
Abstract
Potato is among the most important food crops round the world. Potato yield and quality is constrained by a myriad of insect pests and pathogens. Beyond doubt, the late blight of potato continues to be the most important disease of potato which can cause loses worth billions of dollars annually. Apart from the changing composition and intensity of the pests and diseases, the emergence of newer species/strains continues at an alarming rate. The impact of climate change is going to be profound on the crop as well as the pests and pathogens, of course the response is species specific. The risk of the spread of invasive organisms of quarantine significance has increased many folds due to international trade and exchange of germplasm. On the other hand, the potato pest and disease management has come a long way, and several innovative approaches are being adopted for diagnostics and detection, monitoring and forecasting and management of various pest and diseases. In this chapter, an outline of the current issues of the potato pest and disease management is given along with a discussion on sustainability of the management practices.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Pest management
- Quarantine
- Pesticide resistance
- Economic losses
- Emerging diseases
- Climate change
- Disease diagnosis
- Innovation
- Sustainability
1.1 Introduction
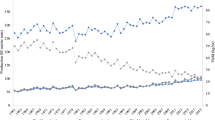
Potato (Solanum tuberosum) is regarded as one of the most important crops in addressing the challenge of food security, especially in developing countries. Presently, the crop is raised in 19 million hectares with 378 million tonnes global production. The world has witnessed notable increase in potato production in many countries during the last two decades because of area expansion and improvements in yield. Global statistics indicate that potato production is shifting towards developing countries, especially in Asia and Africa, and potato production in the developing countries has surpassed the developed world (FAOSTAT 2020). Worldwide, in 2018, China was the largest producer of potatoes, with India at second number (Fig. 1.1) (FAOSTAT 2020). More than a billion people consume potato on a regular basis, and it is a vital source of income for millions of farmers (Devaux et al. 2014). Though potato production has increased enormously since the 1960s in the developing world, but the sustainability of potato production globally is threatened by adverse abiotic conditions, pests and pathogens. Potato is prone to more than 100 pests and diseases including insects, nematodes, viruses, bacteria, oomycete and fungi which cause direct yield losses and decrease of farmer’s incomes by downgrading the quality of affected tubers. Due to climate change, global trade and increasing cropping intensity, pathogens are evolving at a faster rate and adapting to the new climate and hosts. As a consequence, health management is becoming more and more complex. Therefore, knowledge about the pathogens as well as factors influencing disease severity is needed to setup efficient control strategies.

Potato production from 2000 to 2019 (Source: FAOSTAT 2020)
The pest and disease management in potato is heavily reliant on the use of synthetic pesticides. Continued use of such plant protection products has led to development of resistance, secondary pest outbreaks and pest resurgence on frequent basis. Besides, the use of pesticides is generally harmful to mankind and the environment. Safer alternatives of pest and disease management are continuously being explored. Besides, the pest and disease management in potato is constrained due to the effects of climate change and the risk of invasive pests. In this chapter, current issues of pest and disease management in potato are discussed to lay open the platform for thorough discussions on the sustainable management of potato pets and diseases which are described throughout the book.
1.2 Pests and Diseases of Potato: Economic Importance
The potato is susceptible to many diseases. The oomycetes and the fungi ranging from moulds to the smuts and rusts perhaps constitute the most important biotic threats to potato. Potato is susceptible to numerous bacterial diseases, phytoplasmas, more than three dozen viruses of the yellow and mosaic groups and several parasitic nematodes. Diseases of potatoes include arguably the most historically significant crop disease, late blight, which is still the most important potato disease. Haverkort et al. (2009) estimated that the global costs and losses due to late blight may take 16% of all global potato production. At 100 €/t, the world potato production represents a value of €38 billion today. The 16% loss then represents an annual financial loss of €6.1 billion per annum today. After potato late blight, early blight represents one of the most important fungal diseases of potato today. Enormous yield losses are reported worldwide: 18–39% in the United States (Harrison and Venette 1970), 2–40% in parts of Europe (Leiminger and Hausladen 2014), 20–50% in South Africa (Van der Waals et al. 2001) and >20% in Australia (Horsfield et al. 2010). In India, yield loss to the tune of 79% is recorded under severe condition. An increasing emphasis on the cosmetic appearance of potatoes has recently brought hitherto non-significant diseases into prominence. Unless effective methods of control are practiced, some of the diseases have the potential to cause the total loss of a crop. The economic importance and losses caused by various fungal, bacterial, viral and nematode diseases in potato crops are described in detail in respective chapters.
Insect pests are among the major constraints affecting the yield and quality of produce in potato. Variable losses are estimated across locations, with global losses estimated on average at 16% (Oerke et al. 1994). Losses as high as 30% and 70% are recorded for various pests under different crop management regimes; complete crop losses or complete loss of quality seed crops is not a surprise if proper management practices are not adopted (Kroschel and Schaub 2013; Mujica and Kroschel 2013; Kroschel et al. 2020).
A large number of insect pests infest potato crops. The insect may either damage the tubers directly or feed on leaves or stems and transmit debilitating pathogens (Radcliffe 1994). The latter result in loss of seed potato quality with severe consequences for the seed chain at multiple levels. Insect pests are also assigned to classes as per convenience, namely, above-ground (indirect) and below-ground (direct) pests. Kroschel et al. (2020) described a total of 49 species of insect pests infesting potato crops in different parts of the world. Out of these, 6 major and 32 minor species are prevalent throughout the temperate, tropical and subtropical regions; 9 major species are prevalent in tropical and subtropical regions and 2 major species affect potato crops in the temperate regions. Although the composition of major pests varies with many other factors, the Colorado potato beetle, aphids, leafhoppers, the potato psyllid, potato tuber moth, whitefly and wireworms are considered the deadliest insect pests of potato (Kroschel et al. 2012).
Some of these pest species specialize on Solanaceae and have moved around the world along with the spread of potato cultivation. The Colorado potato beetle and potato tuber moth are the classic examples of this phenomenon. Other species are generalists with near cosmopolitan distribution like the aphids and mites. Most regions in the world are also affected by regionally important pests such as the beet leafhopper (Circulifer tenellus (Baker)) in the north-west USA and the 28 spotted ladybird (Henosepilachna vigintioctopunctata (Fabr.)) in China. On the other hand, aphids are the major concern in prominent potato production areas in Europe, and the Colorado potato beetle is of secondary importance. The reverse is true in most states of the United States and Canada. In India, aphids, whitefly (Bemisia tabaci), numerous species of leaf hoppers, white grubs and cutworms are major pests (Shah et al. 2020). Among the global pests of potato, aphids are among the most important. Aphids are sap feeding insects, but the major damage inflicted by aphids in potato crops is by transmission of numerous potato viruses. The resulting viral diseases lead to considerable yield reductions, limit the production of disease-free seed potatoes and cause a progressive degeneration of seed stocks. Worldwide, the most important concern for the production of quality seed potatoes is the aphid-transmitted Potyvirus, potato virus Y (PVY). Being a non-persistent virus that has been demonstrated of being transmitted by more than 60 species of aphids, PVY infection is the most difficult to contain and is the most actively researched problem in potato pest management (Gray et al. 2010; Karasev and Gray 2013).
Waters and Jensen (2014) proposed that the insect pest scenario for potatoes at a given location are related to the end-use markets (e.g. subsistence agriculture, table stock, processing, long-term storage etc.) and the level of industrialization and access to insecticides and other crop protection technology. For example, potato tuber worm can be a pest of life-threatening proportions for subsistence farmers using rustic storage for food preservation, whereas it is a pest controlled easily by one or two well-timed insecticide treatments for technologically advanced farms with refrigerated storage. Conversely, an aphid-related disease such as potato virus Y (PVY) may be a minor yield-reducing concern for subsistence farming, but represents a nearly intractable problem and the difference between profit and loss for technologically intensive farms.
1.2.1 Emerging Pests and Diseases
Many of the important pests and diseases affecting potato can be regarded as emerging due to the attributes: (1) increase in incidence, geographical distribution or host range; (2) change in pathogenesis; (3) have newly evolved or (4) have been discovered or newly described (Anderson et al. 2004). The primary reasons for the occurrence of emerging pests and diseases are related to increased trade and travel, intensified and expanded land use, changes in agricultural practices, planting of new varieties and extreme weather events linked to climate change (Fry et al. 2015). Though late blight (Phytophthora infestans) has historically been an important disease of potatoes, it is regarded as re-emerging disease due to regular emergence of novel strains of the pathogen with increasing virulence and appearance in new locations with surprising intensity (Fry et al. 2015). Multiple clonal lineages have been found across the globe that indicates the history of the displacement of lineages over time. Migrations of exotic clonal lineages contribute substantially to change in the population composition in most locations worldwide (Fry 2020). A complex population structure is observed in Europe with ~70% population dominated by a few widely disseminated clonal lineages (www.euroblight.net). Recently, declining trend in the combined frequency of the clones EU_13_A2, EU_6_A1 and EU_1_A1 and increasing trend in the frequency of EU_36_A2, EU_37_A2, EU_41_A2 and EU_43_A2 have been observed in Europe (Cooke et al. 2019). Migration of the pathogen followed by selection has been the major causes of population change. The epidemiological consequences of sexually reproducing populations have made late blight management more difficult due to early set of epidemics thereby demanding more fungicide applications (Hannukkala et al. 2007). The occurrence of fungicide resistance to fluazinam (EU_37_A2) has further aggravated its management. Since the pathogen population is continually evolving and novel clonal lineages with new traits are emerging, it is necessary to tailor management schedule to the local pathogen population.
In recent years with warm and dry summers, early blight has become widespread and turned into one of most important diseases after late blight. In the past, potato diseases caused by Alternaria species were described as early blight (caused by A. solani) and brown spot (caused by A. alternata) disease. Recent studies, however, showed that the Alternaria population on potato is much more elaborate than a two-species disease complex. Ten species of Alternaria have been implicated to cause foliar diseases of potato worldwide. These include A. solani, A. alternata, A. tenuissima, A. dumosa, A. arborescens, A. infectoria, A. grandis, A. interrupta, A. longipes and A. arbusti (Taheri et al. 2009; Ardestani et al. 2010; Rodrigues et al. 2010; Shoaib et al. 2014; Leiminger et al. 2015; Tymon et al. 2016).
Bacterial diseases are other important biotic constraints, especially in tropical and subtropical regions and in some warm temperate regions of the world. About seven bacterial diseases affect potato worldwide and cause severe damages especially on tubers. Bacterial wilt and black leg are considered the most important diseases, whereas potato ring rot, pink eye and common scab are the minor ones. Recently, Ralstonia species complex has been reclassified on the basis of whole genome comparisons into three distinct species: R. solanacearum (Phylotype II), R. pseudosolanacearum (Phylotypes I and III) and R. syzygii (Phylotype IV) (Safni et al. 2014; Prior et al. 2016). Phylotype I strains are regarded to be of Asian origin, Phylotype II strains are thought to be of South American origin, whereas Phylotype III appears to have evolved in Africa and Phylotype IV in Indonesia. The R. solanacearum species complex is widely designated as a quarantine organism in many countries in an effort to prevent its movement across geographical borders. Nevertheless, the PIIB1 strain has spread from its origin to many potato-growing areas worldwide, presumably with movement in trade of infected seed tubers (Elphinstone 2005).
Pectobacterium spp. are important bacterial potato pathogens and can be aggressive on tubers and stems causing wilting and eventually plant death. The Pectobacterium species most commonly found on potato include P. atrosepticum, P. brasiliense, P. carotovorum, P. odoriferum, P. parmentieri, P. peruviense, P. polaris and P. punjabense. Though there are some regional differences in the species distribution, some appear to be ubiquitous. For example, P. atrosepticum, P. brasiliense, P. parmentieri and P. carotovorum are found on multiple continents (Pérombelon and Kelman 1987; Duarte et al. 2004; Kim et al. 2009; Pitman et al. 2008, 2010; van der Merwe et al. 2010; De Boer et al. 2012; Ngadze et al. 2012; She et al. 2017; van der Wolf et al. 2017; Wang et al. 2017a, b, c). In Europe, P. atrosepticum has been the predominant species responsible for blackleg disease on potato, while P. carotovorum is often associated with soft rot in storage. P. brasiliense was originally identified as causing disease on potato in Brazil (Duarte et al. 2004) and has been common in the United States since 2001, as also P. parmentieri (Yap et al. 2004; Kim et al. 2009). This species was not known to cause disease on potato in Europe prior to 2012–2013 but has since increased greatly in its incidence in many European countries (de Werra et al. 2015) and is now recognized as an important pathogen in Africa as well (van der Merwe et al. 2010).
Dickeya species (formerly Erwinia chrysanthemi) have emerged as a new threat to potato production in Europe. Like Pectobacterium, Dickeya species have also a wide host range with global distribution (Samson et al. 2005). Out of eight Dickeya species, only D. dianthicola and D. solani are of concern to potato (Toth et al. 2011). In some cases, D. dianthicola replaced P. atrosepticum as the dominant blackleg pathogen (Parkinson et al. 2009; Toth et al. 2011). D. solani has been recognized as a new Dickeya pathogen on potato by several groups from 2004 through 2010 (Laurila et al. 2008; Parkinson et al. 2009; Slawiak et al. 2009) and spreading across Europe on seed tubers.
Zebra chip (ZC), a new and economically important disease of potato in the United States, Mexico, Central America and New Zealand, is caused by the bacterium “Candidatus Liberibacter solanacearum (Lso)” and transmitted to potato by the potato psyllid, Bactericera cockerelli. Although Lso is only spread in potato by B. cockerelli, it can also be found in other Bactericera species, suggesting that vector feeding preferences limit the species of vectors important for zebra chip and not Lso-vector interactions (Borges et al. 2017). These observations suggest that the bacterium can easily be introduced in many parts of the world along with its insect vectors.
Insect pests such as the potato tuber moth, Phthorimaea operculella (Zeller), and the leafminer fly, Liriomyza huidobrensis (Blanchard), have become invasive and occur today as serious pests in many tropical and subtropical regions. Around 2006, the tomato leafminer, Tuta absoluta Meyrick, although a minor pest in potato, was unintentionally introduced to Spain, from where it continued its devastating journey across Africa and into Asia and reached India within less than 10 years (Rahman et al. 2012; Caparros Megido et al. 2013; Sridhar et al. 2015; Kanle Satishchandra et al. 2019). As farmers had not been prepared and no control measures had been in place, the pest caused large production losses in tomato (Lycopersicon esculentum Mill.); under certain conditions, potato was more heavily infested as known from South America. The bud midge, Prodiplosis longifolia Gagne, currently with a restricted distribution in Florida and Virginia and South America (Colombia, Peru, and Ecuador) could become an invasive species supported by its very polyphagous feeding habit. Lately, the fall armyworm (Spodoptera frugiperda J. E. Smith) (Lepidoptera: Noctuidae) is threatening crops across globe with very fast rate of spread. Potato could be a potential host for the fall armyworm in tropical and subtropical potato crops, e.g. in India (Yu 1982; Sidana et al. 2018; Nagoshi et al. 2019).
Potato is a temperate crop; however, efforts at breeding day neutral and high temperature tolerance have enabled tropicalisation of the crops. This phenomenon exposed the potato crops to some of the insect pests which are mainly prevalent in the warmer regions located in tropics or the subtropics. The potato cultivation in India is a major example of this type of pest shift. More than 85% of potato production in India is realized from the tropical and subtropical regions where the sweetpotato whitefly, Bemisia tabaci, is a major pest of the potato now (Shah et al. 2021a). To worsen the situation, we saw the evolution of the potato-specific Begomovirus, tomato leaf curl New Delhi virus [potato] around two decades back (Usharani et al. 2004; Kumar et al. 2021). The whitefly-virus complex is the major concern for quality seed potato production in India (Shah et al. 2020, 2021a).
1.3 Effect of Climate Change on Occurrence and Distribution of Potato Pests and Diseases
Climate change is a reality, and it has impact on infection, reproduction, dispersal and survival between seasons and other critical stages in the life cycle of a pathogen. Studies have shown that, assuming global temperature rise by 2 °C, there will be lower risk of late blight in warmer areas (<22 °C) and higher risk in cooler areas (>13 °C) at global level. However, earlier onset of warm temperatures could result in an early appearance of late blight disease in temperate regions with the potential for more severe epidemics and increased number of fungicide applications needed for its control. Similar predictions have been made for Finland where for each 1 °C warming, late blight would occur 4–7 days earlier, and with the susceptibility period extended by 10–20 days (Kaukoranta 1996) resulting in 1–4 additional fungicide applications. In United Kingdom, hotter and drier summers are likely to reduce the importance of late blight, although earlier disease onset may act in opposite direction. An empirical climate-disease model has suggested that under the climate change scenario of 1 °C temperature increase with 30% reduction in precipitation in Germany, incidence of potato late blight will decrease to a mere 16% of its current level. Increase in both temperature and RH has added new dimension to late blight across the world. Under such a situation, P. infestans is likely to attack potato stems more often than foliage. In fact, in recent years it is more of “stem blight” than the foliar blight. This phase of the disease is more serious than the foliar stage as it affects the very crop plant. In India, Lahaul valley of HP, which was earlier free from late blight because of lack of precipitation, has now experienced attack of late blight due to occurrence of rainfall (Singh et al. 2013). Studies on the potential impact of climate change on late blight outbreak in Punjab and western Uttar Pradesh revealed that a higher number of sprays would be required in Punjab, whereas there would be no change or it is likely to be reduced in western Uttar Pradesh under future scenario (Dua et al. 2015). Shortest incubation period at higher temperature (28 °C) is on records for Phytophthora infestans (Becktell et al. 2005).
The effect of climate change on soilborne pathogens would vary from pathogen to pathogen. Black scurf and common scab diseases are favoured by moderate temperature and are likely to remain insulated from global warming in the near future. However, as the ambient temperatures are likely to increase by 1.4–5.8 °C by the end of the century, the severity of these two diseases may decrease substantially. Charcoal rot, which is favoured by high temperature and moisture, is currently endemic in eastern Uttar Pradesh, Bihar, Chhattisgarh and Madhya Pradesh, and the severity of this disease is likely to increase in these regions under global warming. Moreover, it is also expected that it may expand to other parts of north-central plains as well. Sclerotium wilt is restricted to plateau regions and is favoured by high temperature and moisture. With the increase in temperature due to global warming, the disease may also become prevalent in eastern Indo-Gangetic plains and may also enter into other areas like mid hills. Similarly, bacterial wilt may also advance to higher altitudes in hilly regions due to global warming, making them unfit for seed production.
Diseases caused by Synchytrium endobioticum (wart) and Spongospora subterranea (powdery scab) are favoured by low temperature and high soil moisture. Although wart spores can cause infection in the range of 10–28 °C with an optimum of 21 °C, there is hardly any infection beyond 23 °C. Therefore, warmer climates are likely to reduce wart infestation. Similarly, powdery scab infestation is also likely to be reduced with increase in temperature and reduction in rainfall under climate change scenario. The optimum temperature for powdery scab is 12 °C, and moisture requirement is 100%; the global warming may either lead to elimination of this disease or be pushed to higher altitudes making high hills (2500 m asl) free of powdery scab (Singh et al. 2013). The Pectobacterium species also differ in optimal and upper limits of growth temperatures. For example, P. atrosepticum and P. parmentieri die above 33 °C, while P. carotovorum and P. brasiliense can grow at temperatures up to 39 °C; hence the former two species may get eliminated, whereas the later may expand to new horizons (Charkowski 2015) under climate change scenario.
Among other phenomena, the impact of climate change on the crop and the associated insect pest complex is likely to be substantial (Hijmans 2003; Minhas et al. 2018; Raymundo et al. 2018; Rana et al. 2020; Shah et al. 2021b). It is being realized that insect species can respond to climate change in a multitude of ways, mainly in anticipation of a generally warmer and drier environment rich in CO2 predicted through climate change general circulation models (IPCC 2013). Insect species may shift their geographic distributions or phenology in an attempt to track changes in their optimal conditions. Within ectotherms and endotherms alike, there is substantial evidence of range shifts already occurring, particularly towards the poles where temperatures are increasing; with recent evidence also suggesting, this is the case for pest species (Bebber et al. 2013). There are several reported cases of phenological change with the environment in insect pests. Crop damage due to insect pests may increase due to higher number of generations in a season, i.e. increased voltinism. Based on a day-degree model, an increased number of suitable days for development could allow faster generation time and, therefore, an additional generation (or possibly even two) to develop within a growing season (Barton and Terblanche 2014).
Climatic response phenology models have been used to assess the effect of temperature increase under projected changes in global temperature for the year 2050 and beyond for a wide range of potato pests (as summarized by Kroschel et al. 2020): P. operculella (Kroschel et al. 2013, 2016); L. huidobrensis (Mujica et al. 2016); Guatemalan potato tuber moth, Tecia solanivora (Povolny) (Schaub et al. 2016); Andean potato tuber moth, Symmetrischema tangolias (Gyen) (Sporleder et al. 2016); the whiteflies, Bemisia tabaci (Gennadius); and Trialeurodes vaporariorum (Westwood) (Gamarra et al. 2016a, b). As mentioned above, the geographical range of most of these is likely to expand with increased damage potential. However, the response is likely to be species specific. The deadly leafminer fly, L. huidobrensis, will expand in range to temperate regions of Asia, North and South America and Europe, as well as into subtropical and tropical mountainous regions with a moderate increase of its establishment and damage potential (Mujica et al. 2016). Even smaller changes in temperature predicted for tropical regions compared to temperate regions will have stronger consequences on pest development due to already higher existing metabolism rates of organisms such as insects (Dillon et al. 2010).
1.4 Pests and Pathogens of Quarantine Significance
All the pests and pathogens are not present everywhere. However, chances of their introduction into new geographical areas are very high due to globalisation. Challenges like population growth, globalization, climate change, bioterrorism and changing agribusiness infrastructure hamper plant bio-security at the local, regional and global levels. It is important for each nation to develop a plant bio-security infrastructure that ensures a safe and constant supply of food, feed and fibre. It is equally important to develop an international framework for cooperation that maintains plant bio-security without compromising trade. The devastating effects resulting from diseases and pests introduced along with international movement of planting material, agricultural produce and products are well documented. The historical Irish famine of 1845, caused by late blight of potato introduced from Central America, coffee rust introduced in Sri Lanka in 1875, fluted scale on citrus, San Jose scale in apples, banana bunchy top, the dreaded Golden nematode infesting potatoes, wart pathogen and the noxious weed Lantana camara introduced in 1809 from Central America are glaring examples that clearly demonstrate that introduction and establishment of quarantine pests including weeds into new areas can severely damage the crop production and economy of a region/country. Movement of plants and plant products between bio-geographical zones by human activities is now generally accepted to be the primary mode of introduction of exotic pathogens and pests.
Potato is usually propagated through tubers, and this vegetative mode of propagation is beset with many problems; hence, there is continuous threat of their introduction into new areas with planting material. In most of the countries, the following pests and pathogens are regarded as quarantined viz., potato tuber nematode (Ditylenchus destructor), stem and bulb nematode (Ditylenchus dipsaci), potato cyst nematodes (Globodera rostochiensis, G. pallida), gangrene (Phoma exigua var. foeta), potato wart (Synchytrium endobioticum), potato smut (Thecaphora solani), bacterial ring rot (Clavibacter michiganensis ssp. sepedonicus), Colorado potato beetle (Leptinotarsa decemlineata), Andean potato weevil (Premnotrypes spp.) and viruses (Andean potato latent, Andean potato mottle, arracacha B virus, potato deforming mosaic, potato T, potato yellow dwarf, potato yellow vein, potato calico strain of tobacco ring spot virus, potato strains of tobacco streak virus and potato purple-top wilt and stolbur Phytoplasmas), which might establish in the country and hamper potato production programme. The Colorado potato beetle, Leptinotarsa decemlineata (Say), native to Mexico, has spread across most of the United States and was introduced into France in the 1920s from where it spread further reaching also parts of China (CABI 2017). Insect pests such as the potato tuber moth, Phthorimaea operculella (Zeller), and the leafminer fly, Liriomyza huidobrensis (Blanchard), with their centre of origin in the Andes have spread to and established in many tropical and subtropical regions.
1.5 Innovations in Disease Diagnostics
The conventional pathogen detection is totally dependent on morphological, serological and molecular based technologies, which are complicated, skill intensive and time consuming and require sophisticated lab facilities. However, with acceleration of computer technology and proliferation of smartphones coupled with software-based information and widespread access, the plant disease diagnosis and management can be deployed in an effective manner (Mohanty et al. 2016). An innovative artificial intelligence (AI)-based mobile phone app may provide an effective, easy-to-use diagnostic and low-cost solution. The recent advances in artificial intelligence coupled with increasing global smartphone penetration have paved the way for smartphone-assisted disease diagnosis (Johnson et al. 2021). There are android-based mobile apps available for download on smartphones which can diagnose a disease based on the pictures of the disease symptoms captured on the device. These apps have an expert-based algorithm of expert system which not only diagnoses the disease but gives advisories for the suitable management. One such plant disease diagnosis app “plantix” can detect more than 400 pathogens of 30 crops and has about ten million users worldwide (https://plantix.net).Interestingly, the app may further provide the advisories to minimize the disease outbreak in the next cropping season.
The first ever deep learning neural network platform in detection of potato diseases has been developed recently. Oppenheim et al. (2019) reported the detection of four economically important potato diseases viz., black scurf, common scab, silver scurf and black dot with more than 90% accuracy. The two devastating diseases of potato viz., early blight and late blight, were also being detected using this neural network. Recently, a cost-effective smartphone-based volatile organic compound (VOC) fingerprinting platform has been developed which allows non-invasive diagnosis of late blight caused by Phytophthora infestans (Li et al. 2019). This VOC-sensing platform is portable and easy to handle and can perform multiplex detection by classification of ten plant volatiles at a time. In fact, the symptomless plants having fungal infection may also be detected using this sensor-based platform. The approach will supplement the lab-based routine molecular detection and will become a viable additional method to help prevent major yield losses in potato.
E-nose technology has gained popularity for its applications in a field of human diagnostics, food quality and environmental safety. The technology is a quick and easily operated, rapid responding flexible tool to recognize signature gas samples (Cellini et al. 2017). The main operating principle relies in chemical interaction and electrical conductivity of sensors and resultant variations in the signature molecules. The e-nose basically comprises of an array of sensors with different sensibilities against diverse chemicals with diverse functional groups. The sensor generates electrical signals which correspond to the gas composition of the sample, and hence unique e-nose profiles are created which may be matched with a diverse pool of reference samples. Several plant materials infected with pathogenic bacteria or fungi have been subjected to e-nose-based detection, and potato is no exception. The earliest detection of potato pathogen using e-nose tool was utilized in 2004 where researchers developed generic system for detection of statuary potato pathogen causing ring rot and bacterial wilt in potato (Stinson 2007). An advancement in e-nose-based detection system of the same disease came in 2014, when a commercial electronic nose (e-nose) having metal oxide sensor array was developed that recognizes volatile compounds emitted by potatoes experimentally infected with Ralstonia solanacearum or Clavibacter michiganensis subsp. sepedonicus, which are bacterial agents of potato brown and ring rot, respectively (Cellini et al. 2017). An e-nose gas chamber and sampling device was designed to detect the volatile profiles in soft rot infected potato tubers. The developed device utilized RBF NN algorithm and SVM algorithm and detected soft rot in tubers with accuracy as high as 89%. Likewise, there is a report of detection of soft rot in potatoes caused by the bacterium Pectobacterium carotovorum through the use of an array of low-cost gas sensors. These researchers utilized the strong odour emitted by rot infected potato in the cold stores and devised a low-cost gas sensor to detect the disease based on signature molecules. Under lab conditions, they demonstrated that out of 11 sensors used, 3 were able to detect the soft rot infected tubers with 100% accuracy (Rutolo et al. 2018). The identified sensors therefore offer promise for an automated in-store monitoring system. E-nose technology has significantly progressed since the first attempts of application to plant diagnosis and may become mature in a near future.
It is worthwhile to point to some valuable publications on the subject of potato entomology, e.g. Giordanengo et al. (2013), the insect-related portions of Potato Health Management (Johnson 2007) and the basic biology of several insect-transmitted diseases as discussed in Stevenson et al. (2001). Numerous field manuals for identifying potato pests have been published, e.g. Zehnder et al. (1994), Strand and Rude (2006) and Johnson (2007). Other suggested readings include Navarre and Pavek (2014) and Kroschel et al. (2020).
1.6 Development in the Area of Pest/Disease Forecasting
Though the chemical-based management of pests and diseases are still dominating, their use can be rationalized. This can be achieved through the use of forecasting models and decision support systems (DSSs). Forecasting allows a better control of a disease and a more efficient use of fungicides by making informed disease management decisions; whereas, the DSS-based strategy can deliver general or site-specific information to the stakeholders enabling them to take firm decisions on the management thereby resulting in economic gains and environment protection (Cooke et al. 2011; Sharma 2019). Various late blight forecasting models and DSSs have been developed across the globe for the management of late blight in different agro-ecologies and are discussed in Chap. 7.
Degree-day models are routinely used for predicting the emergence or build-up of insect population for various insect pests like potato tuber moth, the Andean tuber worms (Symmetrischema tangolias and Tecia solanivora), Colorado potato beetle, other potato beetles, potato psyllids, leaf hoppers etc. in different parts of the world (Keller 2003; Sporleder et al. 2004; Giordanengo et al. 2013). The most impactful is perhaps monitoring of the flight activity of aphids and forecasting the extent of the risk of PVY spread in seed potato crops. Extensive aphid monitoring programmes using suction traps have been running successfully in European countries, the United States and New Zealand, for example. The oldest network is in the United Kingdom, which has been running for more than 50 years. Each week results of trap catch (species composition and abundance) with a cumulative vector pressure index are published and made available to the farmers and others involved with this sector. This index is designed to give the user an assessment of the risk to their crop of PVY spread and helps in decision-making processes when considering the need for insecticide treatments and to decide the best time to burn down/cutting of haulms of potato crops (details in Chap. 9).
1.7 Innovations in Pest and Disease Management
Dynamics of pathogens and insect pests keep on changing over time and space. This warrants the development of novel and future-oriented management strategies which are rapid, easy to operate and implicate, cost effective, widely applicable and well automated in the era of computer-based modernization. There is certainly good development in this direction, and novel futuristic techniques are emerging in potato farming, some are being adopted, while others are expected to be followed in the near future (Sharma and Tiwari 2021). The development and manufacture of effective, safe to humans and environmentally friendly pesticides have been a challenge and an important target. The general trends and strategies for novel pesticides include development of pesticides that are effective at an extremely low dosage, development of pesticides that are readily degradable and less residual in the environment and development of selective toxic agrochemicals. These development strategies have become increasingly prominent. The most developed fungicides are succinate dehydrogenase inhibitors (SDHI), demethylation inhibitors (DMI) and inhibitors of the mitochondrial electron transport chain complex III, i.e. quinone outside inhibitors (QoI) and quinone inside inhibitors (QiI). Due to the development of resistance to fungicides with existing modes of action, the second general trend is the development of fungicides with a novel mode of action and a unique chemical structure, and many fungicides possessing these traits have been launched or are under development. Other trends are the development of novel plant defence activators and novel natural product origin fungicides (Umetsu and Shirai 2020). “Oxathiapiprolin” (Zorvec™, Orondis™) is a new class of piperidinyl thiazole isoxazoline fungicide effective against several fungal diseases, including downy mildew and late blight on crops such as vegetables, ornamentals and turf. Its mode of action involves binding to the oxysterol-binding protein in oomycetes (Hagiwara et al. 2019).
Plant-associated bacteria contribute to their host’s health in diverse ways, among which the emission of disease inhibiting volatile organic compounds (VOCs) is one option. Volatile organic compounds (VOCs) produced by the plant microbiota have been demonstrated to elicit plant defences and inhibit the growth and development of numerous plant pathogens. The inhibitory impact of volatiles emitted by Pseudomonas species against late blight has been shown by impeding mycelial growth and sporangia germination of P. infestans (Bailly and Weisskopf 2017). The VOCs containing sulphur compound S-methyl methane thiosulfonate (MMTS) had shown high in planta protective potential against late blight without phytotoxic effects. This protective activity of MMTS is not mediated by the plant immune system but is due to its anti-oomycete activity (Chinchilla et al. 2019). This provides new perspectives for plant protection by opening new research avenues on the role of VOCs in the interaction between plants and their microbiome and thus could help select for efficient biocontrol strategies and lead to a greener chemical disease management in the field.
Genome editing or gene editing has opened new opportunities to introduce sequence-specific modifications into the genomes of organisms and in the identification, characterization and validation of resistant genes coupled with their deployment in suitable cultivars for the development of disease resistance (Schenke et al. 2020). Genome editing technologies such as transcription activator-like effector nucleases (TALENs) and clustered regularly interspaced short palindromic repeats/CRISPR-associated protein 9 (Cas9) have become powerful genetic tools for increasing pathogen resistance in plants (Zhan et al. 2019). The novel opportunities provided by genome editing, synthetic biology and gene drive may give immense support in managing diseases caused by viral, bacterial and fungal pathogens in potato. A recent study revealed that mutation of a single gene in Arabidopsis, DMR6 (downy mildew resistance 6), led to increased salicyclic acid levels and resistance to several plant pathogens, including bacteria and oomycetes (Zeilmaker et al. 2014). Interestingly, the tomato orthologous SIDMR6–1 is also upregulated in response to infection by Pseudomonas syringae pv tomato and Phytophthora capsici. Null mutants of SIDMR6–1 generated via CRISPR/Cas9 system showed resistance to P. syringae, P. capsici and Xanthomonas spp. without detrimental effects on tomato growth and development (Thomazella et al. 2016). Together, these results suggest that knocking out DMR6 may be a promising strategy to confer broad spectrum disease resistance to plants. A related but advanced system CRISPR/Cas13a is effective to cleave single-stranded RNA, thus providing protection against RNA viruses in plant. Recently, a study reported the reframing of CRISPR/Cas13a to protect the potato plants from Potato virus Y (PVY) (Zhan et al. 2019). Four of the PVY target region P3 protein (potyviral membrane protein), CI (cytoplasmic inclusion bodies), NIb (RNA-dependent RNA polymerase) and CP (coat protein) were selected and used for sgRNA design. There was a suppressed PVY accumulation, and symptoms were attenuated in transgenic potato lines expressing small guide RNA (Cas13a/sgRNA). Another innovative approach is to target susceptibility genes (S-genes) which are beneficial for pathogens’ growth and development. Silencing of S genes attenuates the disease symptoms as observed in several economically important crops such as rice, wheat, tomatoes, citrus etc. A recent study by Kieu et al. (2021) reported that CRISPR/Cas9 genome editing-mediated induced mutations in susceptibility genes confer increased late blight resistance in potatoes. A tetra-allelic deletion mutant (conferring co-expression of two guide RNAs) was generated using CRISPR/Cas9 system targeting seven putative S genes including two DMR6 potato homologues. The mutant plants assayed for late blight resistance have shown significantly high level of resistance as compared to control.
Presently, the exogenous application of double-stranded RNA, small interfering RNA (siRNA) and hpRNA-mediated post-transcriptional gene silencing (i.e. RNA interference or gene silencing) have emerged as sustainable strategies in activating plant defence against phytopathogenic diseases (Machado et al. 2018). The management of late blight disease in potato was also made possible using dsRNA-based spray formulations. A latest study reported that spraying of Phytophthora infestans derived dsRNA molecules as an effective plant protection strategy for the management of potato late blight (Sundaresha et al. 2021). Overall dsRNA-based spray formulation was highly effective in mitigating late blight disease symptoms, and this can be an effective strategy alternative to chemical pesticides. In fact, many studies have been found effective in managing vector populations using dsRNA-based spray, and this can be utilized in managing whiteflies and aphids in potato production system (Arif et al. 2012; Thakur et al. 2014). It is well reported that whitefly mortality has been induced via oral delivery of dsRNA, targeting five important genes namely actin ortholog, ADP/ATP translocase, α-tubulin, ribosomal protein L9 (rpl9) and v-ATPase A. There was enhanced whitefly resistance in transgenic tobacco plants expressing dsRNA of v-ATPase gene (Thakur et al. 2014). Similarly, dsRNA-based spray formulations targeting potential RNAi target genes (TREH, ATPD, ATPE and CHS1) were selected and cloned and further tested through the transdermal dsRNA delivery system against soybean aphids. The delivered dsRNA silenced the gene expressions of target genes with mortality rate up to 81.67%. Such kind of novel, ecofriendly and highly target-specific formulations are the need of the hour for simultaneous management of virus and vector populations.
Unmanned aerial vehicles (UAVs) and drones are becoming increasingly popular in the era of precision agriculture. The drones are excellent in terms of speed of its coverage of large area much faster than humans so they can be a valuable scouting tool especially if other sensors are also equipped in it. The advancement in image processing and machine learning tools has evolved to such an extent that it can generate useful information for the management of the farm (Sugiura et al. 2018). There are several successful examples of drone-based monitoring and redressal of various abiotic and biotic stresses in commercial crop plants. Multispectral, thermal and hyperspectral images are the preferred methods of acquiring information in disease monitoring. The drone-based disease diagnostics has been used for potato diseases. Sugiura et al. (2018) devised RGB sensor-based UAV for the detection of PVY-infected plants. Likewise, the late blight phenotyping, disease progression was also assessed in potato using similar RGB-based UAVs (Sugiura et al. 2016). The spectral imaging based system also captured disease progression and provided area under disease progress curve (AUDPC) as phenotypic data. Identification of the onset of potato black leg disease (Pectobacterium atrosepticum) within using off-the-shelf digital cameras equipped on UAV was also made possible in United Kingdom (Gibson-Poole et al. 2017). Besides disease monitoring, drones are being used for application of pesticides. The advantages of drone-based application of pesticides are on account of savings on pesticides and spraying volume per unit area, less application time and more penetration and coverage of pesticides.
Among the pest control options, potato crops are highly reliant on the use of synthetic pesticides. Potato has been infamously nicknamed “one of the most chemically-dependent crops in the world”. Since the mid-twentieth century, intense use of insecticides has led to the selection of resistant insect pest populations. Potato pests include some of the species that are most prone to evolving resistance to a wide variety of chemicals. The Arthropod Pesticide Resistance Database (2018) lists 469 cases of green peach aphid (Myzus persicae (Sulzer)) resistance to a total of 80 active ingredients, 300 cases of Colorado potato beetle (L. decemlineata) resistance to a total of 56 active ingredients, 111 cases of greenhouse whitefly (T. vaporariorum) resistance to 27 active ingredients and 501 cases of two-spotted spider mite (Tetranychus urticae Koch) resistance to rather impressive 95 active ingredients (as reported in Kroschel et al. 2020). The extent of resistance is likely to be underestimated because not every case of its development is entered into the database. It is possible that the ability to deal with toxic glycoalkaloids contained in potato foliage serves as a preadaptation to resisting synthetic pesticides (Alyokhin and Chen 2017). Not every population of a given pest species is resistant to all compounds that have been recorded to fail against that species. However, these statistics vividly illustrate the seriousness of the problem. On several occasions, potato growers already experienced the situation when virtually all commercially available chemicals failed to control their targets (Alyokhin et al. 2013). Today, as with other major crops, potato culture has to deal with increasing environmental and public concerns that lead to the reduction of new chemical discoveries and development, while also supporting a rapidly rising world demand. In recent decades, there has been a shift in insecticide development to safer and more targeted (or narrow spectrum) insecticides that are often less toxic to non-target species. Today, there is a wide diversity of insecticide mode of actions and products that can aid in a more sustainable approach to pest management for potatoes (details in Chap. 16).
Despite the fact that chemical control is the most popular form of insect pest management in the potato industry, yet, this does not necessarily have to be the case. Very basic understanding of the pest population dynamics revealed the periods of maximum spread of virus like PVY. This information is routinely used to decide the time of haulms-cutting or destruction. This simple cultural practice is perhaps the most important virus disease control tactic that is widely adopted round the world in seed potato crops. Similarly, in subtropical conditions in India, studies on population dynamics of the most efficient vector of potato viruses, the peach-potato aphid, M. persicae, were used to decide the dates of planting and haulms-cutting during a period of lowest aphid activity for quality seed potato production (Pushkarnath 1967). Thorough understanding of the potato ecosystem using the systems approach is the way forward. Similarly, high hopes are associated with novel genomics techniques to provide breakthroughs for major problems in potato pest management, along with other aspects.
The management tactics are rendered ineffective due to contiguous evaluation of pathogen variants, resistance development and emergence of new pests and diseases which usually were characterized as of minor importance or were under the radar. Climate change is/will complicate the situation further. And needless to say, the synthetic plant protection chemicals have always been under criticism due to their ill effects to human beings and the environment. In this book, an attempt has been made to bring together information on such aspects of pest management which are believed to be more sustainable in the long run.
1.8 Conclusion
Potato is among the most important food crops which has a pivotal role to play in food and nutritional security of the populace in the foreseeable times. The yield and quality of potato is contained by a myriad of insect pests, fungi, bacterial, viruses and nematode pathogens. The pest management regime is currently almost entirely dependent on the use of plant protection products, the use of which is not sustainable in the long run. Newer issue like that of climate change, invasive species and newer species/strains of pest and pathogens are straining the pest management practices further. As time witnessed, the potato pets and disease management has come a long way, its high time to think on the lines to make the practices sustainable for the future generations.
References
Alyokhin A, Chen YH (2017) Adaptation to toxic hosts as a factor in the evolution of insecticide resistance. Curr Opin Insect Sci 21(1):33–38
Alyokhin A, Chen YH, Udalov M, Benkovskaya G, Lindström L (2013) Evolutionary considerations in potato pest management. In: Giordanengo P, Vincent C, Alyokhin A (eds) Insect pests of potato: global perspectives on biology and management. Academic Press, Oxford, pp 543–571
Anderson PK, Cunningham AA, Patel NG et al (2004) Emerging infectious diseases of plants: pathogen pollution, climate change and agrotechnology drivers. Trends Ecol Evol 19:535–544. https://doi.org/10.1016/j.tree.2004.07.021
Ardestani ST, Sharifnabi B, Zare R, Moghadam AA (2010) New Alternaria species associated with potato leaf spot in various potato growing regions of Iran. Iranian J Plant Pathol 45:83–86
Arif M, Azhar U, Arshad M et al (2012) Engineering broad-spectrum resistance against RNA viruses in potato. Transgenic Res 21:303–311
Arthropod Pesticide Resistance Database (2018). https://www.pesticideresistance.org. Accessed 3 Jul 2018
Bailly A, Weisskopf L (2017) Mining the volatilomes of plant-associated microbiota for new biocontrol solutions. Front Microbiol 8:1638. https://doi.org/10.3389/fmicb.2017.01638
Barton MG, Terblanche JS (2014) Predicting performance and survival across topographically heterogeneous landscapes: the global pest insect Helicoverpa armigera (Hübner, 1808) (Lepidoptera: Noctuidae). Austral Entomol 53(3):249–258
Bebber DP, Ramotowski MA, Gurr SJ (2013) Crop pests and pathogens move pole wards in a warming world. Nat Clim Change 3(11):985
Becktell M, Daughtrey M, Fry W (2005) Temperature and leaf wetness requirements for pathogen establishment, incubation period, and sporulation of Phytophthora infestans on Petunia hybrida. Plant Dis 89(9):975–979
Borges KM, Cooper WR, Garczynski SF, Thinakaran J, Jensen AS, Horton DR et al (2017) “Candidatus Liberibacter solanacearum” associated with the psyllid, Bactericera maculipennis (Hemiptera: Triozidae). Environ Entomol 46(2):210–216. https://doi.org/10.1093/ee/nvw174
CABI (2017) Leptinotarsa decemlineata (Colorado potato beetle) data sheet. In: Invasive species compendium. CAB International, Wallingford
Caparros Megido R, Brostaux Y, Haubruge E et al (2013) Propensity of the tomato leafminer, Tuta absoluta (Lepidoptera: Gelechiidae), to develop on four potato plant varieties. Am J Potato Res 90:255–260. https://doi.org/10.1007/s12230-013-9300-9
Cellini A, Blasioli S, Biondi E et al (2017) Potential applications and limitations of electronic nose devices for plant disease diagnosis. Sensors 17:2596
Charkowski AO (2015) Biology and control of Pectobacterium in potato. Am J Potato Res 92(2):223–229
Chinchilla D, Bruisson S, Meyer S et al (2019) A sulfur-containing volatile emitted by potato-associated bacteria confers protection against late blight through direct anti-oomycete activity. Sci Rep 9(1):18778
Cooke DEL, Kessel GJT, Lassen P, Hansen JG (2019) The European population of Phytophthora infestans in a global context. Seventeenth Euroblight workshop, York, 12-15 May, 2019. WUR-Special Report No 19, pp 35–36
Cooke LR, Schepers HTAM, Hermansen A et al (2011) Epidemiology and integrated control of potato late blight in Europe. Potato Res 54:183–222
De Boer SH, Li X, Ward LJ (2012) Pectobacterium spp. associated with bacterial stem rot syndrome of potato in Canada. Phytopathology 102(10):937–947
de Werra P, Bussereau F, Keiser A, Ziegler D (2015) First report of potato blackleg caused by Pectobacterium carotovorum subsp Brasiliense in Switzerland. Plant Dis 99(4):551
Devaux A, Kromann P, Ortiz O (2014) Potatoes for sustainable global food security. Potato Res 57:185–199. https://doi.org/10.1007/s11540-014-9265-1
Dillon ME, Wang G, Huey RB (2010) Global metabolic impacts of recent climate warming. Nature 467:704–706
Dua VK, Singh BP, Islam A, Sanjeev S (2015) Potential impact of climate change on late blight outbreak in western Uttar Pradesh and Punjab using Jhulsacast model. Potato J 42(1):58–71
Duarte V, De Boer SH, Ward LJ, De Oliveira AMR (2004) Characterization of atypical Erwinia carotovora strains causing blackleg of potato in Brazil. J Appl Microbiol 96(3):535–545
Elphinstone JG (2005) The current bacterial wilt situation: a global overview. In: Allen C, Piror P, Hayward AC (eds) Bacterial wilt disease and the Ralstonia solanacearum species complex. APS Press, St Paul, pp 9–28
FAOSTAT (2020) Food and Agriculture Organization of the United Nations. http://www.fao.org/faostat/en/#data/QC
Fry WE (2020) Phytophthora infestans: the itinerant invader; “late blight”: the persistent disease. Phytoparasitica 48:87–94. https://doi.org/10.1007/s12600-019-00778-3
Fry WE, Birch PRJ, Judelson HS et al (2015) Five reasons to consider Phytophthora infestans a re-emerging pathogen. Phytopathology 105:966–981
Gamarra H, Carhuapoma P, Mujica N, Kreuze J, Kroschel J (2016b) Greenhouse whitefly, Trialeurodes vaporariorum (Westwood 1956). In: Kroschel J, Mujica N, Carhuapoma P, Sporleder M (eds) Pest distribution and risk atlas for Africa—potential global and regional distribution and abundance of agricultural and horticultural pests and associated biocontrol agents under current and future climates. International Potato Center (CIP), Lima, pp 154–168. https://doi.org/10.4160/9789290604761
Gamarra H, Mujica N, Carhuapoma P, Kreuze J, Kroschel J (2016a) Sweetpotato whitefly, Bemisia tabaci (Gennadius 1989) (biotype B). In: Kroschel J, Mujica N, Carhuapoma P, Sporleder M (eds) Pest distribution and risk atlas for Africa—potential global and regional distribution and abundance of agricultural and horticultural pests and associated biocontrol agents under current and future climates. International Potato Center (CIP), Lima, pp 85–99. https://doi.org/10.4160/9789290604761
Gibson-Poole S, Humphris S, Toth I, Hamilton A (2017) Identification of the onset of disease within a potato crop using a UAV equipped with un-modified and modified commercial off-the-shelf digital cameras. Adv Anim Biosci 8:812–816
Giordanengo P, Vincent C, Alyokhin A (eds) (2013) Insect pests of potato. Elsevier, Amsterdam
Gray S, De Boer S, Lorenzen J, Karasev A, Whitworth J, Nolte P, Singh R, Boucher A, Xu H (2010) Potato virus Y: an evolving concern for potato crops in the United States and Canada. Plant Dis 94(12):1384–1397
Hagiwara H, Ezaki R, Hamada T et al (2019) Development of a novel fungicide, tolprocarb. J Pestic Sci 44:1–6
Hannukkala AO, Kaukoranta T, Lehtinen A, Rahkonen A (2007) Late-blight epidemics on potato in Finland, 1933-2002; increased and earlier occurrence of epidemics associated with climate change and lack of rotation. Plant Pathol 56(1):167–176
Harrison MD, Venette JR (1970) Chemical control of potato early blight and its effect on potato yield. Am J Potato Res 47(3):81–86
Haverkort AJ, Struik PC, Visser RGF, Jacobsen E (2009) Applied biotechnology to combat late blight in potato caused by Phytophthora infestans. Potato Res 52(3):249–264
Hijmans RJ (2003) The effect of climate change on global potato production. Am J Potato Res 80:271–279. https://doi.org/10.1007/BF02855363
Horsfield A, Wicks T, Davies K, Wilson D, Paton S (2010) Effect of fungicide use strategies on the control of early blight (Alternaria solani) and potato yield. Australas Plant Pathol 39(4):368–375
IPCC (2013) Climate change 2013: mitigation of climate change, vol 3. Cambridge University Press, Cambridge
Johnson DA (ed) (2007) Potato health management. The American Phytopathological Society Press, St Paul
Johnson J, Sharma G, Srinivasan S et al (2021) Enhanced field-based detection of potato blight in complex backgrounds using deep learning. Plant Phenomics 2021:1–13. https://doi.org/10.34133/2021/9835724
Kanle Satishchandra N, Chakravarthy AK, Özgökçe MS, Atlihan R (2019) Population growth potential of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) on tomato, potato, and eggplant. J Appl Entomol 143(5):518–526
Karasev AV, Gray SM (2013) Continuous and emerging challenges of potato virus Y in potato. Annu Rev Phytopathol 51:571–586
Kaukoranta T (1996) Impact of global warming on potato late blight: risk, yield loss, and control. Agric Food Sci Finl 5:311–327
Keller S (2003) Integrated pest management of the potato tuber moth in cropping systems of different agro-ecological zones. In: Kroschel J (ed) Advances in crop research, vol 1. Margraf, Weikersheim
Kieu NP, Lenman M, Wang ES et al (2021) Mutations introduced in susceptibility genes through CRISPR/Cas9 genome editing confer increased late blight resistance in potatoes. Sci Rep 11:4487
Kim H-S, Ma B, Perna NT, Charkowski AO (2009) Phylogeny and virulence of naturally occurring type III secretion system-deficient Pectobacterium strains. Appl Environ Microbiol 75(13):4539–4549
Kroschel J, Mujica N, Alcazar J, Canedo V, Zegarra O (2012) Developing integrated pest management for potato: experiences and lessons from two distinct potato production systems of Peru. In: He Z, Larkin RP, Honeycutt CW (eds) Sustainable potato production: global case studies. Springer, London, pp 419–450
Kroschel J, Mujica N, Carhuapoma P, Sporleder M (2016) Pest distribution and risk atlas for Africa—potential global and regional distribution and abundance of agricultural and horticultural pests and associated biocontrol agents under current and future climates. International Potato Center (CIP), Lima, p 650. https://doi.org/10.4160/9789290604761
Kroschel J, Mujica N, Okonya J, Alyokhin A (2020) Insect pests affecting potatoes in tropical, subtropical, and temperate regions. In: Campos H, Ortiz O (eds) The potato crop. Springer, Cham, pp 251–306
Kroschel J, Schaub B (2013) Biology and ecology of potato tuber moths as major pests of potato. In: Giordanengo P, Vincent C, Alyokhin A (eds) Insect pests of potato: global perspectives on biology and management. Academic Press, Oxford, pp 165–192
Kroschel J, Sporleder M, Tonnang HEZ, Juarez H, Carhuapoma P, Gonzales JC, Simon R (2013) Predicting climate change caused changes in global temperature on potato tuber moth Phthorimaea operculella (Zeller) distribution and abundance using phenology modeling and GIS mapping. Agric For Meteorol 170:228–241
Kumar R, Tiwari RK, Jeevalatha A et al (2021) Potato apical leaf curl disease: current status and perspectives on a disease caused by tomato leaf curl New Delhi virus. J Plant Dis Prot 128:897–911. https://doi.org/10.1007/s41348-021-00463-w
Laurila J, Ahola V, Lehtinen A et al (2008) Characterization of Dickeya strains isolated from potato and river water samples in Finland. Eur J Plant Pathol 122(2):213–225. https://doi.org/10.1007/s10658-008-9274-5
Leiminger J, Bassler E, Knappe C (2015) Quantification of disease progression of Alternaria spp. on potato using real-time PCR. Eur J Plant Pathol 141:295–309
Leiminger JH, Hausladen H (2014) Untersuchungen zur Befallsentwicklung und Ertragswirkung der Dürrfleckenkrankheit (Alternaria spp.) in Kartoffelsorten unterschiedlicher Reifegruppe. Gesunde Pflanzen 66:29–36
Li Z, Paul R, Ba Tis T et al (2019) Non-invasive plant disease diagnostics enabled by smartphone-based fingerprinting of leaf volatiles. Nature Plants 5:856–866
Machado AK, Brown NA, Urban M et al (2018) RNAi as an emerging approach to control fusarium head blight disease and mycotoxin contamination in cereals. Pest Manag Sci 74:790–799
Minhas JS, Kumar P, Kumar D, Dua VK, Gupta YK (2018) Response of potato to elevated CO2 under short days: growth, physiological parameters and tuber yield. Indian J Hortic 75(1):82–86
Mohanty SP, Hughes DP, Salathé M (2016) Using deep learning for image-based plant disease detection. Front Plant Sci 7:1419
Mujica N, Carhuapoma P, Kroschel J (2016) Serpentine leafminer fly, Liriomyza huidobrensis (Blanchard 1926). In: Kroschel J, Mujica N, Carhuapoma P, Sporleder M (eds) Pest distribution and risk atlas for Africa—potential global and regional distribution and abundance of agricultural and horticultural pests and associated biocontrol agents under current and future climates. International Potato Center (CIP), Lima, pp 114–125. https://doi.org/10.4160/9789290604761
Mujica N, Kroschel J (2013) Pest intensity-crop loss relationships for the leafminer fly Liriomyza huidobrensis (Blanchard) in different potato (Solanum tuberosum L.) varieties. Crop Prot 47:6–16
Nagoshi RN, Dhanani I, Asokan R, Mahadevaswamy HM, Kalleshwaraswamy CM, Sharanabasappa et al (2019) Genetic characterization of fall armyworm infesting South Africa and India indicate recent introduction from a common source population. PLoS One 14(5):e0217755. https://doi.org/10.1371/journal.pone.0217755
Navarre R, Pavek MJ (eds) (2014) The potato: botany, production and uses. CAB International, Wallingford
Ngadze E, Brady CL, Coutinho TA et al (2012) Pectinolytic bacteria associated with potato soft rot and blackleg in South Africa and Zimbabwe. Eur J Plant Pathol 134:33–549. https://doi.org/10.1007/s10658-012-0036-z
Oerke EC, Dehne HW, Schönbeck F, Weber A (1994) Crop production and crop protection. Estimated losses in major food and cash crops. Elsevier, Amsterdam, p 808
Oppenheim D, Shani G, Erlich O, Tsror L (2019) Using deep learning for image-based potato tuber disease detection. Phytopathology 109:1083–1087
Parkinson N, Stead D, Bew J et al (2009) Dickeya species relatedness and clade structure determined by comparison of RecA sequences. Int J Syst Evol Microbiol 59(10):2388–2393. https://doi.org/10.1099/ijs.0.009258-0
Pérombelon MCM, Kelman A (1987) Blackleg and other potato diseases caused by soft rot erwinias: proposal for revision of terminology. Plant Dis 71(3):283–285
Pitman AR, Harrow SA, Visnovsky SB (2010) Genetic characterisation of Pectobacterium wasabiae causing soft rot disease of potato in New Zealand. Eur J Plant Pathol 126(3):423–435
Pitman AR, Wright PJ, Galbraith MD, Harrow SA (2008) Biochemical and genetic diversity of pectolytic enterobacteria causing soft rot disease of potatoes in New Zealand. Australas Plant Pathol 37(6):559–568
Prior P, Ailloud F, Dalsing BL et al (2016) Genomic and proteomic evidence supporting the division of the plant pathogen Ralstonia solanacearum into three species. BMC Genomics 17(1):90
Pushkarnath (1967) Seed potato production in the sub-tropical plains of India. Am Potato J 44:429–441
Radcliffe EB, Johnson KB (1994) Biology and management of leafhoppers on potato. In: Zehnder GW, Powelson ML, Jansson RK, Raman KV (eds) Advances in potato pest biology and management. APS Press, Saint Paul, pp 71–82
Rana A, Dua VK, Chauhan S et al (2020) Climate change and potato productivity in Punjab—impacts and adaptation. Potato Res 63:597–613. https://doi.org/10.1007/s11540-020-09460-2
Raymundo R, Asseng S, Robertson R, Petsakos A, Hoogenboom G, Quiroz R, Hareau G, Wolf J (2018) Climate change impact on global potato production. Eur J Agron 100:87–98
Rehman A, Saeed AM, Khan AM, Rafique A, Ashraf M (2012) Potato: a new host plant of Tuta absoluta Povolny (Lepidoptera: Gelechiidae) in Turkey. Pak J Zool 44(4):1183–1184
Rodrigues TTMS, Berbee ML, Siommons EG et al (2010) First report of Alternaria tomatophila and A. grandis causing early blight on tomato and potato in Brazil. New Dis Rep 22:28
Rutolo MF, Clarkson JP, Covington JA (2018) The use of an electronic nose to detect early signs of soft-rot infection in potatoes. Biosyst Eng 167:137–143
Safni I, Cleenwerck I, De Vos P et al (2014) Polyphasic taxonomic revision of the Ralstonia solanacearum species complex: proposal to emend the descriptions of Ralstonia solanacearum and Ralstonia syzygii and reclassify current R. syzygii strains as Ralstonia syzygii subsp. syzygii subsp. nov., R. solanacearum phylotype IV strains as Ralstonia syzygii subsp. indonesiensis subsp. nov., banana blood disease bacterium strains as Ralstonia syzygii subsp. celebesensis subsp. nov. and R. solanacearum phylotype I and III strains as Ralstonia pseudosolanacearum sp. nov. Int J Syst Evol Microbiol 64:3087–3103. https://doi.org/10.1099/ijs.0.066712-0
Samson R, Legendre JB, Christen R et al (2005) Transfer of Pectobacterium chrysanthemi (Burkholder et Al. 1953) Brenner et Al. 1973 and Brenneria paradisiaca to the genus Dickeya gen. nov. as Dickeya chrysanthemi comb. nov. and Dickeya paradisiaca comb. nov. and delineation of four novel species, Dickeya dadantii sp. nov., Dickeya dianthicola sp. nov., Dickeya dieffenbachiae sp. nov. and Dickeya zeae sp. nov. Int J Syst Evol Microbiol 55(Pt 4):1415–1427. https://doi.org/10.1099/ijs.0.02791-0
Schaub B, Carhuapoma P, Kroschel J (2016) Guatemalan potato tuber moth, Tecia solanivora (Povolny 1973). In: Kroschel J, Mujica N, Carhuapoma P, Sporleder M (eds) Pest distribution and risk atlas for Africa—potential global and regional distribution and abundance of agricultural and horticultural pests and associated biocontrol agents under current and future climates. International Potato Center (CIP), Lima, pp 24–38. https://doi.org/10.4160/9789290604761
Schenke D, Cai D (2020) Applications of CRISPR/Cas to improve crop disease resistance: beyond inactivation of susceptibility factors. iScience 23:101478
Shah MA, Aarti B, Naga KC, Subhash S et al (2020) Important potato pests and their management. In: Singh AK, Chakrabarti SK, Singh B, Sharma J, Dua VK (eds) Potato science and technology for sub-tropics. NIPA, New Delhi, pp 295–326
Shah MA, Al-Tawaha AR, Naga KC, Sharma S (2021b) Insect pest management in the era of climate change. In: Fahad S, Sonmez O, Saud S, Wang D, Wu C, Adnan M, Tarun V (eds) Climate change and plants: biodiversity, growth and interactions. CRC Press, New York, pp 133–146
Shah MA, Sharma S, Kumar R (2021a) Seasonal population dynamics and flight activity of sweetpotato whitefly, Bemisia tabaci on potato in the sub-tropical plains of India. Arthropod-Plant Interact 15:605–613. https://doi.org/10.1007/s11829-021-09835-3
Sharma S (2019) Scheduling fungicide application for economic management of late blight using indo-Blightcast model. Potato J 46(2):124–131
Sharma S, Tiwari RK (2021) Futuristic strategies for management of potato diseases. In: Bharat NK, Gautam HR (eds) Technology strides in plant health management. Neoti Book Agency, New Delhi, pp 63–90
She XM, Lan GB, Tang YF, He ZF (2017) Pectobacterium carotovorum subsp. brasiliense causing pepper black spot disease in China. J Plant Pathol 99(3):769–772
Shoaib A, Akhtar N, Akhtar S, Hafeez R (2014) First report of Alternaria longipes causing leaf spot of potato cv. Sante in Pakistan. Plant Dis 98:1742
Sidana JA, Singh BI, Sharma O (2018) Occurrence of the new invasive pest, fall armyworm, Spodoptera frugiperda (JE smith) (Lepidoptera: Noctuidae), in the maize fields of Karnataka, India. Curr Sci 115:621–623
Singh BP, Dua VK, Govindakrishnan PM, Sharma S (2013) Impact of climate change on potato. In: Singh HP (ed) Climate resilient horticulture: adaptation and mitigation strategies. Springer, New Delhi, pp 125–135
Sławiak M, van Beckhoven JRCM, Speksnijder AGCL et al (2009) Biochemical and genetical analysis reveal a new clade of biovar 3 Dickeya spp. strains isolated from potato in Europe. Eur J Plant Pathol 125(2):245–261. https://doi.org/10.1007/s10658-009-9479-2
Sporleder M, Carhuapoma P, Kroschel J (2016) Andean potato tuber moth, Symmetrischema tangolias (Gyen 1913). In: Kroschel J, Mujica N, Carhuapoma P, Sporleder M (eds) Pest distribution and risk atlas for Africa—potential global and regional distribution and abundance of agricultural and horticultural pests and associated biocontrol agents under current and future climates. International Potato Center (CIP), Lima, pp 39–53. https://doi.org/10.4160/9789290604761
Sporleder M, Kroschel J, Gutierrez Quispe MR, Lagnaoui A (2004) A temperature-based simulation model for the potato tuberworm, Phthorimaea operculella Zeller (Lepidoptera: Gelechiidae). Environ Entomol 33:477–486
Sridhar V, Nitin KS, Nagaraja T (2015) Comparative biology of south American tomato moth, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) on three solanaceous host plants. Pest Manag Hortic Ecosys 21(2):159–161
Stevenson WR, Loria R, Franc GD, Weingartner DP (2001) Compendium of potato diseases, 2nd edn. The American Phytopathological Society, St Paul
Stinson JA (2007) Sensing potato pathogens: an automated approach. The University of Manchester, Manchester
Strand LL, Rude PA (2006) Integrated pest management for potatoes in the western United States, 2nd edn. University of California Agriculture and Natural Resources Publications, Davis
Sugiura R, Tsuda S, Tamiya S et al (2016) Field phenotyping system for the assessment of potato late blight resistance using RGB imagery from an unmanned aerial vehicle. Biosyst Eng 148:1–10
Sugiura R, Tsuda S, Tsuji H, Murakami N (2018) Virus-infected plant detection in potato seed production field by UAV imagery. American Society of Agricultural and Biological Engineers, Washington, p 1
Sundaresha S, Sharma S, Bairwa A et al (2021) Spraying of dsRNA molecules derived from Phytophthora infestans, as a plant protection strategies for the management of potato late blight
Taheri AS, Sharif NB, Zare R, Abbasi MA (2009) Alternaria interrupta, a new pathogen causing potato early blight in Iran. Rostaniha 10:72–73
Thakur N, Upadhyay SK, Verma PC et al (2014) Enhanced whitefly resistance in transgenic tobacco plants expressing double stranded RNA of v-ATPase a gene. PLoS One 9:e87235
Thomazella DPT, Brail Q, Dahlbeck D, Staskawicz B (2016) CRISPR-Cas9 mediated mutagenesis of a DMR6 ortholog in tomato confers broad-spectrum disease resistance bioRxiv https://doi.org/10.1101/064824
Toth IK, van der Wolf JM, Saddler et al (2011) Dickeya species: an emerging problem for potato production in Europe. Plant Pathol 60(3):385–399. https://doi.org/10.1111/j.1365-3059.2011.02427.x
Tymon LS, Peever TL, Johnson DA (2016) Identification and enumeration of small-spored Alternaria species associated with potato in the US northwest. Plant Dis 100:465–472
Umetsu N, Shirai Y (2020) Development of novel pesticides in the 21st century. J Pestic Sci 45(2):54–74
Usharani KS, Surendranath B, Paul-Khurana SM, Garg ID, Malathi VG (2004) Potato leaf curl—a new disease of potato in northern India caused by a strain of tomato leaf curl New Delhi virus. Plant Pathol 253:235
van der Merwe JJ, Coutinho TA, Korsten L, van der Waals JE (2010) Pectobacterium carotovorum subsp. brasiliensis causing blackleg on potatoes in South Africa. Eur J Plant Pathol 126(2):175–185
Van der Waals JE, Korsten L, Aveling TAS (2001) A review of early blight of potato. Afr Plant Protect 7(2):91–102
Van der Wolf JM, de Haan EG, Kastelein P et al (2017) Virulence of Pectobacterium carotovorum subsp. brasiliense on potato compared with that of other Pectobacterium and Dickeya species under climatic conditions prevailing in the Netherlands. Plant Pathol 66(4):571–583
Wang J, Haapalainen M, Schott T et al (2017a) Genomic sequence of ‘Candidatus Liberibacter solanacearum’ haplotype C and its comparison with haplotype a and B genomes. PLoS One 12(2):e0171531
Wang J, Wang YH, Dai PG et al (2017c) First report of tobacco bacterial leaf blight caused by Pectobacterium carotovorum subsp. brasiliense in China. Plant Dis 101(5):830–830
Wang N, Pierson EA, Setubal JC et al (2017b) The Candidatus Liberibacter–host interface: insights into pathogenesis mechanisms and disease control. Annu Rev Phytopathol 55:451–482
Wates TD, Jensen AS (2014) Insect pests of potato. In: Navarre R, Pavek MJ (eds) The potato, botany, production and uses. CABI, Wallingford, pp 133–147
Yap M-N, Barak JD, Charkowski AO (2004) Genomic diversity of Erwinia carotovora subsp. carotovora and its correlation with virulence. Appl Environ Microbiol 70(5):3013–3023
Yu SJ (1982) Host plant induction of glutathione S-transferase in the fall armyworm. Pestic Biochem Physiol 18(1):101–106
Zehnder GW, Powelson ML, Jansson RK, Raman KV (1994) Advances in potato pest biology and management. The American Phytopathological Society Press, St Paul
Zeilmaker T, Ludwig NR, Elberse J et al (2014) Downy mildew resistant 6 and DMR6-like oxygenase 1 are partially redundant but distinct suppressors of immunity in Arabidopsis. Wiley Online Libr 81:210–222
Zhan X, Zhang F, Zhong Z et al (2019) Generation of virus-resistant potato plants by RNA genome targeting. Plant Biotechnol J 17:1814–1822
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Chakrabarti, S.K., Sharma, S., Shah, M.A. (2022). Potato Pests and Diseases: A Global Perspective. In: Chakrabarti, S.K., Sharma, S., Shah, M.A. (eds) Sustainable Management of Potato Pests and Diseases. Springer, Singapore. https://doi.org/10.1007/978-981-16-7695-6_1
Download citation
DOI: https://doi.org/10.1007/978-981-16-7695-6_1
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-16-7694-9
Online ISBN: 978-981-16-7695-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)