the information on the importance of stomatal closure is controversial. Most frequently, UV-B radiations were shown to have adverse effects on stomatal movements (Eisinger et al. 2003; He et al. 2013). The detrimental effects on CO2 assimilation were particularly severe when combining UV-B stress with other stresses, such as drought or low nutrient stress (Musil and Wand 1994; Nogues et al. 1998; Tian and Lei 2007; Lu et al. 2009; Arroniz-Crespo et al. 2011; Doupis et al. 2016). On the other hand, the cross-tolerance observed at the stomata closure level was demonstrated when UV-B was combined with high-temperature stress (Ibrahim et al. 2013). In woody plants, the long-term increase of UV-B radiation influenced stomatal closure and the stomatal density, both contributing to changes in canopy transpiration and water use efficiency (WUE) (Keiller and Holmes 2001).
The effects of UV-B on stomata opening strongly depend on doses. At the same time, the low UV-B stimulate stomatal opening, but high levels of UV-B lead to stomata closure (Eisinger et al. 2003; Tossi et al. 2014), with adverse effects on CO2 assimilation rate (Jansen and Noort 2000; Lu et al. 2009; Reddy et al. 2013).
The stomata on the adaxial part are more sensitive to UV-B than the guard cells located on the abaxial side of the leaf (Nogues et al. 1999). Whereas decreased stomatal conductance represents a typical stress response, the decrease in stomatal density and changes in the distribution of stomata on the leaf surface represent typical acclimation response (Gitz et al. 2013).
11.6 The Effects of UV-B on Chlorophyll and Carotenoid Content
The effects of UV-B radiation are highly dependent on UV-B doses, plant species, and plant acclimation level. Pigment degradation (chlorophylls and carotenoids) and thylakoid disruption were described as typical symptoms of UV-B stress (Strid and Porra 1992; Gaberscik et al. 2002; Kataria et al. 2013; Leon-Felix 2017; Chen et al. 2019). A UV-induced loss of chlorophyll a, b, and total chlorophyll was found in numerous plant species, such as Zea mays, Hordeum vulgare (Tevini et al. 1981), Pisum sativum (Vu et al. 1982, 1983), Amygdalus dulcis (Ranjbarfordoei et al. 2011), and Capsicum annum (Hoffmann et al. 2015).
Severe damage of assimilation apparatus associated with the chlorophyll breakdown and the decline of biochemical and physiological indicators was observed in Triticum durum exposed to UV-A, UV-B, and UV-C radiations. In addition to chlorophylls, the other pigments, such as carotenoids, flavonoids, and anthocyanins, were negatively influenced, and the proline concentration decreased due to excessive values of UV (Balouchi et al. 2009).
Based on the study of Marwood and Greenberg (1996), chlorophyll a tends to be more affected by the UV-induced damage than chlorophyll b. In similar, Zhang and Chen (2013) showed a lover chlorophyll a/b ratio in Oryza sativa exposed to UV-B treatment, supporting the predominant reduction of chlorophyll a.
Carotenoids were shown to be less affected by UV-B treatment than chlorophylls (Pfundel et al. 1992; Sharma et al. 1998; León-Chan et al. 2017). It may be associated with the fact that carotenoids are considered directly associated with the photoprotection of photosynthetic function under UV-B (Middleton and Teramura 1993). However, some studies also reported a significant decrease in some plant species (Muzafarov et al. 1995; Cicek et al. 2012; Hoffmann et al. 2015), and reduction in carotenoids could have a serious impact on pigments (Agrawal and Rathore 2007; Mishra et al. 2003).
Some authors associated the reductions in photosynthetic pigments with the loss of photosynthetic yield (Jordan et al. 1994). However, the effects on photosynthetic functions seem to be the more probable reason for the decrease in photosynthesis than a simple decrease of photosynthetic absorption due to a decrease in photosynthetic pigment concentrations.
11.6.1 The Effects of UV-B on Thylakoid Membrane Complexes
The damaging effects of UV-B on the chloroplast level are associated with a lower chloroplast number and changes in chloroplast ultrastructure (Fagerberg and Bornman 2005; Holzinger et al. 2004). UV-B causes disintegration of the envelope around chloroplasts (He et al. 1994), as well as dilation of thylakoid membranes (He et al. 1994), resulting in leakage of the membrane, which increases ion permeability (Doughty and Hope 1973; Vass et al. 2005). The thylakoid membrane is the main target of UV-B radiation, leading to reduced functioning and alterations in the membrane organization (Petroluleas 2002). Structural and functional effects of UV-B have been predominantly observed at the level of chloroplast structures, especially at the level of thylakoid membranes (Lidon et al. 2012).
11.6.1.1 The Effects of UV-B on PSII Reaction Centers and Oxygen-Evolving Complex
The PSII reaction centers contain the main proteins D1 and D2, which can be damaged by light, but, under normal conditions in light, the repair and synthesis of these proteins, especially of D1, are fast enough to keep the degradation and synthesis in equilibrium. However, under UV-B exposure, their degradation rate of D1 and D2 is fast (Greenberg et al. 1989; Trebst and Depka 1990; Melis et al. 1992; Jansen et al. 1993; Barbato et al. 1995; Friso et al. 1995; Spetea et al. 1996; Vass 1997; Jansen et al. 1998; Kataria et al. 2014; Faseela and Puthur 2018), which disturbs the equilibrium (Friso et al. 1994; Savitch et al. 2001; Vass et al. 2005; Lidon et al. 2012). During photosynthetic electron transport driven by light, tri-molecular oxygen is produced continuously in OEC, producing superoxide radical (O2•−), hydroxyl radical (•OH), and hydrogen peroxide (H2O2) (Apel and Hirt 2004). The damage to D1 and D2 proteins can also induce the semiquinone radicals induced by UV (Brosche and Strid 2003; Zvezdanovic et al. 2013). The inactivation of PSII reaction center and specifically oxygen-evolving complex are vital for the decrease of PSII quantum efficiency, leading to a significant decrease of oxygen evolution in UV-B conditions (Renger et al. 1986; Renger et al. 1989; Bornman 1989; Barbato et al. 1995; Jordan 1996; Segui et al. 2000; Vass 2012). The damage of D1 protein by UV-B seems to be located close to the lumenal end or in the middle of the second transmembrane helix (Friso et al. 1993), which may affect catalysis of water oxidation in OEC (Svensson et al. 1990; Kamiya and Shen 2003). Overreduction of QA leads to significant damage of D2 protein (Friso et al. 1994; Jansen et al. 1996). However, the damage of D2 protein in conditions of elevated UV-B has not been observed to the extent typical for D1 protein (Friso et al. 1994). In addition to faster damage, UV-B was shown to downregulate the turnover of D1 and D2 proteins (Jordan 1996; Wong et al. 2015).
Nevertheless, other components of the electron transport chain are sensitive to UV-B, as well. For example, the UV-B-mediated impairment of 43 and 47 kDa pigment-protein complexes associated with PSII was clearly demonstrated (Gupta et al. 2008). In addition, changes in quinone binding sites caused by UV were observed (Renger et al. 1989), especially the sensitivity of the QB binding site of the electron transport chain between PSII and PSI (Bornman 1989; Jordan 1996; Cai et al. 2016). The acceptor side of PSII may be affected by UV radiation also via the direct damage of molecules of plastoquinone (PQ) electron carriers (Bornman and Teramura 1993).
Analyses of the UV-B effects on PSII membrane fragments demonstrated that the donor side of PSII, the oxygen-evolving system, is one of the major and primary targets of UV-B damage (Renger et al. 1989; Hideg et al. 1993; Vass 1997; Lidon and Ramalho 2011). The resistance of the reaction center of purple bacteria lacking the water-oxidizing complex to UV-B confirms the major contribution of OEC damage in the inactivation of PSII reaction centers (Tandori et al. 1996). Although the exact mechanism of OEC inactivation by UV-B is not fully clear, it was found that the tetra-nuclear Mn complex of OEC is the primary site of the adverse effects (Hideg et al. 1993; Vass 1997; Szilard et al. 2007).
11.6.1.2 The Effects of UV-B on Light-Harvesting Complexes
The light-harvesting complex of PSII (LCH II) is essential to absorb the incident light and transfer the energy to the PSII reaction center. It also influences the organization of thylakoids. The light-harvesting complex of PSII (LCH II) may be adversely affected by UV-B stress associated with changes in the composition of binding proteins. This effect can be ascribed to a decrease of the transcriptional level of cab genes (Vass et al. 2005) and disconnection of LHC II from PSII, which significantly modifies LHC II function (Lidon et al. 2012; Ashraf and Harris 2013). The damage of the light-harvesting complex of PSII (LCH II) is associated with a reduction of chlorophyll a/b ratio in binding proteins. Whereas chlorophyll a occurs in the core complex of both photosystems (PSI and PSII), chlorophyll b is located in their antenna systems (more in PSI). Thus, the decrease of chlorophyll a to b ratio demonstrates a higher UV-B susceptibility of the core complex compared to the peripheral antenna complexes (Zhang and Chen 2013).
11.6.1.3 The Effects of UV-B on Photosystem I and Cytochrome b6/f
Variability in the effects of UV-B irradiation on photosystem I has been observed, and many studies report only minor or no effects on PSI (Brandle et al. 1977; Kulandaivelu and Noorudeen 1983; Iwanzik et al. 1983; Turcsanyi and Vass 2000). The reason for the lower susceptibility of PSI compared to PSII can be not only the absence of a water-splitting complex in PSI but also the lack of redox-active tyrosine (Hansson and Wydrzynski 1990; Yadav et al. 2017). A slight decline in PSI activity was observed as an acclimation response associated with adjusting the PSI/PSII ratio after UV-B-induced damage of PSII occurred (Yadav et al. 2017). In high UV-B conditions, the destruction of PSI reaction centers was observed, which was reflected by the decrease of fraction of oxidizable reaction centers of PSI (lower P700+), indicated by lower amplitude of absorption change at 700 nm. Alternatively, this effect was demonstrated by the analyses of OJIP transient, indicating a decrease of the amplitude of the I–P phase (ΔVIP), which was shown to be sensitive to UV-B radiation (Çiçek et al. 2020).
In similar to photosystem I, cytochrome b6/f complex is considered as relatively resistant to UV-B (Bornman et al. 1984; Teramura and Ziska 1996; Biswal et al. 1997; Mishra et al. 2008; Lidon et al. 2012), and recent studies pay minimum attention to cytochrome b6/f activity associated with UV-B stress.
11.6.1.4 The Effects of UV-B on Photosynthetic Enzymes
UV-B radiation has significant adverse effects on the activity as well as the content of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) in plants (Vu et al. 1982; Correia et al. 1998; Takeuchi et al. 2002; Savitch et al. 2001; Fedina et al. 2010; Kataria et al. 2013, 2019; Kataria and Guruprasad 2015). The most abundant leaf protein in plants, ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco, EC 4.1.1.39), was found to be sensitive to UV-B damage. The Rubisco protein contains the aromatic amino acids absorbing the UV-B band efficiently and, hence, it represents a typical target of direct physical impairment of the peptide structures (Yu et al. 2013). The alterations of the molecular chain and UV-induced degradation result in Rubisco inactivation and an overall decrease of photosynthetic activity. The reactive oxygen species present in the chloroplast in UV-B conditions may cause division of the larger Rubisco subunit into two polypeptides (Takeuchi et al. 2002; Bouchard et al. 2008; Singh et al. 2017).
An indirect reason for the Rubisco decrease in UV-B stress conditions may be the expression of senescence-associated genes (SAGs), namely, SAG12, as observed in Arabidopsis sp. They serve as encoders of cysteine protease, an enzyme responsible for the upregulation of Rubisco degradation (John et al. 2001).
In conditions of elevated UV radiation, the decline in Rubisco activity correlates with lowering the mRNA level of Rubisco subunits. Diminution is also observed in photosynthetic genes’ expressions (Mackerness et al. 1997; Casati and Walbot 2003; Lv et al. 2021). On the other hand, the UV-B activates the genes encoding antioxidative protection (Strid et al. 1994; Singh et al. 2017).
In addition to Rubisco, the following steps of the Calvin cycle, including the stage of RuBP regeneration, were inhibited by excessive UV-B radiation (Allen et al. 1998). Adverse effects on malic dehydrogenase (MDH), phosphoenolpyruvate carboxylase (PEPC), and chlorophyllase were also observed (Kataria et al. 2019). The studies also indicate other enzymes, such as RuBP and sedoheptulose 1,7-bisphosphatase, which are degraded or downregulated by UV-B (Savitch et al. 2001; Lee et al. 2014). A reduction in photosynthetic under UV-B radiation was attributed to the decline of sucrose biosynthesis, regeneration rate of RuBP, and consumption of triose phosphate. Reduced activities were also observed in catalase and nitrate reductase (Kataria and Guruprasad 2015). In turn, the activity of nitrate reductase was stimulated in leaves of wheat, barley, and common bean grown in experiments, in which the UV-B was excluded (Pal et al. 2006; Moussa and Khodary 2008). In addition to nitrate reductase, the UV-B was found to affect also nitrogen metabolism by inhibiting nitrogenase activity (Dohler et al. 1987).
The chloroplast ATP synthase belongs to the group of components the thylakoid membrane impaired by UV-B radiation. Both reduction of amount (Murphy et al. 1985) and activity of ATP synthase (Zhang et al. 1994; Lee et al. 2014) were reported in response to UV-B irradiation. Analogical results were obtained when the expression level of ATP synthase subunits was investigated (Wang et al. 2015).
11.6.2 Effects of UV-B Radiation on Photosynthetic Apparatus Measured by the Non-invasive Techniques
UV-B exposure of plants influences the emission of chlorophyll fluorescence signals originating from the photosystems and related antenna complexes in thylakoids. As in the case of other stresses, the UV-B-induced photoinhibition of PSII can be indicated according to the values of effective quantum yield (ΔF/Fm′) measured in light-adapted samples (Gómez et al. 1998; Fabón et al. 2012; Inostroza-Blancheteau et al. 2016) or maximum quantum yield of PSII photochemistry (Fv/Fm) measured in dark-adapted samples (Ziska et al. 1993; Šprtová et al. 2000; Ranjbarfordoei et al. 2011; Li et al. 2012; Yoon et al. 2020; Mosadegh et al. 2021). In addition, the UV-induced changes of parameters, such as increase of basal fluorescence(F0) or decrease variable fluorescence (Fv), were also reported, indicating adverse effects on PSII photochemistry, in addition to the decrease of photosynthetic rate and chlorophyll content upon UV-B exposure (Ranjbarfordoei et al. 2011). The application of modulated fluorescence technique was found to be very efficient in UV-B studies (Tevini et al. 1988; van Rensen et al. 2007; Kalaji et al. 2017) as this method enables to analyze the mechanistic aspects of the regulation of electron transport in stress conditions, applying various protocols and parameters (Brestic and Zivcak 2013; Kalaji et al. 2014). For example, applying this method, van Rensen et al. (2007) indicated a decrease in photosystem II efficiency with increasing intensity and the length of UV-B exposure. Studies of photoprotective responses indicated an increase of NPQ parameter (Li et al. 2011; Liu et al. 2012), representing an acclimating response. On the other hand, exclusion of UV-B may also lead to NPQ rise (Láposi et al. 2009), which may be well explained by an increase of the proton motive force due to enhanced ATP synthase activity.
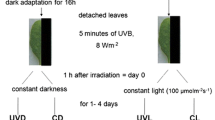
Despite numerous advantages of modulated fluorescence technique, the measurements are time consuming, which limits its use in screening experiments (Brestic and Zivcak 2013; Kalaji et al. 2017). Therefore, an alternative fluorescence technique represented by the analyses of fast chlorophyll a fluorescence transient was proposed (Strasser et al. 1995), which sensitively reflects the damage or modifications in PSII photochemistry. The analysis is based on recordings of polyphasic curves of florescence rise during 1 s light pulse, showing visual changes due to UV-B stress (Fig. 11.1).

OJIP transient measured in control variant without UV (green), moderate UV (red), high UV (brown), and very high UV (purple). The figure left shows the short-term response (day 1, after 4 h of UV exposure); the figure right shows the long-term effects (after 7 days of treatments). The fluorescence curves were processed and visualized using the Biolyzer Software (version 3.06, R. Rodriguez R. Strasser, University of Geneva, Switzerland). Unpublished data by the authors
The functional changes may be estimated based on proportional changes of variable fluorescence values in particular time threshold levels (O, K, J, I, P steps), as shown in Fig. 11.2.

Double normalized OJIP transient (Fo–Fm normalization) measured in control variant without UV (green), moderate UV (red), high UV (brown), and very high UV (purple). The figure left shows the short-term response (day 1, after 4 h of UV exposure); the figure right shows the long-term effects (after 7 days of treatments). The fluorescence curves were processed and visualized using the Biolyzer Software (version 3.06, R. Rodriguez R. Strasser, University of Geneva, Switzerland). Unpublished data by the authors
The numerical analyses of fast chlorophyll kinetics using the model of Strasser et al. (2004) identified the decrease in the number of active reaction centers (Essemine et al. 2012; Mathur and Jajoo 2015), decrease in efficiency of energy trapping per reaction center (TRo/RC) (Albert et al. 2011), the efficiency of light absorption (Yamane et al. 2000; Hollosy 2002; Yu et al. 2013), decrease of quantum efficiency of PSII photochemistry (Guo et al. 2005; Guidi et al. 2007), and inhibition of the electron transfer at the PSII acceptor side (Lidon et al. 2012; Yu et al. 2013; Mathur and Jajoo 2015).
Wang et al. (2010) observed significant inhibition in CO2 assimilation rate and the contents of chlorophyll and carotenoids. The chlorophyll fluorescence indices, such as quantum yield of primary photochemistry (ΦPo), electron transport (ΦEo), and efficiency per trapped excitation (Ψo), declined in conditions of a high UV-B. In parallel, the number of active reaction centers of PSII per functional cross-section (RC/CS) and per absorbed light unit (RC/ABS) changed significantly, as well. It indicates that the photosynthetic function at the level of CO2 assimilation and PSII photochemistry was impaired under the high irradiance of UV-B (Wang et al. 2010).
UV-B causes a reduction in quantum yields and electron transport at both donor and acceptor sides of photosystems, resulting in a decrease of the performance indices, indicating UV-B effects at different levels on the photosynthetic functionality of plants (Çiçek et al. 2020). The values of parameters characterizing the redox state of the PSII acceptor site and the presence of closed reaction centers indicate the transformation of PSII reaction centers into dissipative sinks for excitation energy in the condition of elevated UV radiation (Pan et al. 2011).
The study of Mathur and Jajoo (2015) demonstrated in plants drastically affected by elevated UV radiation the inactivation of oxygen-evolving complex, functional disconnection of light-harvesting complexes from PSII core, and increase of the fraction of inactive reaction centers, all of them contributing to declining of electron transport process indicated by the specific parameters (e.g., ET0/CS).
In all published studies, the observed effects are dependent on UV-B intensity and the length of the treatment. The interaction of these two factors is demonstrated by the leaf models based on the analysis of OJIP transient chlorophyll fluorescence measured in lettuce leaves cultivated in a growth chamber with supplemental UV-B radiation (Fig. 11.3).

Leaf models showing phenomenological energy fluxes per excited cross-section (CS) of lettuce leaves exposed to a different level of UV-B radiation. The models left (a, c, e, g) show the short-term response (day 1, after 4 hours of UV exposure), the models right (b, d, f, h) show the long-term effects (after 7 days of treatments). ABS/CSm absorption flux per excited CS approximated by Fm, TR/CSm trapped energy flux per CS, ET/CSm electron transport flux per CS, DI/CSm dissipated energy flux per CS. Each relative value is represented by the size of proper parameters (arrow), empty circles represent reducing QA reaction centers (active), full black circles represent non-reducing QA reaction centers (inactive or silent). The color intensity of leaves is proportional to its chlorophyll content calculated by Biolyzer software. The models were produced using the Biolyzer Software (version 3.06, R. Rodriguez R. Strasser, University of Geneva, Switzerland). Unpublished data by the authors
The results indicate that whereas the short-term response is associated mainly with the decrease of electron transport at the PSII acceptor side, the long-term UV-B treatment causes accumulation of inactive PSII reaction centers and decrease of light absorbance per leaf cross-section, which reflects the photooxidation of the photosynthetic structures, altogether leading to the drastic decrease of electron transport at the PSII acceptor side.
The fast fluorescence kinetic analysis also enables detection of a specific limitation of electron transport at the PSII donor side associated with impairment of oxygen-evolving complex. That can be well recognized by the double normalization of the initial O–J phase (Fig. 11.4), in which an increase of relative variable fluorescence at 0.3 ms (K step of the transient) represents the specific indicator of OEC damage (Strasser 1997).

Double normalized O-J step of OJIP transient (Fo–Fj normalization) measured in control variant without UV (green), moderate UV (red), high UV (brown), and very high UV (purple). The figure left shows the short-term response (day 1, after 4 hours of UV exposure); the figure right shows the long-term effects (after 7 days of treatments). The fluorescence curves were processed and visualized using the Biolyzer Software (version 3.06, R. Rodriguez R. Strasser, University of Geneva, Switzerland). Unpublished data by the authors
It is evident that short-term UV-B stress in lettuce was not associated with significant damage of OEC, but the elevated K step observed after several days of elevated UV-B indicated that the damage of OEC might be responsible for a substantial number of inactive PSII reaction centers. A decrease in the fraction of OEC measured by an increase of K step in fluorescence transient after exposure to UV-B in wheat was also observed by Mathur and Jajoo (2015). It corresponds to molecular studies and physical methods identifying the OEC as a major target of UV-B damage (Renger et al. 1989; Vass 1997; Lidon and Ramalho 2011).
The alternative fluorescence method used frequently in UV-B studies is the multispectral analysis of the fluorescence signal obtained after excitation by the different monochromatic lights, followed by calculations of fluorescence excitation ratios (Cerovic et al. 2002; Ghozlen et al. 2010). Based on previous knowledge on UV-induced fluorescence (Bilger et al. 1997; Cerovic et al. 1999; Bilger et al. 2001), the ratio of the fluorescence values measured after excitation by red and UV-B light sources was used as an estimate of UV-absorbing compounds in plant epidermis, mainly belonging to the group of flavonols. Indeed, the epidermis with a low content of flavonols transmits the UV light to the chloroplast, leading to a high fluorescence signal. In turn, the epidermis with a high content of flavonols absorbs most UV-B, and the fluorescence signal is low. The absorbance of red light by flavonols is very low; hence, the flavonols’ concentration does not significantly influence the fluorescence signal induced by red light. Therefore, the ratio between the red light-induced fluorescence and UV-light-induced fluorescence is proportional to the flavonol concentration. Analogically, using the green light instead of UV, the anthocyanins with absorption maximum in the green band can be assessed. Due to a non-linear trend of relationships, the logarithms of the ratios were introduced as FLAV and ANTH indices enabling fast and non-invasive estimates of flavonoid and anthocyanin contents in leaves and other aboveground plant organs (Cerovic et al. 2002; Sytar et al. 2016). The most promising were the applications in assessing the quality and maturity level of grapes (Baluja et al. 2012; Tuccio et al. 2020; Agati et al. 2020a, b) or fruits (Lafontaine and Freund 2013; Pinelli et al. 2013; Groher 2019; Agati et al. 2020a, b), but the applications in vegetables (Bruckova et al. 2016; Chaturvedi et al. 2021) or medicinal plants (Müller et al. 2013a, b; Sytar et al. 2015, 2020) were also reported. Thanks to the ability to assess the compounds protecting plants against UV-B stress, the method was successfully used in several studies evaluating the UV-B acclimation responses and co-occurring environmental effects in lettuce (Zivcak et al. 2017; Sytar et al. 2018) or medicinal plant Centella asiatica L. (Müller et al. 2013a, b) or tobacco (Mátai et al. 2019). Some examples of the observed trends in conditions of different UV-B doses will be presented in the next chapter as an acclimation response.
11.7 Acclimation Responses of Photosynthetic Apparatus to UV-B Radiation
11.7.1 Acclimatory and Protection Mechanisms Against UV-B Stress
Jansen et al. (1998) stated that UV-B tolerance depends on the balance between damage reactions and both repair and regulation of the general stress tolerance pathways.
UV-B radiation represents a significant stress factor affecting the growth and productivity of crop plants. The plants dispose of protective mechanisms that eliminate the UV damages, thus maintaining productivity and yield (Kakani 2003; Ballaré et al. 2011; Wargent and Jordan 2013; Ballaré and Austin 2017). The grasses and other monocots use to resist UV-B radiation more efficiently, as their leaves are better arranged to protect apical meristem and leaf sheath. One of the protective responses is lowering of the leaf area (Caldwell and Flint 1994), which was observed in various crops, such as oat (Skórska and Lewandowski 2003), sugar beet (Panagopoulos et al. 1990), maize, and sunflower (Saile-Mark and Tevini 1997). This acclimation helps the plants survive the UV-B stress by reducing exposure to damaging radiation. Additional acclimation response is represented by the synthesis of UV-absorbing compounds, especially flavonoids. They use to be located in the epidermal layer of leaves, and they protect the sensitive cell structures inside leaves by absorbing the excessive UV-B radiation and thus protecting the photosynthetically active tissues against the harmful effects (Tevini et al. 1991; Braun and Tevini 1993; Solovchenko and Schmitz-Eiberger 2003; Hideg and Strid 2017). Flavonoids act as a UV filter absorbing the radiation in the region of 280–320 nm (Singh et al. 2012; Schaller et al. 2013). There are also other compounds accumulated in conditions of elevated UV-B, protecting the photosynthetic tissues from damage, such as the products of the shikimic acid pathway (furanocoumarins, polyketides) and terpenoids, such as cannabinoids (Treutter 2005). In addition, carotenoids have antioxidant properties protecting against harmful effects of UV-B radiation (Middleton and Teramura 1993; Brosche and Strid 2003; Sandmann 2019).
The antioxidative defense supplements the UV-absorbing protection; these two acclimation responses represent the main two pillars of the defense against UV-B damages in higher plants (Caldwell and Flint 1994; Jordan 1996; Piri et al. 2011; Szwarc and Skórska 2007; Vidović et al. 2017). Multiple experimental studies are supporting the importance of the antioxidant defense by the activity of not only the antioxidant enzymes––superoxide dismutase SOD (EC 1.15.1.1), catalase (CAT; EC 1.11.1.6), APX (EC1.11.1.11), glutathione reductase (GR; EC1.6.4.2), guaiacol peroxidase (POD; EC 1.11.1.7), and dehydroascorbate reductase (DHAR; EC1.8.5.1) (Alexieva et al. 2001; Szwarc and Skórska 2007; Wang et al. 2008; Varga et al. 2012; Mishra et al. 2013; Kong et al. 2014; Tripathi et al. 2016)––but also the phenolic antioxidants (Li et al. 1993; Lois and Buchanan 1994), as well as UV-screening effects of the flavonoids (Ziska et al. 1992; Liu et al. 1995; Ravindran et al. 2010; Agati et al. 2020a, b).
Although there is general agreement in the role of antioxidant enzymes in acclimation against UV-B, the experimental studies provide in equal responses for individual antioxidative enzymes in different crops. For example, Agrawal and Rathore (2007) found an increase of SOD and POD but decreased CAT in a condition of high UV-B in wheat. Singh et al. (2012) found an increase of SOD and APX, suggesting the vital role of APX in controlling endogenous hydrogen peroxide content. In the same crop species, Ibrahim et al. (2013) also observed an increase of GPX. In turn, the inverse trend was observed by Tripathi et al. (2016) in wheat, where SOD and APX activities decreased, whereas CAT and POD were enhanced by elevated UV-B. Comparison of the antioxidative enzyme activity in plants grown under ambient and low UV-B indicated higher SOD, APX, POD, and GR activities in UV-rich environment (Kanungo et al. 2013), whereas CAT was found to be lower (Romanatti et al. 2019).
Non-enzymatic antioxidants, such as phenolic acids, ascorbate, reduced glutathione, or α-tocopherol, also play an essential role in alleviating the oxidative stress in high UV environments (Kataria et al. 2007; Selvakumar 2008). The synthesis of UV-B-protecting compounds strongly depends on UV doses. Moderate UV-B intensity stimulates the expression of genes involved in UV-B protection and, hence, synthesis of compounds from the flavonoid family (Brosche and Strid 2003; Ulm et al. 2004). On the other hand, too high UV-B concentrations are not efficient in increasing the synthesis of UV-protecting compounds, and the damage reaction prevails, leading to a decline in the synthesis of antioxidants (Kliebenstein et al. 2002).
The biosynthesis of secondary metabolites plays a significant role in protecting plants from UV-B damage. The dynamics of flavonoid and anthocyanin synthesis after the onset of UV-B radiation in red lettuce grown under monochromatic LED light are shown in Fig. 11.5.

The values of flavonoid index (a) and anthocyanin index (b) measured in the control variant without UV (green), moderate UV (red), high UV (brown), and very high UV (purple). The figure left shows the short-term response (day 1, after 4 h of UV exposure); the figure right shows the long-term effects (after 7 days of treatments). Unpublished results by the authors
Whereas the increase in flavonol content was very fast and steep, the anthocyanin content remained low in the absence of blue light. It confirms that the flavonols play a vital role in protection against UV-B (Agati and Tattini 2010), whereas the anthocyanins serve as regulators of spectral light distribution in visible bands, absorbing quanta in the green region of the solar spectrum (Kytridis and Manetas 2006; Lev-Yadun and Gould 2007; Gould et al. 2010; Agati et al. 2020a, b). The accumulation of UV-absorbing compounds, such as flavonoids, eliminates the transmittance of UV-B to the epidermis, with only minimal effect on transmitting photosynthetically active radiation necessary for photosynthesis (Day et al. 1994; Mazza et al. 1999; Bidel et al. 2007).
There is also the hypothesis that the biosynthesis of phenylpropanoids and flavonoid glycosides represents an energy escape valve using excessive electrons from the photosynthetic electron transport in conditions of stress (Grace and Logan 2000; Hernandez and Van Breusegem 2010). Moreover, Dobrikova and Apostolova (2015) suggested that the flavonoid quercetin present in the chloroplast protects photosynthetic structures against UV-B damage. It provides protection by increasing the production of antioxidants, absorbing the UV-B increasing membrane fluidity, and protecting the Mn cluster, thus decreasing the adverse effect of UV-B.
The production of UV-absorbing compounds is an important UV acclimation response, but it is associated with significant energetic expenses (Snell et al. 2009; Guidi et al. 2011; Hofmann and Jahufer 2011), which causes the differences between the species and environments (Day et al. 1992; Qi et al. 2010; Randriamanana et al. 2015). On the other hand, it is crucial to point out that the production of secondary metabolites as a response to UV-B is linked with cross-tolerance to other abiotic and biotic stresses (Mewis et al. 2012; Bandurska et al. 2013; Zavala et al. 2015), providing additional value to this synthesis.
The evidence on the protective role of flavonoids in lettuce leaves is demonstrated in Fig. 11.6.

(a) The decrease of parameter Fv/Fm parameters after 4 h of UV exposition (difference of Fv/Fm between morning and afternoon measurements) in control variant without UV (green), moderate UV (red), high UV (brown), and very high UV (purple). (b) The relationship between the values of flavonoid index (FLAV index) and decrease of parameter Fv/Fm in various levels of UV-B radiation. Unpublished data by the authors
It is evident that the decrease of Fv/Fm is lowering with an increase of flavonol concentration shown in Fig. 11.5, representing the same experiment with lettuce. Thus, the flavonoid concentration was found to be inversely correlated with the decrease of photochemical efficiency by UV-B. However, it is important to point out that the protection was insufficient at very high UV-B doses, and significant damage occurred despite a very high flavonol accumulation.
In addition to protection, the repair of sensitive targets of UV-B plays an essential role in survival in adverse environments (Favory et al. 2009; Hectors et al. 2009; Schreiner et al. 2017). Moreover, plants possess additional defense mechanisms to protect photosynthetic machinery, such as increased length of epidermal cells, production of a waxy cuticle, and some other morphological and functional adjustments (Hideg et al. 2002).
11.7.2 Positive Effects of UV-B on Productivity, Stress Resistance, and Quality of Production
The acclimation to UV-B may also bring some benefits to plants, with consequences on crop production.
Some studies demonstrate the positive effects of ambient UV-B radiation on photosynthetic rate (Musil and Wand 1994; Favory et al. 2009; Davey et al. 2012; Vidović et al. 2015). It may be associated with UV-B signaling pathways and their regulatory role in expressing specific genes directly linked to photosynthesis (Singh et al. 2014). An increase in electron transport rate was also observed, which was explained as an additional energy need for the biosynthesis of UV-absorbing compounds (Vidović et al. 2015). Kumari et al. (2009) observed in the experiments with the sweet flag that appropriate doses of UV-B radiation led to increased photosynthetic rate, stomatal conductance, and WUE, leading to higher productivity and yield. In similar, wheat trials have shown that curtailing UV-B radiation can improve photosynthesis and productivity of wheat varieties (Kataria and Guruprasad 2015). On the other hand, it is needed to note that most studies report decreased productivity and yield due to UV-B. Nevertheless, the existence of variability in UV-B responses provides a good scope for the subsequent research aimed at molecular and physiological studies of relationships between the UV-B signaling and crop yield.
Compared to the positive effects of UV-B on photosynthesis and yield, much more knowledge exists on the indirect effects of UV-B on physiological responses and qualitative traits of plants and crops. For example, UV-B increased the expression of genes related to pathogenesis-related (PR) protein synthesis, directly promoting resistance to pathogens (Barka et al. 2000; Fujibe et al. 2000; Charles et al. 2009).
More frequently reported are the effects of UV-B on the quality and nutritional value of plant products. In a study by Brzozowska et al. (2014), elevated UV-B during germination increased the content of L-ascorbic acid and polyphenols, and it enhanced the antioxidant activity and improved the sensory properties of germs. UV-B stress increased the content of beta-carotene and lycopene in Cuminum cyminum L. (Ghasemi et al. 2019). UV-B improves the nutritional quality of fruits, potentially making these fruits more attractive (Mariz-Ponte et al. 2019).
The UV-B-induced oxidative stress initiates protective cascades leading to the increase of phenolic compounds and antioxidants (Lobo et al. 2010; Agati et al. 2012; Ilić and Fallik 2017). Typically, the phenylalanine ammonia-lyase (PAL) (de Oliveira et al. 2016), chalcone synthase (CHS), and flavonol synthase (FLS) (Heijde and Ulm 2012) are stimulated, increasing the levels and proportion of different polyphenols in the crops or fruits (Heijde and Ulm 2012). It may be associated with elevated antioxidant content leading to the nutritional benefits to consumer’s health (Lobo et al. 2010). The UV-B may stimulate the production of some volatile organic compounds contributing to fruit flavor and taste (Severo et al. 2016). There are several attempts to apply the UV as a treatment to increase of quality of plant products, especially in the case of vegetables produced in greenhouses (Schreiner et al. 2012; Brazaitytė et al. 2015; Bian et al. 2015; Urban et al. 2016; Dzakovich et al. 2016; Neugart and Schreiner 2018).
11.8 Summary and Conclusions
UV-B radiation represents an important environmental factor. As the photosynthetic apparatus is almost entirely exposed to a dynamic light environment also containing the UV component with its diurnal and seasonal dynamics, the UV-B radiation significantly influences the photosynthetic performance, as well as quantity and quality of crop production. Being an essential regulatory factor, UV-B influences plant morphology and anatomy of photosynthetic organs and structures, with possible effects on leaf area, plant surface structures, including stomatal characteristics. Thus, the UV-B may directly influence the radiation and WUE by plants. In addition to the regulatory role, the UV-B represents an important stress agent. Numerous studies indicate the adverse effects of UV-B on different plant structures, including those essential for the photosynthetic processes. The plant species and genotypes differ in responses and susceptibility to UV-B stress, which highlights the role and importance of acclimation processes and protective mechanisms, such as the building of the efficient antioxidative system, synthesis of UV-absorbing compounds creating the efficient UV screen in plant epidermis, and dynamic regulation of photosynthetic processes toward efficient photoprotection and high efficiency of solar light conversion and assimilation. Graphical examples proposed in this chapter demonstrate that the rapid, non-invasive, chlorophyll fluorescence-based methods may provide valuable information on the actual functional state of photosynthetic apparatus related to the processes of light energy conversion in the chloroplast, including monitoring of the UV-sensitive sites of the photosynthetic system. In addition to the damages, it is possible to monitor also the acclimation processes to UV-B, such as the accumulation of UV-screening compounds in plant epidermis. Thanks to the rapidity, non-invasiveness, and simple applications of the techniques, the scope for future screening programs aimed at identifying highly UV-resistant genotypes opens, which may lead to higher and more stable yields, especially in crops identified as sensitive to UV-B. Moreover, the accumulation of UV-absorbing compounds may be associated with nutritional and health benefits to consumers, especially in fresh vegetables and fruits. Therefore, the future applications in UV-B research should be aimed not only at eliminating the harmful effects of UV-B radiation on photosynthetic processes and overall plant productivity by enhancing the proper acclimation responses but also at efficient exploitations of benefits related to the positive effects of UV radiation on quality and nutritional value of fresh plant products.
This chapter was created thanks to the support of the grant programs, within the projects VEGA 1-0683-20, EPPN2020-OPVaI-VA-ITMS313011T813 and APVV-18-0465.
References
Agati G, Tattini M (2010) Multiple functional roles of flavonoids in photoprotection. New Phytol 186:786–793
Agati G, Azzarello E, Pollastri S, Tattini M (2012) Flavonoids as antioxidants in plants: location and functional significance. Plant Sci 196:67–76
Agati G, Bilger W, Cerovic ZG (2020a) Fluorescence tools for sensing of quality-related phytochemicals in fruits and vegetables. In: Sensor-based quality assessment systems for fruits and vegetables. Apple Academic Press, pp 79–109
Agati G, Brunetti C, Fini A, Gori A, Guidi L, Landi M, Sebastiani F, Tattini M (2020b) Are flavonoids effective antioxidants in plants? Twenty years of our investigation. Antioxidants 9:1098
Agrawal SB, Rathore D (2007) Changes in oxidative stress defense system in wheat (Triticum aestivum L.) and mung bean (Vigna radiata L.) cultivars grown with and without mineral nutrients and irradiated by supplemental ultraviolet-B. Environ Exp Bot 59:21–33
Agrawal SB, Rathore D, Singh A (2004) Combined effects of enhanced UV-B radiation and additional nutrients on two cultivars of wheat (Triticum aestivum L.). Physiol Mol Biol Plants 10:99–108
Albert KR, Mikkelsen TN, Ro-Poulsen H, Arndal MF, Michelsen A (2011) Ambient UV-B radiation reduces PSII performance and net photosynthesis in high Arctic salixarctica. Environ Exp Bot 72:439–447
Alexieva V, Sergiev I, Mapelli S, Karanov E (2001) The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ 24:1337–1344
Allen DJ, Mckee IF, Farage PK, Baker NR (1997) Analysis of limitations to CO2 assimilation on exposure of leaves of two Brassica napus cultivars to UV-B. Plant Cell Environ 20:633–640
Allen DJ, Nogués S, Baker NR (1998) Ozone depletion and increased UV-B radiation: is there a real threat to photosynthesis? J Exp Bot 49:1775–1788
Amudha P, Jayakumar M, Kulandaivelu G (2005) Impacts of ambient solar UV (280–400 nm) radiation on three tropical legumes. J Plant Biol 48:284–291
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress and signal transduction. Ann Rev Plant Biol 55:373–399
Aphalo PJ (2017) Chapter 2. UV-B measurement and methodology. In: Jordan BR (ed) The role of UV-B radiation in plant growth and development. CABI Press, Oxford
Arroniz-Crespo M, Gwynn-Jones D, Callaghan TV, Nunez-Olivera E, Martinez-Abaigar J, Horton P, Phoenix GK (2011) Impacts of long-term enhanced UV-B radiation on bryophytes in two sub-Arctic heathland sites of contrasting water availability. Ann Bot 108:557–565
Ashraf M, Harris PJC (2013) Photosynthesis under stressful environments: an overview. Photosynthetica 51:163–190
Bais AF, Mckenzie RL, Bernhard G, Aucamp PJ, Ilyas M, Madronich S, Tourpali K (2015) Ozone depletion and climate change: impacts on UV radiation. Photochem Photobiol Sci 14:19–52
Ballaré CL, Austin AT (2017) Chapter 3. A perspective on UV-B and terrestrial ecosystems. In: Jordan BR (ed) The role of UV-B radiation in plant growth and development. CABI Press, Oxford
Ballaré CL, Caldwell MM, Flint SD, Robinson SA, Bornman JF (2011) Effects of solar ultraviolet radiation on terrestrial ecosystems. Patterns, mechanisms, and interactions with climate change. Photochem Photobiol Sci 10:226–241
Balouchi HR, Sanavy SAMM, Emam Y, Dolatabadian A (2009) UV radiation, elevated CO2 and water stress effect on growth and photosynthetic characteristics in durum wheat. Plant Soil Environ 55:443–453
Baluja J, Diago MP, Goovaerts P, Tardaguila J (2012) Assessment of the spatial variability of anthocyanins in grapes using a fluorescence sensor: relationships with vine vigour and yield. Precis Agric 13(4):457–472
Bandurska H, Niedziela J, Chadzinikolau T (2013) Separate and combined responses to water deficit and UV-B radiation. Plant Sci 213:98–105
Barbato R, Frizzo A, Friso G, Rigoni F, Giacometti GM (1995) Degradation of the D1 protein of photosystem II reaction centre by UV-B light requires the presence of functional manganese on the donor side. Eur J Biochem 227:723–729
Barka EA, Kalantari S, Makhlouf J, Arul J (2000) Impact of UV-C irradiation on the cell wall-degrading enzymes during ripening of tomato (Lycopersicon esculentum L.) fruit. J Agric Food Chem 48:667–671
Barnes PW, Flint SD, Caldwell MM (1990) Morphological responses of crop and weed species of different growth forms to ultraviolet-B radiation. Am J Bot 77:1354–1360
Barnes PW, Ballare CL, Caldwell MM (1996) Photomorphogenic effects of UV-B radiation on plants: consequences for light competition. J Plant Physiol 148:15–20
Barnes PW, Kersting AR, Flint SD, Beyschlag W, Ryel RJ (2013) Adjustments in epidermal UV-transmittance of leaves in sun-shade transitions. Physiol Plant 149:200–213
Berli FJ, Bottini R (2013) UV-B and abscisic acid effects on grape berry maturation and quality. J Berry Res 3:1–14
Bian ZH, Yang QC, Liu WK (2015) Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: a review. J Sci Food Agric 95:869–877
Bidel LPR, Meyer S, Goulas Y, Cadot Y, Cerovic ZG (2007) Responses of epidermal phenolic compounds to light acclimation: in vivo qualitative and quantitative assessment using chlorophyll fluorescence excitation spectra in leaves of three woody species. J Photochem Photobiol B 88:163–179
Bilger W, Veit M, Schreiber L, Schreiber U (1997) Measurement of leaf epidermal transmittance of UV radiation by chlorophyll fluorescence. Physiol Plant 101(4):754–763
Bilger W, Johnsen T, Schreiber U (2001) UV-excited chlorophyll fluorescence as a tool for the assessment of UV-protection by the epidermis of plants. J Exp Bot 52(363):2007–2014
Biswal B, Joshi PN, Kulandaivelu G (1997) Changes in leaf protein and pigment contents and photosynthetic activities during senescence of detached maize leaves: influence of different ultraviolet radiations. Photosynthetica 34:37–44
Bornman JF (1989) Target sites of UV-B radiation on photosynthesis of higher plants. J Photochem Photobiol B 4:145–158
Bornman JF, Teramura AH (1993) Effects of ultraviolet-B radiation on terrestrial plants. In: Young AR et al (eds) Environmental UV photobiology. Plenum Press, New York, pp 427–477
Bornman JF, Vogelman TC (1991) Effect of UV-B radiation on leaf optical properties measured with fiber optics. J Exp Bot 41:547–554
Bornman JF, Bjorn LO, Akerlund H-E (1984) Action spectrum for inhibition by ultraviolet radiation of photo-system II activity in spinach thylakoids. Photobiochem Photobiophys 8:305–313
Bouchard JN, Longhi ML, Roy S, Campbell DA, Ferreyra G (2008) Interaction of nitrogen status and UV-B sensitivity in a temperate phytoplankton assemblage. J Exp Mar Biol Ecol 359:67–76
Brandle JR, Campbell WF, Sisson WB, Caldwell MM (1977) Net photosynthesis, electron transport capacity and ultrastructure of Pisum sativum L. exposed to ultraviolet-B radiation. Plant Physiol 60:165–169
Braun J, Tevini M (1993) Regulation of UV-protective pigment synthesis in the epidermal layer of rye seedlings (Secale cereale L.). Photochem Photobiol 57:318–323
Brazaitytė A, Viršilė A, Jankauskienė J, Sakalauskienė S, Samuolienė G, Sirtautas R, Novičkovas A, Dabašinskas L, Miliauskienė J, Vaštakaitė V, Bagdonavičienė A (2015) Effect of supplemental UV-A irradiation in solid-state lighting on the growth and phytochemical content of microgreens. Int Agrophys 29(1):13–22
Brestic M, Zivcak M (2013) PSII fluorescence techniques for measurement of drought and high temperature stress signal in crop plants: protocols and applications. In: Rout GR, Das AB (eds) Molecular stress physiology of plants. Springer, Berlin
Brosche M, Strid A (2003) Molecular events following perception of ultraviolet-B radiation by plants: UV-B induced signal transduction pathways and changes in gene expression. Physiol Plant 117:1–10
Bruckova K, Sytar O, Zivkak M, Barestic M, Lebeda A (2016) The effect of growth conditions on flavonols and anthocyanins accumulation in green and red lettuce. J Cent Eur Agric 17:986–997
Brzozowska E, Gałązka-Czarnecka I, Krala J (2014) Wpływrozproszonegopromieniowaniasłonecznego na wybranewłaściwościkiełkówkoniczynyczerwonej (Trifolium pratense L.). Effect of diffuse solar radiation on selected properties of red cover sprouts (Trifolium pratense L.). Żywność Nauka Technologia Jakość 6, 67–80 (in Polish)
Cai W, Gao X, Hu J, Chen L, Li X, Liu Y, Wang G (2016) UV-B radiation inhibits the photosynthetic electron transport chain in Chlamydomonas reinhardtii. Pak J Bot 48:2587–2593
Caldwell MM, Flint SD (1994) Stratospheric ozone reduction, solar UV-B radiation and terrestrial ecosystems. Clim Chang 28:375–394
Caldwell MM, Bornman JF, Ballaré CL, Flint SD, Kulandaivelu G (2007) Terrestrial ecosystems, increased solar ultraviolet radiation and interactions with other climatic change factors. Photochem Photobiol Sci 6:252–266
Casati P, Walbot V (2003) Gene expression profiling in response to ultraviolet radiation in maize genotypes with varying flavonoid content. Plant Physiol 132:1739–1754
Cerovic ZG, Samson G, Morales F, Tremblay N, Moya I (1999) Ultraviolet-induced fluorescence for plant monitoring: present state and prospects. Agronomie 19(7):543–578
Cerovic ZG, Ounis A, Cartelat A, Latouche G, Goulas Y, Meyer S, Moya I (2002) The use of chlorophyll fluorescence excitation spectra for the non-destructive in situ assessment of UV-absorbing compounds in leaves. Plant Cell Environ 25(12):1663–1676
Charles MT, Tano K, Asselin A, Arul J (2009) Physiological basis of UV-C induced resistance to Botrytis cinerea in tomato fruit. V. Constitutive defence enzymes and inducible pathogenesis-related proteins. Postharvest Biol Technol 51:414–424
Chaturvedi AK et al (2021) Exploring growth, physiological status, yield and water use efficiency of vegetables grown under wick method of irrigation. Plant Physiol Rep 26(1):64–73
Chen Z, Ouyang X, Chen X, Deng XW, Huang X (2019) Two E3 ligases antagonistically regulate the UV-B response in Arabidopsis. Proc Natl Acad Sci U S A 116:4722
Choudhary KK, Agrawal SB (2015) Effect of elevated ultraviolet-B on four tropical soybean cultivars: quantitative and qualitative aspects with special emphasis on gas exchange, chlorophyll fluorescence, biomass and yield. Acta Physiol Plant 37:31–43
Christie JM, Arvai AS, Baxter KJ, Heilmann M, Pratt AJ, O’Hara A, Kelly SM, Hothorn M, Smith BO, Hitomi K, Jenkins GI, Getzoff ED (2012) Plant UVR8 photoreceptor senses UV-B by tryptophan-mediated disruption of cross-dimer salt bridges. Science 335:1492–1496
Cicek N, Fedina I, Cakirlar H, Velitchkova M, Georgieva K (2012) The role of short term high temperature pretreatment on the UV-B tolerance of barley cultivars. Turk J Agric For 36:153–165
Çiçek N, Kalaji HM, Ekmekçi Y (2020) Probing the photosynthetic efficiency of some European and Anatolian scots pine populations under UV-B radiation using polyphasic chlorophyll a fluorescence transient. Photosynthetica 58(2):468–478
Coffey A, Jansen MAK (2019) Effects of natural solar UV-B radiation on three Arabidopsis accessions are strongly affected by seasonal weather conditions. Plant Physiol Biochem 134:64–72
Comont D, Winters A, Gomez LD, McQueen-Mason SJ, Gwynn-Jones D (2013) Latitudinal variation in ambient UV-B radiation is an important determinant of Lolium perenne forage production, quality, and digestibility. J Exp Bot 64:2193–2204
Correia CM, Areal ELV, Torres-Pereira MS, Torres-Pereira JMG (1998) Intraspecific variation in sensitivity to ultraviolet-B radiation in maize grown under field conditions. I. Growth and morphological aspects. Field Crops Res 59(2):81–89
Czégény G, Mátai A, Hideg É (2016) UV-B effects on leaves-oxidative stress and acclimation in controlled environments. Plant Sci 248:57–63
Davey MP, Susanti NI, Wargent JJ, Findlay JE, Paul QW, Paul ND, Jenkins GI (2012) The UV-B photoreceptor UVR8 promotes photosynthetic efficiency in Arabidopsis thaliana exposed to elevated levels of UV-B. Photosynth Res 114:121–131
Day TA, Vogelmann TC, Delucia EH (1992) Are some plant life forms more effective than others in screening out ultraviolet-B radiation? Oecologia 92:513–519
Day TA, Howells BW, Rice WJ (1994) Ultraviolet absorption and epidermal-transmittance spectra in foliage. Physiol Planta 92:207–218
de Oliveira IR, Crizel GR, Severo J, Renard CM, Chaves FC, Rombaldi CV (2016) Preharvest UV-C radiation influences physiological, biochemical, and transcriptional changes in strawberry cv. Camarosa. Plant Physiol Biochem 108:391–399
Dobrikova AG, Apostolova EL (2015) Damage and protection of the photosynthetic apparatus from UV-B radiation. II. Effect of quercetin at different pH. J Plant Physiol 184:98–105
Dohler G, Worrest RC, Biermann I, Zink J (1987) Photosynthetic14CO2fixation and (15N)-amino assimilation during UV-B radiation of Lithodesmium variable. Physiol Plant 70:511–515
Doughty JC, Hope AB (1973) Effects of ultraviolet radiation on the membranes of Characoralline. J Membr Biol 13:185–197
Doupis G, Bosabalidis AM, Patakas A (2016) Comparative effects of water deficit and enhanced UV-B radiation on photosynthetic capacity and leaf anatomy traits of two grapevine (Vitis vinifera L.) cultivars. Theor Exp Plant Physiol 28:131–141
Dzakovich MP, Ferruzzi MG, Mitchell CA (2016) Manipulating sensory and phytochemical profiles of green house tomatoes using environmentally relevant doses of ultraviolet radiation. J Agric Food Chem 64(36):6801–6808
Eisinger WR, Bogomolni RA, Taiz L (2003) Interactions between a blue green reversible photoreceptor and a separate UV-B receptor in stomatal guard cells. Am J Bot 90:1560–1566
Essemine J, Govindachary S, Ammar S, Bouzid S, Carpentier R (2012) Enhanced sensitivity of the photosynthetic apparatus to heat stress in digalactosyl-diacylglycerol deficient Arabidopsis. Environ Exp Bot 80:16–26
Fabón G, Monforte L, Tomás-Las-Heras R, Núñez-Olivera E, Martínez-Abaigar J (2012) Dynamic response of UV-absorbing compounds, quantum yield and the xanthophyll cycle to diel changes in UV-B and photosynthetic radiations in an aquatic liver wort. J Plant Physiol 169(1):20–26
Fagerberg WR, Bornman JF (2005) Modification of leaf cytology and anatomy in Brassica napus grown under above ambient levels of supplemental UV radiation. Photochem Photobiol Sci 4:275–279
Faseela P, Puthur JT (2018) The imprints of the high light and UV-B stresses in Oryza sativa L. ‘Kanchana’ seedlings are differentially modulated. J Photochem Photobiol B 178:551–559
Favory JJ, Stec A, Gruber H, Rizzini L, Oravecz A, Funk M, Albert A, Cloix C, Jenkins GI, Oakeley EJ, Seidlitz HK, Nagy F, Ulm R (2009) Interaction of COP1 and UVR8 regulates UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis. EMBO J 28:591–601
Fedina I, Hidema J, Velitchkova M, Georgieva K, Nedeva D (2010) UV-B induced stress responses in three rice cultivars. Biol Plant 54:571–574
Foyer CH, Lopez-Delgado H, Dat JF, Scott IM (1997) Hydrogen peroxide-and glutathione-associated mechanisms of acclimatory stress tolerance and signalling. Physiol Plant 100(2):241–254
Friso G, Spetea C, Giacometti GM, Vass I, Barbato R (1993) Degradation of photosystem II reaction center D1 protein induced by UV-B irradiation in isolated thylakoids. Identification and characterization of C- and N-terminal breakdown products. Biochim Biophys Acta 1184:78–84
Friso G, Barbato R, Giacometti GM, Barbar J (1994) Degradation of D2 protein due to UVB irradiation of the reaction centre of PSII. FEBS Lett 339:217–221
Friso G, Vass I, Spetea C, Barber J, Barbato R (1995) UV-B induced degradation of the D1 protein in isolated reaction centres of photosystem II. Biochim Biophys Acta 1231:41–46
Fujibe T, Watanabe K, Nakajima N, Ohashi Y, Mitsuhara I, Yamamoto KT, Takeuchi Y (2000) Accumulation of pathogenesis-related proteins in tobacco leaves irradiated with UV-B. J Plant Res 113:387–394
Gaberscik A, Voncina M, Trost T, Germ M, Bjorn LO (2002) Growth and production of buckwheat (Fagopyrum esculentum) treated with reduced, ambient and enhanced UV-B radiation. J Photochem Photobiol B 66:30–42
Gao W, Zheng Y, Slusser JR, Gordon M, Heisler GM (2003) Impact of enhanced ultraviolet-B irradiance on cotton growth, development, yield, and qualities under field conditions. Agric For Meteorol 120:241–248
Gerhardt KE, Wilson MI, Greenberg BM (2005) Ultraviolet wavelength dependence of photomorphological and photosynthetic responses in Brassica napus and Arabidopsis thaliana. Photochem Photobiol 81(5):1061–1068
Ghasemi S, Kumleh HH, Kordrostami M (2019) Changes in the expression of some genes involved in the biosynthesis of secondary metabolites in Cuminum cyminum L. under UV stress. Protoplasma 256:279–290
Ghozlen NB, Cerovic ZG, Germain C, Toutain S, Latouche G (2010) Non-destructive optical monitoring of grape maturation by proximal sensing. Sensors 10(11):10040–10068
Gitz DC, Britz SJ, Sullivan JH (2013) Effect of ambient UV-B on stomatal density, conductance and isotope discrimination in four field grown soybean [Glycine max (L.) Merr.] isolines. Am J Plant Sci 4:100–108
Gómez I, Pérez-Rodríguez E, Viñegla B, Figueroa FL, Karsten U (1998) Effects of solar radiation on photosynthesis, UV-absorbing compounds and enzyme activities of the green alga Dasycladus vermicularis from southern Spain. J Photochem Photobiol B Biol 47(1):46–57
González-Villagra J, Marjorie RD, Alberdi M et al (2020) Solar UV irradiation effects on photosynthetic performance, biochemical markers, and gene expression in high bush blueberry (Vaccinium corymbosum L.) cultivars. Sci Hortic 259:108816
Gould KS, Dudle DA, Neufeld HS (2010) Why some stems are red: cauline anthocyanins shield photosystem II against high light stress. J Exp Bot 61:2707–2717
Grace SC, Logan BA (2000) Energy dissipation and radical scavenging by the plant phenylpropanoid pathway. Philos Trans R Soc B Biol Sci 355:1499–1510
Greenberg BM, Gaba V, Canaani O, Malkin S, Mattoo AK, Edelmann M (1989) Separate photosensitizers mediate degradation of the 32 KD PSII reaction centre protein in the visible and UV spectral region. Proc Natl Acad Sci U S A 86:6617–6620
Groher T (2019) Suitability of non-destructive sensors for monitoring physiological and biochemical responses of tomato leaves and fruits to abiotic stresses
Guidi L, Mori S, Degl’Innocenti E, Pecchia S (2007) Effects of ozone exposure or fungal pathogen on white lupin leaves as determined by imaging of chlorophyll a fluorescence. Plant Physiol Biochem 45:851–857
Guidi L, Degl’Innocenti E, Remorini D, Biricolti S, Fini A, Ferrini F, Nicese FP, Tattini M (2011) The impact of UV-radiation on the physiology and biochemistry of Ligustrum vulgare exposed to different visible-light irradiance. Environ Exp Bot 70:88–95
Guo DP, Guo YP, Zhao JP, Hui L, Peng Y, Wang QM, Chen JS, Rao GZ (2005) Photosynthetic rate and chlorophyll fluorescence in leaves of stem mustard (Brassica juncea var. tsatsai) after turnip mosaic virus infection. Plant Sci 168:57–63
Gupta R, Bhadauriya P, Chauhan VS, Bisen PS (2008) Impact of UV-B radiation on thylakoid membrane and fatty acid profile of Spirulina platensis. Curr Microbiol 56:156–161
Guruprasad K, Bhattacharjee S, Kataria S, Yadav S, Tiwari A, Baroniya S, Rajiv A, Mohanty P (2007) Growth enhancement of soybean (Glycine max) upon exclusion of UV-B and UV-A components of solar radiation, characterization of photosynthetic parameters in leaves. Photosynth Res 94:299–306
Hansson O, Wydrzynski T (1990) Current perceptions of photosystem II. Photosynth Res 23:131–162
Harm W (1980) Biological effects of ultraviolet radiation. Cambridge University Press, Cambridge
He J, Huang LK, Whitecross M (1994) Chloroplast ultrastructure changes in Pisum sativum associated with supplementary ultraviolet (UV-B) radiation. Plant Cell Environ 17(6):771–775
He JM, Ma XG, Zhang Y, Sun TF, Xu FF, Chen YP, Liu X, Yue M (2013) Role and interrelationship of Ga protein, hydrogen peroxide, and nitric oxide in ultraviolet B-induced stomatal closure in Arabidopsis leaves. Plant Physiol 161:1570–1583
Hectors K, Prinsen E, Van Oevelen S, Guisez Y, Jansen M (2009) Interactions between two UV-B protective responses: morphogenesis and UV screening. Comp Biochem Physiol A Physiol 153A:S202
Heijde M, Ulm R (2012) UV-B photoreceptor-mediated signalling in plants. Trends Plant Sci 17:230–237
Heisler GM, Grant RH, Gao W (2003) Ultraviolet radiation and its impacts on agriculture and forests. Agric For Meteorol 120:3–7
Herman JR (2010) Global increase in UV irradiance during the past 30 years (1979–2008) estimated from satellite data. J Geophys Res Atmos 115(D4)
Hernandez I, Van Breusegem F (2010) Opinion on the possible role of flavonoids as energy escape valves: novel tools for nature’s Swiss army knife? Plant Sci 179:297–301
Hideg É, Strid Å (2017) The effects of UV-B on the biochemistry and metabolism in plants. In: UV-B radiation and plant life molecular biology to ecology, pp 90–110
Hideg E, Vass I (1996) UV-B induced free radical production in plant leaves and isolated thylakoid membranes. Plant Sci 115:251–260
Hideg E, Sass L, Barbato R, Vass I (1993) Inactivation of photosynthetic oxygen evolution by UVB radiation: a thermoluminescence study. Photosynth Res 38:455–462
Hideg E, Barta C, Kalai T, Vass I, Hideg K, Asada K (2002) Detection of singlet oxygen and superoxide with fluorescent sensors in leaves under stress by photoinhibition or UV radiation. Plant Cell Physiol 43:1154–1164
Hideg E, Jansen MAK, Strid A (2013) UV-B exposure, ROS, and stress: inseparable companions or loosely linked associates? Trends Plant Sci 18:107–115
Hoffmann AM, Noga G, Hunsche M (2015) High blue light improves acclimation and photosynthetic recovery of pepper plants exposed to UV stress. Environ Exp Bot 109:254–263
Hofmann RW, Jahufer MZZ (2011) Trade off between biomass and flavonoid accumulation in white clover reflects contrasting plant strategies. PLos One 6:e18949
Hollosy F (2002) Effects of ultraviolet radiation on plant cells. Micron 33:179–197
Holzinger A, Lutz C, Karsten U, Wiencke C (2004) The effect of ultraviolet radiation on ultrastructure and photosynthesis in the red macro algae Palmaria palmata and Odonthalia dentate from Arctic waters. Plant Biol 6:568–577
Ibrahim MM, Alsahli AA, Al-Ghamdi AA (2013) Cumulative abiotic stresses and their effect on the antioxidant defense system in two species of wheat, Triticum durum Desf and Triticum aestivum L. Arch Biol Sci 65:1423–1433
Ilić ZS, Fallik E (2017) Light quality manipulation improves vegetable quality at harvest and postharvest: a review. Environ Exp Bot 139:79–90
Inostroza-Blancheteau C, Acevedo P, Loyola R, Arce-Johnson P, Alberdi M, Reyes-Díaz M (2016) Short-term UV-B radiation affects photosynthetic performance and antioxidant gene expression in high bush blueberry leaves. Plant Physiol Biochem 107:301–309
Ivanov AG, Miskiewicz E, Clarke AK, Greenberg BM, Huner NPA (2000) Protection of photosystem I1 against UV-A and UV-B radiation in the cyanobacterium Plectonema boryanum: the role of growth temperature and growth irradiance. Photochem Photobiol 72:772–779
Iwanzik W, Tevini M, Dohnt G, Voss M, Weiss W, Graber P, Renger G (1983) Action of UV-B radiation on photosynthetic primary reactions in spinach chloroplasts. Physiol Plant 58:401–407
Jansen MAK, Depka B, Trebst A, Edelman M (1993) Engagement of specific sites in the plastoquinone niche regulates degradation of the D1 protein in photosystem II. J Biol Chem 268:21246–21252
Jansen MAK, Greenberg BM, Edelman M, Mattoo AK, Gaba V (1996) Accelerated degradation of the D2 protein of photosystem II under ultraviolet radiation. Photochem Photobiol 63:814–817
Jansen MAK, Gaba V, Greenberg BM (1998) Higher plants and UV-B radiation: balancing damage, repair and acclimation. Trends Plant Sci 3:131–135
Jansen M, van den Noort RE (2000) Ultraviolet-B radiation induces complex alterations in stomatal behaviour. Physiol Plant 110:189–194
Jenkins GI (2009) Signal transduction in responses to UV-B radiation. Annu Rev Plant Biol 60:407–431
Jiao Y, Lau OS, Deng XW (2007) Light-regulated transcriptional networks in higher plants. Nat Rev Genet 8:217–230
John CF, Morris K, Jordan BR, Thomas B, Mackerness SAH (2001) Ultraviolet exposure leads to up-regulation of senescence associated genes in Arabidopsis thaliana. J Exp Bot 52:367–1373
Jordan BR (1996) The effect of ultraviolet-B radiation on plants: a molecular perspective. Adv Bot Res 22:97–162
Jordan BR, James PE, Strid A, Anthony RG (1994) The effect of ultraviolet-B radiation on gene expression and pigment composition in etiolated and green pea leaf tissue UV-B induced changes are gene-specific and dependent upon the developmental stage. Plant Cell Environ 17:45–54
Kakani VG (2003) Field crop responses to ultraviolet-B radiation: a review. Agric For Meteorol 120:191–218
Kakani VG, Reddy KR, Zhao D, Sailaja K (2003) Field crop responses to ultraviolet-B radiation: a review. Agric For Meteorol 120:191–218
Kalaji HM, Schansker G, Ladle RJ, Goltsev V, Bosa K, Allakhverdiev SI, Brestič M, Bussotti F, Calatayud A, DąbrowskiElsheery NI, Ferroni L, Guidi L, Hogewoning SW, Jajoo A, Misra AN, Nebauer SG, Pancaldi S, Penella C, Poli DB, Pollastrini M, Romanowska-Duda ZB, Rutkowska B, Serôdio J, Suresh K, Szulc W, Tambussi E, Yanniccari M, Živčák M (2014) Frequently asked questions about chlorophyll fluorescence: practical issues. Photosynth Res 122:121–158
Kalaji MH, Goltsev VN, Żuk-Gołaszewska K, Brestic M, Živčák M (2017) Chlorophyll fluorescence: understanding crop performance–basics and applications. CRC Press, Boca Raton, p 222
Kamiya N, Shen JR (2003) Crystal structure of oxygen evolving photosystem II from Thermosynechococcus vulcanusat 3.7A° resolution. Proc Natl Acad Sci U S A 100:98–103
Kanungo M, Dubey A, Kataria S (2013) Solar UV-B and UV-A/B exclusion affects growth and antioxidant enzymes in cucumber and wheat. Indian J Plant Sci 2:63–72
Karvansara PR, Razavi SM (2019) Physiological and biochemical responses of sugar beet (Beta vulgaris L.) to ultraviolet-B radiation. Peer J 7:e6790
Kataria S, Guruprasad KN (2015) Exclusion of solar UV radiation improves photosynthetic performance and yield of wheat varieties. Plant Physiol Biochem 97:400–411
Kataria S, Jain K, Guruprasad KN (2007) UV-B induced changes in antioxidant enzymes and their isoforms in cucumber (Cucumis sativus L) cotyledons. Indian J Biochem Biophys 44:31–37
Kataria S, Dehariya P, Guruprasad KN, Pandey GP (2012) Effect of exclusion of ambient solar UV-A/B components on growth and antioxidant response of cotton (Gossypium hirsutum L.). Acta Biol Cracov Series Bot 54:47–53
Kataria S, Guruprasad KN, Ahuja S, Singh B (2013) Enhancement of growth, photosynthetic performance and yield by exclusion of ambient UV components in C3 and C4 plants. J Photochem Photobiol B 127:140–152
Kataria S, Jajoo A, Guruprasad KN (2014) Impact of increasing ultraviolet-B (UV-B) radiation on photosynthetic processes. J Photochem Photobiol B 137:55–66
Kataria S, Jain M, Kanungo M, Sharma S (2019) Wheat responses and tolerance to UV-B radiation: an overview. In: Hasanuzzaman M, Nahar K, Hossain M (eds) Wheat production in changing environments. Springer, Singapore, pp 175–196
Keiller DR, Holmes MG (2001) Effects of long-term exposure to elevated UV-B radiation on the photosynthetic performance of five broad-leaved tree species. Photosynth Res 67:229–240
Khudyakova AY, Kreslavski VD, Shmarev AN et al (2019) Impact of UV-B radiation on the photosystem II activity, pro-/antioxidant balance and expression of light-activated genes in Arabidopsis thaliana hy4 mutants grown under light of different spectral composition. J Photochem Photobiol B 194:14–20
Kim BC, Tennessen DJ, Last RL (1998) UV-B induced photomorphogenesis in Arabidopsis thaliana. Plant J 15:667–674
Kliebenstein DJ, Lim JE, Landry LG, Last RL (2002) Arabidopsis UVR8 regulates ultraviolet-B signal transduction and tolerance and contains sequence similarity to human regulator of chromatin condensation 1. Plant Physiol 130:234–243
Kong L, Wang F, Si J, Feng B, Zhang B, Li S, Wang Z (2014) Increasing in ROS levels and callose deposition in peduncle vascular bundles of wheat (Triticum aestivum L) grown under nitrogen deficiency. J Plant Interact 8:109–116
Kotilainen T, Lindfors A, Tegelberg R, Aphalo PJ (2011) How realistically does outdoor UV-B supplementation with lamps reflect ozone depletion: an assessment of enhancement errors. Photochem Photobiol 87:174–183
Kulandaivelu G, Noorudeen AM (1983) Comparative study of the action of ultraviolet-C and ultraviolet-B radiation on photosynthetic electron transport. Physiol Plant 58:389–394
Kumari R, Singh S, Agrawal SB (2009) Effects of supplemental ultraviolet-B radiation on growth and physiology of Acorus Calamus L. (Sweetflag). Acta Biol Cracov Ser Bot 51:19–27
Kytridis VP, Manetas Y (2006) Mesophyll versus epidermal anthocyanins as potential in vivo antioxidants: evidence linking the putative antioxidant role to the proximity of oxy-radical source. J Exp Bot 57:2203–2210
Lafontaine M, Freund M (2013) Improving optical fruit sorting by non-destructive determination of quality parameters affecting wine quality. In: Optical characterization of materials, pp 115–126
Láposi R, Veres S, Lakatos G, Olah V, Fieldsend A, Mészáros I (2009) Responses of leaf traits of European beech (Fagus sylvatica L.) saplings to supplemental UV-B radiation and UV-B exclusion. Agric For Meteorol 149(5):745–755
Lee SE, Yoo SY, Kim DY, Ko TS, Ok YS, Kim TW (2014) Proteomic evaluation of the response of soybean (Glycine max var Seoritae) leaves to UV-B. Plant Omics 7(3):123–132
León-Chan RG, López-Meyer M, Osuna-Enciso T, Sañudo-Barajas JA, Heredia JB, León-Félix J (2017) Low temperature and ultraviolet-B radiation affect chlorophyll content and induce the accumulation of UV-B-absorbing and antioxidant compounds in bell pepper (Capsicum annuum) plants. Environ Exp Bot 139:143–151
Leon-Felix J (2017) Low temperature and ultraviolet-B radiation affect chlorophyll content and induce the accumulation of UV-B-absorbing and antioxidant compounds in bell pepper (Capsicum annuum) plants. Environ Exp Bot 139:143–151
Lev-Yadun S, Gould KS (2007) What do red and yellow autumn leaves signal? Bot Rev 73:279–289
Li J, Qu-Lee TM, Raba R, Amundson RG, Last RL (1993) Arabidopsis flavonoid mutant are hypersensitive to UV-B irradiation. Plant Cell 5:171–179
Li Q, Liu X, Yue M, Zhang XF, Zhang RC (2011) Effects of physiological integration on photosynthetic efficiency of Trifolium repens in response to heterogeneous UV-B radiation. Photosynthetica 49(4):539–545
Li X, Zhang L, Li Y, Ma L, Bu N, Ma C (2012) Changes in photosynthesis, antioxidant enzymes and lipid peroxidation in soy bean seedlings exposed to UV-B radiation and/or Cd. Plant Soil 352(1):377–387
Lidon FC, Ramalho JC (2011) Impact of UV-B irradiation on photosynthetic performance and chloroplast membrane components in Oryza sativa L. J Photochem Photobiol B 104(3):457–466
Lidon FC, Teixeira M, Ramalho JC (2012) Decay of the chloroplast pool of ascorbate switches on the oxidative burst in UV-B irradiated rice. J Agron Crop Sci 198:130–144
Lingakumar K, Kulandaivelu G (1993) Changes induced by ultraviolet-B radiation in vegetative growth, foliar characteristics and photosynthetic activities in Vigna ungiculata. Aust J Plant Physiol 20:299–308
Liu L, Gitzi DC, McClure JW (1995) Effects of UV-B on flavonoids, ferulic acid, growth and photosynthesis in barley primary leaves. Physiol Plant 93:725–733
Liu X, Chi H, Yue M, Zhang X, Li W, Jia E (2012) The regulation of exogenous jasmonic acid on UV-B stress tolerance in wheat. J Plant Growth Regul 31(3):436–447
Liu FF, Chen HZ, Han R (2015) The effects of He-Ne laser and enhanced ultraviolet-B radiation on proliferating-cell nuclear antigen in wheat seedlings. Am J Plant Sci 6:1206–1214
Llorens L, Neugart S, Vandenbussche F, Castagna A (2020) Ultraviolet radiation: friend or foe for plants? Front Plant Sci 11:541
Lobo V, Patil A, Phatak A, Chandra N (2010) Free radicals, antioxidants and N. Mariz-Ponte et al. Sci Hortic 246(2019):777–784
Lois R, Buchanan BB (1994) Severe sensitivity to ultraviolet radiation in an Arabidopsis mutant deficient in flavonoids accumulation. II. Mechanisms of UV-resistance in Arabidopsis. Planta 194:504–509
Lopez ML, Palancar GG, Toselli BM (2009) Effect of different types of clouds on surface UV-B and total solar irradiance at southern mid-latitudes: CMF determinations at Cordoba, Argentina. Atmos Environ 43:3130–3136
Lu YB, Duan X, Zhang H, Korpelainen F, Li BC (2009) Intraspecific variation in drought response of Populus cathayana growth under ambient and enhanced UV-B radiation. Ann For Sci 66:613–624
Lv M, Su HY, Li ML, Yang DL, Yao RY, Li MF, Wei JH (2021) Effect of UV-B radiation on growth, flavonoid and podophyllotoxin accumulation, and related gene expression in Sinopodophyllum hexandrum. Plant Biol 23:202–209
Madronich S, Wagner M, Groth P (2011) Influence of tropospheric ozone control on exposure to ultraviolet radiation at the surface. Environ Sci Technol 45:6919–6923
Manetas Y, Petropoulou Y, Stamatakis K, Nikolopoulos D, Levizou E, Psaras G, Karabourniotis G (1997) Beneficial effects of enhanced UV-B radiation under field conditions: improvement of needle water relations and survival capacity of Pinus pinea L. seedlings during the dry Mediterranean summer. Plant Ecol 128:101–108
Mackerness S, Thomas B, Jordan BR (1997) The effect of supplementary ultraviolet-B radiation on mRNA transcripts, translation and stability of chloroplast proteins and pigment formation in Pisum sativum L. J Exp Bot 48:729–738
Mannuss A, Trapp I, Puchta H (2012) Gene regulation in response to DNA damage. Biochim Biophys Acta 1819:154–165
Mariz-Ponte N, Martins S, Gonçalves A, Correia CM, Ribeiro C, Dias MC, Santos C (2019) The potential use of the UV-A and UV-B to improve tomato quality and preference for consumers. Sci Hortic 246:777–784
Marwood CA, Greenberg BM (1996) Effect of supplementary UVB radiation on chlorophyll systems during chloroplast development in Spirodela oligarrhiza. J Photochem Photobiol 64:664–670
Mátai A, Nagy D, Hideg É (2019) UV-B strengthens antioxidant responses to drought in Nicotiana benthamiana leaves not only as supplementary irradiation but also as pre-treatment. Plant Physiol Biochem 134:9–19
Mathur S, Jajoo A (2015) Investigating deleterious effects of ultraviolet (UV) radiations on wheat by a quick method. Acta Physiol Plant 37(6):1–7
Mazza CA, Battista D, Zima AM, Szwarcberg-Bracchitta M, Giordano CV, Acevedo A, Scopel AL, Ballare CL (1999) The effects of solar ultraviolet-B radiation on the growth and yield of barley are accompanied by increased DNA damage and antioxidant responses. Plant Cell Environ 22:61–70
Melis A, Nemson JA, Harrison MA (1992) Damage to functional components and partial degradation of photosystem II reaction center proteins upon chloroplast exposure to ultraviolet-B radiation. Biochim Biophys Acta 1100:312–320
Mewis I, Schreiner M, ChauNhi N, Krumbein A, Ulrichs C, Lohse M, Zrenner R (2012) UV-B irradiation changes specifically the secondary metabolite profile in broccoli sprouts: induced signaling overlaps with defense response to biotic stressors. Plant Cell Physiol 53:1546–1560
Middleton EM, Teramura AH (1993) The role of flavonol glycosides and carotenoids in protecting soybean from ultraviolet-B damage. Plant Physiol 103(3):741–752
Mishra V, Srivastava G, Prasad SM, Abraham G (2003) Growth, photosynthetic pigments and photosynthetic activity during seedling stage of cowpea (Vigna unguiculata) in response to UV-B and dimethoate. Pestic Biochem Physiol 92:30–37
Mishra V, Srivastava G, Prasad SM, Abraham G (2008) Growth, photosynthetic pigments and photosynthetic activity during seedling stage of cowpea (Vigna unguiculata) in response to UV-B and dimethoate. Pestic Biochem Physiol 92:30–37
Mishra AK, Rai R, Agrawal SB (2013) Individual and interactive effects of elevated carbon dioxide and ozone on tropical wheat (Triticum aestivum L.) cultivars with special emphasis on ROS generation and activation of antioxidant defense system. Indian J Biochem Biophys 50:139–149
Mosadegh H, Trivellini A, Maggini R, Ferrante A, Incrocci L, Mensuali A (2021) In-vivo in-vitro screening of Ocimum basilicum L. ecotypes with differential UV-B radiation sensitivity. Horticulturae 7(5):101
Moussa HR, Khodary SDK (2008) Changes in growth and 14CO2 fixation of Hordeum vulgare and Phaseolus vulgaris induced by UV-B radiation. J Agric Soc Sci 4:59–64
Müller K, Engesser R, Schulz S, Steinberg T, Tomakidi P, Weber CC, Ulm R, Timmer J, Zurbriggen MD, Weber W (2013a) Multi-chromatic control of mammalian gene expression and signaling. Nucleic Acids Res 41(12):e124. https://doi.org/10.1093/nar/gkt340
Müller V, Albert A, Winkler JB, Lankes C, Noga G, Hunsche M (2013b) Ecologically relevant UV-B dose combined with high PAR intensity distinctly affect plant growth and accumulation of secondary metabolites in leaves of Centella asiatica L. Urban. J Photochem Photobiol B 127:161–169
Murphy TM, Hurrel HC, Sasaki TL (1985) Wavelength dependence of ultraviolet radiation-induced mortality and K+ efflux in cultured cells of Rosa damascena. Photochem Photobiol 42:281–286
Musil CF, Wand SJE (1994) Differential stimulation of an arid-environment winter ephemeral Dimorphotheca pluvialis (L.) Moench by ultraviolet-B radiation under nutrient limitation. Plant Cell Environ 17:245–255
Muzafarov EN, Kreslavsky VD, Nazarova GN (1995) Light and hormonal regulation of photosynthesis and plants growth. ONTI, Pushchino, 124 p
Nagel LM, Bassman JH, Edwards GE, Robberecht R, Franceshi VR (1998) Leaf anatomical changes in Populus trichocarpa, Quercus rubra, Pseudotsuga menziesii and Pinus ponderosa exposed to enhanced ultraviolet-B radiation. Physiol Plant 104:385–396
Neugart S, Schreiner M (2018) UVB and UVA as eustressors in horticultural and agricultural crops. Sci Hortic 234:370–381
Nogues S, Baker NR (1995) Evaluation of the role of damage to photosystem II in the inhibition of CO2 assimilation in pea leaves on exposure to UV-B radiation. Plant Cell Environ 18:781–787
Nogues S, Allen DJ, Morison JI, Baker NR (1998) Ultraviolet-B radiation effects on water relations, leaf development, and photosynthesis in droughted pea plants. Plant Physiol 117:173–181
Nogues S, Allen DJ, Morison JIL, Baker NR (1999) Characterization of stomatal closure caused by ultraviolet-B radiation. J Plant Physiol 121:489–496
Pal M, Zaidi PH, Voleti SR, Raj A (2006) Solar UV-B exclusion effect on growth and photosynthetic characteristics of wheat and pea. J New Seeds 8:19–34
Pan XL, Zhang DY, Li L (2011) Responses of photosystem II of white elm to UV-B radiation monitored by OJIP fluorescence transients. Russ J Plant Physiol 58(5):864–870
Panagopoulos L, Bornman JF, Bjorn LO (1990) Effects of ultraviolet radiation and visible light on growth, fluorescence induction, ultra weak luminescence and peroxidise activity in sugar beet plants. J Photochem Photobiol 8:73–87
Petroluleas V (2002) The mechanism of UV-A radiation induced inhibition of photosystem II electron transport studied by EPR and chlorophyll fluorescence. Biochemistry 41:10200–10208
Pfundel EE, Pan RS, Dilley RA (1992) Inhibition of violax anthinde epoxidation by ultraviolet-B radiation in isolated chloroplasts and intact leaves. Plant Physiol 98(4):1372–1380
Pinelli P, Romani A, Fierini E et al (2013) Characterization of the polyphenol content in the kiwifruit (Actinidia deliciosa) exocarp for the calibration of a fruit sorting optical sensor. Phytochem Anal 24(5):460–466
Piri E, Babaeian M, Tavassoli A, Esmaeilian Y (2011) Effects of UV irradiation on plants. Afr J Microbiol Res 5:1710–1716
Pollastrini M, DiStefano V, Ferretti M, Agati G, Grifoni D, Zipoli G, Orlandini S, Bussotti F (2011) Influence of different light intensity regimes on leaf features of Vitis vinifera L. in ultraviolet radiation filtered condition. Environ Exp Bot 73:108–115
Poulson ME, Boeger MRT, Donahue RA (2006) Response of photosynthesis to highlight and drought for Arabidopsis thaliana grown under a UV-B enhanced light regime. Photosynth Res 90:79–90
Qi Y, Heisler GM, Gao W, Vogelmann TC, Bai S (2010) Characteristics of UV-B radiation tolerance in broad leaf trees in southern USA. In: Gao W, Schmoldt DL, Slusser JR (eds) UV radiation in global climate change. Measurements, modeling and effects on ecosystems. Springer, Berlin
Randriamanana TR, Nissinen K, Moilanen J, Nybakken L, Julkunen-Tiitio R (2015) Long-term UV-B and temperature enhancements suggest that females of Salix myrsinifolia plants are more tolerant to UV-B than males. Environ Exp Bot 109:296–305
Ranjbarfordoei A, Samson R, Van Damme P (2011) Photosynthesis performance in sweet almond [Prunus dulcis (Mill) D. Webb] exposed to supplemental UV-B radiation. Photosynthetica 49:107–111
Ravindran KC, Indrajith A, Pratheesh PV, Sanjiviraja K, Balakrishnan V (2010) Effect of ultraviolet-B radiation on biochemical and antioxidant defence system in Indigofera tinctoria L. seedlings. Int J Eng Sci 2:226–232
Reddy KR, Singh SK, Koti S, Kakani VG, Zhao D, Gao W, Reddy VR (2013) Quantifying the effects of corn growth and physiological responses to ultraviolet-B radiation for modeling. Agron J 105:1367–1377
Renger G, Voss M, Graber P, Schulze A (1986) Effects of UV radiation on different partial reactions of the primary processes of photosynthesis. In: Worrest RC, Caldwell MM (eds) Stratospheric ozone depletion, solar UV radiation and plant life. Springer, Berlin, pp 171–184
Renger G, Volker M, Eckert HJ, Fromme R, Hohm-Viet S, Graber P (1989) On the mechanism of photosystem II deterioration by UV-B radiation. Photochem Photobiol 49:97–105
Romanatti PV, Rocha GA, Júnior VV, SantosFilho PR, de Souza TC, Pereira FJ, Polo M (2019) Limitation to photosynthesis in leaves of egg plant under UVB according to anatomical changes and alterations on the antioxidant system. Sci Hortic 249:449–454
Ruhland CT, Xiong FS, Clark WD, Day TA (2005) The influence of ultraviolet radiation on growth, hydroxy cinnamic acids and flavonoids of Deschampsia antarctica during spring time ozone depletion in Antarctica. Photochem Photobiol 81:1086–1093
Ryel RJ, Barnes PW, Beyschlag W, Catdwell MM, Flint SD (1990) Plant competition for light analyzed with a multispecies canopy model. I Model development and influence of enhanced UV-B conditions on photosynthesis in mixed wheat and wild oat canopies. Oecologia 82:304–310
Saile-Mark M, Tevini M (1997) Effects of solar UV-B radiation on growth, flowering and yield of central and southern European bush bean cultivars (Phaseolus vulgaris L.). Plant Ecol 128:115–125
Sandmann G (2019) Antioxidant protection from UV-and light-stress related to carotenoid structures. Antioxidants 8(7):219
Savitch LV, Pocock T, Krol M, Wilson KE, Greenberg BM, Huner NPA (2001) Effects of growth under UV-A radiation on CO2 assimilation, carbon partitioning, PSII photochemistry and resistance to UV-B radiation in Brassica napus cv. Topas. Aust J Plant Physiol 28:203–212
Schaller J, Brackhage C, Bäucker E, Dudel E (2013) UV-screening of grasses by plant silica layer? J Biosci 38:413–416
Schmidt AM, Ormrod DP, Livingston NJ, Misra S (2000) The interaction of ultraviolet-B radiation and water deficit in two Arabidopsis thaliana genotypes. Ann Bot 85:571–575
Schreiner M, Mewis I, Huyskens-Keil S, Jansen MAK, Zrenner R, Winkler JB, O’Brien N, Krumbein A (2012) UV-B-induced secondary plant metabolites—potential benefits for plant and human health. Crit Rev Plant Sci 31:229–240
Schreiner M, Neugart S, Wiesner M, Baldermann S (2017) Chapter 4. Plant defence against UV-B. In: Jordan BR (ed) The role of UV-B radiation in plant growth and development. CABI Press, Oxford
Schultze M, Bilger W (2019) Acclimation of Arabidopsis thaliana to low temperature protects against damage of photosystem II caused by exposure to UV-B radiation at 9°C. Plant Physiol Biochem 134:73–80
Searles PS, Flint SD, Caldwell MM (2001) A meta-analysis of plant field studies simulating stratospheric ozone depletion. Oecologia 127:1–10
Seckmeyer G, Pissulla D, Glandorf M, Henriques D, Johnsen B, Webb A et al (2008) Variability of UV irradiance in Europe. Photochem Photobiol 84:172–179
Segui JA, Maire V, Gabashvili IS, Fragata M (2000) Oxygen evolution loss and structural transitions in photosystem II induced by low intensity UV-B radiation of 280 nm wavelength. J Photochem Photobiol B 56:39–47
Selvakumar V (2008) Ultraviolet-B radiation (280–315 nm) invoked antioxidant defense systems in Vigna unguiculata (L.) Walp and Crotalaria juncea (L). Photosynthetica 46:98–106
Severo J, de Oliveira IR, Bott R, Le Bourvellec C, Renard CM, Page D, Chaves FC, Rombaldi CV (2016) Preharvest UV-C radiation impacts strawberry metabolite content and volatile organic compound production. LWT Food Sci Technol 85:390–393
Sharma PK, Anand P, Sankhalkar S (1998) Oxidative damage and changes in activities of antioxidant enzymes in wheat seedlings exposed to ultraviolet-B radiation. Curr Sci 75:359–366
Singh M, Singh S, Agrawal SB (2012) Intraspecific responses of six cultivars of wheat (Triticum aestivum L) to supplemental ultraviolet-B radiation under field conditions. Acta Physiol Plant 34:65–74
Singh VP, Kumar J, Singh S, Prasad SM (2014) Dimethoate modifies enhanced UV-B effects on growth, photosynthesis and oxidative stress in mung bean (Vigna radiata L) seedlings: implication of salicylic acid. Pestic Biochem Physiol 116:13–23
Singh A, Bashri G, Prasad SM (2017) Major influence on phytochrome and photosynthetic machinery under UV-B exposure. In: Singh VP, Singh S, Prasad SM, Parihar P (eds) UV-B radiation: from environmental stressor to regulator of plant growth. Wiley-Blackwell, West Sussex, pp 123–142
Skórska E, Lewandowski R (2003) Porównaniereakcjiroślintrzechodmianowsa na promieniowanie UV-B. Comparison of reactions of three oat varieties to UV-B radiation. Biul IHAR 229, 199–204. (in Polish)
Snell KRS, Kokubun T, Griffiths H, Convey P, Hodgson DA, Newsham KK (2009) Quantifying the metabolic cost to an Antarctic liver wort of responding to an abrupt increase in UVB radiation exposure. Glob Chang Biol 15:2563–2573
Solovchenko A, Schmitz-Eiberger M (2003) Significance of skin flavonoids for UV-B protection in apple fruits. J Exp Bot 54:1977–1984
Spetea C, Hideg E, Vass I (1996) The quinone electron acceptors are not the main sensitizers of UV-B induced protein damage in isolated photosystem II reaction centre and core complexes. Plant Sci 115:207–215
Šprtová M, Nedbal U, Marek MV (2000) Effect of enhanced UV-B radiation on chlorophyll a fluorescence parameters in Norway spruce needles. J Plant Physiol 156(2):234–241
Srivastava PK, Singh VP, Prasad SM (2014) Low and high doses of UV-B differentially modulate chloropyrifos-induced alterations in nitrogen metabolism of cyanobacteria. Ecotoxicol Environ Saf 107:291–299
Strasser BJ (1997) Donor side capacity of photosystem II probed by chlorophyll a fluorescence transients. Photosynth Res 52(2):147–155
Strasser RJ, Srivastava A, Govindjee (1995) Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria. Photochem Photobiol 61:32–42
Strasser RJ, Tsimilli-Michael M, Srivastava A (2004) Analysis of the chlorophyll a fluorescence transient. In: Papageorgiou GC, Govindjee (eds) Chlorophyll a fluorescence: a signature of photosynthesis. Advances in photosynthesis and respiration. Springer, Dordrecht, pp 321–362
Strid A, Hideg E (2017) Chapter 7. The effects of UV-B on the biochemistry and metabolism of plants. In: Jordan BR (ed) The role of UV-B radiation in plant growth and development. CABI Press, Oxford
Strid Å, Porra RJ (1992) Alterations in pigment content in leaves of Pisum sativum after exposure to supplementary UV-B. Plant Cell Physiol 33(7):1015–1023
Strid A, Chow WS, Anderson JM (1994) UVB damage and protection at the molecular level. Photosynth Res 39:475–489
Svensson B, Vass I, Cedergren E, Styring S (1990) Structure of donor-side components in photosystem II predicted by computer modelling. EMBO J 9:2051–2059
Sytar O, Bruckova K, Hunkova E, Zivcak M, Konate K, Brestic M (2015) The application of multiplex fluorimetric sensor for the analysis of flavonoids content in the medicinal herbs family Asteraceae, Lamiaceae, Rosaceae. Biol Res 48(1):1–9
Sytar O, Zivcak M, Brestic M (2016) Noninvasive methods to support metabolomic studies targeted at plant phenolics for food and medicinal use. In: Plant omics: trends and applications. Springer, Cham, pp 407–443
Sytar O, Zivcak M, Bruckova K, Brestic M, Hemmerich I, Rauh C, Simko I (2018) Shift in accumulation of flavonoids and phenolic acids in lettuce attributable to changes in ultraviolet radiation and temperature. Sci Hortic 239:193–204
Sytar O, Zivcak M, Neugart S, Brestic M (2020) Assessment of hyperspectral indicators related to the content of phenolic compounds and multispectral fluorescence records in chicory leaves exposed to various light environments. Plant Physiol Biochem 154:429–438
Szilard A, Sass DZ, Vass I (2007) The sensitivity of photosystem II to damage by UV-B radiation depends on the oxidation state of the water-splitting complex. Biochim Biophys Acta 1767:876–882
Szwarc W, Skórska E (2007) The influence of UV-B radiation on some physiological features of soybean cv. Augusta plants. Biuletyn IHAR 245:215–221
Takeuchi A, Yamaguchi T, Hidema J, Strid A, Kumagai T (2002) Changes in synthesis and degradation of RuBisCO and LHCII with leafage in rice (Oryza sativa L.) growing under supplementary UV-B radiation. Plant Cell Environ 25:695–706
Tandori J, Mate Z, Vass I, Maroti P (1996) The reaction centre of the purple bacterium Rhodopseudomonas sphaeroides R-26 is highly resistant against UV-B radiation. Photosynth Res 50:171–179
Teramura AH, Ziska LH (1996) Ultraviolet-B radiation and photosynthesis. In: Baker NR (ed) Photosynthesis and the environment. Kulwer Academic Publishers, Dordrecht, pp 435–450
Teramura AH, Ziska LH, Sztein AE (1991) Changes in growth and photosynthetic capacity of rice with increased UV-B radiation. Physiol Plant 83:373–380
Tevini M, Iwanzik W, Thoma U (1981) Some effects of enhanced UV-B irradiation on the growth and composition of plants. Planta 153(4):388–394
Tevini M, Grusemann P, Fieser G (1988) Assessment of UV-B stress by chlorophyll fluorescence analysis. In: Lichtenthaler HK (ed) Applications of chlorophyll fluorescence. Kluwer Academic Publishers, Dordrecht, pp 229–238
Tevini M, Braun J, Fieser F (1991) The protective function of the epidermal layer of rye seedlings against ultraviolet-B radiation. Photochem Photobiol 53:329–333
Thiel S, Dohring T, Kofferlein M, Kosak A, Martin P, Seidlitz HK (1996) A phytotron for plant stress research: how far can artificial lighting compare to natural sunlight? J Plant Physiol 148:456–463
Tian X, Lei Y (2007) Physiological responses of wheat seedlings to drought and UV-B radiation effect of exogenous sodium nitroprusside application. Russ J Plant Physiol 54:676–682
Tilbrook K, Arongaus AB, Binkert M, Heijde M, Yin R, Ulm R (2013) The UVR8 UV-B photoreceptor: perception, signaling and response. In: The Arabidopsis. American Society of Plant Biologists, 11, e0164
Tossi V, Lamattina L, Jenkins GI, Cassia RO (2014) Ultraviolet-B-induced stomatal closure in Arabidopsisis regulated by the UV RESISTANCE LOCUS8 photoreceptor in a nitric oxide-dependent mechanism. Plant Physiol 164:2220–2230
Tossi VE, Regalado JJ, Iannicelli J, Laino LE, Burrieza HP, Escandón AS, Pitta-Álvarez SI (2019) Beyond Arabidopsis: differential UV-B response mediated by UVR8 in diverse species. Front Plant Sci 10:780
Trebst A, Depka B (1990) Degradation of the D-1 protein subunit of photosystem II in isolated thylakoids by UV light. Zeitschriftfur Naturforschung 45c:765–771
Treutter D (2005) Significance of flavonoids in plant resistance and enhancement of their biosynthesis. Plant Biol 7:581–591
Tripathi DK, Singh S, Singh VP, Prasad SM, Dubey NK, Chauhan DK (2016) Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol Biochem 110:70–81
Tuccio L, Cavigli L, Rossi F, Dichala O, Katsogiannos F, Kalfas I, Agati G (2020) Fluorescence-sensor mapping for the in vineyard non-destructive assessment of crimson seedless table grape quality. Sensors 20(4):983
Turcsanyi E, Vass I (2000) Inhibition of photosynthetic electron transport by UV-A radiation targets the photosystem II complex. Photochem Photobiol 72:513–520
Ulm R, Baumann A, Oravecz A, Mate Z, Adam E, Oakeley EJ, Schafer E, Nagy F (2004) Genome-wide analysis of gene expression reveals function of the bZIP transcription factor HY5 in the UV-B response of Arabidopsis. Proc Natl Acad Sci U S A 101:1397–1402
Ulm R, Nagy F (2005) Signalling and gene regulation in response to ultraviolet light. Curr Opin Plant Biol 8:477–482
Urban L, Charles F, de Miranda MRA, Aarrouf J (2016) Understanding the physiological effects of UV-C light and exploiting its agronomic potential before and after harvest. Plant Physiol Biochem 105:1–11
Valkama E, Kivimaenpaa M, Hartikainen H, Wulff A (2003) The combined effects of enhanced UV-B radiation and selenium on growth, chlorophyll fluorescence and ultrastructure in strawberry (Fragaria × ananassa) and barley (Hordeum vulgare) treated in the field. Agric For Meteorol 120:267–278
van Rensen JJ, Vredenberg WJ, Rodrigues GC (2007) Time sequence of the damage to the acceptor and donor sides of photosystem II by UV-B radiation as evaluated by chlorophyll a fluorescence. Photosynth Res 94(2):291–297
Varga B, Janda T, Laszlo E, Veisz O (2012) Influence of abiotic stresses on the antioxidant enzyme activity of cereals. Acta Physiol Plant 34:849–858
Vass I (1997) Adverse effects of UV-B light on the structure and function of the photosynthetic apparatus. In: Pessarakli M (ed) Handbook of photosynthesis. Marcel Dekker, New York, pp 931–949
Vass I (2012) Molecular mechanisms of photodamage in the photosystem II complex. Biochim Biophys Acta 1817:209–217
Vass I, Andras S, Sicora C (2005) Adverse effects of UV-B light on the structure and function of the photosynthetic apparatus. Taylor Francis Group, LLC
Vidović M, Morina F, Milić S, Zechmann B, Albert A, Winkler JB, Veljović Jovanović S (2015) UV-B component of sunlight stimulates photosynthesis and flavonoid accumulation in variegated Plectranthus coleoides leaves depending on background light. Plant Cell Environ 38:968–979
Vidović M, Morina F, Veljović Jovanović S (2017) Stimulation of various phenolics in plants under ambient UV-B radiation. In: UV-B radiation: from environmental stressor to regulator of plant growth. Wiley-Blackwell, Chichester
Vincent WF, Roy S (1993) Solar ultraviolet radiation and aquatic primary production: damage, protection and recovery. Environ Rev 1:1–12
Vu CV, Allen LH, Garrard LA (1982) Effects of supplemental UV-B radiation on primary photosynthetic carboxylating enzymes and soluble proteins in leaves of C3 and C4 crop plants. Physiol Plant 55:11–16
Vu CV, Allen LH, Garrard LA (1983) Effects of enhanced UV-B radiation (280–320 nm) on ribulose-1, 5-bisphosphate carboxylase in pea and soybean. Environ Exp Bot 24:141–143
Vyšniauskienė R, Rančelienė V (2014) Effect of UV-B radiation on growth and antioxidative enzymes activity in Lithuanian potato (Solanum tuberosum L.) cultivars. Zemdirbyste-Agriculture 101:51–56
Wang ZY, Li FM, Xiong YC, Xu BC (2008) Soil-water threshold range of chemical signals and drought tolerance was mediated by ROS homeostasis in winter wheat during progressive soil drying. J Plant Growth Regul 27:309–319
Wang G, Hao Z, Anken RH, Lu J, Liu Y (2010) Effects of UV-B radiation on photosynthesis activity of Wolffia arrhiza as probed by chlorophyll fluorescence transients. Adv Space Res 45:839–845
Wang Y, Yu G, Li K, Wu M, Ma J, Xu J, Chen G (2015) Responses of photosynthetic properties and antioxidant enzymes in high-yield rice flag leaves to supplemental UV-B radiation during senescence stage. Environ Sci Pollut Res 22(6):4695–4705
Wargent JJ (2017) Chapter 11. Turning UV-B photobiology into commercial reality. In: Jordan BR (ed) The role of UV-B radiation in plant growth and development. CABI Press, Oxford
Wargent JJ, Jordan BR (2013) From ozone depletion to agriculture: understanding the role of UV radiation in sustainable crop production. New Phytol 197:1058–1076
Wong CY, Teoh ML, Phang SM, Lim PE, Beardall J (2015) Interactive effects of temperature and UV radiation on photosynthesis of Chlorella strains from polar, temperature and tropical environments: differential impacts on damage and repair. PLoS One 10(10):e0139469
Wu D, Hu Q, Yan Z, Chen W, Yan C, Huang X, Zhang J, Yang P, Deng H, Wang J, Deng X, Shi Y (2012) Structural basis of ultraviolet-B perception by UVR8. Nature 484:214–219
Yadav S, Shrivastava AK, Agrawal C, Sen S, Chatterjee A, Rai S, Rai R, Singh S, Rai L (2017) Impact of UV-B exposure on phytochrome and photosynthetic machinery: from cyanobacteria to plants. In: Singh S, Prasad SM, Parhar P (eds) UV-B radiation: from environmental stressor to regulator of plant growth. Wiley, West Sussex, pp 259–277
Yadav A, Singh D, Lingwan M, Yadukrishnan P, Masakapalli SK, Datta S (2020) Light signaling and UV-B-mediated plant growth regulation. J Integr Plant Biol 62(9):1270–1292
Yamane Y, Shikanai T, Kashino Y, Koike H, Satoh K (2000) Reduction of QA in the dark: another cause of fluorescence Fo increases by high temperatures in higher plants. Photosynth Res 63:23–34
Yannarelli GG, Gallego SM, Tomaro ML (2006) Effect of UV-B radiation on the activity and isoforms of enzymes with peroxidase activity in sunflower cotyledons. Environ Exp Bot 56:174–181
Yin R, Ulm R (2017) How plants cope with UV-B: from perception to response. Curr Opin Plant Biol 37:42–48
Yoon HI, Kim D, Son JE (2020) Spatial and temporal bioactive compound contents and chlorophyll fluorescence of kale (Brassica oleracea L.) under UV-B exposure near harvest time in controlled environments. Photochem Photobiol 96(4):845–852
Yu GH, Li W, Yuan ZY, Cui HY, Lv CG, Gao ZP, Han B, Gong YZ, Chen GX (2013) The effects of enhanced UV-B radiation on photosynthetic and biochemical activities in super-high-yield hybrid rice Liangyou peijiu at the reproductive stage. Photosynthetica 51:33–44
Zavala JA, Mazza CA, Dillon FM, Chludil HD, Ballaré CL (2015) Soybean resistance to stinkbugs (Nezara viridula and Piezodorus guildinii) increases with exposure to solar UV-B radiation and correlates with isoflavonoid content in pods under field conditions. Plant Cell Environ 38:920–928
Zhang M, Chen G (2013) Influence of enhanced ultraviolet-B radiation on photosynthesis in flag leaves of a super-high-yield hybrid rice during senescence. Photosynthesis: research for food, fuel and future—15th international conference on photosynthesis. Zhejiang University Press, Hangzhou
Zhang J, Hu X, Henkow L, Jordan BR, Strid A (1994) The effects of ultraviolet-B radiation on the CF0F1-ATPase. Biochim Biophys Acta 1185:295–302
Zhao D, Reddy KR, Kakani VG, Mohammed AR, Read JJ, Gao W (2004) Leaf canopy photosynthetic characteristics of cotton (Gossypium hirsutum) under elevated CO2 concentration and UV-B radiation. J Plant Physiol 161:581–590
Ziska LH, Teramura AH, Sullivan JH (1992) Physiological sensitivity of plants along an elevational gradient to UV-B radiation. Am J Bot 79:863–871
Ziska LH, Teramura AH, Sullivan JH, McCoy A (1993) Influence of ultraviolet-B (UV-B) radiation on photosynthetic and growth characteristics in field-grown cassava (Manihot esculentum Crantz). Plant Cell Environ 16(1):73–79
Zivcak M, Brestic M, Balatova Z, Drevenakova P, Olsovska K, Kalaji MH, Allakhverdiev SI (2013) Photosynthetic electron transport and specific photoprotective responses in wheat leaves under drought stress. Photosynth Res 117:529–546
Zivcak M, Brückova K, Sytar O, Brestic M, Olsovska K, Allakhverdiev SI (2017) Lettuce flavonoids screening and phenotyping by chlorophyll fluorescence excitation ratio. Planta 245(6):1215–1229
Zu YG, Pang HH, Yu JH, Li DW, Wei XX, Gao YX, Tong L (2010) Responses in the morphology, physiology and biochemistry of Taxuschinensis var mairei grown under supplementary UV-B radiation. J Photochem Photobiol B 98:152–158
Zuk-Golaszewska K, Upadhyaya MK, Golaszewski J (2003) The effect of UV-B radiation on plant growth and development. Plant Soil Environ 49:135–140
Zvezdanovic J, Markovic DZ, Jovanovic SV, Radovanovic B (2013) UV-induced oxygen free radicals production in spinach thylakoids. Adv Technol 2:45–50
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Brestic, M. et al. (2022). Acclimation of Photosynthetic Apparatus to UV-B Radiation. In: Kataria, S., Singh, V.P. (eds) UV-B Radiation and Crop Growth. Plant Life and Environment Dynamics. Springer, Singapore. https://doi.org/10.1007/978-981-19-3620-3_11
Download citation
DOI: https://doi.org/10.1007/978-981-19-3620-3_11
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-19-3619-7
Online ISBN: 978-981-19-3620-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)