
Abstract
Suicide is a complex and multifactorial disorder. Two important risk factors, genetic and environment, contribute significantly to the psychopathology and aetiology of suicidal behaviour. In the last two decades, family, twins and adoption studies have been conducted for the heritability of suicidal behaviour. Recently, molecular genetic approaches, including Genome Wide Association (GWAS) and Whole Exome Sequencing (WES) have been applied into the genetics of suicidal behaviour. Using these techniques, several new common and rare variants have been identified. In this chapter, we summarize and critically evaluate the recent developments in the field of genetics of suicide. The obtained data suggests that suicide behaviour is a highly complex and polygenic disorder. Despite a significant progress in this area, future studies are needed in a larger population. We have also discussed the clinical translation of suicide genetics and future directions.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
Suicidal behaviour is a complex and multifactorial behaviour. About million people commit suicide each year. Suicide is ranked second among cause of death in adolescent. Although, suicidal behaviour has been attributed to various factors including, psychological and economical, there is a great deal of evidence point to interaction of genetics and environment that may significantly contribute to the suicidal behaviour. In this chapter, we will first discuss epidemiology of suicidal behaviour describing family, twin, and adoption studies. We will then examine the major candidate gene studies. Lastly, we will review recent progress in genome-wide association study and next-generation sequence in the suicidal behaviour. We will also address the new ideas and future directions in order to find the susceptible genetic factors associated with suicidal behaviour.
Genetic Epidemiology
Familial clustering of suicidal behaviour and psychopathology has been reported for adolescent suicide attempters. Mittendorfer-Rutz, Rasmussen, and Wasserman (2008) conducted the largest epidemiology studies with 14,440 suicide attempters and 14,440 healthy controls that were born in Sweden between 1968 and 1980. For youth suicide attempt, familial risk factors were siblings (OR 3.4; 2.8–4.1), maternal (OR 2.7; 2.5–3.1), and paternal (OR 1.9; 1.7–2.1). The important risk factors included were familial personality, substance abuse disorders, maternal schizophrenia, non-affective psychoses, organic disorders, and parental neurotic stress-related and somatoform disorders (1.9–3.2 fold increase), and paternal (OR 1.9; 1.6–2.3) and maternal (OR 1.8; 1.3–2.4) suicide completion. It was found that familial suicide completion had stronger effect on suicide attempt of earlier onset and in boys. About half of all suicide attempts were attributed to familial psychopathology (13%), family suicide attempt (7%), suicide completion (1%), and own psychopathology. This familial study directly supports the evidence that genetic risk factors contribute to the suicidal behaviour.
Twin studies are a useful way for analysing concordance. Individuals share 100% genes between monozygotic twins and share 50% genes between dizygotic twins. Twins study can separate environmental variance from genetic factor. Roy, Segal, Centerwall, and Robinette (1991) examined 176 twin pairs in which one or both twins had committed suicide. Interestingly, they found concordant difference between monozygotic twin pairs (11.3%) and dizygotic twin pairs (1.8%). The presence of psychiatric disorder in the twins and their families was examined in 11 twin pairs, two of whom were concordant for suicide. Out of 13, 11 twin suicide victims had been treated for psychiatric disorders. In addition, twins in 10 pairs had other first- or second-degree relatives who had been treated for psychiatric disorders. These twin data suggest that genetic factors related to suicide may largely represent a genetic predisposition for the psychiatric disorders associated with suicide. Statham et al. (1998) examined the epidemiology and genetics of suicidality, which included reports of persistent suicidal thoughts or a plan or suicide attempt in a large number of community-based samples of monozygotic and dizygotic twin pairs (5995 twins). They found that lifetime prevalence of suicidal thoughts and attempts was remarkably constant across birth cohorts, and across gender. History of suicide attempts or persistent thoughts in the respondent’s co-twin remained a powerful predictor in monozygotic pairs, but was not consistently predictive in dizygotic pairs. Overall, genetic factors accounted for approximately 45% of the variance in suicidal thoughts and behaviour. Voracek and Loibl (2007) conducted a systematic review of twins study and they collected 32 studies, which included 19 case reports, 5 register-based studies, 4 population-based epidemiological studies, and 4 surviving co-twins study. From register-based studies and case reports, concordance for completed suicide was significantly more frequent among monozygotic (24.1%) than dizygotic twin pairs (2.3%). From population-based epidemiological studies, heritability of suicidal behaviour was estimated at 30–55%. This twin study suggests strong genetic contributions to the suicide risk.
Adoption studies have also been conducted in the suicide behaviour. Schulsinger, Ketty, Rosenthal, and Wender (1979) examined 57 suicide victims, which were adopted in the Danish family. They compared the suicide rates between biological relatives of suicide victims and non-suicidal adoptee control group, which were found to be 4.46 and 0.74%, respectively. None of the relatives of the adopted families were affected by suicide. Another study reported by Von Borczyskowski, Lindblad, Vinnerljung, Reintjes, and Hjern (2011) was conducted in the Swedish registry with a national cohort of 2,471,496 individuals, including 176,000 adoptees for suicide during 1987–2001. They found that biological parent suicide and severe psychiatric morbidity which were the major risk factors for suicide in adopted and in non-adopted offspring.
Candidate Gene Association Studies of Suicidal Behaviour
Gene studies of most candidates are based on case-control association study design. The basic principle of genetic association studies is the hypothesis of common disease and common variants. The groups of cases and controls are enrolled in the association study and the allele or genotype frequencies are determined and compared statistically. If the polymorphisms are more frequent in one group than the other group, the genetic variants show association with the disease and the candidate gene may be the susceptible genes for suicide or psychiatric diseases. The selections of genetic variants for the study are usually based on the plausible hypothesis, such as serotonin transporter for antidepressants. It is noticeable that the association studies have several limitations. First, spurious positive findings and false-positive findings are unavoidable due to the population stratification. Second, if the sample sizes are small, then the statistical power is very low. Lastly, most of common variants are located within the intron regions and elucidation of function relevance is confronted. Such limitations also include clinical and diagnostic heterogeneity, low statistical power if sample sizes are small, often limited biological evidence of candidate gene selection, and unknown functional relevance of tested single nucleotide polymorphisms (SNPs), as well as population stratification within the sample leading to spurious positive findings or false-negative associations. Despite these limitations, the candidate gene association studies are still an effective solution in finding for susceptible genes in suicide victims. Here we summarize the candidate gene studies in suicidal behaviour examined using association studies.
Serotonergic System
Studies showing the role of serotonin, a monoamine neurotransmitter, in suicidal behaviour date back to the 70s of the last century. Serotonergic functions have been demonstrated to be critical for the regulation of impulsive, aggressive, and suicidal behaviour (Koller, Bondy, Preuss, Bottlender, & Soyka, 2003; Virkkunen et al., 1989). For example, several studies have reported that the expression of genes associated with serotonergic system is altered in the post-mortem brains of suicide victims, especially in the prefrontal cortex. These include 5HT2A, 5HT1A, and 5HT2C (Celada, Puig, Amargós-Bosch, Adell, & Artigas, 2004; Holmes, 2008; Sugden et al., 2009). Clinical studies also found that the concentration of the serotonin metabolite 5-hydroxyindoleacetic acid (5-HIAA) is decreased in the cerebrospinal fluid of the suicidal patients (Asberg, 1997; Placidi et al., 2001). Most of the genetic association studies are conducted using the genes of serotonergic neurotransmission pathway.
Tryptophan Hydroxylase (TPH1)
TPH1 is located on the chromosome 11p15.3-p14 and the full-length fragment covers 29 kbp. Serotonin is catalyzed from l-Tryptophan by tryptophan hydroxylase (THP), which is a rating-limiting enzyme (Boularand, Darmon, Ravassard, and Mallet, 1995). To identify the susceptible risk single nucleotide polymorphism (SNP)s for suicidal behaviour, several studies focused on common variants associated with THP1 gene. Early studies focused on the two SNPs: A779C (rs1799913) and A218C (rs1800532). Both these SNPs are located on intron 7. Nielsen et al. (1994) reported variation on a polymorphism in intron 7 with an A to C substitution at nucleotides 779 (A779C) which was associated with a history of violent suicide attempts. Following this, two studies found C allele as the risk variant (Nielsen et al., 1998; Roy et al., 2001) and one study showed increased frequency of the A allele in suicide attempters (Mann et al., 1997). The first meta-analysis conducted by Lalovic and Turecki (2002) reported no association between A218C with suicide behaviour. Another meta-analysis investigated A218C in Caucasians, which showed that suicidal patients frequently carried A218 allele (Rujescu, Giegling, Sato, Hartmann, & Möller, 2003). A meta-analysis summarizing 9 studies reported an association between A218C polymorphisms and suicidal behaviour (Bellivier, Chaste, and Malafosse, 2004). Li, Duan, and He (2006) performed meta-analysis of 22 studies and confirmed this evidence. Another meta-analysis was conducted by Clayden, Zaruk, Meyre, Thabane, and Samaan (2012) which further supported the involvement of A218C and suicidal behaviour. Recently, González-Castro et al. (2014) collected 37 literatures about genetic association studies of THP1 and reviewed 2 polymorphisms (one is A218C, the other is A779C). This meta-analysis study involved a total number of sample 5683 cases and 11,652 controls. Meanwhile, subgroup analyses were performed in the Caucasian and Asian populations. The results demonstrated that both A218C and A779C were susceptible to suicide behaviour. Other studies examining the A allele of the A218C showed frequent association with suicide attempters than non-attempters (Abbar et al., 2001; Galfalvy et al., 2009; Souery et al., 2001; Tsai et al., 1999). On the other hand, the C allele was associated with suicide attempts in two studies (Paik, Toh, Kim, & Lee, 2000; Stefulj, Kubat, Balija, & Jernej, 2006). Other common variants identified within the promoter region are T7180G, C7065T, A6526G, and G5806T. The A allele of the A6526G was found to be in linkage disequilibrium with A779C and was associated with suicide attempts (Galfalvy et al., 2009; Rotondo et al., 1999). A study by Turecki et al. (2001) showed risk haplotype associated with suicide completion (6526G–5806T–218C). An association between the haplotype TCAAA of T7180G-C7065T-A6526G-A218C-A779C SNPs and suicide attempts have also been reported (Liu et al., 2006).
Several studies support the evidence that TPH1 is involved in the personality trait, especially anger and aggression. Rujescu et al. (2002) reported that individuals with “A” allele had higher scores on the trait Anger Scale of the State-Trait Anger Expression Inventory, STAX in the healthy control and suicide attempters. Baud et al. (2009) also reported that suicide attempters with “AA” genotype had lower score on the anger than with CC genotype. Cicchetti, Rogosch, and Thibodeau (2012) examined common variants in the TPH1 in child maltreatment and non-maltreatment low-income children. Interestingly, they found that TPH1 polymorphisms moderated the effects of maltreatment subtype on adult reports of antisocial. Andre et al. (2013) analysed the interaction between the TPH1 genotype, treatment response, and the different temperament dimensions in 97 subjects with major depression who was treated serotonin reuptake inhibitors. They found TPH1 genotype and treatment response had interactive effects on both HA1 and HA2 scores. Interaction between CC genotype and remission or treatment status differentiates between different temperament profiles. Karanović et al. (2016) assessed TPH1 variant rs1800532 and its possible interaction with recent stressful life events as risk factors for suicide attempt in Serbian psychiatric patients, including 165 suicide attempters and 188 suicide non-attempters. They found that rs1800532 and recent stressful life events were independently associated with suicide attempt, while rs1800532 did not moderate the effect of recent stressful life events on suicide attempt vulnerability.
Tryptophan Hydroxylase (TPH2)
Walther et al. (2003) first reported a second TPH isoform in mice and termed it TPH2. Breidenthal, White, and Glatt (2004) screened the whole genomic region and identified several common variants in this gene, which served for several association studies. Zill et al. (2004) Investigated the SNPs and haplotype in 263 suicide victims and 266 healthy controls with 10 SNPs in the TPH2 gene. One SNP was detected to be significantly associated with suicide (p = 0.004). Haplotype analysis also supported for this association (p = 0.001). Several case-control studies reported association between TPH2 gene and suicide. Some studies showed positive results, but others showed negative results. Lately, González-Castro, Juárez-Rojop, López-Narváez, and Tovilla-Zárate (2014) conducted a meta-analysis focusing three SNPs (G603T, A473T, and G19918A) in 4196 cases and 5990 controls. The meta-analysis did not find any significant association with patients showing suicidal behaviour. Choong, Tee, and Tang (2014) selected 9 SNPs and conducted a meta-analysis with a total of 2536 cases and 3101 controls in 11 case-control studies. No significant results were detected, suggesting that TPH2 may not play a significant role in suicidal behaviour. Interestingly, polymorphisms in the TPH2 gene were found in relation to personality traits like aggression or impulsivity, emotional regulation, and neuroticism (Perez-Rodriguez et al. 2010; Hong et al. 2011).
Serotonin Transporter
The function of serotonin transporter (5-HTT) is to reuptake the released serotonin from the synaptic cleft. A function insertion/deletion variant in the promoter region contain two or three alleles called short(s) and long (IA and IG). Polymorphisms in HTTLPR have been associated with numerous neuropsychiatric disorders, including schizophrenia, depression, and aggression. These disorders in turn increase the susceptibility of suicidal behaviour. Anguelova, Benkelfat, and Turecki (2003) conducted a meta-analysis pooling 12 studies investigating the 5-HTT promoter polymorphism. A total of 1,168 cases (suicide completers and suicide attempters) and 1,371 controls (compromised of 10 Caucasian populations, 1US population and 1 Chinese population) were studied. A significant association of the s allele was demonstrated with suicidal behaviour. Lin and Tsai (2004) also performed a meta-analysis but reported no association between 5-HTTLPR polymorphism and suicidal behaviour (p = 0.379). When they compared subjects with the same psychiatric diagnosis, the genotypes carrying the s allele were significantly more frequent in suicide attempters than in non-attempters (p = 0.004). In addition, the s allele was associated with violent suicide (p = 0.0001) but not with non-violent suicide (p = 1.00). Li and He (2007) provided a comprehensive meta-analysis pooling 39 studies and found that long alleles to be associated with decreased risk of suicide attempts (OR 0.88; 95% CI 0.80–0.97). Clayden et al. (2012) analysed the association between the serotonin transporter s allele and suicidal behaviour in 31 primary case-control studies (6324 cases and 10,285 controls). The results revealed no significance association, however, subgroups of suicide attempters demonstrated significance association with serotonin transporter (SLC6A4) 5-HTTLPR [OR 1.13 (95% CI 1.05–1.21), p = 0.001], suggesting that the 5-HTTLPR and rs1800532 polymorphisms are significantly associated with suicide attempts, but not with completed suicides. A recent meta-analysis by Schild, Pietschnig, Tran, and Voracek (2013) included 2536 cases and 3984 controls and further supported the involvement of the 5-HTTLPR in suicidal behaviour. Anguelova, Benkelfat, and Turecki (2003) combined 12 studies investigating 5-HTT promoter 44 bp insertion/deletion polymorphism in a total of 1,599 and 2,539 subjects. The combined evidence was significant association with the 5-HTT locus (OR 1.1795%, CI 1.04–1.32, p = 0.009), suggesting that 5-HTT may play a role in the predisposition to suicide. Several other studies found an association between the SS genotype/S allele and increased lethality (Saiz et al., 2011; Wasserman et al., 2007) and violent attempts (Neves et al., 2008, 2010). In a large cohort of 5608 controls and subjects suffering from various psychiatric disorders, Perroud et al. (2010) studied the frequency of haplotypes and corresponding genotypes combining the 5-HTTLPR and the other serotonin transporter promoter functional variant (rs25531). They found that a very rare G-14/G-16 genotype was carried by only three subjects who were women with a history of suicide attempt. Taken together, these studies provide evidence that 5-HTTLPR may play crucial role in the suicidal behaviour.
Serotonin Receptors
Serotonin receptors, a G protein-coupled receptor super family, consist of receptors named 5-HT1 to 5-HT7 (Cowen, 1991). Here, we focus on the genetics of serotonin receptors in the context of suicidal behaviour.
5-HT1A Receptor
Lemonde et al. (2003) conducted common variant of C1109G on the promoter region in a sample of suicide victims and control subjects. They found that G allele was significantly over-represented in the suicide group. In a total of 191 suicide and 218 healthy control subjects, Samadi et al. (2012) studied association between polymorphisms C1019G, C47T (Pro16Leu), and G815A (Gly272Asp) and suicidal behaviour taking into account age, gender, and the presence of stressful life, and loss events in 1 year prior to suicide. They found that the distribution of C1019G genotypes was significantly different in suicide victims and healthy controls (p = 0.002), and the GG genotype was associated with a significantly higher number of more stressful life and loss events in the suicide victims (p = 0.017, p = 0.037, respectively). Two missense polymorphisms, Pro16Leu and Gly 272 Asp, have also been investigated in the Japanese subjects and the results revealed no significant differences in genotype distribution or allele frequencies between suicide victims and controls. Ohtani, Shindo, and Yoshioka (2004) conducted a meta-analysis for the A652G polymorphisms and reported no association and three further meta-analyses for the C1019G variants were also negative (Rivera, Bermúdez Ocaña, Camarena Medellín, & Tovilla-Zárate, 2012, Schild et al., 2013)
5-HT1B Receptor
The 5-HT1B gene is located at 6q13 which includes 1.17 kb of axons regions. The 5-HT1B has received attention due to its association with aggressive and impulsive behaviour. Kia-Keating, Glatt, and Tsuang (2007) performed a meta-analysis of 6 studies with various study designs and did not find significant results for the G861C allele with suicidal behaviour. Another meta-analysis also did not find significant results in a sample of 2,948 cases and 4,066 controls (Clayden et al., 2012). Jensen et al. (2009) reported that a mirSNP, regulated by mir-96, was significantly associated with aggressive behaviour. Conner et al. (2010) genotyped 5 polymorphisms in the HTR1B gene (rs11568817, rs130058, rs6296, rs6297, rs13212041) in 361 University students. They found rs13212041 polymorphism, which modulates miRNA-mediated regulation of HTR1B expression, can predict self-reported anger and hostility among young men. Recently, Hakulinen et al. (2013) examined a connection between polymorphism of HTR1B in 967 participants from a large population-based sample with 27-year follow-up. They found rs6296 was associated with childhood aggressive behaviour but not with adulthood anger or hostility.
5-HT2A Receptor
Earlier, the density of brain and platelet 5HT2A receptors has been shown to be increased in suicidal patients (Du, Bakish, Lapierre, Ravindran, & Hrdina, 2000). Thus, 5-HT2A receptor has been considered to be a major candidate gene in suicidal behaviour (Bachus et al., 1997). A first meta-analysis with 9 studies, which included 596 suicidal completers or attempters and 1003 healthy controls, showed no association with T102C located in an axon (Anguelova et al., 2003). Later, Li et al. (2006) carried out a meta-analysis of 25 studies on suicidal behaviour and they also did not find significant association with suicide attempter (OR 0.98; 95% CI 0.83–1.16). On the other hand, another SNP in the promoter region A1438G in seven studies showed that A-allele has protective risk of suicide (OR 0.67, 95% CI 0.59–0.89). Fanous et al. (2009) investigated the tagSNPs, spanning HTR2A genome regions, in the 270 families with suicidal ideation. They did not find significant association with the any tagSNPs or haplotype and suicidal ideation. Interestingly, Giegling, Hartmann, Möller, & Rujescu, 2006) found that the HTR2A variants rs594242-rs6311: G-C and rs6311 C were associated with increased non-violent and impulsive suicidal behaviour, respectively. On the other hand, the CC genotype of the T102C SNP was more frequent among suicide attempters (Vaquero-Lorenzo et al., 2008; Wrzosek et al., 2011). In 227 suicide attempters, 686 non-suicidal psychiatric patients, and 420 healthy controls from a homogeneous Spanish Caucasian population, Saiz et al. (2011) found no differences in genotype frequencies between the three groups.
Dopaminergic System and Catecholamine’s
Soderstrom, Blennow, Manhem, and Forsmen (2001) reported that high cerebrospinal fluid (CSF) levels of the dopamine metabolite HVA correlates with human aggression, whereas Pitchot, Hansenne, and Ansseau (2001) found the involvement of D2-dopaminergic functions in the biology of suicide by demonstrating a smaller growth hormone response to apomorphine, a dopaminergic agonist. These investigators also found that reduced dopamine function in depressed patients was related to suicidal behaviour but not its lethality (Pitchot et al., 2001b). Catechol-O-Methyltransferase (COMT), a major enzyme which can inactivate the activity of dopamine and noradrenalin has received much attention in psychiatric illnesses because of the functional polymorphisms associated with val158met (Chen et al., 2004). Kia-Keating et al. (2007) performed a meta-analysis of a total 519 cases and 933 healthy controls and included 6 studies. They found Val158Met was significantly associated with suicidal behaviour. Additionally, Calati et al. (2011) summarized 10 studies with 1324 patients but found no association between val158met and suicidal behaviour. Tovilla-Zárate et al. (2011) further confirmed no difference in this functional polymorphism between suicidal patients and healthy control.
Brain-Derived Neurotrophic Factors
BDNF is a member of the neurotrophin family of growth factors (Laske & Eschweiler, 2006). Several studies have revealed that the expression of BDNF is lower in postmortem brain of suicide victims (Dwivedi et al., 2003) as well as in patients with suicidal behaviour (Dwivedi et al., 2003; Lee, Kim, Park, & Kim, 2007). Because of these findings, several studies attempted to study polymorphism in BDNF gene in suicide. The most studied polymorphism in BDNF gene is Val66Met. This polymorphism is a missense mutation at position 66 resulting in a valine to methionine substitution. Previous studies demonstrated that the BDNF val66met polymorphism was associated with impaired cognitive functioning and worse memory functioning in humans (Egan et al., 2003). Hong et al. (2003) found that V66M polymorphism in the BDNF was not associated with suicide attempts in bipolar disorder or major depressed patients. No association between V66M and suicide victims was also reported by Zarrilli et al. (2009). However, Vincze et al. (2008) found the frequency of Met allele in violent bipolar disorder patients was higher than the healthy controls. Kim et al. (2008) studied the allelic and genotypic distributions of BDNF Val/Met in a population of 169 bipolar patients and 251 normal controls and found that the rate of suicide attempts among the Val/Val (11.3%), Val/Met (28.8%), and Met/Met (38.9%) genotype groups were significantly different. Relative to patients with the Val/Val genotype had a 4.9-fold higher risk of suicide attempts than those with the Met/Met genotype. A recent study with combining 11 published studies reported that Met allele was associated with suicide victims (p = 0.032, ORmet = 1.16) (Zai et al., 2012). Iga et al. (2007) studied BDNF Val66Met polymorphism in depressed patients in a Japanese population. They found similar genotypic distributions and allele frequencies among the patients and control subjects. However, when the relationships of the polymorphism with several clinical variables were examined, Met allele had significant effects on psychotic feature and suicidal behaviour and family history. These results suggest that the BDNF Val66Met polymorphism is not related to the development of MDD but related to clinical features of MDD. Another meta-analysis with 8 published studies showed that the met carrier have a higher trend for suicide. The Met allele and genotype show significance with suicide attempt (Zai et al., 2012). Gonzalez-Castro et al. (2015) studied the association of Val66Met BDNF polymorphism in bipolar disorder in a Mexican population. Although their results from the meta-analysis (9,349 cases and 7,437 controls) did not show a significant association in any of the models, however, when they analysed the association between rs6265 and lifetime history of suicidal behaviour, they found an association between genotype Val–Val and suicide attempt. Recently, Bresin, Sima Finy, and Verona (2013) explored the interaction between retrospective reports of childhood emotional environment and the BDNF Val66Met polymorphism in relation to a history of two main types of self-injurious behaviours: suicide attempt and non-suicidal self-injury, in a sample of individuals with a history of involvement in the criminal justice system. For individuals with two Val alleles, these investigators found a significant direct relationship between emotional maltreatment and self-injurious behaviours. However, the relationship was not significant for Met carriers.
Genome-Wide Association Studies of Suicide Behaviour
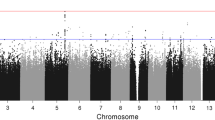
With the rapid development of technological advances in genomics, Genome-wide association study (GWAS) can scan more than 1 million SNPs across the genome in parallel and provide an in-depth knowledge of the genetic information. Several GWAS studies have emerged in the suicidal behaviour.
Treatment-Emergent Suicidal Ideation
Laje et al. (2009) collected 90 white participants who were treatment-emergent suicidal ideators and matched healthy controls. Based on these subjects, the authors performed a GWAS and 109365 SNPs were genotyped on the Illumina Human-1 Bead chip. One SNP in the gene PALPLN reached genome-wide significant and another SNP in the gene IL28RA showed a trend. PAPLN encodes papil in, a protoglycan-like sulfated glycoprotein. IL28RA encodes an interleukin receptor. Perroud et al. (2012) examined to identify genetic variants involved in increasing suicidal ideation during escitalopram and nortriptyline treatment. They studied a total number of 706 major depressed patients treated with escitalopram or nortriptyline and genotyped those using platform based on the Illumina 610-Quad Beadchips. The genetic marker most significantly associated with increasing suicidality was a SNP (rs11143230) located 30 kb down stream of a gene encoding guanine deaminase on chromosome 9q21.13. Two suggestive drug-specific associations within KCNIP4 (Kv channel-interacting protein 4; chromosome 4p15.31) and near ELP3 (elongation protein 3 homolog; chromosome 8p21.1) were found in subjects treated with escitalopram. The most significant association with in a set of 33 candidate genes was in the neurotrophic tyrosine kinase receptor type 2 genes. They also found a trend for an association within genes such as GRIP1, NXPH1 and ANK3, previously associated with psychiatric phenotypes indirectly linked to suicidal behaviour. The results suggest novel pathways involved in increasing suicidal ideation during antidepressant treatment and can help to target treatment to reduce the risk of this dramatic adverse event. A third GWAS on treatment-emergent suicidal ideators was provided by Menke et al. (2012) in depressed patients (n = 397) with suicidal thoughts. They did not find any genome-wide significant variant. The best associated SNP was rs1630535 (p value = 1.3 × 10−7). Fourteen variants showed nominal significant association with the same risk allele in the replication sample. The study suggests that a combination of genetic markers maybe used to identify patients at risk for treatment-emergent suicidal ideation.
Suicide Attempts
Perlis et al. (2010) reported the first GWAS on suicide attempts. Data on lifetime suicide attempts were extracted from GWAS of bipolar I and II disorder as well as MDD. For suicide attempt in bipolar disorder, rs1466864 was identified to be the most significant. Five loci also showed suggestive association. For suicide attempt in major depressive disorder, rs2576377 in ABI3BP gene showed the strongest association where as six loci showed nominal association. Schosser et al. (2011) performed a genome-wide association scan in 251 depressed patients with serious suicidal attempts. No SNP reached the cutoff of genome-wide significance level. For the quantitative trait, GFRA1 was detected to be associated with suicide attempts (p = 2 × 10−6). For the discrete trait of suicide attempt, SNPs in KIAA1244 and RGS18 showed suggestive associations. A polymorphism in NTRK2, previously associated with suicidality (Kohli et al., 2010; Perroud et al., 2009; Ropret, Zupanc, Komel, & Videtic Paska, 2015), was also validated. Willour et al. (2012) conducted GWAS and compared the SNP genotypes between 1201 bipolar subjects with a history of suicide attempts and 1,497 bipolar subjects without a history of suicide attempts. In all, 2,507 SNPs were identified with an evidence for association at p < 0.001. These associated SNPs were subsequently tested for association in a large and independent bipolar sample set. None of these SNPs showed significantly associated in the replication sample after correcting for multiple testing, but the combined analysis of the two sample sets produced an association signal on 2p25 (rs300774) at the threshold of genome-wide significance (p = 5.07 × 10−8). The associated SNPs on 2p25 fall in a large linkage disequilibrium block containing the ACP1 (acid phosphatase 1) gene, whose expression is significantly elevated in bipolar subjects who completed suicide. Furthermore, the ACP1 protein is a tyrosine phosphatase that influences Wnt signalling, a pathway regulated by lithium, making ACP1 a functional candidate for involvement in the phenotype. Mullins et al. (2014) conducted a genome-wide association and meta-analysis. They reported no significant evidence for association at any SNP in the GWAS or meta-analysis. Meanwhile, polygenic score analyses were performed to suicide attempts in the mood disorder cohorts or ideation status in the GENDEP study. Polygenic scores for suicidal behaviour showed no predictive ability for suicidal ideation.
A recent SNP GWAS on severity of suicidal behaviour was published by Zai et al. (2015). This GWAS was conducted in three independent bipolar patients. No genome-wide significant association of any tested markers was found in any of the bipolar samples, although a number of common variants located on the chromosomes 8 and 10 showed suggestive association. A more recent GWAS on suicidal behaviour was performed by Galfalvy et al. (2015). A consortium of US, Canadian, and German teams assembled 2 groups of cases: suicide attempters and suicides (n = 577) and non-attempter psychiatric and healthy controls (n = 1233). The test was repeated separating suicide attempter and completed suicide as outcomes. No SNP reached genome-wide significance but several SNPs within STK3, ADAMTS14, PSME12, and TBX20 genes reached p < 1 × 10−5 level.
In summary, although the results of these GWAS in suicide were all essentially negative, they do suggest interesting candidate genes that may be worthwhile to follow up in future studies. It is becoming increasingly clear that individual’s genetic susceptibility factors for suicide are likely to have only minor effects, and very large pooled analyses of cases and controls will be necessary to identify them.
Conclusions and Future Directions
Suicide is a multifactorial and complex disorder. Apart from biological and psychosocial factors, environmental factors are also critical in predisposition of suicidal behaviour. As discussed above, a large number of genetic studies have been conducted in psychiatric disorder patients dissecting suicidal behaviour. Majority of the studies are focused on serotonergic pathways which is logical given that this neurotransmitter is the most implicated in suicide. Several studies link TPH1 gene with suicide. More recently, TPH2 gene shows promising results only when haplotypic analysis is done. 5-HTTLPR (S (L) allele) is another candidate gene which has shown an association with violent suicide attempts. The results of serotonin receptor subtypes are mixed. Several factors may contribute to conflicting results. As pointed out earlier, some studies are positive in Caucasians but negative in Asian population or vice versa. There are several technical and statistical limitations to the candidate gene approach. This has led to GWAS, which has provided some interesting results, although majority of them are negative. There are several issues with GWAS as discussed elegantly by Ward and Kellis (2012). They have discussed the influence of cumulative predictive power, noncoding variants, detection of rare variants, and reproducibility on GWAS studies. At present, most of the susceptible variants are demonstrated to be located on the intron regions on the chromosome. Thus, it is difficult to explain the function of these identified susceptible variants. Whole exome and whole genome sequencing can detect rare variants and functional mutations. Objectively, whole exome and whole genome sequencing have been applied into the many complex diseases, such as cancer, schizophrenia, autism, etc., however, there is yet to find any report in suicide behaviour. Environmental and/or socioeconomic factors need to be considered carefully that can affect phenotype and assessing noncoding mutation is a challenging task. Even with these limitations, the filed is moving forward and effort needs to be made to form a consortium where a large number of patients and healthy controls can be recruited and examined in a single study.
References
Abbar, M., Courtet, P., Bellivier, F., Leboyer, M., Boulenger, J., Castelhau, D., … Paoloni-Giacobino, A. (2001). Suicide attempts and the tryptophan hydroxylase gene. Molecular Psychiatry, 6(3), 268–273.
Andre, K., Kampman, O., Viikki, M., Illi, A., Setälä-Soikkeli, E., Poutanen, O., & Lehtimaki, T. (2013). TPH1 A218C polymorphism and temperament in major depression. BMC Psychiatry, 13(1), 118.
Anguelova, M., Benkelfat, C., & Turecki, G. (2003). A systematic review of association studies investigating genes coding for serotonin receptors and the serotonin transporter: II. Suicidal behaviour. Molecular Psychiatry, 8(7), 646–653.
Asberg, M. (1997). Neurotransmitters and suicidal behaviour. The evidence from cerebrospinal fluid studies. Annals of the New York Academy of Sciences, 836(1), 158–181.
Bachus, S. E., Hyde, T. M., Akil, M., Weickert, C. S., Vawter, M. P., & Kleinman, J. E. (1997). Neuropathology of suicide. Annals of the New York Academy of Sciences, 836(1), 201–219.
Baud, P., Perroud, N., Courtet, P., Jaussent, I., Relecom, C., Jollant, F., & Malafosse, A. (2009). Modulation of anger control in suicide attempters by TPH 1. Genes, Brain and Behaviour, 8(1), 97–100.
Bellivier, F., Chaste, P., & Malafosse, A. (2004). Association between the TPH gene A218C polymorphism and suicidal behaviour: A meta analysis. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 124(1), 87–91.
Boularand, S., Darmon, M. C., Ravassard, P., & Mallet, J. (1995). Characterization of the human tryptophan hydroxylase gene promoter transcriptional regulation by camp requires a new motif distinct from the camp-responsive element. Journal of Biological Chemistry, 270(8), 3757–3764.
Breidenthal, S. E., White, D. J., & Glatt, C. E. (2004). Identification of genetic variants in the neuronal form of tryptophan hydroxylase (TPH2). Psychiatric Genetics, 14(2), 69–72.
Bresin, K., Sima Finy, M., & Verona, E. (2013). Childhood emotional environment and self-injurious behaviours: The moderating role of the BDNF Val66Met polymorphism. Journal of Affective Disorders, 150(2), 594–600.
Calati, R., Porcelli, S., Giegling, I., Hartmann, A. M., Moller, H. J., De Ronchi, D., & Rujescu, D. (2011). Catechol-o-methyltransferase gene modulation on suicidal behaviour and personality traits: Review, meta-analysis and association study. Journal of Psychiatric Research, 45(3), 309–321.
Celada, P., Puig, M. V., Amargós-Bosch, M., Adell, A., & Artigas, F. (2004). The therapeutic role of 5-HT1A and 5-HT2A receptors in depression. Journal of Psychiatry & Neuroscience, 29(4), 252–265.
Chen, J., Lipska, B. K., Halim, N., Ma, Q. D., Matsumoto, M., Melhem, S., & Weinberger, D. R. (2004). Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): Effects on mRNA, protein, and enzyme activity in postmortem human brain. American Journal of Human Genetics, 75(5), 807–821.
Choong, M. Y., Tee, S. F., & Tang, P. Y. (2014). Meta-analysis of polymorphisms in TPH2 gene and suicidal behaviour. Psychiatry Research, 220(3), 1163–1166.
Cicchetti, D., Rogosch, F. A., & Thibodeau, E. L. (2012). The effects of child maltreatment on early signs of antisocial behaviour: Genetic moderation by tryptophan hydroxylase, serotonin transporter, and monoamine oxidase A genes. Development and Psychopathology, 24, 907–928.
Clayden, R., Zaruk, A., Meyre, D., Thabane, L., & Samaan, Z. (2012). The association of attempted suicide with genetic variants in the SLC6A4 and TPH genes depends on the definition of suicidal behaviour: A systematic review and meta-analysis. Translational Psychiatry, 2(10), e166.
Conner, T. S., Jensen, K. P., Tennen, H., Furneaux, H. M., Kranzler, H. R., & Covault, J. (2010). Functional polymorphisms in the serotonin 1B receptor gene (HTR1B) predict self-reported anger and hostility among young men. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 153(1), 67–78.
Cowen, P. (1991). Serotonin receptor subtypes: implications for psychopharmacology. The British Journal of Psychiatry, 159(12), 7–14.
Du, L., Bakish, D., Lapierre, Y. D., Ravindran, A. V., & Hrdina, P. D. (2000). Association of polymorphism of serotonin 2A receptor gene with suicidal ideation in major depressive disorder. American Journal of Medical Genetics, 96(1), 56–60.
Dwivedi, Y., Rizavi, H. S., Conley, R. R., Roberts, R. C., Tamminga, C. A., & Pandey, G. N. (2003). Altered gene expression of brain-derived neurotrophic factor and receptor tyrosine kinase B in postmortem brain of suicide subjects. Archives of General Psychiatry, 60(8), 804–815.
Egan, M. F., Kojima, M., Callicott, J. H., Goldberg, T. E., Kolachana, B. S., Bertolino, A., & Weinberger, D. R. (2003). The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell, 112(2), 257–269.
Fanous, A. H., Chen, X., Wang, X., Amdur, R., O’Neill, F. A., Walsh, D., & Kendler, K. S. (2009). Genetic variation in the serotonin 2A receptor and suicidal ideation in a sample of 270 Irish high-density schizophrenia families. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 150(3), 411–417.
Galfalvy, H., Haghighi, F., Hodgkinson, C., Goldman, D., Oquendo, M. A., Burke, A., & Mann, J. J. (2015). A genome-wide association study of suicidal behaviour. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 168(7), 557–563.
Galfalvy, H., Huang, Y.-Y., Oquendo, M. A., Currier, D., & Mann, J. J. (2009). Increased risk of suicide attempt in mood disorders and TPH1 genotype. Journal of Affective Disorders, 115(3), 331–338.
Giegling, I., Hartmann, A. M., Möller, H.-J., & Rujescu, D. (2006). Anger-and aggression-related traits are associated with polymorphisms in the 5-HT-2A gene. Journal of Affective Disorders, 96(1), 75–81.
González-Castro, T. B., Juárez-Rojop, I., López-Narváez, M. L., & Tovilla-Zárate, C. A. (2014). Association of TPH-1 and TPH-2 gene polymorphisms with suicidal behaviour: A systematic review and meta-analysis. BMC Psychiatry, 14(1), 196.
Gonzalez-Castro, T. B., Nicolini, H., Lanzagorta, N., Lopez-Narvaez, L., Genis, A., Pool Garcia, S., & Tovilla-Zarate, C. A. (2015). The role of brain-derived neurotrophic factor (BDNF) Val66Met genetic polymorphism in bipolar disorder: A case-control study, comorbidities, and meta-analysis of 16,786 subjects. Bipolar Disorder, 17(1), 27–38.
Hakulinen, C., Jokela, M., Hintsanen, M., Merjonen, P., Pulkki-Råback, L., Seppälä, I., & Viikari, J. (2013). Serotonin receptor 1B genotype and hostility, anger and aggressive behaviour through the lifespan: The Young Finns study. Journal of Behavioural Medicine, 36(6), 583–590.
Holmes, A. (2008). Genetic variation in cortico-amygdala serotonin function and risk for stress-related disease. Neuroscience & Biobehavioural Reviews, 32(7), 1293–1314.
Hong, C. J., Huo, S. J., Yen, F. C., Tung, C. L., Pan, G. M., & Tsai, S. J. (2003). Association study of a brain-derived neurotrophic-factor genetic polymorphism and mood disorders, age of onset and suicidal behaviour. Neuropsychobiology, 48(4), 186–189.
Hong, K. W., Weiss, A., Morimura, N., Udono, T., Hayasaka, I., Humle, T., et al. (2011). Polymorphism of the tryptophan hydroxylase 2 (TPH2) gene is associated with chimpanzee neuroticism. PloS ONE, 6(7), e22144.
Iga, J., Ueno, S., Yamauchi, K., Numata, S., Tayoshi-Shibuya, S., Kinouchi, S., & Ohmori, T. (2007). The Val66Met polymorphism of the brain-derived neurotrophic factor gene is associated with psychotic feature and suicidal behaviour in Japanese major depressive patients. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 144b(8), 1003–1006.
Jensen, K. P., Covault, J., Conner, T. S., Tennen, H., Kranzler, H. R., & Furneaux, H. M. (2009). A common polymorphism in serotonin receptor 1B mRNA moderates regulation by miR-96 and associates with aggressive human behaviours. Molecular Psychiatry, 14(4), 381–389.
Karanović, J., Ivković, M., Jovanović, V. M., Pantović, M., Pavlović-Janković, N., Damjanović, A.,… Savić-Pavićević, D. (2016). Tryptophan hydroxylase 1 variant rs1800532 is associated with suicide attempt in serbian psychiatric patients but does not moderate the effect of recent stressful life events. Suicide and Life-Threatening Behaviour. Ahead of print. doi:10.1111/sltb.12246.
Kia-Keating, B. M., Glatt, S. J., & Tsuang, M. T. (2007). Meta-analyses suggest association between COMT, but not HTR1B, alleles, and suicidal behaviour. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 144(8), 1048–1053.
Kim, B., Kim, C. Y., Hong, J. P., Kim, S. Y., Lee, C., & Joo, Y. H. (2008). Brain-derived neurotrophic factor Val/Met polymorphism and bipolar disorder. Association of the Met allele with suicidal behaviour of bipolar patients. Neuropsychobiology, 58(2), 97–103.
Kohli, M. A., Salyakina, D., Pfennig, A., Lucae, S., Horstmann, S., Menke, A., & Binder, E. B. (2010). Association of genetic variants in the neurotrophic receptor-encoding gene NTRK2 and a lifetime history of suicide attempts in depressed patients. Archives of General Psychiatry, 67(4), 348–359.
Koller, G., Bondy, B., Preuss, U. W., Bottlender, M., & Soyka, M. (2003). No association between a polymorphism in the promoter region of the MAOA gene with antisocial personality traits in alcoholics. Alcohol and Alcoholism, 38(1), 31–34.
Laje, G., Allen, A. S., Akula, N., Manji, H., John Rush, A., & McMahon, F. J. (2009). Genome-wide association study of suicidal ideation emerging during citalopram treatment of depressed outpatients. Pharmacogenetics and Genomics, 19(9), 666–674.
Lalovic, A., & Turecki, G. (2002). Meta-analysis of the association between tryptophan hydroxylase and suicidal behaviour. American Journal of Medical Genetics, 114(5), 533–540.
Laske, C., & Eschweiler, G. (2006). Brain-derived neurotrophic factor. Der Nervenarzt, 77(5), 523–537.
Lee, B. H., Kim, H., Park, S. H., & Kim, Y. K. (2007). Decreased plasma BDNF level in depressive patients. Journal of Affective Disorders, 101(1–3), 239–244.
Lemonde, S., Turecki, G., Bakish, D., Du, L., Hrdina, P. D., Bown, C. D., & Basak, A. (2003). Impaired repression at a 5-hydroxytryptamine 1A receptor gene polymorphism associated with major depression and suicide. The Journal of Neuroscience, 23(25), 8788–8799.
Li, D., Duan, Y., & He, L. (2006). Association study of serotonin 2A receptor (5-HT2A) gene with schizophrenia and suicidal behaviour using systematic meta-analysis. Biochemical and Biophysical Research Communications, 340(3), 1006–1015.
Li, D., & He, L. (2007). Meta-analysis supports association between serotonin transporter (5-HTT) and suicidal behaviour. Molecular Psychiatry, 12(1), 47–54.
Lin, P. Y., & Tsai, G. (2004). Association between serotonin transporter gene promoter polymorphism and suicide: Results of a meta-analysis. Biological Psychiatry, 55(10), 1023–1030.
Liu, X., Li, H., Qin, W., He, G., Li, D., Shen, Y., & He, L. (2006). Association of TPH1 with suicidal behaviour and psychiatric disorders in the Chinese population. Journal of Medical Genetics, 43(2), e04.
Mann, J. J., Malone, K. M., Nielsen, D. A., Goldman, D., Erdos, J., & Gelernter, J. (1997). Possible association of a polymorphism of the tryptophan hydroxylase gene with suicidal behaviour in depressed patients. American Journal of Psychiatry, 154(10), 1451–1453.
Menke, A., Domschke, K., Czamara, D., Klengel, T., Hennings, J., Lucae, S., & Holsboer, F. (2012). Genome-wide association study of antidepressant treatment-emergent suicidal ideation. Neuropsychopharmacology, 37(3), 797–807.
Mittendorfer-Rutz, E., Rasmussen, F., & Wasserman, D. (2008). Familial clustering of suicidal behaviour and psychopathology in young suicide attempters. Social Psychiatry and Psychiatric Epidemiology, 43(1), 28–36.
Mullins, N., Perroud, N., Uher, R., Butler, A. W., Cohen Woods, S., Rivera, M., & Tansey, K. E. (2014). Genetic relationships between suicide attempts, suicidal ideation and major psychiatric disorders: A genome-wide association and polygenic scoring study. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 165(5), 428–437.
Neves, F., Silveira, G., Romano-Silva, M., Malloy Diniz, L., Ferreira, A., De Marco, L., & Correa, H. (2008). Is the 5-HTTLPR polymorphism associated with bipolar disorder or with suicidal behaviour of bipolar disorder patients? American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 147(1), 114–116.
Neves, F. S., Malloy-Diniz, L. F., Romano-Silva, M. A., Aguiar, G. C., de Matos, L. O., & Correa, H. (2010). Is the serotonin transporter polymorphism (5-HTTLPR) a potential marker for suicidal behaviour in bipolar disorder patients? Journal of Affective Disorders, 125(1), 98–102.
Nielsen, D. A., Goldman, D., Virkkunen, M., Tokola, R., Rawlings, R., & Linnoila, M. (1994). Suicidality and 5-hydroxyindoleacetic acid concentration associated with a tryptophan hydroxylase polymorphism. Archives of General Psychiatry, 51(1), 34–38.
Nielsen, D. A., Virkkunen, M., Lappalainen, J., Eggert, M., Brown, G. L., Long, J. C., & Linnoila, M. (1998). A tryptophan hydroxylase gene marker for suicidality and alcoholism. Archives of General Psychiatry, 55(7), 593–602.
Ohtani, M., Shindo, S., & Yoshioka, N. (2004). Polymorphisms of the tryptophan hydroxylase gene and serotonin 1A receptor gene in suicide victims among Japanese. The Tohoku Journal of Experimental Medicine, 202(2), 123–133.
Paik, I.-H., Toh, K.-Y., Kim, J.-J., & Lee, C. (2000). TPH gene may be associated with suicidal behaviour, but not with schizophrenia in the Korean population. Human Heredity, 50(6), 365–369.
Perez-Rodriguez, M. M., Weinstein, S., New, A. S., Bevilacqua, L., Yuan, Q., Zhou, Z., et al. (2010) Tryptophan-hydroxylase 2 haplotype association with borderline personality disorder and aggression in a sample of patients with personality disorders and healthy controls. Journal of Psychiatric Research, 44(15), 1075–1081.
Perlis, R. H., Huang, J., Purcell, S., Fava, M., Rush, A. J., Sullivan, P. F., … Smoller, J. W. (2010). Genome-wide association study of suicide attempts in mood disorder patients. American Journal of Psychiatry, 167(12), 1499–1507.
Perroud, N., Aitchison, K. J., Uher, R., Smith, R., Huezo-Diaz, P., Marusic, A., & Craig, I. (2009). Genetic predictors of increase in suicidal ideation during antidepressant treatment in the GENDEP project. Neuropsychopharmacology, 34(12), 2517–2528.
Perroud, N., Salzmann, A., Saiz, P. A., Baca-Garcia, E., Sarchiapone, M., Garcia-Portilla, M. P., & Malafosse, A. (2010). Rare genotype combination of the serotonin transporter gene associated with treatment response in severe personality disorder. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 153b(8), 1494–1497.
Perroud, N., Uher, R., Ng, M., Guipponi, M., Hauser, J., Henigsberg, N., & Rietschel, M. (2012). Genome-wide association study of increasing suicidal ideation during antidepressant treatment in the GENDEP project. The Pharmacogenomics Journal, 12(1), 68–77.
Pitchot, W., Hansenne, M., & Ansseau, M. (2001a). Role of dopamine in non-depressed patients with a history of suicide attempts. European Psychiatry, 16(7), 424–427.
Pitchot, W., Hansenne, M., Gonzalez Moreno, A., Pinto, E., Reggers, J., Fuchs, S., & Ansseau, M. (2001b). Reduced dopamine function in depressed patients is related to suicidal behaviour but not its lethality. Psychoneuroendocrinology, 26(7), 689–696.
Placidi, G. P., Oquendo, M. A., Malone, K. M., Huang, Y. Y., Ellis, S. P., & Mann, J. J. (2001). Aggressivity, suicide attempts, and depression: relationship to cerebrospinal fluid monoamine metabolite levels. Biological Psychiatry, 50(10), 783–791.
Rivera, A. M., Bermúdez Ocaña, D., Camarena Medellín, B., & Tovilla-Zárate, C. (2012). No association between the HTR1A gene and suicidal behaviour: A meta-analysis. Revista Brasileira de Psiquiatria, 34(1), 38–42.
Ropret, S., Zupanc, T., Komel, R., & Videtic Paska, A. (2015). Investigating the associations between polymorphisms in the NTRK2 and NGFR genes and completed suicide in the Slovenian sample. Psychiatrics Genetics, 25(6), 241–248.
Rotondo, A., Schuebel, K., Bergen, A., Aragon, R., Virkkunen, M., Linnoila, M., … Nielsen, D. (1999). Identification of four variants in the tryptophan hydroxylase promoter and association to behaviour. Molecular Psychiatry, 4(4), 360–368.
Roy, A., Rylander, G., Forslund, K., Asberg, M., Mazzanti, C. M., Goldman, D., & Nielsen, D. A. (2001). Excess tryptophan hydroxylase 17 779C allele in surviving cotwins of monozygotic twin suicide victims. Neuropsychobiology, 43(4), 233–236.
Roy, A., Segal, N. L., Centerwall, B. S., & Robinette, C. D. (1991). Suicide in twins. Archives of General Psychiatry, 48(1), 29–32.
Rujescu, D., Giegling, I., Bondy, B., Gietl, A., Zill, P., & Möller, H. J. (2002). Association of anger-related traits with SNPs in the TPH gene. Molecular Psychiatry, 7(9), 1023–1029.
Rujescu, D., Giegling, I., Sato, T., Hartmann, A. M., & Möller, H.-J. (2003). Genetic variations in tryptophan hydroxylase in suicidal behaviour: Analysis and meta-analysis. Biological Psychiatry, 54(4), 465–473.
Saiz, P. A., García-Portilla, P., Paredes, B., Corcoran, P., Arango, C., Morales, B., & Florez, G. (2011). Role of serotonergic-related systems in suicidal behaviour: Data from a case–control association study. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 35(6), 1518–1524.
Samadi, R. B., Ghasemi, A., Seifi, M., Samadikuchaksaraei, A., Baybordi, F., & Danaei, N. (2012). Serotonin 1A receptor genetic variations, suicide, and life events in the Iranian population. Psychiatry and Clinical Neurosciences, 66(4), 337–343.
Schild, A. H., Pietschnig, J., Tran, U. S., & Voracek, M. (2013). Genetic association studies between SNPs and suicidal behaviour: A meta-analytical field synopsis. Progress in Neuropsychopharmacology and Biological Psychiatry, 46, 36–42.
Schosser, A., Butler, A. W., Ising, M., Perroud, N., Uher, R., Ng, M. Y., & Korszun, A. (2011). Genomewide association scan of suicidal thoughts and behaviour in major depression. PLoS ONE, 6(7), e20690.
Schulsinger, F. K., Kety, S., Rosenthal, D., Rosenthal, D., & Wender, P. H. (1979). A family study of suicide. In M. Schou & E. Stromgren (Eds.), Origin, presentation and treatment of affective disorders (pp. 277–287). London: Academic Press.
Soderstrom, H., Blennow, K., Manhem, A., & Forsman, A. (2001). CSF studies in violent offenders I. 5-HIAA as a negative and HVA as a positive predictor of psychopathy. Journal of Neural Transmission, 108(7), 869–878.
Souery, D., Van Gestel, S., Massat, I., Blairy, S., Adolfsson, R., Blackwood, D., … Mendlewicz, J. (2001). Tryptophan hydroxylase polymorphism and suicidality in unipolar and bipolar affective disorders: a multicenter association study. Biological Psychiatry, 49(5), 405–409.
Statham, D. J., Heath, A. C., Madden, P. A., Bucholz, K. K., Bierut, L., Dinwiddie, S., & Martin, N. (1998). Suicidal behaviour: An epidemiological and genetic study. Psychological Medicine, 28(4), 839–855.
Stefulj, J., Kubat, M., Balija, M., & Jernej, B. (2006). TPH gene polymorphism and aging: indication of combined effect on the predisposition to violent suicide. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 141(2), 139–141.
Sugden, K., Tichopad, A., Khan, N., Craig, I. W., & D’Souza, U. M. (2009). Genes within the serotonergic system are differentially expressed in human brain. BMC Neuroscience, 10(1), 1.
Tovilla-Zárate, C., Juárez-Rojop, I., Ramón-Frias, T., Villar-Soto, M., Pool-García, S., Medellín, B. C., & Humberto, N. (2011). No association between COMT val158met polymorphism and suicidal behaviour: Meta-analysis and new data. BMC Psychiatry, 11(1), 151.
Tsai, S.-J., Hong, C.-J., & Wang, Y.-C. (1999). Tryptophan hydroxylase gene polymorphism (A218C) and suicidal behaviours. NeuroReport, 10(18), 3773–3775.
Turecki, G., Zhu, Z., Tzenova, J., Lesage, A., Seguin, M., Tousignant, M., et al. (2001). TPH and suicidal behaviour: A study in suicide completers. Molecular Psychiatry, 6(1), 98–102.
Vaquero-Lorenzo, C., Baca-Garcia, E., Diaz Hernandez, M., Perez-Rodriguez, M. M., Fernandez-Navarro, P., Giner, L., … Oquendo, M. A. (2008). Association study of two polymorphisms of the serotonin 2A receptor gene and suicide attempts. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 147(5), 645–649.
Vincze, I., Perroud, N., Buresi, C., Baud, P., Bellivier, F., Etain, B., … Malafosse, A. (2008). Association between brain-derived neurotrophic factor gene and a severe form of bipolar disorder, but no interaction with the serotonin transporter gene. Bipolar Disorders, 10(5), 580–587.
Virkkunen, M., De Jong, J., Bartko, J., & Linnoila, M. (1989). Psychobiological concomitants of history of suicide attempts among violent offenders and impulsive fire setters. Archives of General Psychiatry, 46(7), 604–606.
Von Borczyskowski, A., Lindblad, F., Vinnerljung, B., Reintjes, R., & Hjern, A. (2011). Familial factors and suicide: an adoption study in a Swedish National Cohort. Psychological Medicine, 41(4), 749–758.
Voracek, M., & Loibl, L. M. (2007). Genetics of suicide: A systematic review of twin studies. Wiener Klinische Wochenschrift, 119(15–16), 463–475.
Walther, D. J., Peter, J.-U., Bashammakh, S., Hörtnagl, H., Voits, M., Fink, H., & Bader, M. (2003). Synthesis of serotonin by a second tryptophan hydroxylase isoform. Science, 299(5603), 76.
Ward, L. D., & Kellis, M. (2012). Interpreting noncoding genetic variation in complex traits and human disease. Nature Biotechnology, 30(11), 1095–1106.
Wasserman, D., Geijer, T., Sokolowski, M., Frisch, A., Michaelovsky, E., Weizman, A., & Wasserman, J. (2007). Association of the serotonin transporter promotor polymorphism with suicide attempters with a high medical damage. European Neuropsychopharmacology, 17(3), 230–233.
Willour, V. L., Seifuddin, F., Mahon, P. B., Jancic, D., Pirooznia, M., Steele, J., … MacKinnon, D. F. (2012). A genome-wide association study of attempted suicide. Molecular Psychiatry, 17(4), 433–444.
Wrzosek, M., Łukaszkiewicz, J., Wrzosek, M., Serafin, P., Jakubczyk, A., Klimkiewicz, A., … Wojnar, M. (2011). Association of polymorphisms in HTR2A, HTR1A and TPH2 genes with suicide attempts in alcohol dependence: A preliminary report. Psychiatry Research, 190(1), 149–151.
Zai, C. C., Gonçalves, V. F., Tiwari, A. K., Gagliano, S. A., Hosang, G., De Luca, V., … Kennedy, J. L. (2015). A genome-wide association study of suicide severity scores in bipolar disorder. Journal of Psychiatric Research, 65, 23–29.
Zai, C. C., Manchia, M., De Luca, V., Tiwari, A. K., Chowdhury, N. I., Zai, G. C., … Kennedy, J. L. (2012). The brain-derived neurotrophic factor gene in suicidal behaviour: A meta-analysis. International Journal of Neuropsychopharmacology, 15(8), 1037–1042.
Zarrilli, F., Angiolillo, A., Castaldo, G., Chiariotti, L., Keller, S., Sacchetti, S., … Sarchiapone, M. (2009). Brain derived neurotrophic factor (BDNF) genetic polymorphism (Val66Met) in suicide: A study of 512 cases. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 150b(4), 599–600.
Zill, P., Büttner, A., Eisenmenger, W., Möller, H.-J., Bondy, B., & Ackenheil, M. (2004). Single nucleotide polymorphism and haplotype analysis of a novel tryptophan hydroxylase isoform (TPH2) gene in suicide victims. Biological Psychiatry, 56(8), 581–586.
Acknowledgements
This study was partially supported by funding from National Institutes of Mental Health (R01MH082802; 1R01MH101890; R01MH100616; 1R01MH107183-01), American Foundation for Suicide Prevention (SRG-1-042-14) to Dr. Yogesh Dwivedi.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer Nature Singapore Pte Ltd.
About this chapter
Cite this chapter
Wang, Q., Dwivedi, Y. (2017). Genetics of Suicidal Behaviour. In: Kumar, U. (eds) Handbook of Suicidal Behaviour. Springer, Singapore. https://doi.org/10.1007/978-981-10-4816-6_4
Download citation
DOI: https://doi.org/10.1007/978-981-10-4816-6_4
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-10-4815-9
Online ISBN: 978-981-10-4816-6
eBook Packages: Behavioral Science and PsychologyBehavioral Science and Psychology (R0)