
Abstract
The northern and eastern plains of Cambodia support the largest extent of lowland deciduous forest remaining in Southeast Asia. This landscape has also been identified as the highest priority site for tiger Panthera tigris recovery in Indochina. We estimated ungulate tiger prey densities using distance-based line transect sampling from two protected areas in the Eastern Plains Landscape between 2009 and 2011. Densities for large ungulates ranged from 1.1 ± SE 0.2 individuals/km2 for banteng Bos javanicus to 2.2 ± SE 0.2 individuals km2 for red muntjac Muntiacus muntjak. The ungulate activity patterns were correlated with activity patterns of extant large carnivores in the landscape with leopard Panthera pardus and dhole Cuon alpinus showing substantial activity pattern overlap with wild pig Sus scrofa and red muntjac, respectively. Overall tiger prey biomass was more than 540 kg/km2 of which the endangered banteng comprised greater than 80 %. However, ungulate densities were much lower than in ecologically similar sites in South Asia. This was mainly due to the absence of large deer species like Cervus deer, which have historically been extensively hunted. Nevertheless, the Eastern Plains landscape likely supports 50–60 % of the global banteng population and remains a high priority area for the conservation of large herbivores in Southeast Asia.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
9.1 Introduction
The northern and eastern Cambodian provinces of Mondulkiri, Ratanakiri, Stung Treng, and Preah Vihear support some of the largest extents of lowland deciduous forest in Southeast Asia (Todorff et al. 2005). These forests had once been described as one of the “great game-lands of the world; a Serengeti of Asia” and had historically supported a diverse and abundant megafaunal assemblage of ungulates, predators and scavengers (Wharton 1957; Todorff et al. 2005). The ungulate component included at one time an assemblage of four sympatric wild cattle species ( gaur Bos gaurus, banteng Bos javanicus , kouprey Bos sauvelli and wild water buffalo Bubalus arnee); a unique distinction as few regions in the world have supported more than two sympatric wild cattle species.
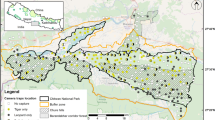
The Eastern Plains Landscape (EPL), Mondulkiri province, in eastern Cambodia (Fig. 9.1) forms part of the Lower Mekong Dry Forest Ecoregion (Olson and Dinerstein 1998; Tordoff et al. 2005). This landscape of rolling lowlands, generally under 150 m asl, is characterized by extensive deciduous dipterocarp forest, including open grassland areas (veal) with a high frequency fire regime, and smaller patches of mixed deciduous and semi-evergreen forests on higher ground and along watercourses. The EPL deciduous forest is bisected by a number of rivers including one major tributary of the Mekong, the Srepok River, and is studded with small seasonal wetlands (trapeang).

Protected areas within the Eastern Plains Landscape Cambodia and Vietnam
The wider landscape in which the EPL is situated suffered considerable political instability and conflict throughout the latter half of the 20th century, which intensified during the Lon Nol (1970–75) and Pol Pot (1975–79) regimes, and persisted into the late 1990s. During this time there was evidence of large declines in the population and distribution of large mammal species including tiger Panthera tigris, leopard P. pardus, Asian elephant Elephas maximus, banteng, gaur and Eld’s deer Cervus eldii (Loucks et al. 2008). These declines were associated with a proliferation of firearms, the development of an external market for wildlife products and, particularly during the Khmer Rouge era, government sponsored hunting (Loucks et al. 2008). This hunting pressure led to the global extinction of one large herbivore species endemic to Indochina, the kouprey (Timmins et al. 2008a, b; Timmins 2011), and more recently the extinction of the tiger from the landscape due to targeted poaching of the remaining individuals (Gray et al. 2012a; O’Kelly et al. 2012).
Almost four–fifths of the EPL (13,730 km2) lie within protected areas; four wildlife sanctuaries managed by the General Department for Administration of Nature Conservation and Protection of the Ministry of Environment, and three protection forests managed by the Forestry Administration of the Ministry of Agriculture, Forestry and Fisheries (Table 9.1). Furthermore, two areas in Vietnam adjacent to the EPL are gazetted as national parks; Bu Gia Map (12.7°N 107.1°E; 260 km2), South of Seima Protection Forest, and Yok Don (12.5°N 107.4°E; 1,155 km2), the largest national park in Vietnam , to the east of Mondulkiri Protected Forest (13.0°N 107.3°E). With improved security resulting from a stable political climate the EPL region has, since the early 2000s, been the focus of conservation activity by government conservation departments supported by international nongovernmental organisations. Since 2002, the World Wide Fund for Nature (WWF) Greater Mekong Program has assisted the Royal Cambodian Government with protected area management activities within two protected areas in the Eastern Plains Landscape: Mondulkiri Protected Forest (MPF) and Phnom Prich Wildlife Sanctuary (PPWS) (Fig. 9.1). These protected areas have received approximately $9,000,000 in international conservation investment during this period. This investment has supported protected area infrastructure development, the recruitment and training of ranger enforcement patrols, biological monitoring, improved judiciary response, and alternative livelihood work with communities adjacent to the protected area.
Mondulkiri Protected Forest (MPF) and Phnom Prich Wildlife Sanctuary (PPWS) cover approximately 7,000 km2 in the heart of the Eastern Plains Landscape and comprise a mosaic of deciduous dipterocarp, mixed deciduous, and semi-evergreen forest (Table 9.1). The two protected areas support at least 19 critically endangered or endangered mammal, bird, and reptile species (Table 9.2). Since 2008 we have been studying large herbivore and predator communities within the core areas of MPF and PPWS (Phan and Gray 2010; Gray and Phan 2011; Gray and Prum 2012; Gray 2012; Gray et al. 2012b, 2013; Gray et al. 2014), largely though distance-based line transect sampling and camera trapping.
9.2 Large Herbivore Densities and Their Conservation Significance in Cambodia’s Deciduous Dipterocarp Forests
We surveyed 110 line transects that were 1–4 km long and randomly distributed within the 3,400 km2 core areas of Mondulkiri Protected Forest and Phnom Prich Wildlife Sanctuary during the dry seasons of 2009/10 and 2010/11. Surveys followed the protocols of Karanth and Nichols (2002) for line transect sampling of large herbivores. Total survey effort was 1,310 km resulting in 325 encounters with large herbivores (Table 9.3). We used the conventional distance sampling (CDS) engine in software DISTANCE 6.0 (Thomas et al. 2010) to estimate densities following the protocols of Thomas et al. (2010). Despite the extensive survey effort the numbers of encounters of three large ungulate species ( gaur , sambar Cervus unicolor, and Eld’s deer ) were too low for density estimation.
Eastern Plains Landscape wide densities for large ungulates ranged from 1.1 ± SE 0.2 individuals/km2 for banteng to 2.2 ± SE 0.2 individuals/km2 for red muntjac Muntiacus muntjak (Table 9.4). Using estimated species weights from Karanth and Sunquist (1992), the densities suggest a prey biomass of approximately 540 kg/km2, of which banteng comprise more than 80 %. Estimated population sizes of species across the entire study area were 2700–5700 banteng, 6000–9000 red muntjac, and 3000–8000 wild pig Sus scrofa (Table 9.4).
As far as we are aware the only other site with similarly robust estimates for ungulates in Indochina (sensu Laos , Cambodia, and Vietnam ) is the adjacent Seima Protected Forest. In 2010 density estimates , from the 1,800 km2 core area of Seima, based on a 1,600 km survey effort of 40 transects lines, were 0.29 wild cattle (both banteng and gaur )/km2, 1.8 red muntjac /km2, 2.0 wild pig /km2, and 0.1 sambar /km2 (O’Kelly and Nut 2010). This corresponds to an overall large ungulate density of 4.2 individuals/km2 providing a biomass of approximately 260 kg/km2.
A number of other studies have attempted to quantify tiger prey densities and biomass, largely in semi-evergreen and evergreen forests, in Southeast Asia . However, only a few of these studies employed the scientifically robust distance-based line transect sampling methodology that had been used to survey the Eastern Plains Landscape (O’Brien et al. 2003; WCS-Thailand 2008). For example, in the mosaic of semi-evergreen, mixed deciduous, and deciduous dipterocarp forests in Huai Kha Khaeng Wildlife Sanctuary, western Thailand , overall large ungulate densities are estimated at 6.2 individuals per km2 (2008 data based on 45+ line transects and 1,000 km+ survey effort; WCS-Thailand 2008). Red muntjac was the most abundant species (2.2 individuals/km2) followed by sambar (1.9 individuals/km2) and wild pig (1.6 individuals/km2). Wild cattle densities were estimated at 0.2 banteng/km2 and 0.4 gaur/km2 (WCS-Thailand 2008). Steinmetz and Mather (1996) estimated Muntiacus spp. density in semi-evergreen and mixed deciduous forest in Thung Yai Naresuan Wildlife Sanctuary, western Thailand, to be 1–2 individuals/km2 based on 360 km of line transect sampling.
Large ungulate density estimates have also been published from lowland rainforest in Bukit Barisan National Park, Sumatra where sambar density was 0.6–1.4 individuals/km2, red muntjac 1.8–4.4 individuals/km2, and wild pig 4.4–6.0 individuals/km2 (O’Brien et al. 2003). Srikosamatara (1993) estimated densities and biomass of wild cattle , sambar , and red muntjac from Huai Kha Khaeng Wildlife Sanctuary based on distance sampling of droppings from line transects calibrated against defecation and decomposition rates. Density estimates were approximately 1.8 wild cattle/km2, 1.9–4.2 sambar/km2, and 3.1 red muntjac/km2 giving an overall biomass of 1,250 kg/km2. By extrapolating of camera-trap encounter rates, Kawanishi and Sunquist (2004) estimated densities of wild pig (3.4–4.6 individuals/km2) and red muntjac (3.2–4.1 individuals/km2), which provided a biomass of 200–400 kg/km2 of tiger prey species in Taman Negara, peninsular Malaysia . Using the Royle-Nichols occupancy heterogeneity model, Vongkhamheng (2011) estimated an ungulate density of 5.3 individuals/km2 (primarily Muntiacus spp. and wild pig with a few sambar ) in largely evergreen forest in Nam Et-Phou Louey NPA, northern Laos . However, calculating abundance from such occupancy models based on sign-encounter rates is dependent upon a large number of assumptions and is unlikely to be as robust, or accurate, as estimates generated from distance-based line transect sampling. In contrast to the paucity of reliable data from South–East Asia, there is, however, a wealth of robust data on large ungulate densities from South Asia, where published densities, based on distance-based line transect sampling, in protected areas range from 7 (Wang 2010) to 250+ individuals per km2 (Wegge and Storaas 2000) with 50+ individual ungulates per km2 being the norm in most Indian tiger reserves (Karanth and Nichols 2000). The EPL estimate of <5 individuals per km2 is clearly at the low end of the ungulate density spectrum for deciduous dipterocarp forests ; for example, in ecologically similar lowland sal deciduous forest in Ranthambore Tiger Reserve ungulate density is approximately 75 animals/km2 (Bagchi et al. 2003). Despite this disparity densities of smaller ungulates (red muntjac and wild pig) within the Eastern Plains Landscape are similar to many South and Southeast Asian protected areas (Table 9.5). Steinmetz et al. (2010) found that in areas with minimal hunting in western Thailand , muntjac densities were generally 1–3 individuals/km2.
Although wild pig have been recorded to reach densities of 40+ individuals per km2 following mass seeding of dipterocarps in lowland evergreen dipterocarp forest in peninsular Malaysia (Ickes 2001), densities of 1–4 individuals/km2 are typical throughout the species’ cosmopolitan world range (Melis et al. 2006). Although O’Kelly and Nut (2010) suggested red muntjac and wild pig densities are lower in Seima Protected Forest than would be expected in sites without hunting , we believe that populations of red muntjac and wild pig are relatively healthy across the Eastern Plains Landscape. Although there was evidence that both species were still being hunted in all protected areas surveyed, the density estimates of both species from the Eastern Plains Landscape were similar to those from ecologically similar protected areas in South and Southeast Asia (Table 9.5), suggesting that both species may be fairly resilient to current levels of hunting pressure in the landscape.
The shortfall in prey densities within the Eastern Plains Landscape in comparison with ecologically similar sites in South Asia and Southeast Asia appears to be mainly due to the low abundances of larger (>45 kg) deer species, like sambar and Eld’s deer (Table 9.5). This is worrying as large cervids make up more than three-quarters of prey consumed by tiger across most of its range (Karanth and Nichols 2002). The reasons for the low densities of sambar and Eld’s deer in the Eastern Plains are unclear, but similar patterns of low densities and slow recoveries of Cervus deer, even when other ungulate species are increasing, have been noted elsewhere in Southeast Asia. For example, in Thung Yai Wildlife Sanctuary, western Thailand , Steinmetz et al. (2010) suggested that the sambar’s mating system , in which females select for prime males, made the sambar less resilient to recovery following hunting when compared to other ungulates (such as red muntjac , wild pig and gaur ) when prime sambar males had been selectively targeted as hunting trophies. Similarly Aung et al. (2001), from a radio tracking study of Eld’s deer in deciduous dipterocarp forest in Myanmar , suggested that the species’ social organization and life history traits differed from other tropical cervids. Eld deer showed tightly synchronized seasonal breeding and low reproductive output by females, which could contribute to low levels of population recovery following hunting. Given the importance of large cervids in tiger diets , additional research into natural history and the role of behavior in these species’ recovery patterns is essential.
9.3 Significance of Banteng Population
The Eastern Plains Landscape supports two globally endangered large ungulates: banteng and Eld’s deer . While we obtained insufficient observations for density estimation of Eld’s deer, our data, also published in Gray et al. (2012b), provides the first robust density and population estimate of banteng from anywhere within the species’ global range. This suggests that Mondulkiri Protected Forest and Phnom Prich Wildlife Sanctuary support the majority of the global population of banteng, and that previous estimates of the species’ Cambodian and global population sizes are low. The IUCN Red List suggests the global population of banteng is ‘unlikely to be more than 8,000 and is quite possibly fewer than 5,000 animals…no subpopulation is believed to exceed 500 individuals and only 6–8 subpopulations of more than 50 animals, are known’ (Timmins et al. 2008a, 2008b). If our estimate of 2,700–5,700 banteng (with a mean of 3,900 individuals) in Mondulkiri Protected Forest and Phnom Prich Wildlife Sanctuary is accurate, then the IUCN information would need to be substantially revised. This study and that of O’Kelly and Nut (2010) in the adjacent Siema Proteced Forest, which suggested a population 100–700 (mean 250) individuals in Siema, are the only banteng population estimates we could find to be derived from robust sampling methodologies accounting for imperfect detection. A review of our data indicates that the assumptions of distance sampling were largely met, and therefore we strongly believe that there is little reason to doubt the accuracy of our estimates. Indeed, the majority of sampling errors which are likely to affect distance sampling of large ungulates, for example evasive movement away from transect lines prior to detection and failing to observe all individuals in groups, will lead to underestimating rather than overestimating density (Wegge and Storaas 2000).
Assessing the global significance of the banteng population in the Eastern Plains Landscape is hindered by the lack of any accurate, scientific population estimates from elsewhere in the species’ range. Table 9.6 summarizes the species’ status from other range countries, though numbers for Myanmar , Laos , Borneo , and the remainder of Cambodia are essentially optimistic estimates of maximum populations based on limited fieldwork. A better global population estimate, therefore, might be 5,600–11,000 individuals, though there is great uncertainty in the population estimates besides those from the Eastern Plains Landscape. We strongly recommend robust surveying for Banteng elsewhere in the species’ range, particularly at sites such as Huai Kha Khaeng Wildlife Sanctuary, Thailand (estimated population 290 individuals; Srikosamatara and Suteethorn 1995), and Ujong Kulon (estimated population 300–800 individuals; Pudyatmoko 2004) and Baluran (estimated population 206 individuals; Pudyatmoko 2004) National Parks, Java , where distance-based line transect surveys appear practical. As in our study areas it is possible that these may result in substantial upwards revisions of known population estimates.
Considering the maximum possible population sizes elsewhere in the species’ range, the protected area complex of Mondulkiri Protected Forest -Phnom Prich Wildlife Sanctuary -Siema Protected Forest supports 50–60 % (upper and lower 95 % confidence intervals respectively) of the global banteng population (Table 9.6). These protected areas are therefore clearly the global stronghold and are irreplaceable for the conservation of this enigmatic and beautiful species. Given the presence of banteng in at least three other protected areas in eastern Cambodia (Lumphat Wildlife Sanctuary and O’Yadao Protected Forest) and adjacent Vietnam (Yok Don National Park—population estimate 30–44 individuals; Pedrono et al. 2009), it is possible that the banteng population in EPL may be even higher than our estimates. However hunting , for wild meat and trophy horns, remains a major threat in these protected areas where enforcement levels are much lower than in our study areas.
9.4 Large Herbivore Activity Patterns and Interactions with Carnivores
Understanding the control that extant large predator populations exert on large herbivore populations is important for conservation management focussed at reviving populations of both threatened large herbivores and their predators . The deciduous dipterocarp forests of the Eastern Plains Landscape, similar to much of tropical Asia , historically supported two Panthera species (tiger and leopard ) together with dhole Cuon alpinus and a suite of smaller cat species (Gray et al. 2014). Where the three species co-occur today, inter-specific competition appears to affect behavior, movement patterns , and prey selection, particularly where large ungulate prey densities are reduced (Stoen and Wegge 1996; Karanth and Sunquist 2000; Odden et al. 2010). The tiger, however, is most likely extinct in the landscape (O’Kelly et al. 2012), but the leopard remains relatively abundant with an estimated density of 3.8 (±SE 1.9) individuals/100 km2 (Gray and Prum 2012), which is comparable with estimates in well protected national parks in Thailand .
We wanted to test, like foraging theory suggests, if whether carnivores synchronize their activity patterns with those of their principal prey species to optimize foraging behavior (Stephens and Krebs 1987; Linkie and Ridout 2011). We, therefore, deployed 50 camera-trap pairs (Reconyx RapidFire Professional PC90; Reconyx, Inc., Holmen, WI) within approximately 210 km2 of the core area of Mondulkiri Protected Forest during the late dry season (March–July 2009) for a total of 3,711 camera-trap pair nights (mean 77.5 per location). Camera-traps were located either side of routes (i.e., motorbike trails, dry-river beds, and ridgelines) designed to maximize encounters with large carnivores (for more details of camera-trap study design see Gray and Prum 2012). All independent encounters (defined as successive photographs separated by >20 min; Phan et al. 2010) of leopard, dhole, and large herbivores were recorded.
Overlap between the activity patterns of leopard , dhole, and the two most frequently photographed large herbivores (red muntjac and wild pig ) were assessed using a statistical model developed by Ridout and Linkie (2009). The Ridout and Linke (2009) model calculates probability density functions of each species’ activity pattern using kernel density estimates (Taylor 2008). Overlap between the density distributions of any two species is estimated using the coefficient of overlapping Δ4 (Weitzman 1970), which ranges from 0 (no overlap) to 1 (complete overlap; Ridout and Linkie 2009). Linkie and Ridout (2011) suggested that Δ4 > 0.8 indicated strongly overlapping activity patterns.
The range of independent encounters of the four target species captured by the camera-traps was between 33 (dhole ) and 442 (red muntjac ) (Table 9.7). Although active throughout the day and night (i.e., cathemeral activity pattern), leopard displayed clear crepuscular activity with 26 % of encounters between 0401–0659 h and 19 % between 1701–1959 h. Dhole were predominantly diurnal (Table 9.7), and wild pig and red muntjac were cathemeral with encounters throughout the 24-h cycle. However, wild pig was distinctly more nocturnal than red muntjac (Table 9.7); red muntjac activity peaked after dawn with 36 % of encounters between 0601–0859 h. The activity pattern of leopard overlapped with wild pig more than it did with red muntjac, while the activity pattern of dhole overlapped more with red muntjac than it did with wild pig (Figs. 9.2 and 9.3).

Activity overlap between leopard and red muntjac (top), and leopard and wild pig (below) based on camera-trapping in Mondulkiri Protected Forest, Cambodia. Δ4, a measure of activity overlap, was calculated based on Linkie and Ridout (2011); Δ4 > 0.8 indicates strongly overlapping activity patterns

Activity overlap between dhole and red muntjac (top) and dhole and wild pig (below) based on camera-trapping in Mondulkiri Protected Forest, Cambodia. Δ4, a measure of activity overlap, was calculated based on Linkie and Ridout (2011); Δ4 > 0.8 indicates strongly overlapping activity patterns
The close temporal overlap between the daily activities of leopard and wild pig, and between dhole and red muntjac suggest that there may be specialization in prey selection by the two large carnivores in Mondulkiri Protected Forest. Although considered a generalist predator, a detailed meta-analysis found that leopard preferentially prey on ungulate species that have a body mass of 10–40 kg and occur in small herds in dense forest habitats (Hayward et al. 2006). However, inter-specific competition with tiger may result in leopard targeting suboptimal prey, for example langurs Semnopithecus spp., particularly when ungulate densities are reduced (Ramakrishnan et al. 1999; Karanth and Sunquist 2000; Steinmetz et al. 2013). In sites such as Mondulkiri Protected Forest, where tigers are absent, wild pig may be an ideal prey for leopard given their relative abundance (approximate density 1.9 individuals per km2), optimal body mass (~38 kg; Karanth and Sunquist 1992), and their herding behavior.
Dhole preferentially prey upon medium to large cervids in South Asia, particularly chital and sambar , and it has been suggested that the primarily diurnal activity patterns of the dhole reflects those of its prey (Karanth and Sunquist 2000; Andheria et al. 2007; Borah et al. 2009). However, in tropical Southeast Asia , where ungulate densities are reduced, muntjac spp., and even mouse-deer Tragulus spp., predominate in dhole scat analysis (Grassman et al. 2005; Kawanishi and Sunquist 2008; Kamler et al. 2012; Steinmetz et al. 2013). The strong temporal overlap found in this study between the activity patterns of the muntjac and dhole in Mondulkiri Protected Forest provides further support for the importance of muntjac in dhole diets in Southeast Asia. Sambar, which dominates dhole diet in other sites, occurs at very low densities in Mondulkiri Protected Forest (4 camera-trap encounters during this study) probably due to hunting pressure. Recovery of sambar populations, therefore, appears to be important for large carnivore conservation in Mondulkiri Protected Forest. Diet studies, involving scat analysis, are recommended for further understanding carnivore–prey interactions in eastern Cambodia.
9.5 Conclusions
The data that our co-workers and us have presented over the past 6 years (Phan and Gray 2010; Gray and Phan 2011; Clements et al. 2012; Gray 2012; Gray and Prum 2012; Gray et al. 2012b; O’Kelly et al. 2012; Wright et al. 2012; Gray et al. 2013) have confirmed the global significance of Mondulkiri Protected Forest, Seima Protection Forest, and Phnom Prich Wildlife Sanctuary for biodiversity conservation . This particularly applies for the long-term conservation of large herbivores in the Southeast Asian region. The documented banteng population is substantially higher than previously estimated and is likely to represent >50 % of the global population of this endangered species. However overall large ungulate densities, <5 individuals per km2, are much lower than the intrinsic carrying capacity of deciduous dipterocarp forest. This appears to be largely due to the scarcity of large deer (sambar and Eld’s deer ).
The landscape goals of WWF Greater Mekong are to recover large ungulate, and thus tiger and other carnivore populations. However, for this to happen, stronger protected area management and a commitment to conservation from the highest levels in the Cambodian government are necessary. All protected areas within the EPL are severely threatened by social and agricultural land concessions and infrastructure projects. While red muntjac and wild pig may be resilient to current levels of hunting and the high population of banteng is encouraging, the low densities of Cervus deer, the high potential for the banteng population to decline, and the high levels of widespread poaching and hunting are major causes for concern. All these factors would need to be addressed for the EPL to be once again the ‘Serengeti of Asia ’.
References
Aung M, McShea WJ, Htung S et al (2001) Ecology and social organization of a tropical deer (Cervus eldi thamin). J Mammal 82:836–847
Andheria AP, Karanth KU, Kumar NS (2007) Diet and prey profiles of three sympatric large carnivores in Bandipur Tiger Reserve. India. J. Zool. 273:169–175
Bagchi S, Goyal SP, Sankar K (2003) Prey abundance and prey selection by tigers (Panthera tigris) in a semi-arid, dry deciduous forest in western India. J Zol Soc Lon 260:285–290
Borah JK, Deka S, Gupta RP (2009) Food habits of dhole (Cuon alpinus) in Satpura Tiger Reserve, Madhya Pradesh, India. Mammalia 73:85–88
Clements T, Gilbert M, Rainey HJ et al. (2012) Vultures in Cambodia: populations, threats and conservation. Bird Cons Int. doi:10.1017/S0959270912000093
Grassman LI, Tewes ME, Silvy NJ et al (2005) Spatial ecology and diet of the dhole Cuon alpinus (Canidae, Carnivora) in North Central Thailand. Mammalia 69:11–20
Gray TNE (2012) Studying large mammals with imperfect detection: status and habitat preferences of wild cattle and large carnivores in eastern Cambodia. Biotropica 44:531–536
Gray TNE, Prum S (2012) Leopard density in post-conflict Cambodia: evidence from spatially explicit capture recapture. J Wil Man 76:163–169
Gray TNE, Phan C (2011) Habitat preferences and activity patterns of the larger mammal community in Phnom Prich Wildlife Sanctuary, Cambodia. R Bull Zool 59:311–318
Gray TNE, Ou R, Huy K et al (2012a) The status of large mammals in eastern Cambodia: a review of camera-trapping data 1999-2007. Cam J Nat His 2012:42–55
Gray TNE, Prum S, Pin C et al (2012b) Distance sampling reveals Cambodia’s Eastern plains landscape supports largest global population of the endangered banteng Bos javanicus. Oryx 46:563–566
Gray TNE, Phan C, Pin C et al (2013) Establishing a monitoring baseline for threatened large ungulates in eastern Cambodia. Wil Biol 18:406–413
Gray TNE, Phan C, Pin C et al. (2014) The status of jungle cat and sympatric small cats in Cambodia’s Eastern Plains Landscape. Cat News Special Issue 8:19-23
Harihar A, Pandav B, Goyal SP (2008) Responses of tiger (Panthera tigris) and their prey to removal of anthropogenic influences in Rajaji National Park, India. Eur J Wil Res 55:97–105
Hayward MW, Henschel P, O’Brien J et al (2006) Prey preferences of the leopard (Panthera pardus). J Zool 270:298–313
Ickes K (2001) Hyper-abundance of Native Wild Pigs (Sus Scorfa) in a lowland dipterocarp rain forest in Penisular Malaysia. Biotropica 33:682–690
Jathanna D, Karanth KU, Johnsingh AJT (2003) Estimation of large herbivore densities in the tropical forests of southern India using distance sampling. J of the Zol Soc Lon 261:285–290
Kamler JF, Johnson A, Vongkhamheng C et al (2012) The diet, prey selection, and activity of dholes (Cuon alpinus) in northern Laos. J Mammal 93:627–633
Karanth KU, Nichols JD (2000) Ecological status and conservation of tigers in India. In: Final technical report to the Division of International Conservation, US Fish and Wildlife Service, Washington DC and Wildlife Conservation Society, New York. Centre for Wildlife Studies, Bangalore, India
Karanth KU, Nichols JD (eds) (2002) Monitoring tigers and their prey: a manual for researchers, managers and conservationistsin tropical Asia. Centre for Wildlife Studies, Bangalore
Karanth KU, Sunquist ME (1992) Population structure, density and biomass of large herbivores in the tropical forests of Nagarhole. India J Trop Trop Ecol 8:21–35
Karanth KU, Sunquist ME (2000) Behavioral correlates of predation by tiger (Panthera tigris), leopard (Panthera pardus) and dhole (Cuon alpinus) in Nagarahole. India J Zool 250:255–265
Kawanishi K, Sunquist ME (2004). Conservation status of tigers in a primary rainforest of Peninsular Malaysia. Biol Cons 120:329–344
Kawanishi K, Sunquist ME (2008) Food habitats and activity patterns of the Asiatic Golden Cat (Catopuma temminckii) and Dhole (Cuon alpinus) in a primary rainforest of Penisular Malaysia. Mam St 33:173–177
Linkie M, Ridout MS (2011) Assessing tiger-prey interactions in Sumatran rainforests. J Zool 284:224–229
Loucks L, Mascia MB, Maxwell A et al (2008) Wildlife decline in Cambodia, 1953–2005: exploring the legacy of armed conflict. Con Let 2:82–92
Melis C, Szafranska PA, Jedrzejewska B et al (2006) Biogeographical variation in the population density of Wild Boar in western Eurasia. J Biogeog 33:803–811
O’Brien TG, Kinnaird MF, Wibisono HT (2003) Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Anim Cons 6:131–139
O’Kelly H, Nut MH (2010) Monitoring of ungulate, primate and peafowl populations using line transect surveys in Seima Protection Forest, Cambodia 2005–2010. Wildlife Conservation Society Cambodia, Phnom Penh
O’Kelly HJ, Evans TDE, Stokes EJ et al (2012) Identify conservation successes, failures and future opportunities; assessing recovery potential of wild ungulates and tiger in eastern Cambodia. PLoS ONE 7(10):e40482. doi:10.1371/journal.pone.0040482
Odden M, Wegge P, Fredriksen T (2010) Do tigers displace leopards? If so, why? Ecol Res 25:875–881
Olson DM, Dinerstein E (1998) The Global 200: a representation approach to conserving the Earth’s most biologically valuable ecoregions. Cons Biol 12:502–515
Pedrono M, Ha MT, Chouteau P et al (2009) Status and distribution of the endangered banteng Bos javanicus birmanicus in Vietnam: a conservation tragedy. Oryx 43:618–625
Phan C, Gray TNE (2010) Ecology and natural history of Banteng in eastern Cambodia: evidence from camera-trapping in Mondulkiri Protected Forest and Phnom Prich Wildlife Sanctuary. Cam J Nat His 2:118–126
Phan C, Prum S, Gray TNE (2010) Recent camera-trap records of globally threatened species from the Eastern Plains Landscape, Cambodia. Cam J Nat His 2:89–93
Pudyatmoko S (2004) Does the banteng (Bos javanicus) have a future in Java? Challenges of the conservation of a large herbivore in a densely populated island. In: Report of the 3rd IUCN World Conservation Congress. Bangkok
Ramakrishnan U, Coss RG, Pelkey NW (1999) Tiger decline caused by the reduction of large ungulate prey: evidence from a study of leopard diets in southern India. Biol Cons 89:113–120
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera-trap data. J Agri Biol Env Stat 14:322–337
Srikosamatara S (1993) Density and Biomass of Large Herbivores and other mammals in a Dry Tropical Forest, Western Thailand. J Trop Eco 9:33–43
Srikosamatara S, Suteethorn V (1995) Populations of Gaur and Banteng and their management in Thailand. Nat His Bul Siam Soc 43:55–83
Steinmetz R, Mather R (1996) Impact of Karen villages on the fauna of Thung Yai Nareseum Wildlife Sanctuary: a participatory research project. Nat His Bul Siam Soc 44:23–30
Steinmetz R, Chutipong W, Seuaturien N et al (2010) Population recovery patterns of Southeast Asian ungulates after poaching. Biol Cons 143:42–51
Steinmetz R, Seuaturien N, Chutipong W (2013) Tigers, leopards, and dholes in a half-empty forest: assessing species interactions in a guild of threatened carnivores. Biol Cons 163:68–78
Stephens DW, Krebs JR (1987) Foraging theory. Princeton University Press, Princeton, USA
Stoen OG, Wegge P (1996) Prey selection and prey removal by tiger (Panthera tigris) during the dry season in lowland Nepal. Mammalia 60:363–373
Taylor CC (2008) Automatic bandwidth selection for circular density estimation. Com Stat Data Anal 52:3493–3500
Thomas L, Buckland ST, Rexstad EA et al (2010) Distance software: design and analysis of distance sampling surveys for estimating population size. J App Eco 47:5–14
Timmins RJ (2011) Searching for the Last Kouprey: report to CEPF. Global Wildlife Conservation, Austin, Texas
Timmins RJ, Hedges S, Duckworth JW (2008) Bos sauveli. IUCN 2009. IUCN red list of threatened species. Version 2009.1. <www.iucnredlist.org>
Timmins RJ, Duckworth WJ, Hedges S, et al. (2008) Bos javanicus IUCN 2010. IUCN red list of threatened species. Version 2010.2. www.iucnredlist.org
Tordoff AW, Timmins RJ, Maxwell A et al (2005) Biological assessment of the Lower Mekong dry forests eco region. WWF-Cambodia, Phnom Penh
Vongkhamheng C (2011) Abundance and distribution of tiger ad prey in montane tropical forest in northern Lao PDR. PhD thesis. University of Florida
Wang SW (2010) Estimating population densities and biomass of ungulates in the temperate ecosystem of Bhutan. Oryx 44:376–382
Thailand WCS (2008) Strengthening conservation and monitoring of Tigers in Haui Kha Khaeng Wildlife Sanctuary and the Western Forest Complex, Thailand: Thailand Tigers Forever annual report 2007–2008. WCS Thailand, Bangkok
Wegge P, Storaas T (2000) Sampling tiger ungulate prey by the distance method: lessons learned in Bardia National Park. Nepal. Anim Cons 12:78–84
Weitzman MS (1970) Measure of the overlap of income distribution of white and begro families in the United States. Technical Report No. 22, US Department of Commerse, Bureau of the Census, Washington DC
Wharton CH (1957) An ecological study of the kouprey, Novibos sauveli (Urbain), vol 5. Monographs of the Institute of Science and Technology, Manila, pp 1–107
Wright HL, Collar NJ, Lake IR et al (2012) First census of white-shouldered ibis Pseudibis davisoni reveals roost-site mismatch with Cambodia’s protected areas. Oryx 46:236–239
Acknowledgments
This study forms part of WWF Greater Mekong Cambodia Country Program’s Eastern Plains Landscape Project. Major funding comes from WWF-US, WWF-Sweden, WWF-Germany, and Humanscale. Work in Mondulkiri Protected Forest is with permission of the Forestry Administration of the Ministry of Agriculture Forestry and Fisheries with support from His Excellency Cheng Kimsun, Men Phymean, Song Keang, and Keo Sopheak. Work in Phnom Prich Wildlife Sanctuary is with permission of the Ministry of the Environment and support from His Excellency Chay Samith, Sanrangdy Vicheth, and Han Sakhan. This project would not have been possible without the hard work and dedication of our field teams: Khaev Oudom, Ing Seangnirithy, Lien Nor, and Van Sonny (MPF) and Sary Tre, Sin Somoan, Se Noun, and Chan Touy (PPWS). Craig Bruce, Pin Chanrattanak, Nick Cox, Keith Metzner, Amy Malling, Pem Maly, Michelle Owen, and Seng Teak provided logistical and administrative support whilst Hannah O’Kelly, Bivash Pandav, Abishek Harihar, Paul Doggerty, Shannon Barber-Meyer, and Barney Long assisted in study design, survey objectives, and provided statistical advice.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Gray, T.N.E., Prum, S., Phan, C. (2016). Density and Activity Patterns of the Globally Significant Large Herbivore Populations of Cambodia’s Eastern Plains Landscape. In: Ahrestani, F., Sankaran, M. (eds) The Ecology of Large Herbivores in South and Southeast Asia. Ecological Studies, vol 225. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-7570-0_9
Download citation
DOI: https://doi.org/10.1007/978-94-017-7570-0_9
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-7568-7
Online ISBN: 978-94-017-7570-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)