
Abstract
Various abiotic stress factors affect plants negatively, affecting their growth and development. These stress factors can be natural or anthropogenic and may have short-term or long-term effects on vegetation. Due to stress, many changes occur in plants, which may be reversible or irreversible and even cause acute damage. The abiotic stress factors are considered as one of the major factors for crop loss worldwide. Plants avoid such stress factors either by acclimatization or by avoidance and have with various defence mechanisms. In this chapter, authors have summarized the effects of temperature, drought, flood, light, and heavy metals as abiotic stress factors on plants along with the molecular mechanisms and signal transduction during such stresses.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1.1 Introduction
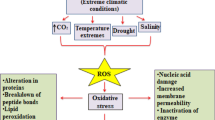
Plants are generally exposed to extremes of biotic and abiotic environmental factors, which have become the integral part of their life cycle too. For its optimum growth a plant requires carbon, energy, light, mineral nutrients, and water. Whenever any of these requirements are either in limited or in excessive amount, there is stress. The stress can be defined as any change in environmental condition having undesirable effects on normal growth and development of plants (Levitt 1980). Stress can also be defined as a non-ideal condition for survival. The first publication on “General Adaptation Syndrome” now known as “Biological Stress” was published in Nature in 1936 by Hans Selye (Lichtenthaler 1998). The concept of stress was originally developed by him and this term has also been used for unfavourable environmental constraints in plants. Plants may experience Eu-stress, which is a positive and growth promoting stress or they may experience De-stress which affects plants negatively (Lichtenthaler 1998). The biological species flourish best in optimum conditions and can tolerate the environmental factors as per Law of Limiting factors (Shelford 1952). Once these factors are in extremes they become a condition which promotes stress. The stress can be also categorized as natural or anthropogenic and can also be distinguished into biotic stress and abiotic stress based on which factor is causing stress (Table 1.1).
The biotic stress is caused by factors such as attacks by herbivores, predators, parasites, pathogens, etc., on the other hand, light, temperature, wind, water (flooding or drought), salinity, relative humidity, nutrition, etc., are responsible for abiotic stress. The effect each factor will have on plant depends on its quality and intensity. Although plants are adapted to changing environmental conditions, but as they cannot move or hide being sessile, they are vulnerable to stress. This co-occurrence of different stresses causes extensive loss to agricultural production; for example, if there is high temperature stress, then the probability of drought and high salinity is high in that area, due to availability of water at low levels. When such conditions prevail there is a cascade of effects at molecular, biochemical, and physiological levels (Mittler 2006).
Once a plant is in stress condition, it responds in different ways as depicted in Fig. 1.1, either it shows resistance and survives or it is susceptible to the stressful environment and dies and the last is avoidance, i.e. ephemeral plants complete their lifecycle before the inception of stress, thus avoids stress and survive (Hopkins and Huner 2009). All these responses of plants are due to complex serial changes at molecular, cellular, and physiological levels. Plants show such responses either to acclimatize with certain adaptations or withstand it. There are changes at biochemical, molecular, physiological, anatomical, morphological levels to survive in a stressful environment (Koyro et al. 2012). Plants have developed various perception and signalling pathways to survive such harsh environmental conditions. Various studies are on-going on Arabidopsis thaliana and Oryza sativa (Rice) as model organisms to understand the insights of plant stress biology.

Biotic and abiotic stresses affecting plant growth and development (Modified after Hopkins and Huner 2009)
Amongst various abiotic stresses, heat and drought affect the plant’s growth the most. Various studies have been carried out on these two factors. Popova et al. (2013) studied the RNA dependent DNA methylation pathway and concluded that this pathway is involved in heat stress resistance in Arabidopsis. Lee et al. (2013) studied the consequences of ABA production during drought conditions. The responses shown by plants towards stress are dynamic, complex and can be plastic (irreversible) or elastic (reversible) (Cramer 2010; Skirycz and Inze 2010; Cramer et al. 2011). Inhibition of protein synthesis, protein folding, processing are one of the first responses to abiotic stresses at molecular level. Plants also show some responses due to abiotic stress at hormone level and produce ABA or ethylene (Cramer et al. 2011).
The stress responses in plants are differentiated into four stages (Lichtenthaler 1998) as mentioned below (Fig. 1.2):
-
1.
Response—considered as commencement of stress and is also considered as Alarm phase.
-
2.
Restitution—stage of confrontation or resistance and in this stage stress continues.
-
3.
End—this is the stage of exhaustion, where stress intensity is too high and cell death may occur.
-
4.
Regeneration—In this phase if stress has not been so prolonged and extensive, regeneration may occur.

Different response phases shown by plants during stress conditions (Modified after Lichtenthaler 1998)
1.2 Types of Abiotic Stress
The abiotic stress includes inanimate environmental components as mentioned above. These abiotic stress factors can occur naturally or may be caused by anthropogenic activities. As plants are sessile and their surrounding environment is ever-changing the occurrence of physiological plasticity is essential for their survival (Gaspar et al. 2002). Plants respond to different types of stresses in different ways as described in different sections:
1.2.1 Temperature as Stress Factor
Plants respond differently to changes in temperature as for every plant species there is an optimum range in which it survives best. In a same temperature range one species may survive and other species may be sensitive as their thermo-sensitive metabolic processes may get hampered due to changes in temperature leading to change in cell processes. Plant species which are growing in hot and dry deserts or high altitudes experience very high temperature during day and low during night. Plants growing in tropical forests or northern forests grow in altogether different temperature ranges so when such plants grow in different climatic conditions they get stressed and are prone to injuries.
1.2.1.1 Low Temperature Stress
Low temperature is considered as one of the major abiotic stress limiting the growth and development. Plants growing in temperate regions are adapted for low temperature conditions, and based on temperature range plants can survive, i.e., cold tolerant or get injury and are classified as chilling-sensitive (10–15 °C); freezing sensitive (0 °C); and freezing resistant plants (able to survive sub-zero temperature) (Pollock and Eagles 1988; Hughes and Dunn 1990). When exposed to low temperature, plants exhibit three types of injuries: desiccation, chilling injury, and freezing injury. Freezing in plants can be induced either from freeing in soil water or freezing of water inside the cells or at intracellular level. Some plants develop freezing tolerance due to formation of antifreeze proteins, which along with some sugars (sucrose) inhibits crystallization at cellular level. Woody or hardy plants are reported to be highly resistant to chilling injuries. The common symptoms of chilling injury are changes in membrane structure, composition, protoplast streaming reduced, increased/decreased respiration, reduced photosynthesis, production of abnormal metabolites, reduced plants growth, curling of leaves, necrosis, die back of stems, abnormal ripening of fruit, decreased N partitioning in young shoots, altered reproductive behaviour (Hashimoto and Komatsu 2007). Generally tropical plants are more susceptible to chilling than the temperate plants; for example, Zea mays, Lycopersicum esculentum, Glycine max show chilling injuries at 10–15 °C, there are some temperate plants which experience chilling injury at 0 °C–5 °C; for example, apple (Malus sp.) and Asparagus sp. (Hopkins and Huner 2009).
Plants also acclimatize themselves for chilling injuries. The acclimatization in plants is associated with changes in several biochemical and physiological processes such as altered gene expressions dehydrins (LEA, late embryogenesis abundant) are induced in various plants, changes in hormone levels (increased ABA), increase in soluble sugars, amino acids, organic acids, increased levels of osmoprotectants, etc. (Palva et al. 2002).
1.2.1.2 High Temperature Stress
Change in global mean temperature and light conditions have significant impact on distribution, abundance, phenology, and physiology of various crop species (Djanaguiraman and Prasad 2014). It has been estimated that climate change decreases the average suitable cultivable area for many plant species such as Arachis, Solanum, and Vigna by 63–100%. It is also predicted that some species might extinct in near future due to reduction in cultivable area (Jarvis et al. 2008). Soil temperature ranges between 45 and 80 °C, where forest soil temperature remains between 40 and 50 °C and in deserts it may reach up to 70–80 °C. Depending on the temperature range, responses exhibited by plants can be categorized as over temperature, intermediate and under temperature responses. The high temperature or heat stress damages the plants and causes the altered phenology, reduced growth and development, scorching of leaves and fruits, sun scabs, abrasions, etc. (Nahar et al. 2015). At cellular and molecular level, high temperature affects stability of various proteins, thus prohibiting or altering many enzymatic activities leading to metabolic disorders, change in membrane and cytoskeleton structure, altered RNA species. Effect of high temperature has been observed in many crop species such as Oryza sativa (rice), Capsicum annuum (capsicum), Triticum aestivum (wheat), Hordeum vulgare (barley), Zea mays (maize), Glycine max (soybean), Abelmoschus esculentus (okra), etc. (Hasanuzzaman et al. 2013). Also, there is formation of Heat Shock Proteins (HSPs), whenever temperature is elevated beyond the threshold limits, which varies from species to species.
1.2.2 Light Stress
Light is an important environmental factor which is essential for CO2 assimilation through photosynthesis, but as it crosses a plants tolerance level it becomes a stress factor and photo inhibition occurs in plants. Plants are exposed to two types of radiations: UV-A (315–400 nm) and UV-B (280–315 nm). UV-A is photo-oxidative and UV-B is photo-oxidative as well as causes photo lesions in bio-membranes. UV damages disulphide bridges in the proteins, dimerization of thymine groups in DNA, and disrupts xanthophyll cycle. When exposed to excessive light stress, plants may sense high photon flux. Shade loving plants get damaged even by brief exposure of strong light. In plants the excessive light damage photosystem II by destroying photosensitive pigments and thylakoid structures, thus inhibiting photosynthesis. There is disruption in electron transport chain and breaking of protein sub-units (Hopkins and Huner 2009). To avoid damage by strong light, plants show some adaptive features such as orienting leaves at an angle to receive less radiation, rolling up of shoots, dense covering of trichomes on upper surface of leaves, thickened cell walls, and production of anthocyanin pigment to shield mesophyll (Balfagón et al. 2019).
In tomato (Solanum lycopersicum) lipidomics analysis identified lipophilic antioxidant molecules, which protects PS II from photodamage (Spicher et al. 2017). The study conducted by Balfagón et al. (2019) showed that in Arabidopsis, in high light intensity combined with heat stress, the stomata opens to increase transpiration and cool the leaf instead of reducing stomatal aperture (Devireddy et al. 2018).
1.2.3 Water Stress
Water which is present in 80–90% in non-woody plants is key molecules for all metabolic processes. It is essential and when it becomes limiting and is not available to roots or when transpiration rate is very high it becomes a stress factor. Water stress may arise when there is water deficit or excess of water (flooding). During flooding oxygen becomes limiting and is not supplied to roots as per requirement. Due to water deficit stress there is desiccation; protoplasm dries up, solute concentration increases, leading to loss in integrity of membranes. Selectivity of membranes is lost. Proteins get denatured and displaced, and finally leading to loss of cellular compartmentalization. Due to water deficit the water potential inside the cells decreases leading to stomatal closure. It is also known that hydropassive stomatal closure occurs in plants to avoid transpirational loss and is evident in all tropical, temperate, or dessert plants (Hopkins and Huner 2009). Stomatal closure limits gaseous exchange, reduced transpiration and photosynthesis, uptake of mineral nutrients and disruption of homeostasis and ions. Drought stress also causes various other adverse effects such as decreased water potential leading to hydro active closure of stomata, lesser number of stomata, thickening of leaf cell wall, decreased cell enlargement, slow or inhibited growth and reproduction, cutinization of leaf surface, poorly developed conducting system, increase in root–shoot ratio, production of osmolytes such as proline, formation of reactive oxygen species (ROS), ascorbate, and glutathione accumulates in cell and further aggravates the condition (Lisar et al. 2012). Water deficits also alter the cell wall confirmations, non-enzymatically through the interaction of pectate and calcium (Boyer 2009; Cramer et al. 2011). Although photosynthesis gets disrupted in both C3 and C4 plants but studies show that C4 plants are more sensitive to drought. Water stress also alters the mRNA expression and thus new proteins are synthesized. Different proteins which are synthesized under stress are late embryogenesis abundant (LEA), dehydrins, desiccation stress proteins, protease, etc. (Lisar et al. 2012).
When there is flooding plants undergo hypoxia, a stage when oxygen concentration is less. During flooding or excessive irrigation water logging occurs in soil, thus limiting the oxygen availability to roots. When water logging is for a prolonged period damage occurs depending on the extent of adaptability and soil conditions. The damage also depends on the type, developmental stage, genotype of plants, extent of severity, and duration of water stress (Fukao and Bailey-Serres 2004; Mariani and Ferrante 2017).
1.2.4 Salinity Stress
Salinity stress is another major problem of world, which decreases crop productivity. High concentration of salt accumulates in the soil either due to improper irrigation or poor drainage. Soil salinity is measured by accumulation of soluble salts such as Cl−, SO42−, Na+, Ca2+, etc., and it harms the plant growth and development through water stress and cytotoxicity (Isayenkov and Maathuis 2019). Soil salinity may be categorized into primary (natural salinity; for example, marshes, salt lake, tidal swamps, etc.) or secondary (due to anthropogenic activities). High saline condition decreases the water potential and causes osmotic stress. Osmotically active compounds or osmolytes (Glycine-Betaine) also known as compatible metabolites accumulate in cells to lower the osmotic potential (Rasool et al. 2013). Salinity is also many times accompanied by the formation of ROS, polyamines (putrescene), and proline. Saline soils also limit the uptake of nutrients by root from soil (Sharma et al. 2016). During high salinity stress, the photosynthetic activities gets affected as thylakoid membranes of chloroplast get damaged, reduction of photosynthetic pigments takes place, further inactivation of electron transport chain, photophosphorylation occurs (Srivastava et al. 2019). Also, the turgor pressure of leaves changes reducing the stomatal conductance and rate of transpiration (Srivastava et al. 2019). ABA accumulates in guard cells, excessive production of hydrogen peroxide occurs which leads to triggering of oxidative stress by influencing the production of reactive oxygen species (Foyer and Noctor 2005). It has been reported that legumes and cereal crops are very sensitive to salinity. Salinity affects nitrogen fixation as it suppresses the formation of root nodules (Ramana et al. 2012). Nutritional imbalance enhances vegetative growth over reproductive growth, thus decreasing crop yield (Munns and Tester 2008). It also causes chlorosis and senescence, DNA, RNA content decreases, mitotic and enzymatic activity slows down (Ali et al. 2004).
The physiological drought condition in plants can be overcome by the application of certain soil microorganisms such as arbuscular mycorrhizal fungi (AMF). Symbiotic association established between AMF and plant roots, mainly in saline environments, could withstand plants under excessive saline conditions as the AMF hyphae run several metres away from the stressed area or from the zone of nutrient depletion, thereby increasing root surface area and facilitating nutrient uptake by the plant. Due to this, physiological processes and various metabolic activities get improved and adverse effects of physiological drought could minimize in plants (Saxena et al. 2017).
1.2.5 Heavy Metal Stress
Extensive activities related to agriculture, industrialization, and urbanization are adding enormous amount of heavy metals; for example, Arsenic (As), Cadmium (Cd), Chromium, Lead (Pb), Mercury (Hg) in environment. Addition of excessive amount of heavy metals damage and alter the soil texture, pH, thus affecting plant growth and development. These toxic elements promote morphological, metabolic abnormalities and thus disrupt cell homeostasis (Amari et al. 2017; Tiwari and Lata 2018).
Some of these metals are vital micronutrients, and are accountable for numerous metabolic processes in a plant, but, if they exceed the threshold limit, these metals can have detrimental effects on metabolic pathways, physiological processes, plant growth and development, and finally senescence (Ghori et al. 2019). As a first line of resistance exhibited, plants reduce the heavy metal uptake, through cellular and root exudates that restrict entry of metals inside the cells (Shahid et al. 2015; Ghori et al. 2019). The second line of resistance involves adopting other mechanisms for detoxification by transports, sequestration of these heavy metal ions, and chelates in the plant’s vacuoles (Ghori et al. 2019). One of the most destructive effects caused by heavy metals in plants is disruption of bio-membranes by lipid per-oxidation (Demiral and Türkan 2005; Yadav 2010). Some of the harmful effects induced by heavy metals are described below:
High concentration of Zinc results in stunted growth, chlorosis, senescence, and changed root and shoot ratio (Fontes and Cox 1998; Yadav 2010). Another heavy metal chromium is released from tanning industry. The excess availability of chromium can bring chlorosis in young leaves, inhibit plant growth, nutrient imbalance, root injury, and wilting of tips (Scoccianti et al. 2006; Yadav 2010). Lead in soil resulting from various sources like municipal sewage sludge, mining and smelting activities, paints, gasoline and explosives, etc. It deteriorates plant growth, morphology, and photosynthesis. Many enzymatic activities are inhibited when lead concentration is very high, membrane permeability is lost, disturbs uptake of mineral, and causes water imbalance (Sharma and Dubey 2005; Yadav 2010). Recently, several studies revealed the physiological and molecular mechanisms of arsenic toxicity, its accumulation, detoxification, and tolerance in various concentrations in plants such as carrot, lettuce, rice, and spinach can cause several physiological disorders (Kumar et al. 2015; Tiwari and Lata 2018).
1.3 Molecular Mechanisms and Signal Transduction in Stress
The three systematic approaches or three “OMICS” (such as Metabolomics, Transcriptomics, Proteomics) studies have enhanced the knowledge on how a plant response at molecular level and what are the complex regulatory mechanisms. The transcriptomics deals with the RNAs and their expressions. Proteomics studies about how proteins modify and metabolomics is a tool to study about the metabolites. The integration of all these three omics has shown various clear pathways related to concepts of plant responses towards stress (Cramer et al. 2011). Signal perception, transduction, and finally expression of stress response genes are the three steps at molecular level once the stress is present in environment (Fig. 1.3). The cascades are associated with reactive oxygen species, Ca2+, Abscisic acid (ABA), Jasmonates (JA), Salicylic acid (SA), Ethylene (ET) which coordinate various signal pathways. Once the signal is perceived, transduction pathways such as calcium dependent protein kinase (CDPK), mitogen activated protein kinase (MAPK) get activated, further down regulating various transcription factors (Baillo et al. 2019). One example of Ca2+ signal pathway can be understood in mechanism of closing of guard cells in drought stress. During drought stress, changes occur in cytosolic concentration of Ca2+ which initiates CPK activity resulting into the release of ABA, which further activates the calcium chelator (BAPTA). When guard cells receive BAPTA, signal comes to control the transpiration and guard cell closes (Zou et al. 2015; Atif et al. 2019). Experimental studies on wheat have shown that CPKs show functional diversity and complexity. The isoforms of closely related CPK genes such as TaCPK 7 and TaCPK 12 in wheat show diversity wherein, the former responds to various stresses such as temperature (low), salinity, drought, H2O2, whereas the latter responds only through ABA signalling (Geng et al. 2011; Atif et al. 2019).

Signalling pathway in response to abiotic stress (Modified after Baillo et al. 2019)
As transcription factors act as key stress tolerance mediators, they can be modified to increase the stress tolerance in various crops. Many studies have been conducted on five major cereal crops such as barley, maize, sorghum, rice, and wheat. Gene modification through these transcription factors (TFs) can improve stress tolerance in transgenic plants. Although boosting tolerance level through TFs is not so easy and is a complicated procedure, as one TF gene during down-regulation may promote or suppress other genes. Various TFs families may be involved in one or multiple stress responses, thus making TF responses very complex and complicated and might have cross-talk between different signal pathways. There are millions of TF at molecular level, and identifying, modifying each TF is a bit challenging task. Application of CRISPR/Cas 9, a gene editing tool is being used to improve the stress tolerance in plants (Baillo et al. 2019).
Abiotic stress also enhances the production, synthesis, and transcription of heat shock proteins (HSPs) in comparison to other normal proteins. Post transcriptional modifications of proteins such as microRNA and alternating splicing also help to cope with abiotic stress. The HSPs have been categorized based on molecular weight into Large (68,000–104,000 Da), Intermediate (20,000–23,000 Da), and Small (15,000–18,000 DA) (Hughes and Dunn 1990). It has now been established that HSPs help newly synthesized protein’s folding and protect them during stress. Therefore HSPs are also known as molecular chaperones (Ul Haq et al. 2019). HSPs are also present in cell and cellular compartments during standard environmental conditions and studies have confirmed their role in normal growth and development apart from being stress responsive (Eck et al. 2007). HSPs not only develop during heat stress but are also found to be present in other abiotic stresses.
HSPs express differentially in different species. Genes encoding HSPs, are present in different cell compartments and thus they are expressed differently and may also be specific depending on the stress intensity (Liu et al. 2006; Ul Haq et al. 2019). Under temperature stress high molecular weight HSPs (HSP 118, −114, −110, −108, −104, −103, −101, −100, and −97) are formed. In Arabidopsis and maize HSPs −100 and −101 are expressed and responsive against the high temperature stress and thermo-tolerance (Queitsch et al. 2000; Nieto-Sotelo et al. 2002). In Pea low molecular weight HSPs 18.1 and −17.9 accumulates after 4 h treatment (Dupuis and Dumas 1990). Low temperature also induces the expression of HSPs in rice, maize, and Arabidopsis to protect them against cold stress (Bae et al. 2003; Kosova et al. 2011; Hlavackova et al. 2013). HSPs have also found to be present during drought stress. Mostly HSPs 70 are upregulated as evident from studies on rice, Arabidopsis, sugar cane, cotton, maize, Cicer sp. (Subba et al. 2013; Reddy et al. 2014; Yer et al. 2018). In Chenopodium rubrum under high light stress HSP 23 gets upregulated (Korotaeva et al. 2001). In marine ecosystems low light stress induced accumulation of HSP 70, ClpBi, and HSP 60 (Kumar et al. 2017). Similarly HSPs have also been found to be upregulated by various other abiotic stresses such as heavy metals, flooding, oxidative stress, etc.
1.4 Conclusions
Various abiotic stresses affect plant’s phenology, disrupt metabolic activities at physiological, cellular, and molecular levels. Such changes unfavourably affect the growth and development of plants and in field crop species even significantly reduce the crop yield. The various stress factors are transduced through signal transduction pathways. The components involved in such pathways are calcium, ROS, protein kinase, etc. The stress signals further regulate transcription factors and thereby controlling gene regulation. Future studies are required focussing on different crop plants, to explore regulatory mechanisms; for example, MiRNA, alternative splicing, and cross-talk between different pathways. Such biotechnological studies using molecular approach would help to develop stress resistant and tolerant crops.
References
Ali Y, Aslam Z, Ashraf MY, Tahir GR (2004) Effect of salinity on chlorophyll concentration, leaf area, yield and yield components of rice genotypes grown under saline environment. Int J Environ Sci Technol 1(3):221–225
Amari T, Ghnaya T, Abdelly C (2017) Nickel, cadmium and lead phytotoxicity and potential of halophytic plants in heavy metal extraction. S Afr J Bot 111:99–110
Atif RM, Shahid L, Waqas M, Ali B, Rashid MAR, Azeem F, Nawaz MA, Wani SH, Chung G (2019) Insights on calcium-dependent protein kinases (CPKs) signaling for abiotic stress tolerance in plants. Int J Mol Sci 20(21):5298. https://doi.org/10.3390/ijms20215298
Bae MS, Cho EJ, Choi E, Park OK (2003) Analysis of the Arabidopsis nuclear proteome and its response to cold stress. Plant J 36:652–663
Baillo EH, Kimotho RN, Zhang Z, Xu P (2019) Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes (Basel) 10(10):771. https://doi.org/10.3390/genes10100771
Balfagón D, Sengupta S, Gómez-Cadenas A, Fritschi FB, Azad RK, Mittler R, Zandalinas SI (2019) Jasmonic acid is required for plant acclimation to a combination of high light and heat stress. Plant Physiol 181(4):1668–1682
Boyer JS (2009) Evans review: cell wall biosynthesis and the molecular mechanism of plant enlargement. Funct Plant Biol 36(5):383–394
Cramer GR (2010) Abiotic stress & plant responses from the whole vine to the genes. Aust J Grape Wine Res 16:86–93
Cramer GR, Urano K, Delrot S, Pezzotti M, Shinozaki K (2011) Effects of abiotic stress on plants: a systems biology perspective. BMC Plant Biol 11:163
Demiral T, Türkan I (2005) Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environ Exp Bot 53:247–257
Devireddy AR, Zandalinas SI, Gómez-Cadenas A, Blumwald E, Mittler R (2018) Coordinating the overall stomatal response of plants: rapid leaf-to-leaf communication during light stress. Sci Signal 11(518):1–9. https://doi.org/10.1126/scisignal.aam9514
Djanaguiraman M, Prasad PVV (2014) High temperature stress. In: Jackson M, Ford-Lloyd B, Parry M (eds) Plant genetic resources and climate change. CABI, Wallingford, pp 201–220
Dupuis I, Dumas C (1990) Influence of temperature stress on in vitro fertilization and heat shock protein synthesis in maize (Zea mays L.) reproductive tissues. Plant Physiol 94:665–670. https://doi.org/10.1104/pp.94.2.665
Eck ERHB, Ettig SEF, Nake CLK, Artig KAH, Hattarai TRB (2007) Specific and unspecific responses of plants to cold and drought stress. J Biosci 32:501–510
Fontes RLS, Cox FR (1998) Zinc toxicity in soybean grown at high iron concentration in nutrient solution. J Plant Nutr 21:1723–1730
Foyer CH, Noctor G (2005) Oxidant and antioxidant signalling in plants: a re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ 28:1056–1071
Fukao T, Bailey-Serres J (2004) Plant responses to hypoxia-is survival a balancing act? Trends Plant Sci 9(9):459–456
Gaspar T, Franck T, Bisbis B, Kevers C, Jouve L, Hausman JF, Dommes J (2002) Concepts in plant stress physiology. Application to plant tissue cultures. Plant Growth Regul 37:263–285
Geng S, Zhao Y, Tang L, Zhang R, Sun M, Guo H, Kong X, Li A, Mao L (2011) Molecular evolution of two duplicated CDPK genes CPK7 and CPK12 in grass species: a case study in wheat (Triticum aestivum L.). Gene 475:94–103
Ghori NH, Ghori T, Hayat MQ, Imadi SR, Gul A, Altay V, Ozturk M (2019) Heavy metal stress and responses in plants. Int J Environ Sci Technol 1:1–22
Hasanuzzaman M, Nahar K, Alam MM, Roychowdhury R, Fujita M (2013) Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int J Mol Sci 14:9643–9684
Hashimoto M, Komatsu S (2007) Proteomic analysis of rice seedlings during cold stress. Proteomics 7:1293–1302
Hlavackova I, Vitamvas P, Santrueek J, Kosova K, Zelenkova S, Prasil IT, Ovesna J, Hynek R, Kodicek M (2013) Proteins involved in distinct phases of cold hardening process in frost resistant winter barley (Hordeum vulgare L.) Cv Luxor. Int J Mol Sci 14:8000–8024
Hopkins WG, Huner NP (2009) Introduction to plant physiology, 4th edn. Wiley, Hoboken, NJ, p 503
Hughes MA, Dunn MA (1990) The effect of temperature on plant growth and development. Biotechnol Genet Eng Rev 8(1):161–188. https://doi.org/10.1080/02648725.1990.10647868
Isayenkov SV, Maathuis FJM (2019) Plant salinity stress: many unanswered questions remain. Front Plant Sci 10:80. https://doi.org/10.3389/fpls.2019.00080
Jarvis A, Lane A, Hijmans RJ (2008) The effect of climate change on crop wild relatives. Agric Ecosyst Environ 126:13–23
Korotaeva NE, Antipina AI, Grabelnykh OI, Varakina NN, Borovskii GB, Voinikov VK (2001) Mitochondrial low-molecular-weight heat-shock proteins and the tolerance of cereal mitochondria to hyperthermia. Russ J Plant Physiol 48:798–803
Kosova K, Vitamvas P, Prasil IT, Renaut J (2011) Plant proteome changes under abiotic stress-contribution of proteomics studies to understanding plant stress response. J Proteome 74:1301–1322
Koyro HW, Ahmad P, Geissler N (2012) Abiotic stress responses in plants: an overview. In: Ahmad P, Prasad MNV (eds) Environmental adaptations and stress tolerance 1 of plants in the era of climate change. Springer, New York. https://doi.org/10.1007/978-1-4614-0815-4_1
Kumar S, Dubey RS, Tripathi RD, Chakrabarty D, Trivedi PK (2015) Omics and biotechnology of arsenic stress and detoxification in plants: current updates and prospective. Environ Int 74:221–230
Kumar M, Padula MP, Davey P, Pernice M, Jiang Z, Sablok G, Contreras-Porcia L, Ralph PJ (2017) Proteome analysis reveals extensive light stress-response reprogramming in the seagrass Zostera muelleri (Alismatales, Zosteraceae) metabolism. Front Plant Sci 7:2023
Lee SC, Lim CW, Lan WZ, He K, Luan S (2013) ABA signaling in guard cells entails a dynamic protein–protein interaction relay from the PYL–RCAR family receptors to ion channels. Mol Plant 6:528–538
Levitt J (1980) Responses of plants to environmental stresses. In: Chilling, freezing, and high temperature stresses, vol 1, 2nd edn. Academic Press, New York, p 497
Lichtenthaler HK (1998) The stress concept in plants: an introduction. Ann N Y Acad Sci 851:187–198
Lisar SYS, Motafakkerazad R, Hossain MM, Rahman IMM (2012) Water stress in plants: causes, effects and responses. Intech, Rijeka. https://doi.org/10.5772/39363
Liu D, Zhang X, Cheng Y, Takano T, Liu S (2006) RHSP90 gene expression in response to several environmental stresses in rice (Oryza sativa L.). Plant Physiol Biochem 44:380–386
Mariani L, Ferrante A (2017) Agronomic management for enhancing plant tolerance to abiotic stresses-drought, salinity, hypoxia, and lodging. Horticulturae 3:52
Mittler R (2006) Abiotic stress, the field environment and stress combination. Trends Plant Sci 11:15–19
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59:651–681
Nahar K, Hasanuzzaman M, Ahamed KA, Hakeem KR, Ozturk M, Fujita M (2015) Plant responses and tolerance to high temperature stress: role of exogenous phytoprotectants. In: Hakeem KR (ed) Crop production and global environmental issues. Springer, Cham. https://doi.org/10.1007/978-3-319-23162-4_17
Nieto-Sotelo J, Martinez LM, Ponce G, Cassab GI, Alagon A, Meeley RB, Ribaut JM, Yang R (2002) Maize HSP101 plays important roles in both induced and basal Thermotolerance and primary root growth. Plant Cell 14:1621–1633
Palva ET, Htiharju ST, Tamminen I, Puhakainen T, Laitinen R, Svensson J, Helenius E, Heino P (2002) Biological mechanisms of low temperature stress response: cold acclimation and development of freezing tolerance in plants. JIRCAS Work Ret 23:9–15
Pollock CJ, Eagles CF (1988) Low temperature and the growth of plants. In: Long SP, Woodward FI (eds) Plants and temperature, vol 42. Society for Experimental Biology Symposium, Company of biologists, Cambridge, pp 157–180
Popova OV, Dinh HQ, Aufsatz W, Jonak C (2013) The RdDM pathway is required for basal heat tolerance in Arabidopsis. Mol Plant 6:396–410. https://doi.org/10.1093/mp/sst023
Queitsch C, Hong SW, Vierling E, Lindquist S (2000) Heat shock protein 101 plays a crucial role in Thermotolerance in Arabidopsis. Plant Cell 12:479–492. https://doi.org/10.1105/tpc.12.4.479
Ramana GV, Padhy SP, Chaitanya KV (2012) Differential responses of our soybean (Glycine max L.) cultivars to salinity stress. Legume Res 35:185–193
Rasool S, Hameed S, Azooz MM, Rehman MU, Siddiqi TO, Ahmed P (2013) Salt stress: causes, types and responses of plants. In: Ahmad P, Azooz MM, Prasad MNV (eds) Ecophysiology and responses of plants under salt stress. Springer, New York, pp 1–24. https://doi.org/10.1007/978-1-4614-4747-4_1
Reddy PS, Kishor PBK, Seiler C, Kuhlmann M, Eschen-Lippold L, Lee J, Reddy MK, Sreenivasulu N (2014) Unraveling regulation of the small heat shock proteins by the heat shock factor HvHsfB2c in barley: its implications in drought stress response and seed development. PLoS One 9(3):e89125. https://doi.org/10.1371/journal.pone.0089125
Saxena B, Shukla K, Giri B (2017) Arbuscular mycorrhizal fungi and tolerance of salt stress in plants. In: Arbuscular mycorrhizas and stress tolerance of plants. Springer, Singapore, pp 67–97. https://doi.org/10.1007/978-981-10-4115-0_4
Scoccianti V, Crinelli R, Tirillini B, Mancinelli V, Speranza A (2006) Uptake and toxicity of Cr (Cr3+) in celery seedlings. Chemosphere 64:1695–1703
Shahid M, Khalid S, Abbas G, Shahid N, Nadeem M, Sabir M, Aslam M, Dumat C (2015) Heavy metal stress and crop productivity. In: Hakeem K et al (eds) Crop production and global environmental issues. Springer, Cham
Sharma P, Dubey RS (2005) Lead toxicity in plants. Braz J Plant Physiol 17:35–52
Sharma V, Rena V, Kumar D, Pandey RN, Singh B (2016) Sulfur regulates iron uptake and iron use efficiency in bread and durum wheat. Ind J Plant Physiol 21:189–196
Shelford VE (1952) Paired factors and master factors in environmental relations. Ill Acad Sci Trans 45:155–160
Skirycz A, Inze D (2010) More from less: plant growth under limited water. Curr Opin Biotechnol 21(2):197–203
Spicher L, Almeida J, Gutbrod K, Pipitone R, Dörmann P, Glauser G, Rossi M, Kessler F (2017) Essential role for phytol kinase and tocopherol in tolerance to combined light and temperature stress in tomato. J Exp Bot 68:5845–5856
Srivastava P, Wu Q-S, Giri B (2019) Salinity: an overview. In: Giri B, Varma A (eds) Microorganisms in saline environments: strategies and functions. Soil biology, vol 56, pp 3–18. https://doi.org/10.1007/978-3-030-18975-4_1
Subba P, Barua P, Kumar R, Datta A, Soni KK, Chakraborty S, Chakraborty N (2013) Phosphoproteomic dynamics of chickpea (Cicer arietinum L.) reveals shared and distinct components of dehydration response. J Proteome Res 12:5025–5047
Tiwari S, Lata C (2018) Heavy metal stress, signalling, and tolerance due to plant associated microbes: an overview. Front Plant Sci 9:452
Ul Haq S, Khan A, Ali M, Khattak AM, Gai WX, Zhang HX, Wei AM, Gong ZH (2019) Heat shock proteins: dynamic biomolecules to counter plant biotic and abiotic stresses. Int J Mol Sci 20(21):5321. https://doi.org/10.3390/ijms20215321
Yadav SK (2010) Heavy metals toxicity in plants: an overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S Afr J Bot 76(2):167–179
Yer EN, Baloglu MC, Ayan S (2018) Identification and expression profiling of all Hsp family member genes under salinity stress in different poplar clones. Gene 678:324–336
Zou JJ, Li XD, Ratnasekera D, Wang C, Liu WX, Song LF, Zhang WZ, Wu WH (2015) Arabidopsis calcium-dependent protein Kinase8 and CATALASE3 function in abscisic acid-mediated signaling and H2O2 homeostasis in stomatal guard cells under drought stress. Plant Cell 27:1445–1460
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Baweja, P., Kumar, G. (2020). Abiotic Stress in Plants: An Overview. In: Giri, B., Sharma, M.P. (eds) Plant Stress Biology. Springer, Singapore. https://doi.org/10.1007/978-981-15-9380-2_1
Download citation
DOI: https://doi.org/10.1007/978-981-15-9380-2_1
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-15-9379-6
Online ISBN: 978-981-15-9380-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)