
Abstract
Potato (Solanum tuberosum L.) is cultivated in diverse agroecosystems, which may harbor different insect pests; accordingly, potato farmers need to have appropriate site-specific pest control solutions. We developed Integrated Pest Management (IPM) strategies for potato production systems of the high Andes and the coast of Peru. This required considering all economically important pests and developing technological innovations to replace farmers’ pesticide applications with equal efficacy. Examples are the use of plastic barriers that effectively prevent infestations of migrating Andean potato weevils (Premnotrypes spp.), the use of attract-and-kill for managing potato tuber moths [Phthorimaea operculella (Zeller), Symmetrischema tangolias (Gyen.)], or the rational use of insecticides to control flea beetles (Epitrix spp.) in the Andean highlands, or the leafminer fly [Liriomyza huidobrensis (Blanchard)] and the bud midge [Prodiplosis longifila (Gagne)] in the coastal lowlands. Moreover, the resilience of potato agroecosystems can be increased through augmentation strategies for natural enemies at the field level and inoculative biological control to recuperate species lost through the intensive use of pesticides. Potato IPM showed clear economic and ecological benefits at pilot sites. Strong public-private partnerships will be crucial for technology delivery, and well-trained field advisors are required to support the specific needs of farmers to adopt IPM.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
The origin of the potato (Solanum tuberosum L.) is the High Andes in South America. Its global distribution began about 300 years ago: first to Europe and then to other parts of the world. Today, potato is grown in over 140 countries, more than 100 of them in tropical and subtropical zones, with China being the largest producer. Losses due to insect pests, which occur during the potato cropping and storage period, are often very high; a 16% of loss was estimated, worldwide (Oerke et al. 1994). Locally, if not routinely controlled, reductions in tuber yield and quality can be between 30% and 70% for various insect pests (Raman and Radcliffe 1992). Potato producing farmers in developing countries worldwide have to contend with about 20 insect pests. With some exceptions, a minimum of 2–4 pests often reach pest status requiring the application of control methods. Many pests have evolved in the center of origin of the potato, and hence farmers in the Andean region are confronted by a higher number of pest species than farmers in Africa or Asia. Some species such as the potato tuber moth [Phthorimaea operculella (Zeller)] and the leafminer fly [Liriomyza huidobrensis (Blanchard)] have become invasive and occur today as serious pests in many tropical and subtropical regions.
In developing countries, the one-sided use of often highly toxic insecticides is a common pest management practice. Farmers receive inadequate training on Integrated Pest Management (IPM) and there is often poor availability and quality of protection equipment and no collection services for used pesticide containers (Orozco et al. 2009; Arica et al. 2006). Serious health threats of pesticides to farmers, consumers and the environment have increased the interest in the search for safer control alternatives through the development of IPM. In addition, a rapid build-up of pest resistance to pesticides has frequently limited their use in potato (Llanderal-Cazares et al. 1996). However, farmers have often been reluctant to adopt IPM for many reasons, including the following: (i) IPM strategies are mainly targeted towards, or available against, single pest species and do not consider all pests in a specific agroecosystem (Trumble 1998; Horne and Page 2008); (ii) Promising IPM technologies are not available on the market or are too expensive; (iii) IPM technologies are not as immediately effective as pesticides, which are relatively cheap, and they are too complex to apply. Farmers can more easily rely on simple and proven pesticide applications (Daxl et al. 1994). Regular insecticide applications are encouraged, even if no pests have been observed, to avoid any risks; (iv) IPM is not understood or applied as a system approach; for this, experienced IPM field advisors would be crucial to support farmers in their IPM decision making. A survey by Horne et al. (1999) showed that the IPM adoption rates of Australian potato growers varied from 0% to 100% depending on the presence or absence of an IPM advisor. Similar observations have been made by Alston and Reading (1998).
By applying a holistic framework for the development of IPM, the objective of our research was to develop practical, economic and ecological solutions to pest management, applicable to resource-poor farmers in the Andean highlands and coastal region of Peru.
2 Pest Problems and Management in Andean Highland and Coastal Lowland Potato Production Systems of Peru
2.1 Andean Highlands
In the Peruvian Andean highlands, potato is mainly produced by small-scale farmers at altitudes between 2,800 and 4,200 m from October to May under rain-fed conditions with an annual precipitation of some 600 mm. Potato cultivation is severely constrained by many pest problems, but farmers’ main response is to control infestations of Andean potato weevils by applying hazardous class Ia and Ib (e.g., carbofuran, metamidophos) insecticides (Orozco et al. 2009). Andean potato weevils belong to the genera Premmnotrypes, Rhigopsidius, and Phyrdenus. With 12 species, the genus Premnotrypes is the most important and the most widely distributed. P. suturicallus Kuschel occurs mainly in the central highlands, P. vorax (Hustache) in the northern and P. latithorax (Pierce) in the southern highlands of Peru (Alcazar and Cisneros 1999). While Andean potato weevils represent the only major biotic insect pest problem at altitudes above 3,800 m, in inner Andean valleys, other important pests are the potato tuber moth (P. operculella) and the Andean potato tuber moth [Symmetrischema tangolias (Gyen.)]; both pests are native to South America and are mainly controlled by farmers in potato storage (Keller 2003). Occasional pests are flea beetles (Epitrix spp.), infestations of which are kept under control by insecticide applications for the Andean potato weevil.
2.2 Coastal Lowlands
The Peruvian coastal region, at altitudes below 500 m, is characterized by an extremely arid subtropical desert climate, with a precipitation of less than 50 mm yearly. Here, the Cañete Valley is one of the most intensive cropping regions, covering some 23,000 ha, which are mostly cultivated by small-to-medium-scale farmers, who produce for the local and export markets. Among the many agricultural and horticultural crops produced in this valley, the potato is an important crop during the winter cropping season. The leafminer fly (L. huidobrensis) is a serious pest on potato and many other vegetable crops, which farmers try to control by the frequent application (8–13 times per season) of mostly highly hazardous pesticides. Insecticides represent the highest input costs (an average of US$600/ha) followed by fertilizers, fungicides, and manure. Without control, yields are commonly reduced by more than 50% (Cisneros and Mujica 1997). The frequent insecticide applications have caused severe secondary infestations by the white mite [Poliphagotarsonemus latus (Banks)] and the bud midge [Prodiplosis longifila (Gagne)]. Depending on the climatic conditions, both pests can seriously affect tuber yield and quality and need to be addressed in an IPM program.
3 Framework for IPM Research and Development in Potato
The International Potato Center (CIP) is investigating safer alternatives to replace toxic chemicals, and anticipates a widespread impact on poverty reduction, food security, human health and environmental protection through the development and adoption of IPM in potato production. In its global research program, a holistic working framework for potato pest management research and development (research outputs) and its application by farmers (research outcomes) was developed (Fig. 25.1).

Framework for integrated pest management (IPM) research and development (research outputs) and its application by farmers (research outcomes)
In this framework, ecosystem research constitutes the basis, aiming at understanding the specific characteristics of potato agroecosystems, the influence of farmers’ crop management interventions on pests and natural enemies, as well as the interactions and relationships among these groups of insects. An inventory of insect communities in potato agroecosystems is being built up with reference material in CIP’s entomological museum. Investigations into the efficacy of the functional diversity in natural pest control are related to landscape fragmentation and farmers’ practices. This research reveals species important in balancing pest problems and self-regulating agroecosystems, and contributes in the long term to the assessment of changes in the stability of systems by identifying relevant bioindicators (Mujica and Kroschel 2011). Pest population ecology and biological studies investigate pest population dynamics influenced by climatic (abiotic), biotic, and external factors such as cultural practices (rotation systems, pest-infested seed, change in sowing date, etc.) applied by farmers (Keller 2003; Kroschel and Koch 1994). This is supported by the development and application of phenology modeling to predict pest population development in different agroecological zones, as well as critical infestation periods of pests (Kroschel and Sporleder 2006; Sporleder et al. 2004; Keller 2003). In combination with Geographic Information Systems (GIS), phenology models make it possible to carry out pest risk assessments under the current and future climate conditions (Kroschel et al. 2011a; Sporleder et al. 2008). Modeling requires insect life-table data, which are collected in autoecological temperature studies at constant and fluctuating temperatures for which insect rearing systems are established and maintained. Control thresholds are defined for targeted IPM interventions (Kroschel 1995). The assessment of intraspecific communication within pest species (pheromones) as well as that of the pest migration capacities and spatial dispersal is fundamental for developing innovative systems for effective pest management (e.g. attract-and-kill, physical control) (Rios and Kroschel 2011; Kroschel and Zegarra 2010; Kroschel et al. 2009). We are aiming at developing environmentally sound pest-management strategies based on manipulating the self-regulating capacity of agro-ecosystems through conservation (adaptation of cultural practices) and augmentation (inoculative/inundative) strategies for pests’ natural antagonists, and biopesticide approaches (entomopathogens). In order to achieve this, an overall understanding and analysis of the antagonists’ biology and ecology is essential, including assessments of their efficacy and the development of systems for formulation and application. At farm level, achieving a compatibility of control methods for different economic pests is important. Interdisciplinary and participatory research is conducted to learn lessons with farmers for adapting innovative technologies to farmers’ situations. Ideally, farm communities are involved, which is often the case in the Andes. For the purpose of up-scaling IPM technologies and outreach, appropriate extension materials and training courses for farmers and extension staff are developed and conducted in collaboration with national programs (Kroschel et al. 2011b).
4 Experiences and Innovations in Potato IPM for the Andean Highlands
4.1 Sustaining the Resilience of Potato Agroecosystems
Maintaining and promoting the natural antagonistic potential is a major element of IPM to stabilize agroecosystems and reduce the incidence of pests. For controlling potato pests, a large number of entomophagous species in families Aphidiidae, Syrphidae, Carabidae, Coccinellidae and Chrysopidae are important natural limiting factors in Europe (Hassan 1989). In developing countries, little information is available as yet on existing beneficial insects present in the wide range of agroecosystems in which the potato is produced. In potato production in the Republic of Yemen, where there is little or no use of insecticides, there are good indications that due to a large number of predators (25) and several parasitoids, the agro-ecosystem functions in a self-regulating manner, keeping aphids (Myzus persicea Sulz., Macrosiphum euphorbiae Thomas) or the cutworm Agrotis ipsilon Hfn. under the control threshold (Kroschel 1995).
The landscape structure and plant diversity play an important role in the conservation and enhancement of natural enemies to keep pest populations under the control threshold. Agroecosystems with a higher structural diversity showed, in most cases, an increased interaction between pests and natural enemies, and provided a higher potential for self-regulation (Tscharntke et al. 2005). Natural vegetation can effectively support beneficial arthropod species (Gurr et al. 2005; Landis et al. 2000) and arable weeds provide important resources for natural enemies such as nectar, pollen and alternative preys and hosts. A possible way to reintroduce insect biodiversity into large-scale monocultures is by establishing vegetation-diverse field margins and/or hedgerows, which may serve as biological corridors (Altieri 1999). Agricultural practices including insecticide application act as disturbances, reducing natural enemy populations and species richness (Croft 1990). Understanding the impacts of common agricultural practices on natural enemies is critical in developing ecologically based pest management (Landis et al. 2000).
For the Andean highlands of Peru, several beneficial insect predators and parasitoids that could play an important role in the control of potato pests have been reported. These include predators in families Carabidae, Coccinelidae, Nabidae, Lygaeidae, Chrysopidae, and Syrphidae (Cisneros 1995). The carabids Harpalus turmalinus Van Emden and Notiobia schmusei Van Emden have been reported as predators of Andean potato weevils (Loza and Bravo 2001; Cisneros 1995); and recently, Kroschel et al. (2009) reported high numbers of carabids of the genera Blennidus, Metius, Pelmatellus, Incagonum, and Notiobia peruviana (Dej.) in the central highlands of Peru that greatly affect the Andean potato weevil. Apanteles subandinus Blanchard, Dolichogenidea gelechiidivoris (Marsh), Copidosoma koehleri Blanchard, and Incamyia cuzcensis Townsend are parasitoids of the potato tuber moths; Gonia peruviana Townsend and Patelloa robusta (Wied.) are larval parasitoids of cutworms (Cisneros 1995).
As the potato is often a pesticide-intensive crop, it is very difficult to gather information on the natural control of potato pests in relation to the natural habitat. To better understand and exploit the potential of habitat management for the promotion and natural conservation of beneficial insects, we started in-depth ecological studies in the Andes of the Mantaro Valley as well as in the Canete Valley in the central coastal region of Peru (see Sect. 25.5.1) to develop practical recommendations for IPM.
4.1.1 The Impact of Insecticides
To explore the effects of insecticide applications on potato pests and natural enemies, ten potato fields were equally divided into insecticide-treated (farmers’ practice) Production Systems of Peru and insecticide-untreated field plots (>1,500 m2) in an altitude gradient between 2,800 and 3,850 m of the Mantaro Valley and the region of Huasahuasi, in the central highlands of Peru (Kroschel and Cañedo 2009). Active and passive evaluation methods were used to monitor arthropods. Phytophagous insects were the most numerous functional groups, accounting for 75.4% of total arthropods. Predatory insects and parasitoids represented 23% and 1.4% of the total insect population, respectively (Table 25.1).
Of 36 identified phytophagous insects, 23 species are phytophagous on potato. Among 23 predator species, ground-dwelling predators were the most numerous (21.7%); plant-inhabiting predatory species represented only 1.3% of the total insect population. Although a total of 16 parasitoids were identified, parasitism of potato tuber moths was low (0.4%). Insecticide treatments significantly reduced about 50% of the arthropod population compared to untreated fields (Table 25.1).
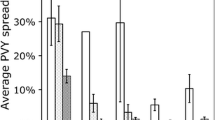
In fields not treated with insecticides, tuber damage by Andean potato weevils and potato tuber moths ranged between 74–95% and 5–35%, respectively. Very striking were the higher infestations with flea beetles (Epitrix yanazara Bechyné), with up to 70% of potato foliar damage causing yield losses of up to 72% (Fig. 25.2, data are shown for the Mantaro Valley only). Insecticide applications were not always effective in controlling Andean potato weevil tuber infestations ranging between 20% and 70%.

Tuber infestation by the potato tuber moth complex, Phthorimaea operculella and Symmetrischem tangolias, and the Andean potato weevil, Premnotrypes suturicallus (a), and total potato yield (b) in five insecticide-treated (I) and non-treated fields (C) in the potato growing regions of the Mantaro Valley. *P ≤ 0.05
4.1.2 The Impact of the Structural Diversity
To identify and to understand plant-beneficial insect interactions and the contribution of plant biodiversity to natural biological control, an inventory of the flora and of arthropods (pests and natural enemies) was made in complex and simple structured landscapes in potato cropping systems of the central Andean highlands at two altitudes (3,250 and 3,850 m a.s.l.). The vegetation composition of grasses, herbs, shrubs and trees was assessed on representative areas as well as arable weeds in potato. Active and passive evaluation methods were used to monitor arthropods.
A total number of 122 plant species comprising grasses (22), herbs (79), shrubs (16) and trees (5) were identified, the distribution of which was highly influenced by altitude: 51% and 42% of the plants occurred only at 3,250 and 3,850 m a.s.l, respectively, with 7% species common in potato systems at both altitudes. There was a clearly higher plant diversity of herbs and shrubs at 3,250 m a.s.l. with a high species diversity of the families Asteraceae, Fabaceae and Brassicaceae. Brassicaceae species (7) were found at 3,250 m a.s.l. only. In contrast, trees were more diverse (5 species) at 3,850 m (e.g., Alnus acuminate Kunth) than at 3,250 m a.s.l. (2 species, e.g., Eucalyptus globules Labill.) (Cañedo et al. 2010).
A total number of 38 weed species were identified, 61% and 29% occurred at 3,250 and 3,850 m a.s.l., respectively, with only 1% found at both altitudes. The most representative families were Poaceae at 3,850 m, and Brassicaceae and Asteraceae at 3,250 m a.s.l. In complex landscapes, Veronica persicae Poiret was predominant during the whole potato cropping period; in contrast, in simple landscapes, Amaranthus hybridus L., Brassica rapa subsp. campestris L., and Chenopodium murale L. were most common in potato at 3,250 m a.s.l.
Thirty-nine plant species were identified as important sources of food for predators, parasitoids and pollinators of 11 insect families. Senecio rudbeckiaefolius Meyen & Walp., distributed at both altitudes, hosted the highest number of parasitoids (7) and predators (13); however, because of their high abundance, plants of the families Asteraceae (e.g., Baccharis penthandii, 4 parasitoids and 3 predators at 3,250 m or Bidens andicola Kunth, 3 parasitoids and 3 predators at 3,850 m a.s.l.) and Brassicaceae (e.g., B. rapa subsp. campestris, 1 parasitoid and 3 predators at 3,250 m) are more important as entomophilous plants attractive to insects.
Phytophagous insects had a significantly higher abundance in simple than in complex landscapes at both altitudes. P. suturicallus dominated in the simple landscape at 3,850 m a.s.l., while E. yanazara was dominant at 3,250 m a.s.l. in the simple landscape. The abundance of parasitoids (3.4%) was very low, but we found 99 species from 10 families; the families Tachinidae (39) and Ichneumonidae (30) were the most representative ones. Predators (17.2% of all insects collected) comprised 75 species; carabids were dominant in the complex landscape at both altitudes. The main carabid species were Pelmatellus columbianus (Reiche), Pelmatellus sp., Blennidus mateui Straneo and Blennidus sp. at 3,850 m a.s.l. and Incagonum sp. (near chilense) at 3,250 m a.s.l. in complex landscapes; unlike Notiobia sp., which was more abundant in simple structured landscapes at 3,250 m a.s.l. At an altitude of 3,250 m, the higher diversity of plants, especially of the families Asteraceae and Brassicaceae, in the surrounding vegetation of potato fields as well as within fields, supported the abundance and diversity of natural enemies more than the vegetation of shrubs and trees forming the complex landscape structure (Cañedo et al. 2010).
4.2 The Development of Technological Innovations
Technological innovations are important to replace insecticide applications by other means of control that are equally or more efficacious at the same or lower costs for farmers. In our research several opportunities for innovations were identified, which could be developed and tested under various agroecological conditions of the Andean highlands.
4.2.1 Plastic Barriers for Andean Potato Weevil Management
Andean potato weevils are flightless and the knowledge of the migratory behavior of weevils, which walk long distances from the previous year’s potato fields to new potato fields, supported our studies to develop simple physical barriers to prevent the migration of these weevils to potato fields (Kroschel et al. 2009). This was further supported by our findings that potato fields are the main source for new weevil infestation and that weevils do not occur – or only in negligible numbers – on fallows (Rios and Kroschel 2011).
In most cases, participatory research with farmers starts after technologies have been developed and tested for many years under laboratory and controlled experimental field conditions. Hence, feedback is seldom received from the end users at the start of the research process regarding the technology already under development; and this has probably often been the cause for low adoption rates of innovations. In our participatory research of testing plastic barriers for Andean potato weevil management, we included farmers from two Andean villages in the evaluation of the technology. Experiments under controlled conditions were conducted merely to prove that adult weevils are unable to climb plastic barriers, and to establish the appropriate height for practical field evaluation. The technology, whereby plastic sheets are fixed to wooden stakes and installed around potato fields (10 cm below and 40–50 cm above soil surface), has been evaluated over several seasons. It proved more effective than several insecticide applications by farmers, where potato is grown in rotation with other crops or after fallow. Ideally, to fully prevent migration a plastic barrier needs to be installed at the time of sowing, or even earlier, but in any case no later than the end of October or beginning of November. In cases where farmers grow potato after potato, plastic barriers prevent new migration, although the potato fields are already a source of infestation. But under these conditions, plastic barriers also proved to be an advantage, since only one initial weevil population needed chemical control, which made it possible to reduce the insecticide applications to only one application (Kroschel et al. 2009). In the High Andes above 3,800 m a.s.l., where Andean potato weevils are the only economic biotic constraint, potato production without the use of insecticides seems to become possible with this simple technology, especially in potato rotational cropping systems (see Sect. 25.4.3). Further, plastic barriers also proved to be effective in controlling other weevil species, such as P. solaniperda Kuschel, P. latithorax and P. vorax in potato agroecologies of Peru, Bolivia, and Ecuador.
4.2.2 Attract-and-Kill for Field and Storage Management of the Potato Tuber Moth Complex
Sex pheromones have been identified for the potato tuber moths, P. operculella and S. tangolias. Since then, they have been mainly deployed for monitoring and studying the flight activity of males, but for practical and economic reasons they have not been used in managing the pests. Attract-and-kill was an innovation successfully developed for controlling other lepidopteran pests (Lösel et al. 2000), a strategy consisting of an insecticide-pheromone co-formulation whereby the male moths are attracted by the pheromone and killed through contact with the insecticide. Considering the small sizes of potato fields in the Andes and in other developing countries, we adapted this technology to be used for the two potato tuber moth species. Attract-and-kill resulted in a 100% kill of both moth species after 3 and 4 days under controlled conditions (Kroschel and Zegarra 2010). In field experiments, at a droplet volume of 100 μl and a droplet density of one drop per 4 m2, equal to 2,500 droplets ha−1, immediately 1 day after the attract-and-kill treatment, daily catches of male moths of either P. operculella or S. tangolias were drastically reduced (>95%) in the pheromone water traps. In contrast, catches continued to be high in the pheromone water traps set up in the respective control fields. Male catches generally remained very low over a long period of time after application (>70 days). At the final evaluation, the total male populations of both P. operculella and S. tangolias were reduced between 90% and 98%, respectively, compared to untreated potato fields (Table 25.2).
Attract-and-kill also proved highly efficacious in reducing the infestation by P. operculella and S. tangolias in potato stores. After storage period of 75 days, the infestation rate and intensity in potato tubers was reduced between 84% and 88% (Table 25.3).
4.2.3 Biological Control
4.2.3.1 Re-naturalization of Lost Biological Control Agents
For decades the frequent use of insecticides has been the farmers’ main practice for managing potato pests in the Andean highlands (Ewell et al. 1990). Considering all study sites, insecticide treatments reduced approximately 50% of the arthropod population compared to non-treated fields within one crop cycle (see Sect. 25.4.1.1). Since the potato is the most widely grown crop, especially at altitudes above 3,800 m, the regular use of insecticides may have caused long-term negative effects on natural enemy populations, especially where no effective recovery from unsprayed fields or hedgerows was given; however, no earlier reference base line studies exist that could provide more evidence for this assumption.
Long-term field studies in the United Kingdom in a range of arable crops found few adverse long-term effects of pesticides on non-target organisms including insects, spiders, earthworms and soil microbes (Young et al. 2001). Here, the application of broad-spectrum insecticides resulted in declines in the number of many non-target arthropods, but these usually recovered within the same growing season. Less temporary effects were seldom noted and affected only soil-dwelling collembolans (springtails). Numbers of these organisms remained comparatively low in treated plots up to 2 years after application (Devine and Furlong 2007). Apart from the persistence of the insecticide, the degree to which affected populations can recover is also dependent upon the recruitment of new individuals from unaffected populations, which often permit the rapid recovery of species in insecticide-treated fields (Jepson and Thacker 1990).
From our research we learned that the landscape structure in the High Andes does not provide effective niches for natural enemies to recuperate, and annual plants (weeds) surrounding potato fields were more supportive than species-poor hedgerows. Further, pest-specific parasitoids, e.g. those of the potato tuber moth complex, can probably not build up sufficient populations due to the use of broad-spectrum insecticide applications, which was also the limiting factor after the introduction of parasitoids for the classical biological control of P. operculella in Australia (Horne and Page 2008). The introduction of plastic barriers as an effective technology to control the Andean potato weevil gives the opportunity to introduce a new IPM program in the Andes, which may fully replace – or will at least drastically reduce – insecticide interventions. This could be accompanied by augmentation strategies for natural parasitoids of the potato tuber moth complex, whose populations were reduced or partly lost through the intensive use of pesticides, to further increase the resilience of potato agroecosystems. For this purpose, CIP maintains and supplies to national programs parasitoids such as Copidosoma koehleri Blanchard, Apanteles subandinus Blanchard, and Orgilus lepidus Muesebeck, which have co-evolved with the potato tuber moth.
4.2.3.2 Bacillus thuringiensis for Low-Cost Storage Management of the Potato Tuber Moth Complex
Bacillus thuringiensis subspec. kurstaki (Btk) was known to be highly effective against P. operculella, but it has hardly been used by small-scale potato farmers because of its relatively high cost. A very low proportion of Btk, 40 g Btk mixed with 960 g fine sand dust containing quartz provided effective storage control for 1 ton of stored potato in the Republic of Yemen (Kroschel and Koch 1994). Alternatively, P. operculella granulovirus (PhopGV) was produced nationally in Peru and Bolivia using low-cost facilities for propagation. A dust formulation, produced by selecting and grinding PhopGV-infected larvae mixed with ordinary talcum, has been used at the rate of 5 kg per ton of stored potatoes (20 infected larvae per kilogram of talcum). Research showed that PhopGV reduced damage in stores by 91% and 78%, 30 and 60 days after application (Raman and Alcázar 1990). However, the pest situation has changed in the High Andes and S. tangolias has become the most prevalent species in potato stores (Keller 2003) for which PhopGV is not effective; it has not been possible to identify a S. tangolias-specific granulovirus. Winters and Fano (1997) pointed out that the market potential of a bioinsecticide directed to only one species of the potato tuber moth complex may limit its potential use and adoption. We found that the commercial product Dipel2X reformulated in magnesium silicate (15 g/1 kg talcum) effectively protects tubers against both tuber moth species, which makes the product highly competitive compared to the current PhopGV-product or to chemical pesticides. Since farmers have adopted the use of a dusted PhopGV formulation to treat potatoes prior to storage, the probability for adoption of a specific Btk-talcum formulation, which controls all species, is high. However, for effective management of potato tuber moths in rustic stores, storage hygiene is also very important. In particular, physical protection is needed to prevent new moths from entering storage facilities and thereby prevent the infestation of tubers through young unprotected potato sprouts.
4.2.3.3 Potential of Entomopathogens
Many studies have been conducted to assess the pathogenicity and practical use of entomopathogenic fungi for the control of Andean potato weevils (Kaya et al. 2009). Kühne (2007) demonstrated that Beauveria bassiana (Balsamo) Vuillmen kills P. suturicallus under laboratory conditions but is not effective when applied in the field, mainly due to the low temperatures at high elevations. In contrast, Cisneros and Vera (2001) obtained good results applying the fungus against weevil larvae under potato tuber storage conditions. However, Winters and Fano (1997) concluded that the benefits of using a biocontrol product based on B. bassiana for Andean potato weevil storage control are low, and farmers are not expected to purchase the product in sufficient quantities to allow a market to develop.
Alcázar and Kaya (2003) isolated an undescribed entomopathogenic nematode in the genus Heterorhabditis, designated as strain Alcázar-1, from last instar P. suturicallus larvae in soil from a commercial potato storage at 2,750 m a.s.l. Parsa et al. (2006) studied the biological activity and demonstrated its high efficacy to infect all stages of P. suturicallus and to protect potato tubers from infestation by neonate larvae; further, it showed adaptability to low temperatures. In field experiments, applied in suspension at a rate of 50 infective juveniles/cm2, the entomopathogenic nematode significantly reduced tuber damage and plant infestation by Andean potato weevil larvae compared to the control; tuber damage was reduced by 41.4% and the larval infestation by 53.2%, respectively (Alcazar et al. 2007). To commercialize entomopathogenic nematodes requires investments in large-scale production facilities, which may not yet be economically viable for small-scale farmers; however, small-holder potato production systems of the high Andes could profit from a wider use of nematode-based bioinsecticides in high value crops, e.g., asparagus (Asparagus officinalis L.) along the Peruvian coast.
4.3 Ecological and Economic Benefits of IPM
With regard to the altitude where potato is produced and the temperature-dependent related abundance of insect pests, our studies concluded with two IPM approaches for the Andean Highlands. At altitudes >3,800 m, where Andean potato weevils are the only insect pest problem, the use of plastic barriers may constitute the main IPM technology. In contrast, in inner Andean valleys such as the Mantaro Valley (3,200–3,500 m a.s.l.), in addition to Andean potato weevils potato tuber moths and flea beetles may cause economic damage and losses. Based on our ecological studies different strategies and technological innovations need to be integrated in a pest management program. We evaluated the ecological and economic benefits of the two IPM systems versus farmers’ practice.
With the participation of 40 farmers (i.e., 40 individual on-farm field evaluations) of the villages Ñuñunhuayo (3,800 m a.s.l.) and Aymara (3,900 m a.s.l.), the use of plastic barriers proved effective in reducing Andean potato weevil tuber infestation. Compared to farmers’ practice using 3–4 insecticide applications per season and a tuber infestation of 18.8%, the barriers reduced the infestation by 68% to a total of 6% at potato harvest (Table 25.4).
The EIQ (Environmental Impact Quotient) is a method for measuring the impact of pesticides on the environment. EIQ values ranging from 0 to 20 indicate a low environmental impact; from 20.1 to 40, medium; and >40 high environmental impact (Mazlan and Munford 2005). Because no insecticides were applied in IPM, the environmental impact (EI) of insect pest management was accordingly zero, while farmers’ practice resulted in an EI of 122.7. Finally, IPM resulted in an excellent investment with mean increase in income of about US$600 for farmers of two Andean villages.
In the Mantaro Valley, IPM for potato was tested against farmers’ practice and an untreated control at four locations. The IPM system included the installation of plastic barriers to control Andean potato weevils, the application of attract-and-kill to control the two potato tuber moth species P. operculella and S. tangolias, intercropping of potato with faba bean (Vicia faba L.) and the establishment of field borders (Brassica rapa var. campestris) to increase the overall functional diversity of natural enemies to reduce the incidence of other pests such as flea beetles or aphids (M. persicae, M. euphorbiae). Further, one insecticide application (beta-cyfluthrin) was considered for flea beetle control. In contrast, farmers’ practice included four insecticide applications (3x carbofuran, 1x methamidophos).
Plant damage caused by flea beetles was significantly reduced in the IPM treated fields compared to farmers’ practice and untreated control (Table 25.5). Tuber damage caused by Andean potato weevils was similar in farmers’ practice (1.9%) and IPM (4.7%) plots, but significantly reduced compared to the untreated control (31.3%). Attract-and-kill significantly reduced the flight activity of both potato tuber moth species as well as the infestation of potato plants and tubers at harvest. In the IPM system, one application of beta-cyfluthrin (EIQ = 31.57) was made, while farmers used three applications of carbofuran (EIQ = 50.67) and one application of methamidophos (EIQ = 36.83). Therefore, IPM showed high ecological benefits with an EI of only 8.3 compared to farmers’ practice with an EI of 129.5.
In all IPM plots, there was a positive interaction between the growth of potato and faba bean, demonstrated by land equivalent ratio (LER) values of 1.39–2.52. However, potato intercropping with faba bean or the weed B. rapa var. campestris failed to increase the biological control of pests and the number of parasitoids and predators. Hence, yield calculations were made for the potato area only (Table 25.5).
The higher pesticide costs increased the total potato production costs in farmers’ practice plots. However, due to insignificant higher yields and lower tuber infestations, farmers’ practice and IPM plots showed no differences in total net income. In contrast, clear differences of both systems with the untreated control occurred, demonstrating the overall need for an effective pest management system in potato. It has to be considered that calculations do not include health related costs of the use of highly toxic pesticides, which are another important social cost factor.
5 Experiences and Innovations in Potato IPM for the Coastal Lowlands
5.1 Sustaining the Resilience of Potato Agroecosystems
The leafminer fly (L. huidobrensis ) is regulated by numerous hymenoptera parasitoids, some of which were successfully used in biological control programs (Murphy and LaSalle 1999). Likewise, along the Peruvian coast a high diversity of 42 parasitoids were identified on L. huidobrensis, and the most abundant species are the ectoparasitoid Diglyphus websteri (Crawford) (Eulophidae) and the endoparasitoids Halticoptera arduine (Walter) (Pteromalidae) and Chrysocharis caribea Boucek (Eulophidae) (Mujica and Kroschel 2011). For the bud midge (Prodiplosis longifila) no efficient parasitoids have been detected. Only the parasitoid Synopeas sp. (Platygasteridae) has been recovered from asparagus in the north coast with a parasitism that hardly exceeds 20% (Cisneros 1995). Because both the leafminer fly and the bud midge pupate in the soil, ground-dwelling predators play an important role in the regulation of populations of these insects. Generalist predators are regarded as an indispensable complement to specialized parasitoids, buffering outbreaks of pests, when populations of parasitoids and oligophagus predators are still low (Riechert and Lawrence 1997). Only few studies on soil dwelling predators in agroecosystems of the central coast of Peru have been conducted, showing the dominance of spiders (Araneae), ground beetles (Carabidae, 4 spp.), rove beetles (Staphylinidae, 16 spp,) and earwigs (Labiduridae, e.g. Labiduria riparia, and Anisolabidae) in potato and other agricultural crops (Schuller and Sanchez 2003; Rondon and Vergara 2003).
5.1.1 The Impact of Insecticides
The effect of broad-spectrum (farmers’ practice) and selective insecticide applications in potato in comparison to untreated fields was studied on the leafminer infestation and the abundance and diversity of its natural enemies. Selective insecticides significantly reduced leafminer fly larvae infestation (75%), compared to the use of broad-spectrum insecticides (35%) (Fig. 25.3a). Populations of the two endoparasitoids H. arduine and C. caribea were greatly reduced by >80% by either kind of insecticide. In the arthropod community, Isopods were the most sensitive to the applications of insecticides and were reduced by 58.7% and 89.9% by broad-spectrum and selective insecticides, respectively. Species of the orders Coleoptera and Hymenoptera were most abundant in non-treated (46.6% and 22.3%) and selective insecticide (50.8% and 15.6%) treated plots; however, they were greatly reduced by 86% and 78% through the application of broad-spectrum insecticides. In contrast, in farmers’ fields Collembolla (64.3%) and Diptera (16.8%) were the most abundant orders. Broad-spectrum insecticides also significantly reduced the density of ground-beetles (Carabidae) (95%), earwigs (Labiduridae and Anisolabidae) (90%) and spiders (Aranea) (49.2%) (Fig. 25.3b), but no differences were found between non-insecticide and selective insecticide treated plots. The density of rove beetles was not affected by any treatment. The use of selective insecticides constitutes a particularly effective conservation biological control strategy, because it combines direct targeted reduction in pest numbers with predator conservation.

Effect of selective and broad-spectrum (farmers’ practice) insecticides on (a) leafminer infestation and parasitism, and on (b) the abundance of ground-dwelling arthropods and spiders, in the Cañete Valley, Peru
5.1.2 The Impact of the Structural Diversity
The landscape structure is a key factor for maintaining the abundance, community structure, and effectiveness of natural enemies in agricultural landscapes. The lower Cañete Valley (Ihuanco and Cañete) was characterized based on vegetation diversity and percent of arable land. The region of Cañete represents 81.6% of arable land associated to large-scale monoculture and a structurally simple landscape; instead in Ihuanco, arable land covers only 20.9% and most of the non-cropped and structurally complex landscape was made up of temporary herbaceous vegetation (72.3%). Although no differences in the vegetation diversity were found between the two landscapes, a higher percentage of field margins and a higher abundance of trees and shrubs were found in the complex landscape.
Leafminer infestation in potato and parasitism of leafminer fly larvae were not affected by the landscape structure, although a higher abundance and diversity of leafminer fly parasitoids was found in the complex landscape. Neither did the landscape structure affect the relative abundance of the species H. arduine and D. websteri, which are the most important parasitoid species, representing 72.4% and 17.9% of the total parasitoid population, respectively. The arthropod community composition was also similar between landscapes. Hexapoda was the dominant taxa in complex (81%) and simple (90%) landscapes, but Isopoda was more abundant in the complex landscape. By contrast, the insect community structure varied greatly between the two agricultural landscapes. Hymenoptera, for instance, was more abundant in the complex (72%) than in the simple (17%) landscape. Ground-dwelling predators were primarily ground beetles (Carabidae), rove beetles (Staphylinidae), earwigs (Labiduridae) and spiders (Aranea) representing 60.2%, 9.5%, 6.6% and 23.7%, respectively. Densities of each group of predators differed significantly between the two types of landscapes. In the complex landscape, Aranea and Carabidae were the most abundant predators, with 42.8% and 38.5% respectively. In the simple landscape Carabidae (71.8%) was the dominant predator group. The Staphylinidae family included 14 species and was the most species-rich family compared to Carabidae with six and one species for the Labiduridae and Anisolabidae families. The dominant species was the carabid Pterostichus sp. with 59.6% of total predators captured, and abundances of 71.3% and 37.7% in the simple and complex landscape, respectively. The Aranea order was represented by 10 families and 10 morphospecies. In general, for ground-dwelling predators and for all four predator families, species diversity (richness, Shannon and Simpson index) was higher in the complex than in the simple landscape, but a higher activity and abundance of Carabidae and Dermaptera were observed in the simple landscape.
5.1.3 The Impact of Weeds
In the central Peruvian coastal area, we assessed leafminer fly-parasitoid relationships in arable weeds. Twenty-two weed species of 14 plant families were identified as hosts of 11 leafminer fly species. Liriomyza sabaziae Spencer (58.7%) was the dominant species followed by L. huidobrensis (13.6%); however, only L. huidobrensis is regarded as an economically important pest, concluding that weeds associated to potato are mainly a source of leafminer flies with non-economic importance. The weeds Galinsoga parviflora Cav. (host of L. sabaziae), Malva parviflora L. [host of Calycomyza malvae (Burgess)], and Commelina fasciculata R&P [host of Liriomyza commelina (Frost)] are non-hosts of L. huidobrensis, but leafminer flies infesting these species shared the highest number of common parasitoids. On average, parasitism of leafminer flies in weeds reached 48.9% by a total of 50 hymenoptera parasitoids, of which Chrysocharis caribea Boucek, C. flacilla, Halticoptera arduine, Diglyphus begini (Ashmead) and Diglyphus websteri were the most important. In potato, in contrast, L. huidobrensis was parasitized by five parasitoid species only. Considering parasitoid richness, abundance and diversity, L. sabaziae, Cerodontha dorsalis (Loew), L. commelina and C. malvae were the main parasitoid sources for leafminer fly. A rational management of arable weeds associated with potato could therefore support the augmentation of parasitoids and natural control of L. huidobrensis. We tested this hypothesis by establishing intercropping strips of the weed G. parviflora in potato (see Sect. 25.5.2.3). However, weeds can also be sources for other insect pests and diseases of potato which need to be taken into consideration as suggested by Altieri (1999) and Kroschel (1995).
5.2 The Development of Technological Innovations
5.2.1 Trapping Devices
The strong attraction of leafminer flies to yellow surfaces was reported in the early 1980s, and, covered with a sticky substance, these were used for trapping and monitoring adults. CIP tested, optimized and adapted the use of yellow sticky traps for the mass trapping of leafminer flies in potato. Fixed traps (50 × 50 cm) were set up at plant emergence, and after becoming saturated with flies they were replaced at flowering stage. After flowering, the flight activity of the leafminer fly decreased and captures were substantially reduced. With the use of 80 fixed yellow sticky traps per hectare, about 5 million insects were recorded in one potato season representing 91.9% adults of leafminer flies and only 8.1% parasitoids, respectively. Highest captures (80%) were made from potato hilling to potato flowering, which were the recommended stages to use traps. In an evaluation of three trap densities of 60, 80 and 100 traps/ha with two replacements during the cropping season, an average of 1.15, 1.60 and 1.81 millions of adults were trapped, respectively (Mujica et al. 2000). A further development was the use of mobile yellow sticky traps (4 × 1 m) that are passed over the plant canopy covering four or more rows at a time. Farmers adopted and modified them according to their ingenuity. Comparing the efficacy of a combination of 95 fixed traps/ha (one change/cropping season) and 9 mobile trap passages with chemical leafminer fly control, a cumulative capture of 7.28 million adults/ha led to a reduction of 66.7% of the control costs (US$66.7/ha) compared to chemical control (US$200/ha) only with a total of 6 insecticide applications per season. Farmers’ use of yellow sticky traps reduced leafminer fly adult populations effectively and made insecticide sprayings unnecessary to control adults, which reduced costs and did not destroy natural enemies at an early stage of plant growth (Mujica et al. 2000). An impact evaluation of the use of trapping devices in the Cañete valley showed that 90% of farmers reduced insecticide applications. The greatest economic benefits (US$200/ha) were observed when farmers used the combination of fixed and mobile traps. Trapping can replace the use of insecticides; however, this might not be fully effective to prevent the development and mining of larvae in the potato foliage as well as yield reductions, especially if the control by natural enemies is limited. In this case, applications of selective insecticides become necessary.
5.2.2 Selective Insecticides
When insecticide applications are unavoidable they should be used as selectively as possible. Several studies have been conducted on the effect of different groups of insecticides on leafminer larvae and their parasitoids. Cyromazine is an effective and selective larvicide with no effects on adults of leafminer flies or parasitoids. However, applied before and after oviposition of the parasitoid H. arduine it produced high mortality of leafminer larvae (>70%), but did not allow the hatching of parasitoids or development of neonate parasitoid larvae. When cyromazine was applied to leafminer larvae already parasitized by H. arduine (larvae already developed), leafminer fly larvae were killed but parasitoid larvae survived. Abamectine, which is a selective but expensive product, deriving from a soil fungus, affects the feeding behavior and oviposition of leafminer fly adults and also kills first instar larvae of the leafminer fly; its effect decreases as larvae grow. It showed no detrimental effects on populations of leafminer parasitoids (CIP 2004). Under field conditions, adding vegetable oil to abamectin increased its efficacy and reduced treatment costs by 73.5% and 74.2% as compared to the use of abamectin alone (Mujica et al. 2001). Imidacloprid was not effective against leafminer fly adults, but killed the parasitoid H. arduine. Pyrethroids (cypermethrine; cyfluthrine and b-cyfluthrine) affected both leafminer fly adults and parasitoids (CIP 2004). Other translaminar compounds (bensultap, cartap, chlorfluazuron, spinosad) significantly affected the leafminer larvae development. Parasitoids, parasitism and species richness were affected by the application of most pesticides; ectoparasitoids being the most susceptible.
5.2.3 Intercropping System
An increasing complexity of a landscape or cropping system can enhance the functional biodiversity in agroecosystems and can therefore positively contribute to the natural regulation of pests. Potato, as host plant of Liriomyza huidobrensis, was intercropped with conservation strips of maize (Zea mays L.) as host of Liriomyza graminivora (Korytkowski), or the weed Galinsoga parviflora as host of L. sabaziae, both in complex (Cañete Valley) and simple (Ihuanco Valley) landscapes. The structurally complex landscape was associated with a reduced leafminer larvae infestation and leaf damage and also with higher parasitism (Fig. 25.4). In the potato-maize intercropping system a minor larvae infestation, leaf damage and increased parasitism were observed compared to the potato-weed intercropping system. Intercropping strips with the weed G. parviflora or maize contributed to a diversification at the landscape scale, but differences in infestation and parasitism were more evident in the simple than in the complex landscape. Therefore, potato pest management will benefit from intercropping systems mostly in structurally poor agricultural landscapes, in which the leafminer fly is a main pest problem.

Mean variation of Liriomyza huidobrensis larvae infestation, foliar damage, and parasitism affected by two intercropping systems in complex and simple structured landscapes in the Cañete Valley, Peru
Ground-dwelling predators were dominated by the carabid Pterostichus sp. followed by rove beetles and earwigs. Although the seasonal activity pattern of these beneficial species varied spatially and temporally, they consistently preferred potato as compared to the intercropping strips. Predators responded differently to intercropping systems and the landscape context, showing that these variables did not influence all species equally, and interacting communities were made up of species with different spatial strategies. Biological diversity and ecological functions are not only affected by the overall landscape characteristics but also by the cropping systems, and must be considered to improve agro-ecosystems for biodiversity conservation and biological control of the leafminer fly (Schmidt et al. 2004). According to our results, natural control could benefit from early planting of conservation strips (at least 30 days before potato planting) to allow colonization by natural enemies, which later should be disturbed to facilitate the movement of natural enemies towards the crop.
5.2.4 Biological Control Using Entomopathogens
In 2003, natural epizootics of the Hypocreales fungus Isaria fumosorosea (Wize) were found in L. huidobrensis adult populations. Bioassays proved the pathogenicity against adults but not to other life stages. In preliminary field experiments, the application of 1.29 × 1012 conidia ml−1 of the isolate CIP-LMF II reduced the leafminer fly adult population by 68.1% within 8 days compared to population increases by 144% in untreated controls. In order to assess the efficacy of the fungus to cause mortality on a known adult population of L. huidobrensis, semi-field experiments with potatoes grown in screen houses (4.8 × 6.0 m) covered with fine nylon mesh were conducted. The screen houses were artificially infested with 750 adults (400 females and 350 males). Ten days after 4 and 2 fungal applications of 7.38 × 1012 conidia ml−1 at 2-days and 5-days intervals, an Abott mortality of 47.4% and 40.1% as well as a fungal infection of 21% and 13% of adults was observed. However, the reduction of the initial adult population was not sufficient to reduce leafminer fly leaf infestation, and hence the potato yields in all fungal treatments and the control were not significantly different.
We concluded that the biocontrol of adults would need complementary biological control efforts that control other life stages of the leafminer fly. Entomopathogenic nematodes of the families Steinernematidae and Heterorhabditidae have been considered as potential control agents against leafminers in recent years. Nematodes enter infested plants via leafminer oviposition holes and feeding sites (Harris et al. 1990) attacking the larvae through natural openings (Poinar 1990). Lebeck et al. (1993) found that all larval instars of Liriomyza trifolii (Burgess) were susceptible to Steinernema carpocapsae (Filipjev); Heterorhabditis bacteriophora Poinar also showed good efficacy in controlling this pest (Olthof and Broadbent 1991). We assessed 42 strains of entomopathogenic nematodes against L. huidobrensis and found large differences in their efficacy. The Heterorhabditis sp. isolate 3712 showed the highest efficacy to induce larvae mortality between 90–100% within a temperature range of 15–25°C. Its application on infested leaves of potted potato in a concentration of 10,000, 5,000 and 2,500 IJs/ml induced larvae mortality by 83%, 70%, and 55%, respectively. Applied in field experiments on leafminer fly infested faba beans, a mortality of larvae in the lower and middle part of the plants of 69% and 48.3% was achieved. In the upper leaves no mortality of larvae was indicated. The preliminary results are promising but before entomopathogenic nematodes can be considered in an integrated pest management program for the control of L. huidobrensis, further field evaluations, including a combination with the entomopathogenic fungus I. fumosorosea, are needed.
5.3 Ecological and Economic Benefits of IPM
The ecological, environmental, and economic benefits of an IPM program based on the use of action thresholds, trapping devices, and selective insecticides was evaluated for the potato crop in comparison to farmers’ practice in the Cañete Valley of the central coast of Peru. The aim was to evaluate and combine IPM strategies profitable for farmers, but with fewer environmental and human risks.
During the potato season, the leafminer fly L. huidobrensis and the bud midge P. longifila were the most important pests. Mobile traps utilized during the vegetative growth of potato made it possible to reduce the flight activity of the leafminer fly in IPM fields (Table 25.6); further, the larvae infestation of both pests as well as the leafminer foliar damage was significantly reduced through IPM.
The arthropod diversity in sweeping net samples was similar in both pest management systems. In general, the abundance of phytophagous (66.3%, 12 spp.) was higher than that of predators and parasitoids (12.6%, 26 spp.). In farmers’ practice fields the higher abundance of parasitoids was due to a higher leafminer larvae infestation. In IPM fields, the total number of plant-inhabiting predators, ground beetles (Pterostichus sp.) and earwigs (Labiduria riparia), was significantly higher than in farmers’ fields. On average, double the amount of pesticides per season were applied by farmers (16.7 sprays) compared to IPM (8.8 sprays). However, the total amount of pesticides used per season was higher in IPM fields due to the specific control of the bud midge with the insecticide chlorpiriphos (2.5%). Still, the applied IPM strategy reduced the environmental impact compared to farmers’ practice by 70.3%, from 137.4/ha to 40.8/ha, respectively, and gained a 35% higher marketable potato yield. Partial budget analysis showed a clear net profit for IPM, with an average increase in income of US$1410 per farmer.
6 IPM Support Tools
6.1 Insect Simulation Modeling and Climate Change Effects on Pests
Insect life table data, developed under a wide range of temperatures, give good predictions for the best temperature conditions that insects require for optimal growth and development. Based on such life table data, temperature-based phenology models are useful tools to gain an understanding of how temperature affects pest population growth potentials in different agroecologies. CIP develops temperature-based phenology models for potato pests; e.g., for P. operculella, which has been validated both in the laboratory and the field (Sporleder et al. 2004; Keller 2003). By linking this model to Geographic Information Systems (GIS) the pest developmental potential can be visualized for potato regions worldwide in order to support decision-making for appropriate pest management practices (Sporleder et al. 2008).
Global Warming will affect pest abundance and severity in agricultural and horticultural crops, and will most likely increase the yield and quality losses caused by pests. Insect population growth potentials are mainly temperature-driven, and a rise in temperature may either increase or decrease insect development rates and related crop damages depending on the insect species’ optimum temperature range. We used the process-based climatic phenology model for P. operculella and applied three risk indices (establishment index, generation index, and activity index) in a geographic information system (GIS) environment to map and quantify changes for climate-change scenarios of the year 2050 based on downscaled climate-change data of the A1B scenario from the WorldClim database (Kroschel et al. 2011a). All applications and simulations were made using the Insect Life Cycle Modeling (ILCYM) software recently developed by The International Potato Center, Lima, Peru (Sporleder et al. 2011). The study concludes that the P. operculella damage potential will progressively increase in all regions where the pest already prevails today, with an excessive increase in warmer cropping regions of the tropics and subtropics. A range expansion into tropical temperate mountainous regions with a moderate increase of its damage potential is also predicted; i.e., that in Bolivia, Ecuador and Peru 44,322, 9,569, and 39,511 ha of potato will be under new risk of infestation. This information is important to prepare farmers for such change and to develop adaptation strategies to climate change.
7 Technology Transfer
The development of IPM calls for an agroecosystem-/site-specific approach and the most practical research outputs (see CIP’s IPM research framework, Fig. 25.1) need to be combined to define an IPM program that is the most relevant to farmers’ conditions and can have a high rate of adoption. Embedded into the best cultural practices of potato cropping for managing the different potato insect pests by the use of high quality pest-free seed, adequate crop rotation, optimal planting and harvest dates, best practices of weeding and hilling, etc. (Kühne 2007; Kroschel 1995), according to the recent research results, an IPM program for the Andean highlands would ideally consist of new technologies and strategies (i) to inhibit the migration of Andean potato weevils into potato fields by using plastic barriers; (ii) to scout the occurrence of Epitrix spp. and eventually control by one single application of low-toxic insecticides, and (iii) to apply attract-and-kill at a rate of 2,500 droplets/ha at the pre-flowering stage to reduce potato tuber moth tuber infestations (Fig. 25.5a) (Kroschel et al. 2011b). Additional control efforts would be needed for pest management in potato stores by using talcum-Btk or attract-and-kill.

Potato growth stages and most relevant IPM practices for potato croping systems of the Andean highlands (a) and the lowlands (coast) (b) of Peru. These technologies have to be embedded into the best cultural practices for potato which are not included in this sketch. *Application on insecticides might be needed to control flea beetle (Epitrix spp.) at specific locations. LMF = leafminer fly (Liriomyza huidobrensis), BM = bud midge (Prodiplosis longifila)
Likewise, for lowland potato production in the coastal region of Peru, the IPM program would be based on (i) seed treatments, (ii) use of mobile or fixed yellow sticky traps, and (iii) pest scouting for both leafminer fly and bud midge, and the target-specific application of low-toxic, selective insecticides. Mobile and yellow stick traps have been already successfully introduced and are widely adopted by farmers (Fonseca and Mujica 1997; Mujica et al. 2000). Potato storage is not practiced in the lowlands and quality seed derives from highland production regions (Fig. 25.5b). Both IPM systems would also profit from intercropping of potato with other crops, or from establishing field borders of naturally occurring weeds, to increase the floristic diversity and to support natural biological control.
Before the development of plastic barriers, which inhibit the migration of adult weevils to potato fields, insecticide applications were the only direct control method for Andean potato weevil adults to prevent oviposition and tuber damage by larvae. Therefore, previous IPM efforts in the Andes were based on the training of farmers to better understand the weevils’ biology and to use cultural practices appropriately to reduce weevil infestation sources (Ortiz et al. 1996); however, this has achieved a low level of adoption (Ortiz et al. 2009). A better adoption of the proposed new potato IPM program could profit from the high cost-efficacy of plastic barriers and the accessibility to plastic in the local markets. Nevertheless, the wider use and application of all three new IPM technologies – plastic barriers, attract-and-kill, and talcum-Btk – can be achieved only if governmental and non-governmental organizations, as well as the private sector, are interested in the implementation and commercialization of the new products. Training courses have been initiated and the new technologies were well perceived by agricultural extension workers and farmers. Organic potato growers, in particular, have shown immediate interest in adopting plastic barriers with the support of non-governmental organizations. The interest in IPM differs from one village to another, and often depends on how well the farming communities are organized internally. Complementary demonstration trials at the village level will be important so that farmers’ groups can jointly experiment for themselves and gain confidence in the new technologies. Currently, partnerships are being built up with the private sector to introduce and make available plastic in adequate sizes (40 cm wide) and quality (durability of at least 2–3 years), to re-formulate existing B. thuringiensis products with talcum to register and commercialize a low-cost biocontrol product, as well as to register attract-and-kill in Peru. Main limiting factors and constraints for implementing IPM in Peru are the non-existent governmental extension service as well as the heavy competition of agrochemical products to which farmers have easy access through a strong network of pesticide sellers.
Generally, the implementation of IPM is knowledge-demanding and farmers would profit from strong support of experienced advisors with an entomological background, who can adequately assist farmers in their decision making throughout the cropping period. This kind of extension service would require a private IPM potato consultancy. A successful example is given by Horne and Page (2008) for potato production in Australia, which can serve as a model for other countries.
8 Lessons Learned and Conclusions
It is often argued that the international agricultural research centers (IARC) should prioritize research on global or regional plant health problems; in the case of CIP, giving priority to research on regional pests like the Andean potato weevils or globally invasive pests such as potato tuber moths or the leafminer fly. National agricultural research should then adapt technologies to their specific locations. Our lessons from the Andean highlands and coastal lowland potato production systems clearly demonstrated that a system approach is required to develop IPM. This needs to consider all economically important pests in an agroecosystem, i.e., globally minor but locally important pests to be addressed by farmers. In the Andean highlands flea beetles can become more important pests if farmers adopt non-chemical pest management strategies for Andean potato weevils. Developing and applying a system approach in IPM as outlined in CIP’s framework for IPM research and development (Fig. 25.1) requires not only sufficient human capacity but also long-term funding, which in most cases cannot be afforded by national programs.
Research efforts on globally important pests have already generated a lot of information for their management on which basis new and more effective technologies could be developed (such as plastic barriers, attract-and-kill). In contrast, basic information is often missing for local pests such as flea beetles or the bud midge. Therefore, it was not possible to include alternative control methods in our IPM program for these pests, which have to be managed for the time being by the rational use of insecticides. Likewise, it has not so far been possible to adapt highly effective technologies for two potato tuber moth species, such as attract-and-kill, for the control of the Guatemalan tuber moth Tecia solanivora (Povolny), an important potato pest in Colombia and Ecuador.
In our research, agroecological studies were conducted to better understand the impact of farmers’ pest management and of the landscape structure on the abundance and diversity of pests and natural enemies. This revealed important information on how long-term pesticide use might have reduced arthropod populations or how natural biological control could be augmented in the two agroecosystems. Further, such information may be relevant for monitoring changes over time.
On-farm field trials to test IPM versus farmers’ conventional pest management practices demonstrated the environmental and economic viability of IPM programs for both potato cropping systems. Evaluation methods such as ecological assessments of foliage and soil inhabitant arthropods and parasitoids, the use of the environmental impact quotient (EIQ) and partial budgeting proved most practical in assessing the ecological, environmental, and economic impact of the systems under evaluation.
References
Alcazar J, Cisneros F (1999) Taxonomy and bionomics of the Andean potato weevil complex: Premnotrypes spp. and related genera. In: Impact on a changing world. Program Report 1997–1998, International Potato Center (CIP), Peru, pp 141–151
Alcazar J, Kaya H (2003) Hallazgo de un nemátodo nativo del género Heterorhabditis sp., parásito del gorgojo de los Andes Premnotrypes suturicallus en Huasahuasi – Junín. Resumen de la XLV Convención Nacional de Entomología Ayacucho, Peru
Alcazar J, Kroschel J, Kaya H (2007) Evaluation of the efficacy of an indigenous Peruvian entomopathogenic nematode Heterorhabditis sp. to control the Andean Potato weevil Premnotrypes suturicallus Kuschel under field conditions. In: XVI international plant protection congress, 15–18 Oct 2007. Congress proceedings vol II. Glasgow, Scotland, pp 544–545
Alston DG, Reading ME (1998) Factors influencing adoption and educational outreach of integrated pest management. J Ext 36(3)
Altieri M (1999) The ecological role of biodiversity in agroecosystems. Agr Ecosyst Environ 74:19–31
Arica D, Kroschel J, Forbes G, Saint Pere K (2006) Persistent Organic Pollutants (POPs) and Hazardous Pesticides in Andean Farming Communities in Peru. Final report to the Canada POPs Fund, World Bank. International Potato Center, Lima, Peru, 73pp
Cañedo V, Rojas J, Alvarado J, Kroschel J (2010) Efecto de la composición del paisaje sobre las plagas y enemigos naturales de la papa en la sierra central del Perú. In: Proceedings of the XXIV congress of the Latin-American Potato Association (ALAP), 23–28 May 2010, Cusco, Peru, pp 300–301
CIP (2004) Research on selective and nonselective insecticide compounds against Liriomyza huidobrensis and their hymenopteros parasitoids under laboratory and field conditions. Compendium of activity reports from 1997 to 2003. International Potato Center, Peru
Cisneros F (1995) Control de plagas agrícolas. Editorial Impresión, Lima
Cisneros F, Mujica N (1997) Developing IPM components for leafminer fly in the Cañete Valley of Peru. In: International Potato Center (ed) Program report 1995–1996, Lima, Peru, pp 177–184
Cisneros F, Vera A (2001) Mass producing Beauveria brongniartii inoculum, an economical, farm-level method. In: Scientist and farmer: partners in research for the 21st century. Program report, 1999–2000, International Potato Center, Lima, Peru, pp 155–160
Croft BA (1990) Arthropod biological control agents and pesticides. Wiley, New York
Daxl R, von Kayserlingk N, Klein-Koch C, Link R, Waibel H (1994) Integrated pest management: guidelines. Deutsche Gesellschaft für Technische Zusammenarbeit (GTZ) GmbH, Schriftenreihe der GTZ No. 249, Eschborn, Germany, 119pp
Devine GJ, Furlong MJ (2007) Insecticide use: contexts and ecological consequences. Agric Human Values 24(3):281–306
Ewell PT, Fano H, Raman KV, Alcazar J, Palacios M, Carhuamaca J (1990) Farmer management of potato insect pests in Peru. International Potato Center, Lima, 87pp
Fonseca C, Mujica N (1997) Impacto del control etologico sobre la mosca minadora (Liriomyza huidobrensis) en el valle de Cañete. Internal report, International Potato Center, Lima, Peru
Gurr G, Wratten S, Tylianakis J, Kean J, Keller M (2005) Providing plant foods for natural enemies in farming systems: balancing practicalities and theory. In: Wackers FL, van Rijn PCJ, Bruin J (eds) Plant provided food for carnivorous insects: a protective mutualism and its applications. Cambridge University Press, New York
Harris M, Begley J, Warkenstein D (1990) Liriomyza trifolii (Diptera: Agromyzidae) suppression with foliar applications of Steinernema carpocapsae (Rhabditida: Steinernema) and abemactin. J Econ Entomol 83:2380–2384
Hassan SA (1989) Vorstellungen der IOBC-Arbeitsgruppe “Pflanzenschutzmittel und Nutzorganismen” zur Erfassung der Nebenwirkungen von Pflanzenschutzmitteln auf Nutzlinge. Gesunde Pflanze 8:295–302
Horne PA, Page J (2008) IPM dealing with potato tuber moth (PTM) and all other pests in Australian potato crops. In: Kroschel J, Lacey L (eds) Integrated pest management for the potato tuber moth – a potato pest of global proportion. Tropical agriculture 20, Advances in crop research 10. Margraf Publishers, Weikersheim, Germany, pp 111–117
Horne PA, Rae JE, Henderson AP, Spooner-Hart R (1999) Awareness and adoption of IPM by Australian potato growers. Plant Protect Q 14:139–142
Jepson P, Thacker J (1990) Analysis of the spatial component of pesticide side-effects on non-target invertebrate populations and its relevance to hazard analysis. Funct Ecol 4:349–355
Kaya HK, Alcázar J, Parsa S, Kroschel J (2009) Microbial control of the Andean potato weevil complex. In: Tennant P, Benkeblia N (eds) Potato II. Fruit, vegetable and cereal science and biotechnology 3(Special issue 1):39–45
Keller S (2003) Integrated pest management of the potato tuber moth in cropping systems of different agroecological zones. In: Kroschel J (ed) Tropical Agriculture 11, Advances in Crop Research 1, Margraf Publisher, Weikersheim, Germany, 156pp
Keller M, Baker G (2002) Impact of conservation biological control practices on natural enemies: a case study of diamondback moth and its parasitoids. In: USDA Forest Service (ed) First international symposium on biological control of arthropods, USDA Forest Service, Washington, USA, pp 215–219
Kroschel J (1995) Integrated pest management in potato production in the Republic of Yemen with special reference to the integrated biological control of the potato tuber moth (Phthorimaea operculella Zeller). Tropical Agriculture 8, Margraf Verlag, Weikersheim, Germany, 227 pp
Kroschel J, Zegarra O (2010) Attract-and-kill: a new strategy for the management of the potato tuber moths Phthorimaea operculella (Zeller) and Symmetrischema tangolias (Gyen) in potato – laboratory experiments towards optimizing pheromone and insecticide concentration. Pest Manag Sci 66:490–496
Kroschel J, Cañedo V (2009) How do insecticides affect potato yield and ecosystem resilience to manage potato pests? An ecological assessment from the central highlands of Peru. In: 15th triennial symposium of the International Society for Tropical Root Crops, 2–6 Nov 2009, International Potato Center, Lima, Peru, pp 9–22
Kroschel J, Koch W (1994) Studies on the population dynamics of the potato tuber moth (Phthorimaea operculella Zeller) (Lepidoptera: Gelechiidae) in the Republic of Yemen. J App Ent 118:327–341
Kroschel J, Koch W (1996) Studies on the use of chemicals, botanicals and biological control in the management of the potato tuber moth in potato stores. Crop Prot 15(2):197–203
Kroschel J, Sporleder M (2006) Ecological approaches to integrated pest mangement of the potato tuber moth, Phthorimaea operculella (Zeller) (Lepidoptera: Gelechidae). In: Proceedings of the 45th annual Washington State potato conference, 7–9 Feb 2006, Moses Lake, Washington, pp 85–94
Kroschel J, Alcazar J, Pomar P (2009) Potential of plastic barriers to control Andean potato weevil Premnotrypes suturicallus Kuschel. Crop Prot 28:466–476
Kroschel J, Sporleder M, Tonnang HE, Juarez H, Carhuapoma P, Gonzales JC, Simon R (2011a) Predicting climate change caused changes in global temperature on potato tuber moth Phthorimaea operculella (Zeller) distribution and abundance using phenology modeling and GIS mapping. J Agric Forest Meteorology (Special Issue) (under revision)
Kroschel J, Cañedo V, Alcazar J, Miethbauer T (2011b) Manejo de plagas de la papa en la región andina del Perú – Guía de capacitación. International Potato Center, Lima, 85pp
Kühne M (2007) The Andean potato weevil Premnotrypes suturicallus ecology and interactions with the entomopathogenic fungus Beauveria bassiana. Ph.D. thesis, Georg-August-University, Göttingen, Germany, 177pp
Landis D, Wratten S, Gurr G (2000) Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu Rev Entomol 45:175–201
LeBeck L, Gaugler R, Kaya H, Hara A, Jhonson M (1993) Host stage suitability of the leafminer Liriomyza trifolii (Diptera: Agromyzidae) to the entomopathogenic nematode Steinernema carpocapsae (Rhabditida: Steinernematidae). J Invertebr Pathol 62:58–63
Llanderal-Cazares C, Lagunes-Tejada A, Carillo-Sanchez JL, Sosa-Moss C, Vera-Graziano J, Bravo-Mojica H (1996) Susceptibility of Phthorimaea operculella (Zeller) to insecticides. J Entomol Sci 31:420–426
Lösel P, Penners G, Potting R, Ebbinghaus D, Elbert A, Scherkenbeck J (2000) Laboratory and field experiments towards the development of an attract and kill strategy for the control of the codling moth, Cydia pomonella. Entomol Exp Appl 95:39–46
Loza AL, Bravo R (2001) Poblaciones de carabidos (Coleoptera) en agroecosistemas del altiplano peruano. Rev Per Ent 42:79–87
Mazlan N, Munford J (2005) Insecticide use in cabbage pest management in the Cameron Highlands, Malaysia. Crop Prot 24(1):31–39
Mujica N, Kroschel J (2011) Leafminer fly (Diptera: Agromyzidae) occurrence, distribution and parasitoid associations in field and vegetable crops along the Peruvian coast. Environ Entomol 40(2):217–230
Mujica N, Fonseca C, Suarez F, Fabian F, Marchena M, Cisneros F (2000) Reducción del uso de insecticidas en el control de la mosca minadora, Liriomyza huidobrensis Blanchard, por medio del uso de técnicas etológicas. In: Arning I, Lizárraga A (eds) Control Etológico. Uso de feromonas, trampas de colores y luz para el control de plagas en la agricultura sostenible. Red de Acción en Alternativas al Uso de Agroquímicos (RAAA), Lima, Peru
Mujica N, Pravatiner M, Cisneros F (2001) Effectiveness of abamectin and plant-oil mixtures on eggs and larvae of the leafminer fly, Liriomyza huidobrensis Blanchard. In: International Potato Center (ed) Scientist and farmer: partners in research for the 21st century. Program report 1999–2000. CIP, Lima, Peru
Murphy ST, La Salle J (1999) Balancing biological control strategies in IPM of New World invasive Liriomyza in field vegetable crops. Biocontrol News INF 20(3):91–104
Oerke EC, Dehne HW, Schönbeck F, Weber A (1994) Crop production and crop protection. Estimated losses in major food and cash crops. Elsevier Science, Amsterdam, p 808
Olthof T, Broadbent B (1991) Control of chrysanthemums leafminer, Liriomyza trifolii with the entomopathogenic nematode Heterorhabditis heliothidis. Nematologica 36:379
Orozco FA, Cole DC, Forbes G, Kroschel J, Wanigaratne S, Arica D (2009) Monitoring adherence to the International Code of Conduct – highly hazardous pesticides in central Andean agriculture and farmers’ rights to health. J Occup Env Health 15(3):255–268
Ortiz O, Alcazar J, Catalan W, Villano W, Cerna V, Fano H, Walker T (1996) Economic impact of IPM practices on the Andean potato weevil in Peru. In: Walker T, Crissman C (eds) Case studies of the economic impact of CIP-related technologies. International Potato Center, Lima, 157pp
Ortiz O, Kroschel J, Alcázar J, Orrego R, Pradel W (2009) Evaluating dissemination and impact of IPM: lessons from case studies of potato and sweetpotato IPM in Peru and other Latin American countries. In: Peshin R, Dhawan AK (eds) Integrated pest management: dissemination and impact. Springer, Dordecht, pp 419–434
Parsa S, Alcazar J, Salazar J, Kaya H (2006) An indigenous Peruvian entomopathogenic nematode for suppression of the Andean potato weevil. Biol Control 39:171–178
Poinar G (1990) Biology and taxonomy of Steinernematidae and Heterorhabditidae. In: Gaugler R, Kaya H (eds) Entomophathogenic nematodes in biological control. Boca Raton, Florida, pp 23–61
Raman KV, Alcazar J (1990) Peruvian virus knocks Potato tuber moth. In paper presented at the 74th annual meeting of Potato Association of America, Québec, Canada
Raman K, Radcliffe EB (1992) Pest aspects of potato production. Part 2. Insect pests. In: Harris PM (ed) The potato crop: the scientific basis for improvement, 2nd edn. Chapman and Hall, London, pp 476–506
Riechert SE, Lawrence K (1997) Test for predation effects of single versus multiple species of generalist predators: spiders and their insect prey. Entomol Exp Appl 84:147–155
Rios A, Kroschel J (2011) Evaluation and implications of Andean potato weevil infestation sources for its management in the Andean region. J Appl Entomol 135(10):738–748
Rondon SI, Vergara C (2003) Effects of insecticide applications in the population dynamic of Labiduria riparia Phallas (Dermaptera: Labiduridae) in four cultivars of sweet potato. Rev Per Entomol 43:121–127
Schmidt MH, Thies C, Tscharntke T (2004) The landscape context of arthropod biological control. In: Gurr GM, Wratten SD, Altieri MA (eds) Ecological engineering for pest management: advances in habitat manipulation for arthropods. CSIRO Press, Collingwood
Schuller S, Sanchez G (2003) Los artrópodos del suelo depredadores en agroecosistemas de maíz en el valle de Chancay, Lima, Perú. Rev Per Entomol 43:47–58
Sporleder M, Kroschel J, Gutierrez Quispe MR, Lagnaoui A (2004) A temperature-based simulation model for the potato tuberworm, Phthorimaea operculella Zeller (Lepidoptera; Gelechiidae). Environ Entomol 33:477–486
Sporleder M, Simon R, Juarez H, Kroschel J (2008) Regional and seasonal forecasting of the potato tuber moth using a temperature-driven phenology model linked with geographic information systems. In: Kroschel J, Lacey L (eds) Integrated pest management for the potato tuber moth – a potato pest of global proportion. Tropical Agriculture 20, Advances in Crop Research 10. Margraf Publishers, Weikersheim, Germany
Sporleder M, Simon R, Gonzales J, Chavez D, Juarez H, Mendiburu F, Kroschel J (2011) ILCYM: a software for developing insect pest phenology models based on temperature and for pest risk mapping (English version). Manual for ILCYM (Insect Life Cycle Modeling), version 2.0. International Potato Center (CIP), Lima, Peru, 78 pp
Trumble JT (1998) IPM: Overcoming conflicts in adoption. Integr Pest Manage Rev 3:195–207
Tscharntke T, Klein A, Kruess A, Steffan-Dewenter I, Thies C (2005) Landscape perspectives on agricultural intensification and biodiversity – ecosystem service management. Ecol Lett 8:857–874
Winters P, Fano H (1997) The economics of biological control in Peruvian potato production. Social Science Department, Working paper No. 1997-7. International Potato Center (CIP), Lima, Peru
Young JEB, Griffin MJ, Alford DV, Ogilvy SE (2001) Reducing agrochemical use on the arable farm: the TALISMAN and SCARAB projects. DEFRA, London, 416pp
Acknowledgements
The authors gratefully acknowledge the financial support for this research that was provided by the Regional Fund for Agricultural Technology (FONTAGRO), Washington, DC, for the project “Developing and use of ecological approaches in pest management for enhancing sustainable potato production of resource-poor farmers in Andean regions of Bolivia, Ecuador, and Peru”, and the Federal Ministry of Cooperation and Development (BMZ), Germany, for the project “Tackling Liriomyza leafmining flies: invasive pests of global proportions”. Finally, we are thankful to the valuable comments on our manuscript provided by Dr. Oscar Ortiz.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Kroschel, J., Mujica, N., Alcazar, J., Canedo, V., Zegarra, O. (2012). Developing Integrated Pest Management for Potato: Experiences and Lessons from Two Distinct Potato Production Systems of Peru. In: He, Z., Larkin, R., Honeycutt, W. (eds) Sustainable Potato Production: Global Case Studies. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-4104-1_25
Download citation
DOI: https://doi.org/10.1007/978-94-007-4104-1_25
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-4103-4
Online ISBN: 978-94-007-4104-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)