
Abstract
The emanations upon combustion of petroleum fuels cause grave pessimistic impact on surrounding ambiance and global climate change. Therefore, the contemporary stance of scientific fraternity is to generate energy and mercantile products through biological methods with waste as a resource. Biohydrogen production from wastewater seems to be a promising green option for sustainable renewable energy. The process is feasible from practical point of view and can be operated under ambient conditions. It has been attracting attention due to its applicability to different types of wastewaters, and the production costs of biohydrogen can compete economically with other traditional methods. However, the crucial challenges like enhancing rate and yield for sustainable biohydrogen production still persist. During fermentation process, the undissociated volatile fatty acids (VFAs) and alcohols accumulate in the system leading to inhibition and redundancy in substrate degradation. Employing integration strategies with other bioprocesses like photo-fermentation or bio-electrochemical systems is the sanguine option to make the process frugally possible.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
12.1 Present Scenario
Paucity of worldwide petroleum reserves and apprehensions concerning environmental pollution prompt for the advancement of green energy substitutes (Arimi et al. 2015). Bioenergy has the sustainable potential that is appealing great curiosity amongst researchers across the world (Rama Mohan 2015). Recently, biohydrogen is cited to be one of the prospective areas of interest (Venkata Mohan 2009). In 2010, the market demand of international hydrogen production was 53 million metric tons and is anticipated to rise with a growth rate of 5.6% per annum. Worldwide commercial H2 production presently is produced majorly from natural gas, oil and coal and to certain extent by water electrolysis (Markets and Markets 2011; Bhaskar et al. 2013; Rama Mohan 2015). Alternatively, H2 from waste biomass/wastewater using biological methods as renewable resource is gaining much interest. By this way negative-valued organic waste is transformed to green energy abating the pollution (Ghimire et al. 2015). The governing prerequisite for wastewater treatment is an ideal prospect to produce biohydrogen. By this way, it diminishes the overall expenditure towards effluent treatment, and additional revenue could be incurred from biohydrogen as a fuel (Nissilä et al. 2014; Thi et al. 2016).
12.2 Biohydrogen Production
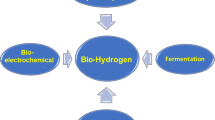
There are several routes for H2 production, viz. physical, chemical, biological and thermochemical (Fig. 12.1). BioH2 production is majorly contributed by anaerobic fermentation which is categorised as light-independent and light-dependent (Venkata Mohan et al. 2007a; Hallenbeck 2013). Light-independent fermentation process is usually referred to as dark fermentation that hires both strict anaerobes and facultative bacteria. This process results a higher rate of H2 and formation of soluble metabolic products (SMPs), viz. volatile fatty acids, solvents, etc. Acidogenic anaerobes are incompetent of consuming these undissociated organic acids resulting in the accumulation of these acids leading to a drop in pH value. Subsequently, H2 production is inhibited and, finally, results in low H2 yields (Guo et al. 2010; Venkata Mohan et al. 2017). Light-dependent processes used by cyanobacteria and green algae are direct and indirect biophotolysis or via photo-fermentation mediated by photosynthetic bacteria. H2 production by photosynthetic bacteria occurs by photo-fermentation of inorganic/organic acids (Lam and Lee 2013). The biochemistry varies considerably based on the biocatalyst, operational conditions, microenvironment and substrate (Lin et al. 2012).

Flowchart representing various routes for biohydrogen production
12.2.1 Photo-Fermentation
Photosynthetic microorganisms, viz. cyanobacteria, photosynthetic bacteria and algae, have varied photosynthetic machinery (anoxygenic or oxygenic) that functions for H2 production (Allakhverdiev et al. 2010). The process involves generation of a proton gradient by energy from sunlight and electron generation either by direct photolysis of water (Fig. 12.2a) or by indirect photolysis involving a parallel photosystem II (PSII)-independent process (Fig. 12.2b). In direct photolysis, the sunlight functions as a powerhouse for PSII ensuing production of oxidising equivalents that are used for the water oxidation into protons, electrons and O2 (Krassen et al. 2009). Then, ferredoxin in reduced state is the electron donor for [FeFe]-hydrogenases in both types of photolysis. Thereafter, these reducing equivalents are transported to the chlorophyll α-dimer (P700) in photosystem I (PSI). The P700 is energised upon absorption of sunlight and then discharges electrons to the acceptor site of PSI containing iron–sulphur clusters via the electron transport chain (Hallenbeck 2013). Later, PSII is driven by water splitting where in the electrons are transported to reducing equivalents which then finally reduced by the [FeFe]-hydrogenase (HydA) to H2. Hydrogenase enzyme performs as a discharge valve in presence of protons and electrons from the reduced ferredoxin to produce H2. The activity of hydrogenase enzyme relies on the number of reducing equivalents from either of two photosynthetic processes (Constant and Hallenbeck 2013).

Schematic illustration of the photosynthetic mechanisms for biohydrogen production – (a) direct photolysis involved in oxygenic photosynthesis; (b) indirect photolysis involved in oxygenic photosynthesis; (c) anoxygenic mechanism involved with photosynthetic bacteria
Microalgae are outstanding for their ability of high translation of solar energy to molecular H2 (12–14%) by the oxidation of water molecules (Sambusiti et al. 2015). In direct biophotolysis, O2 is generated as by-product of PSII that is a major suppressor of the hydrogenase enzyme; therefore, it can be operated for short periods of time (Blankenship et al. 1995). In indirect photolysis, the generated electrons and protons are generated and stored as starch during photosynthesis. Under certain stress conditions, the electrons and protons are fed into the plastoquinone pool and then onto HydA via PSI resulting in H2 production. In case of microalgae and cyanobacteria, compounds for reserve accumulate during the Calvin cycle. At night, the compounds in reserve act as source energy for the cell activities. The change from aerobic to anaerobic condition is complemented by termination of a photosynthetic light reaction and generation of a surplus reductant, which is finally converted to H2 by hydrogenase (Allakhverdiev et al. 2010). Under anaerobic conditions, [FeFe]-hydrogenases (HydA1 and HydA2) catalyse the molecular H2 by proton reduction. In cyanobacteria, H2 is produced during nitrogen fixation by action of nitrogenase enzyme (Greenbaum 1988a, b; Melis et al. 2000).
Photosynthetic bacteria utilise the visible and/or near-infrared spectrum to produce H2 by catabolising organic molecules by anoxygenic photosynthesis (Fig. 12.2c). In these bacteria, the [FeFe]-hydrogenase can easily evade the distress of oxygen, since they do not use water as source for electron generation (Melis and Happe 2001; Krassen et al. 2009). In this process, the bacteriochlorophyll (BChl) elicits for the formation of bacteriopheophytin (BPh) upon light absorption. The electrons from BPh are transferred to quinine pool (QA) and then to the cytochrome subunit of the reaction centre that generates a proton gradient for ATP synthesis and finally to H2 (Hallenbeck and Benemann 2002). The efficacy of light energy translation to H2 is significantly higher in photosynthetic bacteria than cyanobacteria since the quantum of light energy requirement is less than the water photolysis. Thus, the photosynthetic bacteria make photo-fermentation process more feasible, viable and its adaptability in consuming a variety of substrates (Chandra and Venkata Mohan 2011, 2014).
12.2.2 Dark Fermentation
Dark fermentation involves a multitude of biochemical metabolic reactions, viz. hydrolysis, acidogenesis, acetogenesis and methanogenesis (Lin et al. 2012). The composite organic matter is catabolised to simple molecules during hydrolysis and consequently fermented to organic acids during acidogenesis (Dahiya et al. 2015). As a special case, H2 production is feasible from acetic acid by microbial consortia of acetogens and homoacetogens. Further, the acetoclastic methanogens translate the volatile fatty acids to methane during methanogenesis (Thi et al. 2016). Most often, acetogens nurture in syntrophic alliance with the hydrogenotrophic methanogens, thereby retaining low partial pressure to allow acidogenesis to become thermodynamically favourable. Therefore, the methanogenic activity is suppressed to increase the yields of bioH2 as a sole metabolic by-product (Venkata Mohan et al. 2008a, b; Goud and Venkata Mohan 2012a; Sarkar et al. 2016).
The organic sugars are first metabolised during the glycolysis to pyruvate, a key precursor for subsequent microbial fermentation (Venkata Mohan et al. 2007b). Consequently, pyruvate can result in a variety of short-chain organic fatty acids, and H2 is also produced during the metabolism (Fig. 12.3a). Anaerobes that are facultative metabolise pyruvate to acetyl-CoA and then to formate catalysed by pyruvate formate lyase and subsequently to hydrogen by formate hydrogen lyase (Nikhil et al. 2014b). Anaerobes that are obligate oxidise pyruvate to acetyl-CoA through pyruvate ferredoxin oxidoreductase. Metabolites formed during anaerobic substrate metabolism raise the accessibility of reducing equivalents within the bacteria (Wong et al. 2014). Protons from the redox mediators separate in presence NADH dehydrogenase enzyme and subsequently reduced to H2 with electrons offered by the oxidised ferredoxin upon action of hydrogenase enzyme. The membrane-bound NADH dehydrogenase, cytochrome complex and other carrier proteins channel the electrons via the quinone (Q) pool (Fig. 12.3b). Uninterrupted interconversions of quinone and protons assist in the transport of electrons to cytochrome b–c 1 complex and then to cytochrome aa 3. Ultimately, the electrons are transferred from cytochrome aa 3 to iron-containing protein ferredoxin. The ferredoxin in reduced state transfers electrons to the catalytic site of hydrogenase where protons are combined to form H2 (Saratale et al. 2013).

(a) Different biochemical steps involved in the dark fermentation of glucose (organic matter); (b) schematic illustration of substrate conversion and H2 production mechanism during dark fermentation
Dark fermentation has gained significant credit as a practically feasible scheme amongst the other biological ways for H2 production, especially when wastewater is considered as a substrate and assorted bacterial consortium as a biocatalyst (Agler et al. 2011; Nikhil et al. 2014a). The prominent features of this process are: process simplicity, less energy intensive, less carbon footprints, use of broad spectrum of organics and operation at ambient temperatures. Besides, the process is economically steady and robust for large scale of H2 production (Venkata Mohan 2009). If the fermentation favours for the acetic acid synthesis, then the stoichiometric yield of H2 is 4 mol per mol of glucose, whereas if the fermentation pathway is butyric acid, the yield of H2 is 2 mol per mol of glucose. Yet, the experimental H2 yield is lower than the stoichiometric yield, since a fraction of the substrate is consumed for bacterial cell mass and in few cases by-products might reduce the H2 yield. Strategies are developed to attain superior H2 yields at better rates, viz. novel reactor designs, variable mode of operation, inoculum type and pretreatment, nature and pretreatment of substrate and many more (Venkata Mohan et al. 2008a; Monlau et al. 2015; Sarkar et al. 2016).
The potential sources for anaerobic cultures are found almost in all types of natural environments. Anaerobic bacteria under the class of Firmicutes, which are apt biocatalysts for H2 production (Venkata Mohan et al. 2011). A mixture of bacterial flora is largely preferred for continuous H2 production; however, the microbial culture can show a divergence in H2 production competence because of the co-occurrence of H2-consuming bacteria in overall microbial diversity. In such conditions, conditioning of inoculum is performed to augment H2 producers due to their compliance in hostile environments (Sarkar et al. 2013; Goud et al. 2017). Moreover, proton fluxes in/out bacteria affects the enzyme activity, biochemical pathways and substrate decomposition. pH is also acute to sustain ample cellular ATP levels, since surplus H+ ions are pumped out using ATP to ensure cell neutrality (Srikanth and Venkata Mohan 2012). The initial pH can impact the duration of lag phase involved in spore germination and enzyme synthesis. The operational organic load of bioreactor can affect several functional issues which include accumulation of undissociated VFA and pH variations. This subsequently changes the diversity of microbial flora with subsequent amendments of the allied metabolic pathways (Van Ginkel and Logan 2005a; Goud and Venkata Mohan 2012b). Commercial biohydrogen production still poses certain process impede like the inhibitory effect caused by the undissociated VFA (Van Ginkel and Logan 2005b; Goud et al. 2014; Srikanth and Venkata Mohan 2014; Sarkar et al. 2017). Improving H2 production rate and yield are the grave challenges to sustain economic production. In this regard, various strategies were reported in the literature. A few of them are selection and pretreatment of microbial consortia, immobilisation of consortia, statistical techniques for process optimisation, sequencing of bioreactors, bio-electrochemical treatment, multiple process integration and bio-augmentation (Pasupuleti and Venkata Mohan 2015a, b). Commercial operation still poses certain process impedes like inhibitory effect of undissociated VFA (Van Ginkel and Logan 2005b; Goud et al. 2014; Srikanth and Venkata Mohan 2014; Sarkar et al. 2017).
12.2.3 Integration Strategies
12.2.3.1 Biohythane
A proportionate mixture of hydrogen and methane is commercially available as Hythane™, HCNG or methagen (Eden 2010). The recommended composition of H2 with CH4 ranges between 10% (v/v) and 25% (v/v) (Liu et al. 2013). Conventional methods could be unsustainable due to their reliance on exhausting petroleum reserves. Yet, the contemporary trends in bioenergy exploit the different biological routes for the production of both methane and hydrogen from waste organic matter. Literature reports are available on two-step fermentation process to produce a mixture of H2 and CH4 from wastewater (Venkata Mohan et al. 2008b; Mohanakrishna and Venkata Mohan 2013). This biological design assures the composition of biogas mixture (CH4 + H2) by regulating the environments of microbial fermentation. The usual operation is a two-step process that has a few advantages, but operation of two bioreactors is frugally not that sustainable (Cavinato et al. 2012; Willquist et al. 2012). A recent investigation reported biohythane from single-stage bioreactor using spent-wash wastewater (Pasupuleti and Venkata Mohan 2015b). In another study, a unique strategy was reported that augments acidogenesis for the production of biohythane (Sarkar and Venkata Mohan 2016, 2017).
12.2.3.2 Hybrid Dark–Photo Fermentation
A blend of both dark and photo-fermentation is a credible option to accomplish a yield of 12 mol H2/mol glucose. Integrative fermentation can be considered as an operational and competent option to harness maximum H2 yield with simultaneous wastewater treatment (Srikanth et al. 2009; Laurinavichene et al. 2012). A variety of photosynthetic bacteria are capable of H2 production which can uptake the volatile fatty acids as carbon source and light as energy sources. Studies have been carried out with two-stage integration of heterotrophic dark with photo-fermentation for bioH2 production (Chandra et al. 2015). It was noticed that photo-fermentation of acid-rich effluents is considered to be complicated due to light infiltration complications, multifaceted nutritional requirements, obligatory operational conditions, substrate (VFA and ammonia) inhibitions and vulnerability for impurity (Wang et al. 2009; Chandra and Venkata Mohan 2014).
12.2.3.3 Microbial Electrolysis
Microbial electrolysis is an electrically driven H2 production process wherein the exchange of electron equivalents in carbon-based composites to H2 gas involves bio-electrochemical reactions (Call and Logan 2008). A microbial electrochemical cell (MEC) has a prerequisite of additional potential to assist the metabolism of undissociated organic acids into H2 which is obligatory to cross the endothermic barrier (Cheng and Logan 2011). Therefore, the protons and electrons are driven with the supplementation of an external voltage from anode to cathode which is reduced to H2. The typical redox potential for H2 reduction is −0.414 V, and a voltage >0.11 V facilitates H2 formation at the cathode. These applied potentials are moderately low compared to the applied external potential of 1.23 V for electrolysis of water. For example, H2 (−0.414 V) from acetate (−0.279 V) at the cathode occurs upon appliance of voltage (−0.135 V). In reality, a relatively higher voltage is required than the prerequisite; owing to system overpotentials aroused by physicochemical and microbial influences.
It is reported that the applied potentials selectively enrich the growth of electroactive bacteria that efficiently reduce electrons (Arunasri et al. 2016). Operating an MEC can recover more than 90% of H2 as against 33% by the dark fermentation. Thus, the application of MEC can be seen with the usage of acidogenic effluents rich in short-chain fatty acids for recovery of additional H2/CH4 production (Wagner et al. 2009; Modestra et al. 2015). The prospects of MEC over conventional water electrolysis are low-energy consumption, appreciable H2 yields and wastewater treatment. Nevertheless, biocatalyst diversity, electrode materials, membrane, applied potential, substrate loading rate and reactor configuration critically affect the functioning of MEC (Nam et al. 2011). To begin with, studies were carried out in double chamber that allows separate capture of H2 at cathode and prevents fouling by anodic bacteria (Pisciotta et al. 2012). But, separation leads to acidification at the anode chamber; so, removing the partition creates a single-chamber MEC that reduces the applied potential attenuating the pH and energy losses. Electrodes coated with platinum are commonly used as chemical catalysts for H2 evolution as it significantly diminishes the cathode overpotential. Conventional methods use platinum which is costly, consumable and vulnerable to contamination by constituents in the effluents. Recently, research on biocathodes is of prime attention as they are eco-friendly and renewable (Jeremiasse et al. 2010; Nikhil et al. 2015b).
During the operation of MEC, the hydrogen evolution reaction (HER) is linked to the pH. The buffer capacity of wastewater is typically low resulting in accumulation of protons resulting in acidification at anode (Hamelers et al. 2010). The use of chemical buffers could be a possible option that alleviates the pH gradient. The efficacy of a buffer relies on its dissociation constant (pKa) and diffusivity. Buffers with pKa slightly above the operating pH (near neutral) aid to sustain the internal pH close to the external pH (Zhu et al. 2009; Liang et al. 2014). Inappropriate use of buffer (above 300 mM) affects the system and unsuitable for the effluent discharge; besides addition of salts would escalate the operating economics (Ambler and Logan 2011). It was reported that the external voltage exhibited a dual consequence over process performance by regulating the in situ buffering capacity using a biocathode (Lenin Babu et al. 2013b; Nikhil et al. 2015a). Integrating MEC with other processes to harvest biohydrogen generation is the state-of-the-art research amongst the scientific fraternity (Escapa et al. 2014). Small additional voltage with a high current density will be a crucial challenge for scaling-up of MEC to mercantile applications (Lenin Babu et al. 2013a; Venkata Mohan et al. 2014).
12.2.3.4 Bio-based Products
The dark-fermentative effluents are rich in VFA, and these can be used as possible substrate for production of polyhydroxyalkanoates (PHAs) commonly referred to as ‘bioplastics’ (Venkata Mohan et al. 2010; Sarkar et al. 2016). PHAs are biopolyesters of hydroxyalkanoates that amass as cellular storage materials produced under surplus carbon and nutrient-deprived environments (Patel et al. 2012; Fradinho et al. 2013). Commercial scale of PHA is produced using plant-derived resources and pure cultures that rise the production costs. Compared to other substrates (carbohydrates or proteins), the use of VFA facilitates PHA synthesis depriving the participation of glycolysis and β-oxidation pathways. Reports are available on PHA production from synthetic VFA and biohydrogen effluents (VFA-rich) under an anoxic microenvironment using a mixed bacterial culture (Venkateswar Reddy and Venkata Mohan 2012). If acetate and butyrate compositions are relatively higher amounts, then polyhydroxybutyrate (PHB) is the product type, and if propionate and valerate are relatively higher amounts, then polyhydroxyvalerate (PHV) is the product type. The combined production of H2 and PHA followed by methanogenesis is possible option to make the whole process economical and sustainable (Patel et al. 2012; Venkateswar Reddy et al. 2014).
12.3 Waste Treatment vs Biohydrogen Production
Ideally, a unit kilogram of chemical oxygen demand (COD) equals to 5.2 mol of glucose, and upon dark fermentation each mole of glucose results in 4 mol and 2 mol of H2 depending upon acetic acid and butyric acid pathways, respectively (Venkata Mohan et al. 2007c). On the other hand, photo-fermentation yields 12 mol of H2 per mol of glucose. H2 produced by acetate pathway of dark fermentation yields 89.6 l of H2 per mol of glucose which is equivalent to 466.6 l of H2 per kg COD. Assuming with only 40% of COD removal, then H2 yield in dark and photo-fermentation is 125 g and 16.6 g, respectively. For example, if an industry discharging ~3 × 1011 l of effluent per annum containing a typical COD of 20 g/l, then it amounts to 6000 × 106 g of COD for that year. The yields of H2 with dark and photo-fermentation are about 5 × 106 kg and 300 × 106 kg per year that accounts for many million dollars/year (at a rate of $ 4/kg H2) (Markets 2011).
Rapid industrialisation is resulting in immense amounts of waste. Conventional treatment of wastewater is an energy-exhaustive process. Probing means to produce or harness valuable products from wastewater remediation are significantly pursued in recent times (Angenent et al. 2004; Venkata Mohan et al. 2010). The prerequisite requirement for their management markedly makes wastewater a choice for biohydrogen production, besides abating the overall treatment cost (Guo et al. 2010; Elsharnouby et al. 2013). Till date, considerable efforts were put on the application of various wastewaters for the production of biohydrogen through fermentation processes (both light-dependent and light-independent) (Hallenbeck 2013). Both wastewater treatment and H2 production are equally important, and a balance should exist between technical expertise and substrate uptake with operating circumstances (Show and Lee 2013). Assessing these conditions is particularly important in sustaining the financial possibility and environmental competence of the process (Arimi et al. 2015). An illustrative plan of strategies to recover energy and products for value additions along with wastewater treatment is represented in Fig. 12.4.

Schematic details of various strategic routes possible for effective H2 production and other value-added products
12.4 Future Perspectives
Hydrogen is an imperative substitute for energy domain and can be produced from a variety of production technologies (Dunn 2002; Nouni 2012). Presently, investigations are being dedicated on viable and eco-friendly biohydrogen from biomass to substitute fossil fuels (Hallenbeck and Ghosh 2009; Rama Mohan 2015). Waste biomass can be deliberated as the paramount choice and has the foremost prospective that encounters energy supplies and could assure fuel stock in the upcoming future. Utilisation of remaining organic portion after acidogenesis is also the most significant aspect to be paid momentous consideration (Mohanakrishna et al. 2010; Ghimire et al. 2015). Photobiological processes with acidogenic wastewater as substrate are comparatively less studied and need further exploitation. Multi-process combination is effectively evaluated for economic viability and commercialisation (Mohanakrishna and Venkata Mohan 2013; Wang and Ren 2013). Overall, a biorefinery concept with closed-loop approach visualises negative-valued waste as a potential renewable feedstock that will pave new opportunities for the growth of bio-based society with circular economy (Venkata Mohan et al. 2016).
References
Agler MT, Wrenn BA, Zinder SH, Angenent LT (2011) Waste to bioproduct conversion with undefined mixed cultures: the carboxylate platform. Trends Biotechnol 29:70–78. https://doi.org/10.1016/j.tibtech.2010.11.006
Allakhverdiev SI, Thavasi V, Kreslavski VD, Zharmukhamedov SK, Klimo VV, Ramakrishna S, Los DA, Mimurod M, Nishiharae H, Carpentier R (2010) Photosynthetic hydrogen production. J Photochem Photobiol C: Photochem Rev 11:101–113. https://doi.org/10.1016/j.jphotochemrev.2010.07.002
Ambler JR, Logan BE (2011) Evaluation of stainless steel cathodes and a bicarbonate buffer for hydrogen production in microbial electrolysis cells using a new method for measuring gas production. Int J Hydrog Energy 36:160–166. https://doi.org/10.1016/j.ijhydene.2010.09.044
Angenent LT, Karim K, Al-Dahhan MH, Wrenn BA, Domíguez-Espinosa R (2004) Production of bioenergy and biochemicals from industrial and agricultural wastewater. Trends Biotechnol 22:477–485. https://doi.org/10.1016/j.tibtech.2004.07.001
Arimi MM, Knodel J, Kiprop A, Namango SS, Zhang Y, Geißen S-U (2015) Strategies for improvement of biohydrogen production from organicrich wastewater: a review. Biomass Bioenergy 75:101–118. https://doi.org/10.1016/j.biombioe.2015.02.011
Arunasri K, Modestra JA, Yeruva DK, Krishna KV, Venkata Mohan S (2016) Polarized potential and electrode materials implication on electrofermentative di-hydrogen production: microbial assemblages and hydrogenase gene copy variation. Bioresour Technol 200:691–698. https://doi.org/10.1016/j.biortech.2015.10.091
Bhaskar T, Balagurumurthy B, Singh R, Poddar MK (2013) Thermochemical route for biohydrogen production. In: Pandey A, Chang JS, Hallenbeck P, Larroche C (eds) Biohydrogen. Elsevier, Amsterdam, pp 285–316. ISBN: 978-0-444-59555-3
Blankenship RE, Olson JM, Miller M (1995) Antenna complexes from green photosynthetic bacteria. In: Blankenship RE, Madigan Bauer MT (eds) Anoxygenic photosynthetic bacteria. Springer, Dordrecht. ISBN: 978-0-7923-3681-5, pp 399–435. https://doi.org/10.1007/0-306-47954-0_20
Call D, Logan BE (2008) Hydrogen production in a single chamber microbial electrolysis cell lacking a membrane. Environ Sci Technol 42:3401–3406. https://doi.org/10.1021/es8001822
Cavinato C, Giuliano A, Bolzonella D, Pavan P, Cecchi F (2012) Bio-hythane production from food waste by dark fermentation coupled with anaerobic digestion process: a long-term pilot scale experience. Int J Hydrog Energy 37:11549–11555. https://doi.org/10.1016/j.ijhydene.2012.03.065
Chandra R, Venkata Mohan S (2011) Microalgal community and their growth conditions influence biohydrogen production during integration of dark-fermentation and photo-fermentation processes. Int J Hydrog Energy 36:12211–12219. https://doi.org/10.1016/j.ijhydene.2011.07.007
Chandra R, Venkata Mohan S (2014) Enhanced bio-hydrogenesis by co-culturing photosynthetic bacteria with acidogenic process: augmented dark-photo fermentative hybrid system to regulate volatile fatty acid inhibition. Int J Hydrog Energy 39:7604–7615. https://doi.org/10.1016/j.ijhydene.2014.01.196
Chandra R, Nikhil GN, Venkata Mohan S (2015) Single-stage operation of hybrid dark-photo fermentation to enhance biohydrogen production through regulation of system redox condition: evaluation with real-field wastewater. Int J Mol Sci 16:9540–9556. https://doi.org/10.3390/ijms16059540
Cheng S, Logan BE (2011) High hydrogen production rate of microbial electrolysis cell (MEC) with reduced electrode spacing. Bioresour Technol 102:3571–3574. https://doi.org/10.1016/j.biortech.2010.10.025
Constant P, Hallenbeck PC (2013) Hydrogenase. In: Pandey A, Chang JS, Hallenbeck P, Larroche C (eds) Biohydrogen. Elsevier, Amsterdam, pp 75–102. ISBN: 978-0-444-59555-3
Dahiya S, Sarkar O, Swamy Y, Venkata Mohan S (2015) Acidogenic fermentation of food waste for volatile fatty acid production with cogeneration of biohydrogen. Bioresour Technol 182:103–113. https://doi.org/10.1016/j.biortech.2015.01.007
Dunn S (2002) Hydrogen futures: toward a sustainable energy system. Int J Hydrog Energy 27:235–264. https://doi.org/10.1016/S0360-3199(01)00131-8
Eden (2010) Annual Report. Available from: www.edenenergy.com.au
Elsharnouby O, Hafez H, Nakhla G, El Naggar MH (2013) A critical literature review on biohydrogen production by pure cultures. Int J Hydrog Energy 38:4945–4966. https://doi.org/10.1016/j.ijhydene.2013.02.032
Escapa A, San Martin MI, Moran A (2014) Potential use of microbial electrolysis cells in domestic wastewater treatment plants for energy recovery. Front Energy Res 2:1–10. https://doi.org/10.3389/fenrg.2014.00019
Fradinho J, Domingos J, Carvalho G, Oehmen A, Reis M (2013) Polyhydroxyalkanoates production by a mixed photosynthetic consortium of bacteria and algae. Bioresour Technol 132:146–153. https://doi.org/10.1016/j.biortech.2013.01.050
Ghimire A, Frunzo L, Pirozzi F, Trably E, Escudie R, Lens PNL, Esposito G (2015) A review on dark fermentative biohydrogen production from organic biomass: process parameters and use of by-products. Appl Energy 144:73–95. https://doi.org/10.1016/j.apenergy.2015.01.045
Goud RK, Venkata Mohan S (2012a) Acidic and alkaline shock pretreatment to enrich acidogenic biohydrogen producing mixed culture: long term synergetic evaluation of microbial inventory, dehydrogenase activity and bio-electro kinetics. RSC Adv 2:6336–6353. https://doi.org/10.1039/C2RA20526B
Goud RK, Venkata Mohan S (2012b) Regulating biohydrogen production from wastewater by applying organic load-shock: change in the microbial community structure and bio-electrochemical behavior over long-term operation. Int J Hydrog Energy 37:17763–17777. https://doi.org/10.1016/j.ijhydene.2012.08.124
Goud RK, Sarkar O, Chiranjeevi P, Venkata Mohan S (2014) Bioaugmentation of potent acidogenic isolates: a strategy for enhancing biohydrogen production at elevated organic load. Bioresour Technol 165:223–232. https://doi.org/10.1016/j.biortech.2014.03.049
Goud RK, Arunasri K, Yeruva DK, Krishna KV, Dahiya S, Venkata Mohan S (2017) Impact of selectively enriched microbial communities on longterm fermentative biohydrogen production. Bioresour Technol. https://doi.org/10.1016/j.biortech.2017.03.147
Greenbaum E (1988a) Energetic efficiency of hydrogen photoevolution by algal water splitting. Biophys J 54:365–368. https://doi.org/10.1016/S0006-3495(88)82968-0
Greenbaum E (1988b) Interfacial photoreactions at the photosynthetic membrane interface: an upper limit for the number of platinum atoms required to form a hydrogen-evolving platinum metal catalyst. J Phys Chem 92:4571–4574. https://doi.org/10.1021/j100327a001
Guo XM, Trably E, Latrille E, Carrère H, Steyer J-P (2010) Hydrogen production from agricultural waste by dark fermentation: a review. Int J Hydrog Energy 35:10660–10673. https://doi.org/10.1016/j.ijhydene.2010.03.008
Hallenbeck PC (2013) Photofermentative biohydrogen production. In: Pandey A, Chang JS, Hallenbeck P, Larroche C (eds) Biohydrogen. Elsevier, Amsterdam, pp 145–159. ISBN: 978-0-444-59555-3
Hallenbeck PC, Benemann JR (2002) Biological hydrogen production; fundamentals and limiting processes. Int J Hydrog Energy 27:1185–1193. https://doi.org/10.1016/S0360-3199(02)00131-3
Hallenbeck PC, Ghosh D (2009) Advances in fermentative biohydrogen production: the way forward? Trends Biotechnol 27:287–297. https://doi.org/10.1016/j.tibtech.2009.02.004
Hamelers HV, Ter Heijne A, Sleutels TH, Jeremiasse AW, Strik DP, Buisman CJ (2010) New applications and performance of bioelectrochemical systems. Appl Microbiol Biotechnol 85:1673–1685. https://doi.org/10.1007/s00253-009-2357-1
Jeremiasse AW, Hamelers HVM, Buisman CJN (2010) Microbial electrolysis cell with a microbial biocathode. Bioelectrochemistry 78:39–43. https://doi.org/10.1016/j.bioelechem.2009.05.005
Krassen H, Schwarze A, Friedrich B, Ataka K, Lenz O, Heberle J (2009) Photosynthetic hydrogen production by a hybrid complex of photosystem I and [NiFe]-hydrogenase. ACS Nano 3:4055–4061. https://doi.org/10.1021/nn900748j
Lam MK, Lee KT (2013) Biohydrogen production from algae. In: Pandey A, Chang JS, Hallenbeck P, Larroche C (eds) Biohydrogen. Elsevier, Amsterdam, pp 161–184. ISBN: 978-0-444-59555-3
Laurinavichene TV, Belokopytov BF, Laurinavichius KS, Khusnutdinova AN, Seibert M, Tsygankov AA (2012) Towards the integration of darkand photo-fermentative waste treatment. 4. Repeated batch sequential dark- and photofermentation using starch as substrate. Int J Hydrog Energy 37:8800–8810. https://doi.org/10.1016/j.ijhydene.2012.01.132
Lenin Babu M, Sarma P, Venkata Mohan S (2013a) Microbial electrolysis of synthetic acids for biohydrogen production: influence of biocatalyst pretreatment and pH with the function of applied potential. J Microb Biochem Technol S 6:2. https://doi.org/10.4172/1948-5948.S6-003
Lenin Babu M, Subhash GV, Sarma PN, Venkata Mohan S (2013b) Bio-electrolytic conversion of acidogenic effluents to biohydrogen: an integration strategy for higher substrate conversion and product recovery. Bioresour Technol 133:322–331. https://doi.org/10.1016/j.biortech.2013.01.029
Liang D, Liu Y, Peng S, Lan F, Lu S, Xiang Y (2014) Effects of bicarbonate and cathode potential on hydrogen production in a biocathode electrolysis cell. Front Environ Sci Eng 8:624–630. https://doi.org/10.1007/s11783-013-0584-2
Lin C-Y, Lay C-H, Sen B, Chu C-Y, Kumar G, Chen C-C, Chang J-S (2012) Fermentative hydrogen production from wastewaters: a review and prognosis. Int J Hydrog Energy 37:15632–15642. https://doi.org/10.1016/j.ijhydene.2012.02.072
Liu Z, Zhang C, Lu Y, Wu X, Wanf L, Wang L, Han B, Xing QH (2013b) States and challenges for high-value biohythane production from waste biomass by dark fermentation technology. Bioresour Technol 135:292–303. https://doi.org/10.1016/j.biortech.2012.10.027
Markets and Markets (2011) Hydrogen generation market-by merchant and captive type, distributed and centralized generation, application and technology-trends and global forecasts (2011–2016). Report code: EP1708. http://www.marketsandmarkets.com/Market-Reports/hydrogengeneration-market-494.html
Melis A, Happe T (2001) Hydrogen production. Green algae as a source of energy. Plant Physiol 127:740–748. https://doi.org/10.1104/pp.010498
Melis A, Zhang L, Forestier M, Ghirardi ML, Seibert M (2000) Sustained photobiological hydrogen gas production upon reversible inactivation of oxygen evolution in the green alga Chlamydomonas reinhardtii. Plant Physiol 122:127–136. https://doi.org/10.1104/pp.122.1.127
Modestra JA, Babu ML, Venkata Mohan S (2015) Electro-fermentation of real-field acidogenic spent wash effluents for additional biohydrogen production with simultaneous treatment in a microbial electrolysis cell. Sep Purif Technol 150:308–315. https://doi.org/10.1016/j.seppur.2015.05.043
Mohanakrishna G, Venkata Mohan S (2013) Multiple process integrations for broad perspective analysis of fermentative H production from wastewater treatment: technical and environmental considerations. Appl Energy 107:244–254. https://doi.org/10.1016/j.apenergy.2013.01.085
Mohanakrishna G, Venkata Mohan S, Sarma PN (2010) Utilizing acid-rich effluents of fermentative hydrogen production process as substrate for harnessing bioelectricity: an integrative approach. Int J Hydrog Energy 35:3440–3449. https://doi.org/10.1016/j.ijhydene.2010.01.084
Monlau F, Kaparaju P, Trably E, Steyer J-P, Carrere H (2015) Alkaline pretreatment to enhance one-stage CH and two-stage H /CH production from sunflower stalks: mass, energy and economical balances. Chem Eng J 260:377–385. https://doi.org/10.1016/j.cej.2014.08.108
Nam J-Y, Tokash JC, Logan BE (2011) Comparison of microbial electrolysis cells operated with added voltage or by setting the anode potential. Int J Hydrog Energy 36:10550–10556. https://doi.org/10.1016/j.ijhydene.2011.05.148
Nikhil GN, Venkata Mohan S, Swamy YV (2014a) Behavior of acidogenesis during biohydrogen production with formate and glucose as carbon source: substrate associated dehydrogenase expression. Int J Hydrog Energy 39:7486–7495. https://doi.org/10.1016/j.ijhydene.2013.12.169
Nikhil GN, Venkata Venkata Mohan S, Swamy YV (2014b) Systematic approach to assess biohydrogen potential of anaerobic sludge and soil rhizobia as biocatalysts: influence of crucial factors affecting acidogenic fermentation. Bioresour Technol 165:323–331. https://doi.org/10.1016/j.biortech.2014.02.097
Nikhil GN, Subhash GV, Yeruva DK, Venkata Mohan S (2015a) Synergistic yield of dual energy forms through biocatalyzed electrofermentation of waste: stoichiometric analysis of electron and carbon distribution. Energy 88:281–291. https://doi.org/10.1016/j.energy.2015.05.043
Nikhil GN, Venkata Mohan S, Swamy YV (2015b) Applied potentials regulate recovery of residual hydrogen from acid-rich effluents: influence of biocathodic buffer capacity over process performance. Bioresour Technol 188:65–72. https://doi.org/10.1016/j.biortech.2015.01.084
Nissilä ME, Lay C-H, Puhakka JA (2014) Dark fermentative hydrogen production from lignocellulosic hydrolyzates – a review. Biomass Bioenergy 67:145–159. https://doi.org/10.1016/j.biombioe.2014.04.035
Nouni M (2012) Hydrogen energy and fuel cell technology: recent developments and future prospects in India. Renew Energy. Akshay Urja 5:10–14
Pasupuleti SB, Venkata Mohan S (2015a) Acidogenic hydrogen production from wastewater: process analysis with the function of influencing parameters. Int J Energy Res 39:1131–1141. https://doi.org/10.1002/er.3322
Pasupuleti SB, Venkata Mohan S (2015b) Single-stage fermentation process for high-value biohythane production with the treatment of distillery spent-wash. Bioresour Technol 189:177–185. 10.1016/j.biortech.2015.03.128
Patel SK, Singh M, Kumar P, Purohit HJ, Kalia VC (2012) Exploitation of defined bacterial cultures for production of hydrogen and polyhydroxybutyrate from pea-shells. Biomass Bioenergy 36:218–225. https://doi.org/10.1016/j.biombioe.2011.10.027
Pisciotta JM, Zaybak Z, Call DF, Nam JY, Logan BE (2012) Enrichment of microbial electrolysis cell biocathodes from sediment microbial fuel cell bioanodes. Appl Environ Microbiol 78:5212–5219. https://doi.org/10.1128/AEM.00480-12
Rama Mohan S (2015) Structure and growth of research on biohydrogen generation using wastewater. Int J Hydrog Energy 40:16056–16069. https://doi.org/10.1016/j.ijhydene.2015.08.072
Sambusiti C, Bellucci M, Zabaniotou A, Beneduce L, Monlau F (2015) Algae as promising feedstocks for fermentative biohydrogen production according to a biorefinery approach: a comprehensive review. Renew Sustain Energy Rev 44:20–36. https://doi.org/10.1016/j.rser.2014.12.013
Saratale GD, Saratale RG, Chang J-S (2013) Biohydrogen from renewable resources. In: Pandey A, Chang JS, Hallenbeck P, Larroche C (eds) Biohydrogen. Elsevier, Amsterdam, pp 185–221. ISBN: 978-0-444-59555-3
Sarkar O, Venkata Mohan S (2016) Deciphering acidogenic process towards biohydrogen, biohythane, and short chain fatty acids production: multi-output optimization strategy. Biofuel Res J 3:458–469. https://doi.org/10.18331/BRJ2016.3.3.5
Sarkar O, Venkata Mohan S (2017) Pre-aeration of food waste to augment acidogenic process at higher organic load: valorizing biohydrogen, volatile fatty acids and biohythane. Bioresour Technol. https://doi.org/10.1016/j.biortech.2017.05.053
Sarkar O, Goud RK, Subhash GV, Venkata Mohan S (2013) Relative effect of different inorganic acids on selective enrichment of acidogenic biocatalyst for fermentative biohydrogen production from wastewater. Bioresour Technol 147:321–331. https://doi.org/10.1016/j.biortech.2013.08.021
Sarkar O, Kumar AN, Dahiya S, Krishna KV, Yeruva DK, Venkata Mohan S (2016) Regulation of acidogenic metabolism towards enhanced short chain fatty acid biosynthesis from waste: metagenomic profiling. RSC Adv 6:18641–18653. https://doi.org/10.1039/C5RA24254A
Sarkar O, Butti SK, Venkata Mohan S (2017) Acidogenesis driven by hydrogen partial pressure towards bioethanol production through fatty acids reduction. Energy 118:425–434. https://doi.org/10.1016/j.energy.2016.12.017
Show K-Y, Lee D-J (2013) Bioreactor and bioprocess design for biohydrogen production. In: Larroche AP-SCCH (ed) Biohydrogen. Elsevier, Amsterdam. In: Pandey A, Chang JS, Hallenbeck P, Larroche C (eds) Biohydrogen, pp 317–337. Elsevier, Amsterdam. ISBN: 978-0-444-59555-3
Srikanth S, Venkata Mohan S (2012) Regulatory function of divalent cations in controlling the acidogenic biohydrogen production process. RSC Adv 2:6576–6589. https://doi.org/10.1039/C2RA20383A
Srikanth S, Venkata Mohan S (2014) Regulating feedback inhibition caused by the accumulated acid intermediates during acidogenic hydrogen production through feed replacement. Int J Hydrog Energy 39:10028–10040. https://doi.org/10.1016/j.ijhydene.2014.04.152
Srikanth S, Venkata Mohan S, Prathima Devi M, Peri D, Sarma PN (2009) Acetate and butyrate as substrates for hydrogen production through photo-fermentation: process optimization and combined performance evaluation. Int J Hydrog Energy 34:7513–7522. https://doi.org/10.1016/j.ijhydene.2009.05.095
Thi NBD, Lin C-Y, Kumar G (2016) Waste-to-wealth for valorization of food waste to hydrogen and methane towards creating a sustainable ideal source of bioenergy. J Clean Prod 122:29–41. https://doi.org/10.1016/j.jclepro.2016.02.034
Van Ginkel S, Logan BE (2005a) Inhibition of biohydrogen production by undissociated acetic and butyric acids. Environ Sci Technol 39:9351–9356. https://doi.org/10.1021/es0510515
Van Ginkel SW, Logan B (2005b) Increased biological hydrogen production with reduced organic loading. Water Res 39:3819–3826. https://doi.org/10.1016/j.watres.2005.07.021
Venkata Mohan S (2009) Harnessing of biohydrogen from wastewater treatment using mixed fermentative consortia: process evaluation towards optimization. Int J Hydrog Energy 34:7460–7474. https://doi.org/10.1016/j.ijhydene.2009.05.062
Venkata Mohan S, Lalit Babu V, Sarma PN (2007a) Anaerobic biohydrogen production from dairy wastewater treatment in sequencing batch reactor (AnSBR): effect of organic loading rate. Enzym Microb Technol 41:506–515. https://doi.org/10.1016/j.enzmictec.2007.04.007
Venkata Mohan S, Vijaya Bhaskar Y, Murali Krishna P, Chandrasekhara Rao N, Lalit Babu V, Sarma PN (2007b) Biohydrogen production from chemical wastewater as substrate by selectively enriched anaerobic mixed consortia: influence of fermentation pH and substrate composition. Int J Hydrog Energy 32:2286–2295. https://doi.org/10.1016/j.ijhydene.2007.03.015
Venkata Mohan S, Vijaya Bhaskar Y, Sarma PN (2007c) Biohydrogen production from chemical wastewater treatment in biofilm configured reactor operated in periodic discontinuous batch mode by selectively enriched anaerobic mixed consortia. Water Res 41:2652–2664. https://doi.org/10.1016/j.watres.2007.02.015
Venkata Mohan S, Babu VL, Sarma P (2008a) Effect of various pretreatment methods on anaerobic mixed microflora to enhance biohydrogen production utilizing dairy wastewater as substrate. Bioresour Technol 99:59–67. https://doi.org/10.1016/j.biortech.2006.12.004
Venkata Mohan S, Mohanakrishna G, Sarma P (2008b) Integration of acidogenic and methanogenic processes for simultaneous production of biohydrogen and methane from wastewater treatment. Int J Hydrog Energy 33:2156–2166. https://doi.org/10.1016/j.ijhydene.2008.01.055
Venkata Mohan S, Venkateswar Reddy M, Venkata Subhash G, Sarma PN (2010) Fermentative effluents from hydrogen producing bioreactor as substrate for poly (β -OH) butyrate production with simultaneous treatment: an integrated approach. Bioresour Technol 101:9382–9386. https://doi.org/10.1016/j.biortech.2010.06.109
Venkata Mohan S, Agarwal L, Mohanakrishna G, Srikanth S, Kapley A, Purohit HJ, Sarma PN (2011) Firmicutes with iron dependent hydrogenase drive hydrogen production in anaerobic bioreactor using distillery wastewater. Int J Hydrog Energy 36:8234–8242. https://doi.org/10.1016/j.ijhydene.2011.04.021
Venkata Mohan S, Velvizhi G, Vamshi Krishna K, Lenin Babu M (2014) Microbial catalyzed electrochemical systems: a bio-factory with multi-facet applications. Bioresour Technol 165:355–364. https://doi.org/10.1016/j.biortech.2014.03.048
Venkata Mohan S, Nikhil GN, Chiranjeevi P, Reddy CN, Rohit MV, Kumar AN, Sarkar O (2016) Waste biorefinery models towards sustainable circular bioeconomy: crit rev future perspect. Bioresour Technol 215:2–12. https://doi.org/10.1016/j.biortech.2016.03.130
Venkata Mohan S, Srikanth S, Nikhil GN (2017) Augmentation of bacterial homeostasis by regulating in situ buffer capacity: significance of total dissolved salts over acidogenic metabolism. Bioresour Technol 225:34–39. https://doi.org/10.1016/j.biortech.2016.11.027
Venkateswar Reddy M, Venkata Mohan S (2012) Influence of aerobic and anoxic microenvironments on polyhydroxyalkanoates (PHA) production from food waste and acidogenic effluents using aerobic consortia. Bioresour Technol 103:313–321. https://doi.org/10.1016/j.biortech.2011.09.040
Venkateswar Reddy M, Chitanya DNSK, Nikhil GN, Venkata Mohan S, Sarma PN (2014) Influence of co-factor on enhancement of bioplastic production through wastewater treatment. Clean Soil Air Water 42:809–814. https://doi.org/10.1002/clen.201300105
Wagner RC, Regan JM, S-E O, Zuo Y, Logan BE (2009) Hydrogen and methane production from swine wastewater using microbial electrolysis cells. Water Res 43:1480–1488. https://doi.org/10.1016/j.watres.2008.12.037
Wang H, Ren ZJ (2013) A comprehensive review of microbial electrochemical systems as a platform technology. Biotechnol Adv 31:1796–1807. https://doi.org/10.1016/j.biotechadv.2013.10.001
Wang B, Wan W, Wang J (2009) Effect of ammonia concentration on fermentative hydrogen production by mixed cultures. Bioresour Technol 100:1211–1213. https://doi.org/10.1016/j.biortech.2008.08.018
Willquist K, Nkemka VN, Svensson H, Pawar S, Ljunggren M, Karlsson H, Murto M, Hulteberg C, van Niel EWJ, Liden G (2012) Design of a novel biohythane process with high H and CH production rates. Int J Hydrog Energy 37:17749–17762. https://doi.org/10.1016/j.ijhydene.2012.08.092
Wong YM, TY W, Juan JC (2014) A review of sustainable hydrogen production using seed sludge via dark fermentation. Renew Sustain Energy Rev 34:471–482. https://doi.org/10.1016/j.rser.2014.03.008
Zhu H, Parker W, Basnar R, Proracki A, Falletta P, Béland M, Seto P (2009) Buffer requirements for enhanced hydrogen production in acidogenic digestion of food wastes. Bioresour Technol 100:5097–5102. https://doi.org/10.1016/j.biortech.2009.02.066
Acknowledgements
Funding received from the Ministry of New and Renewable Energy (MNRE), Government of India and Council for Scientific and Industrial Research (CSIR) in the form of research grants as MNRE Project No. 103/131/2008-NT, XII 5-year network project (SETCA (CSC-0113), respectively. GNN and OS acknowledge the CSIR for providing Senior Research Fellowship.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Nikhil, G.N., Sarkar, O., Venkata Mohan, S. (2017). Biohydrogen Production: An Outlook of Fermentative Processes and Integration Strategies. In: Purohit, H., Kalia, V., Vaidya, A., Khardenavis, A. (eds) Optimization and Applicability of Bioprocesses . Springer, Singapore. https://doi.org/10.1007/978-981-10-6863-8_12
Download citation
DOI: https://doi.org/10.1007/978-981-10-6863-8_12
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-10-6862-1
Online ISBN: 978-981-10-6863-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)