
Abstract
Model biomembrane vesicles composed of lipids have been widely used to investigate the principles of membrane assembly and organization. A limitation of these vesicles has been that they do not mimic the transbilayer lipid asymmetry seen in many natural membranes, most notably the asymmetry in the plasma membrane of eukaryotic cells. Recently, a number of approaches have been developed to prepare asymmetric membranes and study their properties. This review describes methods to prepare asymmetric model membranes, and the physical properties of asymmetric lipid vesicles. Emphasis is placed on the vesicles prepared by cyclodextrin-catalyzed exchange, which has proven to be a versatile and powerful tool, including for studies manipulating lipid asymmetry in living cells.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
1.1 Lipid Asymmetry: Definition and Origin
When studying biological membrane organization and function, one important aspect to consider is lipid asymmetry. Lipid asymmetry refers to the difference in lipid composition in the outer (exoplasmic, exofacial) leaflet vs. the inner (cytoplasmic, cytofacial) leaflet of a membrane. Many cell membranes possess lipid asymmetry. In mammalian cells, the outer leaflet of the plasma membrane is enriched in sphingomyelin (SM), glycosphingolipids (GSL) and phosphatidylcholine (PC), while the inner leaflet is composed mainly of phosphatidylethanolamine (PE), and anionic lipids such as phosphatidylserine (PS) and phosphatidylinositol (PI) (Fig. 1.1) [1]. Cholesterol is present in both leaflets, but its distribution is still in dispute [2, 3].

Representation of lipid asymmetry in natural biomembranes. In mammalian cells the outer, or exofacial, leaflet is enriched in saturated acyl-chain sphingomyelin and in phosphatidylcholine, while the inner, or cytofacial, leaflet is composed primarily of phosphatidylethanolamine and phosphatidylserine. Cholesterol, shown in gray is present in both leaflets
In cells, lipid asymmetry is maintained by flippases and floppases, enzymes that control the movement of lipids across the bilayer, and by enzymes that synthesize and degrade lipids in one or the other leaflet [4]. Lipid flip-flop, or the transverse diffusion of lipids from one leaflet to another, counteracts asymmetry. The rate of spontaneous phospholipid flip-flop in the absence of proteins is generally slow, and can take days [5,6,7,8,9]. In contrast, cholesterol with its small and weakly polar headgroup, can cross the lipid bilayer in a minute or less [10].
1.2 Biological Function of Asymmetry
The full significance of lipid asymmetry remains elusive, but is known to be important in several biological processes. For example, the loss of PS asymmetry and the resulting display of PS in the outer leaflet of cell membranes is a signal which leads to the consumption of apoptotic cells by phagocytes [11], and is also a signal indicating that the membrane has been damaged, promoting blood clotting [12]. Some viruses even display PS in their outer leaflet to encourage engulfment by macrophages and achieve host infection [13, 14].
Asymmetry may also affect lipid-protein interaction. Transmembrane (TM) proteins typically have a higher positive charge at the cytofacial end of their TM helices relative to their exofacial end. This is known as the positive-inside rule [15]. Lipid charge asymmetry, with a higher negative surface charge at the inner leaflet/cytofacial surface, may help determine TM protein orientation, as well as influence the conformation of positively charged cytofacial juxtamembrane sequences [16]. Importantly, it has recently been observed that plasma membrane TM segments also have an orientational preference as judged from their abundance in natural sequences. The segments of TM helices having amino acids with smaller side chains exhibit a preference to be located in the outer leaflet relative to the inner leaflet [17].
1.3 The Physical State of Membranes: Membrane Domains and How They Might Be Influenced by Asymmetry
Another important aspect of membranes that is likely to be affected by asymmetry is lipid physical state. In a lipid bilayer composed of only phospholipids or sphingolipids there are two common physical states: the gel state and the liquid disordered state (Ld). As the name implies, lipids in the Ld state are disordered and loosely packed. Lipids in the gel state are tightly packed and solid-like, with much less lateral mobility/diffusion than Ld state lipids. In a pure lipid bilayer each individual lipid has its own characteristic melting, or transition temperature (Tm), at which the gel state will reversibly melt to form the Ld state. In lipid bilayers with mixtures of both high Tm lipids and low Tm lipids, gel and Ld states can co-exist in the same bilayer. The addition of cholesterol to gel state lipids generally changes the bilayer’s physical state to what is known as the liquid ordered (Lo) state [18, 19]. Since sphingolipids and other lipids with saturated acyl chains have high Tm values, they readily form the Lo state. The Lo state has lipids that are tightly packed and ordered, as in the gel state, but with fast lateral diffusion, as in the Ld state. By light microscopy it is possible to observe co-existing Lo and Ld lipid phases in giant unilamellar vesicles (GUVs) composed of mixtures of high Tm lipids, low Tm lipids and cholesterol, for example, sphingolipids, unsaturated phospholipids and cholesterol [20, 21], and in giant plasma membrane vesicles (GPMV) derived from eukaryotic plasma membranes, which are rich in sphingolipids and cholesterol [22].
The lipid raft model posits that sphingolipid and cholesterol-rich Lo domains (lipid rafts) co-exist with Ld domains in living cells [18]. Lipid rafts may be an important feature of lipid organization in natural membranes. They have been proposed to play an important role in many cellular processes such as amyloid formation, protein and lipid sorting, cell signal transduction, and pathogen invasion [23,24,25,26,27,28,29,30].
If lipid rafts are an integral component of the machinery for transmitting information through the bilayer from the outside of the cell to the inside, then an interesting question arises: since the inner leaflet of the bilayer contains little to no raft-forming sphingolipids, then how can Lo domains form in the inner leaflet? The answer to this may be that outer leaflet lipids influence the physical properties of the inner leaflet: the physical properties of the two leaflets could be ‘coupled’ [31, 32]. This coupling could transfer information across membranes via the lipids themselves. For example, it is possible that domains are induced in the inner leaflet and these domains could then concentrate cytosolic-anchored proteins which have a high affinity for ordered domains, such as proteins anchored by saturated acyl chains [33, 34]. This could then lead to formation of specific protein-protein interactions (Fig. 1.2). Hints that interleaflet coupling may occur come from studies by the Mayor group which have shown that transmembrane interactions between outer leaflet long acyl-chain lipids and inner leaflet phosphatidylserine are crucial in generating actin dependent clustering of cytofacial lipid-anchored proteins [35]. It should be noted that this would represent a very different mechanism for signal transduction across membranes than that mediated by transmembrane proteins, and might provide a new target for biomedical applications.

Schematic illustration of interleaflet coupling giving rise to lipid-mediated signal transduction across membranes. Lo forming outer leaflet lipids are shown inducing Lo domains in the inner leaflet. This in turn may cause protein clustering in inner leaflet Lo domains which regulates signal transduction
1.4 Symmetric Model Membrane Vesicles
Techniques employing artificial membrane vesicles for the exploration of the principles governing natural membrane structure and function have been well-established. The most commonly used vesicles have lipid symmetry, i.e. the lipid composition in both leaflets is identical, or near-identical. Symmetric model membrane vesicles can be made from a variety of phospholipids and sphingolipids, with or without sterols, and in a variety of sizes. Multi-lamellar vesicles (MLV) can be prepared by adding buffer to a dried lipid film and then agitating. As the name implies MLV are many-layered, and they can be highly variable in size, with 8–15 concentric bilayers enclosed in a vesicle ranging from 0.5 to several microns in diameter [36]. Small unilamellar vesicles (SUV) are a single bilayer and are generally less than 50 nm in diameter. SUVs can be made by the sonication of dried lipids in an aqueous medium, or by dilution from ethanol [37, 38]. Generating SUVs with ethanol injection allows control over the size of SUVs by adjusting the concentration of lipid in ethanol before dilution [37]. Due to their small size, a feature of SUVs is a high level of curvature stress on the lipids and this has been shown to impact lipid packing [39]. Large unilamellar vesicles (LUV), commonly with a diameter of 100–200 nm, are relatively free from the curvature stress of SUVs. They can often be prepared by subjecting MLVs to multiple cycles of freeze-thaw followed by extrusion through membrane filters with pores of desired size, to obtain vesicles with a carefully controlled diameter [40]. Giant unilamellar vesicles (GUV) can approximate the size of living eukaryotic cells, ranging from 10,000 to 50,000 nm in diameter. GUV can be generated by electroformation [41], gentle hydration [42], and a number of oil-in-water techniques [43,44,45,46,47,48,49,50,51]. They are particularly useful for light microscopy studies.
Using NMR, interleaflet coupling was investigated in sonicated, symmetric SM SUV in one early pioneering study [52]. It was found that ion-induced changes in transition (melting) temperatures in outer leaflet could be partly transmitted to inner leaflet, indicating some level of interleaflet coupling, and it was proposed that interleaflet coupling could be a mechanism to transmit information across bilayers. Furthermore, it was found that the SM chain length influenced coupling, which was detected when SM acyl chain length was long. This could reflect some effect of acyl chain interdigitation.
1.5 Asymmetric Model Membranes: Planar Bilayers
Several different methods have been developed for the construction of asymmetric planar supported bilayers. The Langmuir/Blodgett (LB) [53] and Langmuir–Schaeffer (LS) [54] methods involve first depositing a monolayer of lipids onto a silicon substrate, and then adding a second monolayer by dipping the substrate again through an aqueous phase containing second leaflet lipids. Watanabe et al. recently modified supported planar bilayer techniques to develop a system with a capacity for simultaneously generating over 10,000 asymmetric supported lipid bilayers [55].
Fluorescence Interference Contrast Microscopy (FLIC) was used to compare the stability of asymmetry in planar supported bilayers constructed with several techniques [56]. Lipid asymmetry was measured in bilayers with and without cholesterol in Lo and Ld phases. Lipid asymmetry remained stable at 80% for up to 6 h, as judged by fluorescent probe partitioning in supported bilayers generated using the Langmuir-Blodgett/vesicle fusion (LB/VF) method.
Sum frequency vibrational spectroscopy also was used to examine the kinetics and thermodynamics of lipid asymmetry. In cholesterol-free supported planar LB/LS bilayers, asymmetry stability was limited to several hours and was variably dependent on headgroup structure [57], acyl chain length, lateral surface pressure [58], and acyl chain saturation [59]. Later work in LB/LS supported bilayers investigated the effects of sterol structural variation on flip-flop rates [60]. Vibrational spectroscopy revealed that in distearoyl PC (DSPC) bilayers the stability of lipid asymmetry varied considerably with sterol concentration and structure. The lack of stability seen in these studies may be due to the presence of bilayer defects specific to supported membranes [5].
Other efforts have concentrated on preparation of unsupported or cushioned planar bilayers in order to avoid the influence of a supporting substrate upon lipid behavior. Unsupported asymmetric planar bilayers can be made via the Montal-Mueller method [61]. This entails forming two separate lipid monolayers on each side of a moveable Teflon partition containing a small hole. Carefully moving the Teflon partition allows the formation of a bilayer at the hole in the partition where the monolayers make contact. Using asymmetric unsupported bilayers, the Keller group observed that in mixtures of diphytanoyl PC, dipalmitoylPC (DPPC) and cholesterol, Lo domains can form in one leaflet independently of the other and that by adjusting the lipid composition of one leaflet, domain formation in the opposing leaflet could be suppressed [62]. However, the use of hexadecane to prepare the monolayers raises the issue of residual solvent, which if present between the lipid leaflets could influence interleaflet coupling.
Avoiding issues of solvent and support effects by adapting the LB and LS methods, the Tamm and Naumann groups have used asymmetric bilayers cushioned by polymer tethered to lipids. The Tamm group generated a tethered double bilayer system and used single particle tracking to compare the mobility of the transmembrane protein syntaxin-1A in tethered vs. supported bilayers. The work demonstrated that there was no significant difference in syntaxin-1A mobility in tethered vs. supported bilayers [63]. Using the same approach the Naumann group examined the influence of lipid asymmetry on the sequestering and oligomerization behavior of integrins αvβ3 and α5β1 in bilayers [64]. The behavior of these TM proteins in asymmetric bilayers differed significantly from that in symmetric bilayers.
1.6 Asymmetric Lipid Vesicles
Lipid vesicles closely resemble natural biological membranes in that they are unsupported, fully hydrated and the bilayer is unbounded. Because of this, the results of work using lipid vesicles may be most informative for comparison to biological systems. Even more closely resembling natural biomembranes, would be asymmetric vesicles that reflected both the lipid diversity and asymmetry found in the membranes of living cells.
Early work in the development of asymmetric lipid vesicles examined spontaneous fluorescent lipid analog exchange between vesicle populations [65]. It was demonstrated that NBD-PC fluorescent lipid analogs could spontaneously transfer from the outer leaflet of ‘donor’ lipid vesicles to outer leaflet of ‘acceptor’ lipid vesicles. This would result in asymmetric vesicles in the sense that the fluorescent lipid would be restricted to a single leaflet after exchange. In some cases, as much as 50% of fluorescent donor lipids could be transferred to the acceptor vesicles. However, the method was limited in that it required use of fluorescent lipids in which one acyl chain was modified with a somewhat polar group.
1.7 Making Asymmetric Vesicles with Phospholipid Carrier Proteins
In other early work, the Zilversmit group used phospholipid exchange protein isolated from beef heart cytosol to exchange lipids between erythrocyte ghosts and PC/cholesterol liposomes [66]. In doing so they explored the natural lipid asymmetry in red blood cells and confirmed that lipid asymmetry is stable with sphingomyelin (SM) and PC largely limited to the outer leaflet. In studies using phospholipid exchange protein isolated from beef liver, the asymmetric transbilayer distribution of SM, PE, and PC was assessed [67]. The transfer of radiolabeled phospholipids from intact rat erythrocytes to unilamellar vesicles was measured as a function of time. It was found that all of the SM and more than half of the PC were located in the outer leaflet of the erythrocytes. Since nearly no PE was transferred, it appeared to be largely confined to the inner leaflet.
Also using lipid-exchange proteins, the Holloway group prepared asymmetric membranes to examine the depth of insertion of transmembrane protein cytochrome b5 in membranes. They generated model membranes in which only one leaflet contained phospholipids with brominated acyl chains, which quench tryptophan fluorescence. They found that cytochrome b5 orients in the bilayer with the tryptophan in its hydrophobic tail 7 nm from outer leaflet surface [68].
Lipids bound to bovine serum albumin (BSA) were used to generate asymmetry in rat liver endoplasmic reticulum vesicles for the study of lipid flip-flop rates [69]. A series of spin-labeled phospholipids were introduced into the outer leaflets of rat liver ER vesicles by binding them to BSA. To determine lipid flip-flop rates, ESR spectroscopy and kinetics assays were performed. Results showed that in all observed cases, phospholipid flip-flop was fast, with a half time of 20 min at 37 °C, and not dependent on lipid head group type. However, the spin-labelled phospholipids lacked long acyl chains, so the effects of acyl chain length on flip-flop rates could not be gauged.
Holzer et al. used pro-sterol carrier protein (pro-SCP2) to generate vesicles that were asymmetric with regard to negatively charged egg phosphatidylglycerol (PG) [70]. The degree of asymmetry in the vesicles was then measured using free-flow electrophoresis. Membrane curvature was found to be important for lipid transfer efficiency by pro-SCP2. By using both small donor and small acceptor lipid vesicles (50 nm diameter) exchange efficiency was increased by 55% over that achieved with larger vesicles.
1.8 Using pH Gradients to Make Vesicles with Anionic Lipid Asymmetry
The Cullis group induced asymmetry in vesicles containing negatively charged lipids by inducing a pH gradient across the bilayer. Following acidification of the exterior of PG-containing vesicles relative to the luminal cavity, 50% of PG (5% of total outer leaflet lipids) moved to the inner leaflet in within 50 min [71]. Increased acyl chain saturation, chain length and inclusion of cholesterol significantly decreased this effect by decreasing membrane permeability and therefore the ability of PG to move across the bilayer. It was also shown that it was possible to reversibly manipulate the distribution of the negatively charged egg PG and egg phosphatidic acid (PA) in the bilayer [72]. EggPG and eggPA, but not the zwitterionic lipid dioleoylPE (DOPE) could be driven to the inner or outer leaflet, depending on the direction of the proton gradient. Cryoelectron microscopy was used to examine morphological changes in the bilayers of dioleoyl PG (DOPG)-containing LUVs with pH gradient-induced DOPG asymmetry. Flipping of DOPG from the outer to inner to leaflet or vice versa generated inversions and tubular protrusions [73]. This approach is limited to the movement of anionic lipids, and by the small percentages of anionic lipid that could be used (10% of total). Furthermore, the pH gradient necessary to induce movement of anionic lipids would not often be suitable for use in studies of protein conformation or function in asymmetric vesicles.
1.9 Making Asymmetric Vesicles with Water-in-Oil Techniques: Centrifugation Method
The Weitz group developed an oil-in-water technique to generate asymmetric giant unilamellar vesicles (GUV) [74]. In the method, an emulsion of dodecane, inner leaflet lipids and water was first prepared. This was layered over an intermediate phase consisting of dodecane and outer leaflet lipids, which was directly supported on an aqueous phase. As the droplets, which would have inner leaflet lipids in an inverted micelle type of structure, passed via centrifugation through the dodecane/outer leaflet layer into the aqueous phase, a monolayer of outer leaflet lipids was deposited on the outside of the inner leaflet monolayer vesicles, forming asymmetric bilayers. Using this technique, a controllable, high level (95%) of lipid asymmetry was reported.
Studies using vesicles prepared by oil-in-water methods to examine the effect of lipid asymmetry on the mechanical properties of bilayers showed that asymmetry significantly increases membrane rigidity. Two types of fluorescently-labelled asymmetric GUVs: dioleoyl PC (DOPC)in/1-palmitoyl-2-oleoyl PC (POPC)out and POPCin/DOPCout (in = inner leaflet lipid, out = outer leaflet lipid) were generated. Membrane rigidity was calculated by thermal fluctuation analysis of phase contrast micrographs. Results revealed that both types of asymmetric membranes exhibited a significantly higher bending rigidity compared to symmetric membranes consisting of either DOPC, POPC, or 1:1 DOPC:POPC [75].
The Takagi group has developed a method to control the size of water-in-oil asymmetric GUV. By adjusting the density of luminal contents with water-soluble molecules such as sucrose and glucose, the rate of transfer and therefore the size of vesicles could be controlled. Fluorescently labelled lipids were incorporated into the cholesterol-containing asymmetric GUVs and facilitated the observance of microdomain formation and confirmed lipid asymmetry [48].
1.10 Making Asymmetric Vesicles with Water-in-Oil Techniques: Droplet Interface Bilayers
Hwang and colleagues examined the effect of lipid asymmetry on the behavior of membrane proteins in droplet interface bilayers (DIB) [51]. In this method two types of aqueous droplets surrounded by inverted lipid monolayers with different lipids are deposited in an oil mixture. As the droplets meet, and monolayers come into contact, and an asymmetric bilayer forms at their junction. Employing monolayer droplets with differences in lipid charge, bilayers possessing a charge gradient were constructed and used to measure charge gradient-induced changes in the insertion and gating behavior of the outer membrane protein G (OmpG) from Escherichia coli. This method is limited to use with lipids that are soluble in oil, but allows fine control over mixtures of lipids for each monolayer.
1.11 Making Asymmetric Vesicles with Water-in-Oil Techniques: Microfluidic Approaches
The introduction of microfluidic techniques to the production of asymmetric lipid vesicles has enabled methods for the generation of water-in-oil GUV with controllable size, lipid asymmetry, and lumenal content. The Malmstadt group used a microfluidic flow-based layer-by-layer approach to produce asymmetric vesicles with biologically-relevant PS limited to a single leaflet [46]. First sucrose-loaded aqueous droplets were introduced to an oil stream containing dissolved inner leaflet lipids to develop a lipid monolayer. The droplets were then released into a second oil phase containing dissolved outer leaflet lipids floating over an aqueous phase. Following centrifugation, the droplets passed through the aqueous phase and were collected. Lipid asymmetry was confirmed via differential fluorescence quenching and selective labeling with biotinylated lipids. The vesicles were size-selectable because vesicles larger than 120 μm do not survive the centrifugation step and those smaller than 10 μm don’t transfer through the second step. Lipid asymmetry was confirmed with about 85% asymmetry. It was necessary to identify and select oil-free vesicles visually.
The generation of asymmetric vesicles for drug delivery is a promising area of research. In a study of drug cytotoxicity, microfluidics were employed to generate GUVs with a cross-linked, chemically stable inner leaflet composed of trichloro(1H,1H,2H,2H–perfluorooctyl)silane (TPS) and an outer leaflet containing DOPC. When loaded with the cancer therapeutic 5-fluorouacil, the asymmetric vesicles exhibited a pH-dependent threefold higher cytotoxicity relative to the free drug [76].
A high-throughput microfluidic process for generating water-in-oil GUV was recently developed [47, 77]. In the method, water, oil and inner leaflet emulsions were first formed. Outer leaflet lipids were then introduced via replacement of inner leaflet lipid solution with a second solution. Water-in-oil double emulsions were then formed and followed by the extraction of excess oil. Asymmetry and unilamellarity were confirmed with fluorescence quenching assays and transmembrane protein insertion assays. In a second report using this same fabrication strategy, the effects of lipid asymmetry on the mechanical properties of asymmetric bilayers vs. symmetric bilayers were examined. In cholesterol-free symmetric vesicles composed of dimyristoyl PC (DMPC), DOPC a 1:1 mixture of DMPC:DOPC, or asymmetric vesicles with a DMPCin/DOPCout or DOPCin/DMPCout composition results showed that the bending moduli and expansion moduli of asymmetric bilayers are different that those of symmetric bilayers.
1.12 Minimizing Residual Oil Contamination
A complication resulting from oil-in-water techniques used to generate asymmetric GUV is residual oil contamination. This residual oil likely impacts the physical properties and behavior of the bilayer. To circumvent this, methods to minimize oil contamination have been developed.
A microfluidics approach in which lipid monolayers were brought together in a fashion similar to that in droplet interface bilayers was employed to generate asymmetric GUV likely to have only minimal oil and then used to examine the effects of lipid composition and asymmetry on lipid vesicle fusion mediated by SNARE-family transmembrane proteins [45].
In another study, a high-throughput microfluidics method was developed to form asymmetric GUV with minimal oil contamination. In the study, cholesterol- rich asymmetric GUVs with ‘ultrathin shell’ (residual oil) were generated for the observation of lipid microdomain formation. Lipid asymmetry and the presence of lipid domains were observed with the use of fluorescently labelled lipids and fluorescent microscopy [44]. In another attempt to address the issues of solvent contamination, an asymmetric planar lipid bilayer was formed into lipid tubules which were then broken up by applying a microfluidic jet flow of aqueous buffer. Unilamellarity and residual solvent content was evaluated using confocal Raman scattering microscopy [78]. Although these methods may greatly reduce oil contamination, it is not clear whether the residual oil that remains would not perturb lipid behavior to a significant degree.
1.13 Early Studies Using Cyclodextrin Exchange to Alter Lipid Composition
A different strategy that is becoming more widely used is to prepare asymmetric lipid vesicles with cyclodextrins (CDs). CDs are water-soluble cyclic oligosaccharides composed of glucose monomers linked together by α-(1, 4) glycosidic bonds, and are typically composed of 6(α), 7(β), or 8(γ) units. They assume a conical barrel shape with a non-polar cavity and hydrophilic exterior. Hydrophobic guest molecules can complex with CDs. A wide range of functional groups can be conjugated to hydroxyl groups at the edges of CDs to modify their binding specificity [79, 80] (see Fig. 1.3). Changes the number of monomers comprising a CD alters the volume of the hydrophobic cavity, which can be varied to exclude or include larger or smaller guest molecules. For example, the hydrophobic cavity of αCDs is smaller than that of βCDs and therefore binds well to phospholipid acyl chains, but binds cholesterol either not at all or very poorly [80,81,82,83].

α-cyclodextrin chemical structure. The 6-unit 1–4 linked α-D-glucopyranoside ring forms a barrel shape, with a hydrophobic interior cavity and a hydrophilic exterior surface. Functional groups can be conjugated to hydroxyl groups to modify their binding specificity. R = OH or modified OH
Cyclodextrins have long been used to remove sterols from cells, introduce new sterols into cells, or exchange one sterol in cells for another [84,85,86,87,88]. Similarly, CDs have been used to exchange sterols between model membrane vesicles [10, 89].
Early work exploring the properties of CDs as lipid carriers involved interactions of γ-CDs and fluorescent lipids. Carboxyethyl-γ-CD accelerated fluorescent lipid transfer from lipid vesicles to cells in culture [90]. Another study reported that the large hydrophobic cavity of γ-CD could bind more than one lipid acyl chain simultaneously [91]. In addition, stoichiometric measurements using fluorescent binding assays showed that the association constants of pyrene-labeled PC/γ-CD complexes decreased strongly with increasing acyl chain length [91].
Other studies demonstrated that MβCD, among other cyclodextrins, could exchange phospholipids between artificial lipid vesicles, and at high concentrations even dissolve phospholipid vesicles [10, 92].
It was also shown that cyclodextrins can also cause the dissociation of small amounts of phospholipids from cells [86, 93].
1.14 Preparation and Properties of Asymmetric Vesicles Produced by MβCD-Catalyzed Exchange
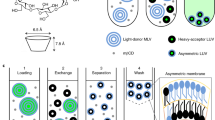
The first study making use of CD to prepare highly asymmetric vesicles (AUV) employed MβCD to exchange lipids between the outer leaflets of two distinct vesicle populations [94]. The mixing of ‘donor lipid’-loaded MβCD solutions (with excess donor lipid vesicles) and symmetric ‘acceptor’ SUV composed solely of the desired inner leaflet lipids was the key feature of the technique (Fig. 1.4). Two lipid exchanges were carried out: The first exchange catalyzed the installation of ‘outer leaflet’ lipids into acceptor SUV – effecting phospholipid asymmetry. Using this approach asymmetric SUV were prepared containing 75–100% SM in their outer leaflet, and DOPC, POPC, 1-palmitoyl-2-oleoyl PS (POPS) or a mixture of 1-palmitoyl-2-oleoyl PE (POPE) and POPS in their inner leaflet. A second, optional exchange installed cholesterol to a final overall cholesterol concentration of approximately 25 mol%. The resulting asymmetric vesicles were purified from donor lipid mixtures by size chromatography. Fluorescence assays measured outer leaflet anisotropy and outer leaflet lipid charge and confirmed lipid asymmetry.

A Scheme for Cyclodextrin-mediated Asymmetric Vesicle Preparation. Outer leaflet donor lipids in complex with cyclodextrin in solution (and some excess donor lipid vesicles) are incubated with sucrose-loaded inner-leaflet acceptor vesicles. The mixture is layered over sucrose at a lower concentration than that inside of the vesicles and ultracentrifuged. The asymmetric vesicle pellets and can be isolated from the other exchange reaction components. Notice that the resulting asymmetric vesicles will have an osmotic imbalance across the bilayer unless the osmotic strength of the external solution is adjusted after the vesicles have been isolated
The thermal stability of ordered state SM domains in the outer leaflet of resulting AUV was found to be much higher than that in symmetric vesicles with the same overall composition, with a Tm approaching that of pure SM vesicles. This suggested that at the Tm of the outer leaflet, coupling to the inner leaflet composed of unsaturated phospholipids had broken down. Analogous properties were observed in asymmetric SUV with cholesterol (vs. corresponding symmetric SUV with the same lipid composition), except that the addition of cholesterol further thermally stabilized ordered domains in both symmetric and asymmetric vesicles. Experiments monitoring the orientation of hydrophobic helices suggested that membrane asymmetry could enhance formation of a transmembrane orientation within the bilayer.
A subsequent study developed a modified method to prepare plasma membrane-mimicking asymmetric LUV [95]. The lesser bilayer curvature of LUVs relieves the lipid packing stresses that exist in SUV. To do this, acceptor LUV were prepared with entrapped sucrose, and then isolated from donor lipids and MβCD after exchange by centrifugation. As with asymmetric SUV, nearly complete replacement of the outer leaflet lipid was achieved. Anisotropy assays revealed that the SM rich outer leaflet of asymmetric LUV melted at about the same temperature as symmetric LUV composed of pure SM, and at a much higher Tm than that of symmetric vesicles with the same overall lipid composition. This indicated that the weak interleaflet coupling at the temperature of outer leaflet melting seen in the prior study was not a result of the high membrane curvature in SUV.
MβCD was next used to prepare asymmetric GUVs for studies of interleaflet coupling with fluorescence correlation spectroscopy (FCS) [96]. In order to prevent the possibility that donor vesicles would stick to the GUV, residual donor vesicles were removed before mixing the donor lipid-MβCD mixture with acceptor GUV. This resulted in a lower, but still substantial, level of lipid exchange in the outer leaflet. Controls with fluorescent lipids demonstrated that stable lipid asymmetry was achieved in osmotically balanced vesicles. Osmotic balance across the bilayer is crucial because even a small imbalance has been shown to induce transient pore formation in GUV, which could potentially lead to a loss of asymmetry [97].
The introduction of SM into the outer leaflet of cholesterol-free DOPC GUV decreased lateral diffusion in the outer leaflet. Interleaflet coupling at room temperature was evaluated from the extent to which it also decreased lateral diffusion in the inner leaflet and was found to be stronger with inner leaflet brain PC (brain PC) than with DOPC.
In a follow-up study, the effect of varying acyl chain length on lateral lipid diffusion and membrane order in asymmetric GUVs and SUVs was further investigated [98]. As previously shown, MβCD-generated asymmetric GUV containing outer leaflet bSM and inner leaflet DOPC showed reduced lateral diffusion in the bSM outer leaflet, and little reduction in diffusion in the DOPC inner leaflet. However, when POPC or 1-oleoyl 2-myristoyl PC (OMPC) (both of which have one saturated and one unsaturated acyl chain) were used in place of DOPC, there was reduced lateral diffusion in both leaflets, with a similar level of reduction of diffusion in both leaflets, indicative of strong interleaflet coupling. Strong interleaflet coupling was further observed with the introduction of longer acyl chain milk SM or synthetic C24:0 SM into the outer leaflet opposing inner leaflet DOPC. This suggested that acyl chain length plays a role in interleaflet coupling. Interestingly, assays measuring inner leaflet lipid order in asymmetric SUV and GUV did not detect an increase in inner leaflet order when SM was introduced into the outer leaflet. For the SUV experiments, a recently developed method to selectively label inner lipids was used [99]. In this method inner leaflets are selectively labelled with fluorescent probes using low levels of MβCD and gel-filtration to remove outer leaflet fluorescent probes.
The difference in coupling with regard to diffusion and order is somewhat puzzling. One possible explanation is that order measurements are the average for an entire leaflet. It is possible that upon introduction of SM into the outer leaflet, an increase in order in inner leaflet ordered domains coupled to outer leaflet ordered domains rich in SM is “cancelled out” by a decrease in order in the remainder of the inner leaflet, which would be in contact with outer leaflet disordered domains rich in DOPC.
1.15 Studies of Lipid Flip-Flop in Asymmetric Vesicles
Next, a systematic study using MβCD exchange was undertaken to examine the effect of acyl chain structure upon the stability of asymmetry in SMout/PCin SUV in which the type of PC used was varied [9]. The level of outer leaflet exchange was quantified by comparing outer leaflet membrane order (assessed via fluorescence anisotropy of TMADPH added to the outer leaflet, TMADPH does not rapidly flip across the lipid bilayer) to that for a standard curve composed of symmetric bSM/PC vesicles with various % SM. Thin layer chromatography was used to measure overall lipid composition. Combining these two methods, the % of inner leaflet bSM, and thus the level of asymmetry, was calculated. In all cases exchange levels indicated, at a minimum, complete replacement of the outer leaflet PC with SM. For AUV containing PC with one saturated and one unsaturated acyl chain, asymmetry was virtually complete if the unsaturated chain had one or two double bonds. However, a small loss of loss of asymmetry was detected when the unsaturated acyl chain in the PC had four double bonds. AUV containing PCs with two monounsaturated acyl chains maintained full asymmetry for all except those with the shortest acyl chain studied (14 carbons), which again showed a small loss of asymmetry. In contrast, exchange-produced vesicles containing PCs with two polyunsaturated chains lost most (with two double bonds per chain) or all (with three or more double bonds per chain) of any asymmetry by the time asymmetry measurements were made, several hours after lipid exchange.
Measurement of transverse diffusion (flip-flop) rates for NBD-lipids in symmetric PC and SM/PC vesicles confirmed that the loss of asymmetry was associated with increased lipid flip-flop, and gave flip-flop values consistent with the literature for the effect of PC acyl chain structure upon lipid flip [100]. Acyl chain polyunsaturation may promote lipid flip-flop due to its effect of decreasing how tightly lipids are packed, and/or due to the increased polarity of the core of the bilayer, as C-C double bonds have increased polarity relative to single bonds [101]. Increased polarity would increase the solubility of lipid polar headgroups within the bilayer core.
A systematic study was also undertaken of the effects of phospholipid polar head group structure on the stability of bilayer asymmetry [8]. MβCD was used to generate asymmetric SUV with outer leaflet bSM and inner leaflet 1-palmitoyl-2-oleoyl-PG (POPG), DOPG, soy PI, tetraoleoylcardiolipin (tetraoleoyl CL), or POPS, in all cases alone or in combination with POPE. Inner leaflet dioleoyl PA (DOPA) was studied in a mixture with POPC. Proton NMR spectroscopy and observed changes in the thermal stability of SM-rich ordered domains helped detect an asymmetric distribution of SM and its decay over time. AUV containing a mixture of PE and anionic lipids maintained nearly full asymmetry that was stable for more than 1 day. However, most exchange vesicles containing anionic lipids without PE only showed partial asymmetry (which was assayed within hours after they were prepared) that decayed further after 1 day of storage. Asymmetry also decayed in 1 day for the vesicles containing PA. Interestingly, exchange vesicles with PS showed almost full asymmetry that was stable over 1 day even in when PE was not present in the inner leaflet. It is possible that lipids with a single charged anionic group undergo transverse diffusion across membranes more easily than lipids combined with multiple charge groups, such as PC, SM, PE and PS.
In a study using a modification of the lipid-exchange method to prepare asymmetric LUV without trapped sucrose (see below), 1H NMR was used to measure the flip-flop rates as a function of temperature using asymmetric vesicles in which chain-perdeuterated dipalmitoyl PC (DPPC-dC) was introduced into the outer leaflet of acceptor LUVs initially composed of headgroup-deuterated acceptor lipid (DPPC-dH) [5]. Flip-flop in gel phase bilayers was undetectable, while in the Ld state, from 50–65 °C, flip-flop half-time rates ranged between days to weeks. Intriguingly, flip-flop in asymmetric vesicles incubated at the main transition temperature (40 °C) for DPPC, was twice as rapid as in fully melted DPPC at 50 °C, indicative of defect-accelerated flip-flop in vesicles when gel and Ld phases co-exist.
1.16 Use of HPαCD to Prepare Asymmetric Vesicles Mimicking Plasma Membranes
Because MβCD binds well to cholesterol, it is difficult to control the levels of cholesterol in AUV during MβCD-mediated phospholipid exchange. To circumvent this, hydroxypropyl-α-cyclodextrin (HPαCD) was used instead. The hydrophobic cavity of the six glucose unit ring of HPαCD is too small to bind cholesterol, but can bind to the acyl chains of phospholipids. In a study comparing MβCD, HPαCD and HPβCD [81], it was found that while only MβCD could dissolve lipid vesicles, all three cyclodextrins could catalyze lipid exchange between vesicles.
HPαCD was used to construct plasma membrane-mimicking asymmetric LUVs with SM and/or POPC in the outer leaflet, POPE and POPS in the inner leaflet, and a wide range of cholesterol concentrations [82]. Efficient exchange (as high as 80–100% replacement of the outer leaflet as judged by thin layer chromatography) could be achieved, especially at high levels of cholesterol. Exchange of POPC donor lipid was somewhat more efficient than that of SM. Asymmetry was confirmed by chemical labelling of externally exposed POPE with TNBS, and by using a cationic peptide binding assay to measure the amount of externally-exposed anionic lipid. Asymmetry was stable for days.
1.17 Domain Formation in Asymmetric GUV Containing Cholesterol
HPαCD-catalyzed lipid exchange was used to prepare and investigate domain forming properties of asymmetric GUV in a following study [102]. Cholesterol-containing GUV with a mixture of SM and DOPC in their outer leaflets and DOPC in their inner leaflet were studied. Surprisingly, SM (and presumably cholesterol)-rich liquid ordered domains in the outer leaflet induced ordered domains in corresponding region of the inner leaflet, despite the strong tendency of DOPC, which has two unsaturated oleoyl chains, to remain in a relatively disordered state, even when mixed with cholesterol [103]. It was proposed that a lateral rearrangement of the inner leaflet was occurring, in which the regions of the inner leaflet in contact with the outer leaflet became enriched in cholesterol and depleted in DOPC. Supporting this, the NBD-DOPE used to probe inner leaflet domain formation, which like DOPC has two unsaturated oleoyl chains, was also depleted in the inner leaflet ordered domains in contact with outer leaflet ordered domains.
The contribution of SM acyl chain interdigitation to interleaflet coupling in the asymmetric GUV was investigated by comparing coupling when the outer leaflet contained egg SM, (predominately having C 16:0 acyl chains) vs. milk SM, which is rich in C 22–24 acyl chains that can interdigitate into the inner leaflet. With both types of SM, inner leaflet ordered domains formed. However, only when milk SM was used did inner leaflet ordered domains show probe partitioning properties similar to those in the outer leaflet, indicating stronger interleaflet coupling. This is consistent with the above-noted coupling seen in GUV lacking cholesterol.
1.18 Studies Using Cyclodextrin-Catalyzed Lipid Exchange to Study the Effect of Phospholipid Composition and Asymmetry Upon Membrane Protein Behavior
Transmembrane protein behavior can be influenced by lipid asymmetry in SUV. An asymmetric distribution of anionic lipids favored formation of a transmembrane configuration relative to a non-transmembrane (but membrane bound) state in which the peptide inserted only shallowly within the lipid bilayer [94].
The conformational behavior of the pore-forming toxin protein perfringolysin (PFO) was studied in AUV prepared using HPαCD to determine if lipid asymmetry could influence the conformation of this full length protein when it is membrane-inserted [21]. In a cholesterol concentration-dependent manner, PFO forms a large, homo-oligomeric β-barrel pore complex that spans the membrane bilayer of mammalian cells. PFO behavior was evaluated in asymmetric LUVs with a POPC outer leaflet, POPE-POPS inner leaflet and varying concentrations of cholesterol. Results were compared to those for PFO mixed with a variety of symmetric vesicles. This included symmetric vesicles with an overall lipid composition identical to that in the AUV, those containing only outer leaflet AUV lipids, and those composed of only the inner leaflet AUV lipids.
PFO binding and insertion were assessed by monitoring intrinsic Trp emission and the fluorescence intensity of an acrylodan-labeled residue. PFO oligomerization was measured with FRET and with SDS-agarose gel electrophoresis, as PFO oligomers do not unfold in SDS once the beta-barrel has formed. Pore-forming capacity was assayed by measuring the increase in externally-added BODIPY fluorescence emission intensity when BODIPY-labeled streptavidin was added to AUV with trapped biocytin that is released upon pore formation.
These assays demonstrated that the dependence of PFO conformation as a function of cholesterol concentration in AUV was different from that in symmetric vesicles of the same overall composition, and also different from that in symmetric vesicles that mimicked the outer or inner leaflets of the AUV. Interestingly, in the AUV a non-pore-forming, but transmembrane embedded, oligomeric conformation of PFO was observed at intermediate cholesterol concentrations. This may represent an intermediate stage in the PFO pore assembly process.
Cyclodextrin-mediated lipid exchange has also been used to investigate the effect of in situ changes in lipid composition upon the topography of lactose permease (LacY) reconstituted into model membrane vesicles. Following MβCD-induced enrichment of outer leaflet PE in proteoliposomes containing LacY in a partly inverted transmembrane conformation, LacY very quickly flipped to its native transmembrane conformation. Conversely, MβCD-mediated exchange introducing PG or CL into PE vesicles induced the flipping of LacY to form the partly inverted transmembrane orientation. The asymmetry of the lipids after the lipid exchange steps was not investigated [104, 105].
Membrane domain localization studies of nicotinic acetylcholine receptor (AChR) reconstituted in proteoliposomes found that localization of AChR within ordered lipid domains can depend upon lipid asymmetry [106]. Using both symmetric and asymmetric model systems containing cholesterol and different SM species, localization of AChR was assessed by FRET and by quantification of the distribution of AChR in detergent resistant and detergent soluble membrane fractions. AChR was reconstituted in symmetric vesicles composed of 1:1 POPC:Chol, or 1:1:1 POPC:Chol:SM. To prepare asymmetric vesicles, SM was exchanged into the outer leaflet of proteoliposomes containing AChR and POPC and cholesterol. In the case of 16:0 SM or 18:0 SM-containing symmetric vesicles, AChR was found to localize in Lo domains. No such Lo preference was found in symmetric vesicles containing bSM or 24:1 SM. However, in asymmetric models with bSM enrichment in the outer leaflet, AChR showed a strong preference for Lo domains.
1.19 Use of MαCD to Prepare Asymmetric Lipid Vesicles
Preparation of asymmetric LUV using MαCD was achieved very recently [as a portion of a study achieving lipid exchange in cells using MαCD, see below] [107]. Since MαCD does not interact well with cholesterol, this again allowed exchange of phospholipids without disturbing cholesterol levels in the acceptor vesicles from which the asymmetric LUV were prepared. MαCD was able to solubilize lipids at an even lower concentration than MβCD, indicating a strong interaction with lipids. In addition, like MβCD, MαCD had a relatively low specificity for lipid structure as judged by the concentration of MαCD needed to solubilize vesicles composed of various lipids. This made it possible to carry out exchange with various lipids at relatively low MαCD concentrations. Efficient exchange was obtained, but in contrast with HPαCD exchange was a bit more efficient for SM than POPC. Most importantly, the combination of strong phospholipid and sphingolipid interactions, coupled with negligible interactions with cholesterol, enabled applications of MαCD-induced lipid exchange to living cells, as described below.
1.20 Studies Using Cyclodextrin-Catalyzed Lipid Exchange to Prepare Asymmetric Planar Bilayers
Cyclodextrin-mediated exchange has also been extended to preparation of asymmetric planar lipid bilayers [108]. Fluorescence correlation spectroscopy was used to measure lipid mobility in each leaflet independently in asymmetric supported bilayers (aSLB). The method was compatible with protein reconstitution. DOPC/GPI-anchored placental alkaline phosphatase (PLAP) proteoliposomes were fluorescently-labelled and applied via a vesicle fusion method to a mica support. Leaflet-specific labeling of the supported leaflet was achieved by destroying NBD-DOPE fluorescence in the solution-exposed outer leaflet using sodium dithionite. Lipid asymmetry was induced using successive incubations with MβCD-loaded bSM. Cholesterol was introduced in additional exchange steps. Asymmetry was stable for several hours. Interestingly, no large scale lipid phase separation was observed in the aSLBs until asymmetry began to break down and bSM was present in both leaflets. Any domains present in the fully asymmetric aSLB were submicroscopic.
1.21 Modifying Cyclodextrin-Catalyzed Lipid Exchange for Study of Lipid Packing Using Small Angle Neutron Scattering
MβCD-catalyzed exchange has also been adapted for small-angle neutron scattering (SANS) studies [109]. This required modifying the protocol for MβCD-mediated exchange for LUV, in which trapping sucrose within the lumen of the vesicle aids AUV isolation, as the trapped sucrose interferes with SANS measurements. To avoid this, sucrose-loaded donor lipid vesicles preincubated with MβCD were mixed with sucrose-free acceptor vesicles. Following lipid exchange, the sucrose-laden donor vesicles were removed from the sucrose-free asymmetric LUV by centrifugation. Cyclodextrin with any bound lipid was then removed by filtration and washing. This allowed generation of asymmetric LUV in which one leaflet contained isotopically (deuterated)-labelled lipids (including fully labeled, or labeling the only the polar headgroup or acyl chains). Unlabeled POPC or DPPC were introduced into the outer leaflet of LUV containing partly or fully deuterated POPC. Similarly, deuterated POPC could be introduced into the outer leaflet of unlabeled POPC LUV. Lipid exchange levels and the degree of lipid asymmetry were assessed with 1H NMR and gas chromatography. Changes in bilayer thickness were assessed with SANS. It was found that DPPC-rich ordered (presumably gel) domains directly opposing the POPC leaflet exhibited reduced lipid packing density compared to typical gel phase lipids, indicating that an inner leaflet composed of Ld favoring lipids can partially fluidize outer leaflet ordered domains. This is indicative of a significant degree of interleaflet coupling.
1.22 Tuning Lipid Asymmetry to Control Phagocytosis of Model Membrane Vesicles
Liposomes can be easily generated that mimic apoptotic cells by containing PS in their outer leaflet. However, their utility in drug delivery is limited due to the speed at which they are detected by macrophages and phagocytosed. It would be useful to generate liposomes with encapsulated therapeutic agents that are protected from detection by the immune system until they have been able to bind their targets. In a recent study aiming to achieve this, engineered antibody-decorated asymmetric liposomes were developed to target pathogens to the immune system and to contain a built in time-delay switch dependent upon the flip-flop rates of PS [110].
Symmetric LUVs containing 1:1 DOPE:POPS and were incubated with MβCD pre-loaded with 1:2 bSM:DOPE (both donor and acceptor lipids contained a small amount of streptavidin-capped PE). Following outer leaflet lipid exchange, the resulting asymmetric vesicles were incubated with polyclonal anti-HIV-1 gp120 that was biotin-conjugated. These vesicles were then capable of binding HIV-1 virus-like particles. The slow flipping of PS to the outer leaflet of the vesicles over a period of 24–48 h then allowed the vesicles to then be targeted for destruction by macrophages. Varying the amount of bSM controlled the time delay before phagocytosis, with the highest outer-leaflet bSM content behaving similarly to LUVs without PS.
1.23 Extending Cyclodextrin-Induced Phospholipid Exchange to Mammalian Cells
Our understanding of the function of lipids in cellular membranes has fallen far behind that of other cellular molecules such as proteins or nucleic acids. This is largely due to the difficulty of manipulating lipid composition in living cells. It has been shown that membrane fatty acid composition, phospholipid composition, and cholesterol content can be modified in many different kinds of cultured mammalian cells by adding exogenous lipids in the form of liposomes or using lipids bound to carrier proteins. This topic has been comprehensively reviewed [111]. Although these methods alter membrane lipid composition, they only can make limited changes, and generally add lipids to cells, rather than substitute endogenous with exogenous lipids. Manipulating lipid composition using synthesis inhibitors can replace the original lipid composition of a cell with an altered one. However, inhibitor molecules act slowly. Furthermore, it is only possible to inhibit the synthesis of a subset of membrane lipids, most commonly sphingolipids or sterols [112, 113]. Mutations can be used to alter sterol structure [114, 115] and using a combination of mutations and introduction of new enzymes for wholesale metabolically re-engineering of lipid content can alter phospholipid composition drastically, but is difficult. To date phospholipid reengineering has been carried out successfully in E. coli [116, 117].
An alternative strategy is to use cyclodextrins to manipulate cell lipids in vivo. As noted above, MβCD has been widely used to manipulate plasma membrane sterol content in cultured cells. This can involve extracting cholesterol using empty MβCD, or loading cells with cholesterol by incubating them with MβCD/cholesterol complexes, or using MβCD/sterol complexes to substitute cholesterol in cells with another sterol [84, 85].
Use of cyclodextrins to alter other cellular lipids has been much less studied. Early studies concentrating upon fluorescently labeled lipids were noted above. In more recent work, a variety of unlabeled phospholipids was introduced into cells using MβCD [118]. One limitation of the approach was that the level of exogenous lipid introduced into cells was dependent upon acyl chain structure. Phospholipids with short 14 carbon acyl chains were mostly highly transferred into cells. It was unclear whether the introduction of lipids involved lipid exchange, because there was a net increase in cellular phospholipid in the transfer process. It was also found that additional cholesterol could to be introduced to compensate for MβCD-induced cholesterol depletion in cells. Interestingly, introduction of exogenous PE and PS with unnatural acyl chains to BHK21 hamster kidney cells led to rapid acyl chain remodeling, with substantial remodeling within 1 h, but remodeling was much more limited and slow when acyl chains matched or were close to those in endogenous lipids [119].
Recently, our group developed a method using MαCD to efficiently replace the plasma membrane outer leaflet phospholipids and sphingolipids in cells with exogenous phospholipids and sphingolipids, including unnatural lipids [107]. One advantage of MαCD relative to MβCD, is its small cavity, which is too small to interact with sterols. As a result, MαCD will not significantly alter cholesterol levels during phospholipid/sphingolipid exchange. To carry out exchange, donor lipids (SM or PC or a mixture of the two) in the form of multilamellar vesicles were incubated with a concentration of MαCD high enough to dissolve at least most of the vesicles at the lipid concentrations used (40 mM MαCD was sufficient for use with 1.5–3 mM lipid in most experiments). The mixture was then incubated with cultured human cells, generally at 37 °C for 1 h. This protocol extracted 70–80% of endogenous SM but only very low (10–15%) amounts of PC and PE from the cultured cells. There was minimal PS, PI, or cholesterol extracted from the cells. Similar results were obtained when an exchange was carried out with an unnatural C17:0 SM as the donor lipid, and the lipid composition in the cells before and after exchange was evaluated by mass spectrometry. Exchange was rapid; under the experimental conditions half-time at 37 °C was 15–20 min both for the removal of endogenous SM and introduction of exogenous fluorescently labeled lipid. The maximal SM exchange level was not affected when MαCD concentration, exogenous lipid concentrations, exogenous lipid compositions or cell types were varied. Given these observations and the fact that it has been reported that a significant fraction of SM is not localized in the plasma membrane [119] suggests almost complete replacement of plasma membrane outer leaflet sphingolipids was achieved. Efficient replacement of outer leaflet lipids was also consistent with the observation that up to 90% of cholera toxin B binding to cells was abolished after MβCD-induced lipid exchange, indicative of removal of the cholera toxin receptor glycosphingolipid GM1 from the plasma membrane outer leaflet upon exchange. In addition, carrying out exchange between model membrane vesicles, it was found that MαCD catalyzed exchange of different phospholipid or sphingolipids was relatively non-specific. Thus, it would be expected that all of the outer leaflet lipids (other than cholesterol) would be replaced upon exchange. This was confirmed by lipid analysis of the radiolabeled lipid removed from the cells upon exchange, which had a composition (SM ∼ PC > PE, and little or no PS + PI) closely matching that previously reported for the outer leaflet of human red blood cells [1].
Mass spectrometry analysis of the difference in lipid composition before and after lipid exchange indicated that plasma membrane outer leaflet was enriched in a subset of PC species that are not highly unsaturated. It also showed that shorter acyl chain SM species were enriched in the plasma membrane outer leaflet, in agreement with prior studies [120].
All these results demonstrate that MαCD could be a useful tool to manipulate cell membrane lipid composition in an efficient manner. This may have many applications in addition to studies of lipid asymmetry. For example, the effect of altering plasma membrane outer leaflet lipid composition upon membrane protein function could be studied with this approach. In addition, the ability to introduce large amounts of unnatural phospholipids and/or sphingolipids into cells could aid analysis of lipid structure/function relationships.
References
Verkleij A, Zwaal R, Roelofsen B, Comfurius P, Kastelijn D, Van Deenen L (1973) The asymmetric distribution of phospholipids in the human red cell membrane. A combined study using phospholipases and freeze-etch electron microscopy. Biochim Biophys Acta Biomembr 323:178–193
Mondal M, Mesmin B, Mukherjee S, Maxfield FR (2009) Sterols are mainly in the cytoplasmic leaflet of the plasma membrane and the endocytic recycling compartment in CHO cells. Mol Biol Cell 20:581–588
Liu SL, Sheng R, Jung JH, Wang L, Stec E, O’Connor MJ, Song S, Bikkavilli RK, Winn RA, Lee D, Baek K, Ueda K, Levitan I, Kim KP, Cho W (2017) Orthogonal lipid sensors identify transbilayer asymmetry of plasma membrane cholesterol. Nat Chem Biol 13:268–274
Clark MR (2011) Flippin’ lipids. Nat Immunol 12:373–375
Marquardt D, Heberle FA, Miti T, Eicher B, London E, Katsaras J, Pabst G (2017) 1H NMR shows slow phospholipid Flip-flop in gel and fluid bilayers. Langmuir 33:3731–3741
McConnell HM, Kornberg RD (1971) Inside-outside transitions of phospholipids in vesicle membranes. Biochemistry 10:1111–1120
Nakano M, Fukuda M, Kudo T, Matsuzaki N, Azuma T, Sekine K, Endo H, Handa T (2009) Flip-flop of phospholipids in vesicles: kinetic analysis with time-resolved small-angle neutron scattering. J Phys Chem B 113:6745–6748
Son M, London E (2013) The dependence of lipid asymmetry upon polar headgroup structure. J Lipid Res 54:3385–3393
Son M, London E (2013) The dependence of lipid asymmetry upon phosphatidylcholine acyl chain structure. J Lipid Res 54:223–231
Leventis R, Silvius JR (2001) Use of cyclodextrins to monitor transbilayer movement and differential lipid affinities of cholesterol. Biophys J 81:2257–2267
Li MO, Sarkisian MR, Mehal WZ, Rakic P, Flavell RA (2003) Phosphatidylserine receptor is required for clearance of apoptotic cells. Science 302:1560–1563
Lentz BR (2003) Exposure of platelet membrane phosphatidylserine regulates blood coagulation. Prog Lipid Res 42:423–438
Morizono K, Chen IS (2014) Role of phosphatidylserine receptors in enveloped virus infection. J Virol 88:4275–4290
Mercer J, Helenius A (2008) Vaccinia virus uses macropinocytosis and apoptotic mimicry to enter host cells. Science 320:531–535
von Heijne G (1992) Membrane protein structure prediction. Hydrophobicity analysis and the positive-inside rule. J Mol Biol 225:487–494
Slusky JS, Dunbrack RL Jr (2013) Charge asymmetry in the proteins of the outer membrane. Bioinformatics 29:2122–2128
Sharpe HJ, Stevens TJ, Munro S (2010) A comprehensive comparison of transmembrane domains reveals organelle-specific properties. Cell 142:158–169
Brown D, London E (1998) Functions of lipid rafts in biological membranes. Annu Rev Cell Dev Biol 14:111–136
Brown D, London E (1998) Structure and origin of ordered lipid domains in biological membranes. J Membr Biol 164:103–114
Korlach J, Schwille P, Webb WW, Feigenson GW (1999) Characterization of lipid bilayer phases by confocal microscopy and fluorescence correlation spectroscopy. Proc Natl Acad Sci 96:8461–8466
Lin Q, London E (2014) The influence of natural lipid asymmetry upon the conformation of a membrane-inserted protein (Perfringolysin O). J Biol Chem 289:5467–5478
Levental I, Grzybek M, Simons K (2011) Raft domains of variable properties and compositions in plasma membrane vesicles. Proc of Natl Acad Sci USA 108:11411–11416
Taylor DR, Hooper NM (2007) Role of lipid rafts in the processing of the pathogenic prion and Alzheimer’s amyloid-beta proteins. Semin Cell Dev Biol 18:638–648
Williamson R, Usardi A, Hanger DP, Anderton BH (2008) Membrane-bound beta-amyloid oligomers are recruited into lipid rafts by a fyn-dependent mechanism. FASEB J 22:1552–1559
Cuadras MA, Greenberg HB (2003) Rotavirus infectious particles use lipid rafts during replication for transport to the cell surface in vitro and in vivo. Virology 313:308–321
Gulbins E, Kolesnick R (2003) Raft ceramide in molecular medicine. Oncogene 22:7070–7077
Lyman MG, Curanovic D, Enquist LW (2008) Targeting of pseudorabies virus structural proteins to axons requires association of the viral Us9 protein with lipid rafts. PLoS Pathog 4:e1000065
Murphy SC, Hiller NL, Harrison T, Lomasney JW, Mohandas N, Haldar K (2006) Lipid rafts and malaria parasite infection of erythrocytes. Mol Membr Biol 23:81–88
Riethmuller J, Riehle A, Grassme H, Gulbins E (2006) Membrane rafts in host-pathogen interactions. Biochim Biophys Acta 1758:2139–2147
Korade Z, Kenworthy AK (2008) Lipid rafts, cholesterol, and the brain. Neuropharmacology 55:1265–1273
Collins MD (2008) Interleaflet coupling mechanisms in bilayers of lipids and cholesterol. Biophys J 94:L32–L34
Kiessling V, Wan C, Tamm LK (2009) Domain coupling in asymmetric lipid bilayers. Biochim Biophys Acta 1788:64–71
Pinaud F, Michalet X, Iyer G, Margeat E, Moore HP, Weiss S (2009) Dynamic partitioning of a Glycosyl-phosphatidylinositol-anchored protein in Glycosphingolipid-rich microdomains imaged by single-quantum dot tracking. Traffic 10:691–712
Lenne PF, Wawrezinieck L, Conchonaud F, Wurtz O, Boned A, Guo XJ, Rigneault H, He HT, Marguet D (2006) Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J 25:3245–3256
Raghupathy R, Anilkumar AA, Polley A, Singh PP, Yadav M, Johnson C, Suryawanshi S, Saikam V, Sawant SD, Panda A, Guo Z, Vishwakarma RA, Rao M, Mayor S (2015) Transbilayer lipid interactions mediate nanoclustering of lipid-anchored proteins. Cell 161:581–594
Olson F, Hunt C, Szoka F, Vail W, Papahadjopoulos D (1979) Preparation of liposomes of defined size distribution by extrusion through polycarbonate membranes. Biochim Biophys Acta Biomembr 557:9–23
Kremer J, Van der Esker M, Pathmamanoharan C, Wiersema P (1977) Vesicles of variable diameter prepared by a modified injection method. Biochemistry 16:3932–3935
Dua J, Rana A, Bhandari A (2012) Liposome: methods of preparation and applications. Int J Pharm Stud Res 3:14–20
Bezrukov SM (2000) Functional consequences of lipid packing stress. Curr Opin Colloid In 5:237–243
Hope MJ, Nayar R, Mayer LD, Cullis PR (1993) Reduction of liposome size and preparation of unilamellar vesicles by extrusion techniques. Liposome Technol 1:123–139
Angelova MI, Dimitrov DS (1986) Liposome electroformation. Faraday Discuss Chem Soc 81:303–311
Akashi K-i, Miyata H, Itoh H, Kinosita K (1996) Preparation of giant liposomes in physiological conditions and their characterization under an optical microscope. Biophys J 71:3242–3250
Sugiura S, Kuroiwa T, Kagota T, Nakajima M, Sato S, Mukataka S, Walde P, Ichikawa S (2008) Novel method for obtaining homogeneous giant vesicles from a monodisperse water-in-oil emulsion prepared with a microfluidic device. Langmuir 24:4581–4588
Arriaga LR, Datta SS, Kim SH, Amstad E, Kodger TE, Monroy F, Weitz DA (2014) Ultrathin shell double emulsion templated giant unilamellar lipid vesicles with controlled microdomain formation. Small 10:950–956
Richmond DL, Schmid EM, Martens S, Stachowiak JC, Liska N, Fletcher DA (2011) Forming giant vesicles with controlled membrane composition, asymmetry, and contents. Proc Natl Acad Sci 108:9431–9436
Hu PC, Li S, Malmstadt N (2011) Microfluidic fabrication of asymmetric giant lipid vesicles. ACS Appl Mater Interfaces 3:1434–1440
Lu L, Schertzer JW, Chiarot PR (2015) Continuous microfluidic fabrication of synthetic asymmetric vesicles. Lab Chip 15:3591–3599
Hamada T, Miura Y, Komatsu Y, Kishimoto Y, Vestergaard M d, Takagi M (2008) Construction of asymmetric cell-sized lipid vesicles from lipid-coated water-in-oil microdroplets. J Phys Chem B 112:14678–14681
Ito H, Yamanaka T, Kato S, Hamada T, Takagi M, Ichikawa M, Yoshikawa K (2013) Dynamical formation of lipid bilayer vesicles from lipid-coated droplets across a planar monolayer at an oil/water interface. Soft Matter 9:9539–9547
Elani Y, Law RV, Ces O (2015) Protein synthesis in artificial cells: using compartmentalisation for spatial organisation in vesicle bioreactors. Phys Chem Chem Phys 17:15534–15537
Hwang WL, Chen M, Cronin B, Holden MA, Bayley H (2008) Asymmetric droplet interface bilayers. J Am Chem Soc 130:5878–5879
Schmidt C, Barenholz Y, Huang C, Thompson T (1978) Monolayer coupling in sphingomyelin bilayer systems. Letters Nature 271:775–777
Blodgett KB (1935) Films built by depositing successive monomolecular layers on a solid surface. J Am Chem Soc 57:1007–1022
Roberts G (2013) Langmuir-blodgett films. Springer, New York
Watanabe R, Soga N, Yamanaka T, Noji H (2014) High-throughput formation of lipid bilayer membrane arrays with an asymmetric lipid composition. Sci Rep 4:7076
Crane JM, Kiessling V, Tamm LK (2005) Measuring lipid asymmetry in planar supported bilayers by fluorescence interference contrast microscopy. Langmuir 21:1377–1388
Anglin TC, Conboy JC (2009) Kinetics and thermodynamics of flip-flop in binary phospholipid membranes measured by sum-frequency vibrational spectroscopy. Biochemistry 48:10220–10234
Anglin TC, Cooper MP, Li H, Chandler K, Conboy JC (2010) Free energy and entropy of activation for phospholipid flip-flop in planar supported lipid bilayers. J Phys Chem B 114:1903–1914
Brown KL, Conboy JC (2013) Lipid flip-flop in binary membranes composed of phosphatidylserine and phosphatidylcholine. J Phys Chem B 117:15041–15050
Allhusen JS, Kimball DR, Conboy JC (2016) Structural origins of cholesterol accelerated lipid flip-flop studied by sum-frequency vibrational spectroscopy. J Phys Chem B 120:3157–3168
Montal M, Mueller P (1972) Formation of bimolecular membranes from lipid monolayers and a study of their electrical properties. Proc Natl Acad Sci 69:3561–3566
Collins MD, Keller SL (2008) Tuning lipid mixtures to induce or suppress domain formation across leaflets of unsupported asymmetric bilayers. Proc Natl Acad Sci 105:124–128
Murray DH, Tamm LK, Kiessling V (2009) Supported double membranes. J Struct Biol 168:183–189
Hussain NF, Siegel AP, Ge Y, Jordan R, Naumann CA (2013) Bilayer asymmetry influences integrin sequestering in raft-mimicking lipid mixtures. Biophys J 104:2212–2221
Pagano RE, Martin OC, Schroit AJ, Struck DK (1981) Formation of asymmetric phospholipid membranes via spontaneous transfer of fluorescent lipid analogs between vesicle populations. Biochemistry 20:4920–4927
Bloj B, Zilversmit D (1976) Asymmetry and transposition rate of phosphatidylcholine in rat erythrocyte ghosts. Biochemistry 15:1277–1283
Crain RC, Zilversmit DB (1980) Two nonspecific phospholipid exchange proteins from beef liver. 1. Purification and characterization. Biochemistry 19:1433–1439
Everett J, Zlotnick A, Tennyson J, Holloway P (1986) Fluorescence quenching of cytochrome b5 in vesicles with an asymmetric transbilayer distribution of brominated phosphatidylcholine. J Biol Chem 261:6725–6729
Herrmann A, Zachowski A, Devaux PF (1990) Protein-mediated phospholipid translocation in the endoplasmic reticulum with a low lipid specificity. Biochemistry 29:2023–2027
Holzer M, Momm J, Schubert R (2010) Lipid transfer mediated by a recombinant pro-sterol carrier protein 2 for the accurate preparation of asymmetrical membrane vesicles requires a narrow vesicle size distribution: a free-flow electrophoresis study. Langmuir 26:4142–4151
Redelmeier T, Hope M, Cullis P (1990) On the mechanism of transbilayer transport of phosphatidylglycerol in response to transmembrane pH gradients. Biochemistry 29:3046–3053
Hope MJ, Redelmeier TE, Wong KF, Rodrigueza W, Cullis PR (1989) Phospholipid asymmetry in large unilamellar vesicles induced by transmembrane pH gradients. Biochemistry 28:4181–4187
Mui B, Döbereiner H, Madden T, Cullis P (1995) Influence of transbilayer area asymmetry on the morphology of large unilamellar vesicles. Biophys J 69:930
Pautot S, Frisken BJ, Weitz DA (2003) Engineering asymmetric vesicles. Proc of Natl Acad Sci USA 100:10718–10721
Elani Y, Purushothaman S, Booth PJ, Seddon JM, Brooks NJ, Law RV, Ces O (2015) Measurements of the effect of membrane asymmetry on the mechanical properties of lipid bilayers. Chem Commun 51:6976–6979
Zhang X, Zong W, Hu Y, Luo N, Cheng W, Han X (2016) A pH-responsive asymmetric lipid vesicle as drug carrier. J Microencapsul 33:663–668
Lu L, Doak WJ, Schertzer JW, Chiarot PR (2016) Membrane mechanical properties of synthetic asymmetric phospholipid vesicles. Soft Matter 12:7521–7528
Kamiya K, Kawano R, Osaki T, Akiyoshi K, Takeuchi S (2016) Cell-sized asymmetric lipid vesicles facilitate the investigation of asymmetric membranes. Nat Chem 8:881–889
Dodziuk H (2006) Cyclodextrins and their complexes: chemistry, analytical methods, applications. Wiley, Weinheim
Somogyi G, Posta J, Buris L, Varga M (2006) Cyclodextrin (CD) complexes of cholesterol–their potential use in reducing dietary cholesterol intake. Die Pharmazie- Int J Pharm Sci 61:154–156
Huang Z, London E (2013) Effect of cyclodextrin and membrane lipid structure upon cyclodextrin-lipid interaction. Langmuir 29:14631–14638
Lin Q, London E (2014) Preparation of artificial plasma membrane mimicking vesicles with lipid asymmetry. PLoS One 9:e87903
Szente L, Fenyvesi É (2017) Cyclodextrin-lipid complexes: cavity size matters. Struct Chem 28:479–492
Kim J, London E (2015) Using sterol substitution to probe the role of membrane domains in membrane functions. Lipids 50:721–734
Zidovetzki R, Levitan I (2007) Use of cyclodextrins to manipulate plasma membrane cholesterol content: evidence, misconceptions and control strategies. Biochim Biophy Acta Biomembr 1768:1311–1324
Ohtani Y, Irie T, Uekama K, Fukunaga K, Pitha J (1989) Differential effects of α-, β-and γ-cyclodextrins on human erythrocytes. Eur J Biochem 186:17–22
Christian A, Haynes M, Phillips M, Rothblat G (1997) Use of cyclodextrins for manipulating cellular cholesterol content. J Lipid Res 38:2264–2272
Legendre J, Rault I, Petit A, Luijten W, Demuynck I, Horvath S, Ginot Y, Cuine A (1995) Effects of β-cyclodextrins on skin: implications for the transdermal delivery of piribedil and a novel cognition enhancing-drug, S-9977. Eur J Pharm Sci 3:311–322
Niu S-L, Mitchell DC, Litman BJ (2002) Manipulation of cholesterol levels in rod disk membranes by methyl-β-cyclodextrin effects on receptor activation. J Biol Chem 277:20139–20145
Tanhuanpää K, Somerharju P (1999) γ-Cyclodextrins greatly enhance translocation of hydrophobic fluorescent phospholipids from vesicles to cells in culture IMPORTANCE OF MOLECULAR HYDROPHOBICITY IN PHOSPHOLIPID TRAFFICKING STUDIES. J Biol Chem 274:35359–35366
Tanhuanpää K, Cheng KH, Anttonen K, Virtanen JA, Somerharju P (2001) Characteristics of pyrene phospholipid/γ-cyclodextrin complex. Biophys J 81:1501–1510
Anderson TG, Tan A, Ganz P, Seelig J (2004) Calorimetric measurement of phospholipid interaction with methyl-beta-cyclodextrin. Biochemistry 43:2251–2261
Kilsdonk EP, Yancey PG, Stoudt GW, Bangerter FW, Johnson WJ, Phillips MC, Rothblat GH (1995) Cellular cholesterol efflux mediated by cyclodextrins. J Biol Chem 270:17250–17256
Cheng H-T, London E (2009) Preparation and properties of asymmetric vesicles that mimic cell membranes effect upon lipid raft formation and transmembrane helix orientation. J Biol Chem 284:6079–6092
Cheng HT, London E (2011) Preparation and properties of asymmetric large unilamellar vesicles: interleaflet coupling in asymmetric vesicles is dependent on temperature but not curvature. Biophys J 100:2671–2678
Chiantia S, Schwille P, Klymchenko AS, London E (2011) Asymmetric GUVs prepared by MbetaCD-mediated lipid exchange: an FCS study. Biophys J 100:L1–L3
Oglęcka K, Rangamani P, Liedberg B, Kraut RS, Parikh AN (2014) Oscillatory phase separation in giant lipid vesicles induced by transmembrane osmotic differentials. elife 3:e03695
Chiantia S, London E (2012) Acyl chain length and saturation modulate interleaflet coupling in asymmetric bilayers: effects on dynamics and structural order. Biophys J 103:2311–2319
Chiantia S, Klymchenko AS, London E (2012) A novel leaflet-selective fluorescence labeling technique reveals differences between inner and outer leaflets at high bilayer curvature. Biochim Biophys Acta Biomembr 1818:1284–1290
Armstrong VT, Brzustowicz MR, Wassall SR, Jenski LJ, Stillwell W (2003) Rapid flip-flop in polyunsaturated (docosahexaenoate) phospholipid membranes. Arch Biochem Biophys 414:74–82
Smith M, Jungalwala F (1981) Reversed-phase high performance liquid chromatography of phosphatidylcholine: a simple method for determining relative hydrophobic interaction of various molecular species. J Lipid Res 22:697–704
Lin Q, London E (2015) Ordered raft domains induced by outer leaflet sphingomyelin in cholesterol-rich asymmetric vesicles. Biophys J 108:2212–2222
Bakht O, Pathak P, London E (2007) Effect of the structure of lipids favoring disordered domain formation on the stability of cholesterol-containing ordered domains (lipid rafts): identification of multiple raft-stabilization mechanisms. Biophys J 93:4307–4318
Vitrac H, MacLean DM, Jayaraman V, Bogdanov M, Dowhan W (2015) Dynamic membrane protein topological switching upon changes in phospholipid environment. Proc Natl Acad Sci 112:13874–13879
Vitrac H, Bogdanov M, Dowhan W (2013) In vitro reconstitution of lipid-dependent dual topology and postassembly topological switching of a membrane protein. Proc Natl Acad Sci 110:9338–9343
Perillo VL, Peñalva DA, Vitale AJ, Barrantes FJ, Antollini SS (2016) Transbilayer asymmetry and sphingomyelin composition modulate the preferential membrane partitioning of the nicotinic acetylcholine receptor in lo domains. Arch Biochem Biophys 591:76–86
Li G, Kim J, Huang Z, Clair JRS, Brown DA, London E (2016) Efficient replacement of plasma membrane outer leaflet phospholipids and sphingolipids in cells with exogenous lipids. Proc Natl Acad Sci 201610705
Visco I, Chiantia S, Schwille P (2014) Asymmetric supported lipid bilayer formation via methyl-β-cyclodextrin mediated lipid exchange: influence of asymmetry on lipid dynamics and phase behavior. Langmuir 30:7475–7484
Heberle FA, Marquardt D, Doktorova M, Geier B, Standaert RF, Heftberger P, Kollmitzer B, Nickels JD, Dick RA, Feigenson GW (2016) Sub-nanometer structure of an asymmetric model membrane: Interleaflet coupling influences domain properties. Langmuir 32:5195–5200
Petazzi RA, Gramatica A, Herrmann A, Chiantia S (2015) Time-controlled phagocytosis of asymmetric liposomes: application to phosphatidylserine immunoliposomes binding HIV-1 virus-like particles. Nanomedicine 11:1985–1992
Spector AA, Yorek MA (1985) Membrane lipid composition and cellular function. J Lipid Res 26:1015–1035
Adada M, Luberto C, Canals D (2016) Inhibitors of the sphingomyelin cycle: sphingomyelin synthases and sphingomyelinases. Chem Phys Lipids 197:45–59
Delgado A, Casas J, Llebaria A, Abad JL, Fabrias G (2006) Inhibitors of sphingolipid metabolism enzymes. Biochim Biophys Acta Biomembr 1758:1957–1977
Aguilar PS, Heiman MG, Walther TC, Engel A, Schwudke D, Gushwa N, Kurzchalia T, Walter P (2010) Structure of sterol aliphatic chains affects yeast cell shape and cell fusion during mating. Proc Natl Acad Sci 107:4170–4175
Ott RG, Athenstaedt K, Hrastnik C, Leitner E, Bergler H, Daum G (2005) Flux of sterol intermediates in a yeast strain deleted of the lanosterol C-14 demethylase Erg11p. Biochim Biophys Acta Mol Cell Biol Lipids 1735:111–118
Dowhan W (2009) Molecular genetic approaches to defining lipid function. J Lipid Res 50:S305–S310
Wikström M, Kelly AA, Georgiev A, Eriksson HM, Klement MR, Bogdanov M, Dowhan W, Wieslander Å (2009) Lipid-engineered Escherichia Coli membranes reveal critical lipid headgroup size for protein function. J Biol Chem 284:954–965
Kainu V, Hermansson M, Somerharju P (2010) Introduction of phospholipids to cultured cells with cyclodextrin. J Lipid Res 51:3533–3541
Kainu V, Hermansson M, Somerharju P (2008) Electrospray ionization mass spectrometry and exogenous heavy isotope-labeled lipid species provide detailed information on aminophospholipid acyl chain remodeling. J Biol Chem 283:3676–3687
Koivusalo M, Jansen M, Somerharju P, Ikonen E (2007) Endocytic trafficking of sphingomyelin depends on its acyl chain length. Mol Biol Cell 18:5113–5123
Acknowledgement
Support from NIH grant GM 112638 and GM 122493 is gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer Nature Singapore Pte Ltd.
About this chapter
Cite this chapter
St. Clair, J.R., Wang, Q., Li, G., London, E. (2017). Preparation and Physical Properties of Asymmetric Model Membrane Vesicles. In: Epand, R., Ruysschaert, JM. (eds) The Biophysics of Cell Membranes. Springer Series in Biophysics, vol 19. Springer, Singapore. https://doi.org/10.1007/978-981-10-6244-5_1
Download citation
DOI: https://doi.org/10.1007/978-981-10-6244-5_1
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-10-6243-8
Online ISBN: 978-981-10-6244-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)