
Abstract
Diatoms commonly called as “Jewels of Sea” are very intricate and beautiful microscopic photosynthetic algae. They make up approximately 90 % of all living organisms in sea and they are expected to produce 20 – 40 % oxygen of earth’s atmosphere. They have distinct cell wall which is silicified and hence called “Algae in Glass house”. There are approximately 20,000 – 2 million species which are present in diverse habitat. They not only photosynthesize and produce O2 but because of this they are able to store lipids and are being considered as a new promising source of biofuel. In the present chapter an attempt has been made to discuss the detailed ecology, classification, structure and reproduction of diatoms.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Diatoms, the members of class Bacillariophyceae are grouped under planktonic algae, as mostly they are free floating microorganism of marine or freshwater environment. Many diatoms live epiphytically on the surface of aquatic organisms including plants, molluscs, turtles, fishes, seaweeds etc. When under water, we are always surrounded with diatoms, but because they are at or below the limit of resolution of naked eye, we can literally swim through them and never know that they are present in surroundings (Nagy 2011). Diatoms are supposed to be originated in late Permian era (about 200 years ago) possibly from Chrysophyceae or Bolidophyceae (Guillou et al. 1999; Lee 2008). Diatoms are considered as the primary producers and thus contribute around 20–25 % of total global primary productivity (Hoek et al. 2009). Diatoms share some features that are typical of the heterokontophyta such as typical tripartite mastigonemes on flagellum, face of Golgi body, continuous chloroplast endoplasmic reticulum with nucleus, presence of periplastidial reticulum, presence of girdle lamellae in chloroplast, presence of chlorophyll a and c 2 . Diatoms have some unique characteristic features which separate them from other Heterokonts such as:
-
Marine and freshwater distribution
-
Unicellular or colonial protists consisting of two overlapping halves
-
Unique silica containing cell wall
-
Male gametes are only flagellated stage in centric diatoms
-
Lack of transitional helix in flagellum
-
Chloroplast usually golden brown (due to fucoxanthin)
-
Ring shaped chloroplast DNA
-
Open mitosis
-
Diplontic life cycle in sexually reproducible species
2 Classification
At present there are over 500 genera of diatoms (350 living and 150 extinct), with around 17,000 species (12,000 living and 5000 extinct) (Hoek et al. 2009; Williams and Kociolek 2011). Diatoms were sought to be unique group of algae by earlier scientist and they were placed as a distinct group by Harvey (1836), Eichler (1883), Engler and Prantle (1912). It was Antonie won Leewenhoek (1703) who recorded the appearance of diatoms, and considered them as animals. He named diatoms as “Animalcule” (The little animal) (Williams and Kociolek 2011). Linnaeus also considered them as animal and placed them under “Vermes”, in Zoophyta in the 12th edition of Systema naturae (1767). Carl Adolph Agardh gave a comprehensive diatom classification in Systema Algarum (1824) and later after few years in Conspectus criticus diatoma cerum (1830–1832) in Sytema Algarum Agardh classified diatoms in one order with 50 species.
Friedrich Traugott Kützing (1843) classified diatoms into two groups: Diatomaceae Liberae and Diatomaceae Inclusae with 16 and 10 genera respectively. Pfitzer (1871) classified diatoms into two divisions Coccochromaticae and Placochromaticae with two subdivisions each. The classification of Pfitzer was based on the structure of valve rather than contents of frustule (Williams and Kociolek 2011).
Petit (1877a, b), followed the classification of diatoms as proposed by Pfitzer and named his two subdivisions as families. H. L. Smith (1872) classified the diatoms based on the presence of raphe on the valve into three tribes with 15 families and 110 genera. This classification was adopted by Henri Van Heurck (1878). Franz Schütt (1896) classified diatoms into two groups Centricae and Pennatae and his classification has been proposed in Engler and Prantl’s Die naturlichen Pflanzenfamilien. With many such classification diatoms are sought to be a unique group of algae by earlier scientist and has always got recognitions as distinct group. Pascher (1931) kept diatoms under division Chrysophyta along with Crysophyceae. Fritsch (1935), proposed the most acceptable and comprehensive diatoms classification in which he kept all the diatoms under class Bacillariophyceae with two taxonomic groups Centrales and Pennales. G. M. Smith (1955) and Prescott (1969) kept Class Bacillariophyceae along with Chrysophyceae and Xanthophyceae in the division Chrysophyta as there are many important similarities between the two groups such as (1) olive green to brown colour chloroplast (2) presence of chlorophyll a and c with fucoxanthin and acetylenic carotenoids (3) oil and chrysolaminarin storage food (4) pleuronematic flagellum of gametes and (5) silicification of cell wall (Chapman and Chapman 1981). Bold and Wynne (1985) include three more classes namely Prymnesiophyceae, Eustigmatophyceae, Raphidophyceae along with Chrysophyceae, Xanthophyceae and Bacillariophyceae in division Chrysophyta. In comparison to other major algal groups classification of diatoms is relatively easy to investigate, because the silica shells bear characteristic patterns of pores and ornamentation. Fritsch (1935), Smith (1955), Prescott (1969), Morris (1971), Bold and Wynne (1985), and Hoek et al. (2009) divided class Bacillariophyceae into two orders namely Centrales and Pennales. A simple way of distinguishing between these two orders is that former one has pill boxes like shape and later one has shape of date boxes (Chapman 1962). Bold and Wynne (1985) further divided these orders in to following subgroups:
-
Order-Centrales
Valve arrangement is usually to a central point on the valve (centric or radial) or to 2, 3 or more points (gonioid) which resulted into biangular, triangular or polygonal valve is evident
-
Suborder: Coscinodiscineae
Valve disciform, flat or convex, without prominent processes or intercalary bands
-
Melosiraceae e.g. Melosira, Stephanopyxis
-
Coscinodiscaceae e.g. Coscinodiscus
-
Asterolampraceae e.g. Asteromphalus
-
Heliopeltaceae e.g. Actinoptychus
-
Thalassiosiraceae e.g. Cyclotella, Planktoniella, Skeletonema, Stephanodiscus, Thalassiosira
-
-
Suborder: Biddulphiineae
Valve bipolar or multipolar, angles of valves provided with spines, elevations, ocelli, or other projections
-
Eupodiscaceae e.g. Eucampia, Odontella, Triceratium, Zygoceros
-
Lithodesmiaceae e.g. Ditylum, Lithodesmium, Streptotheca
-
Stictodiscaceae e.g. Stictodiscus
-
Biddulphiaceae e.g. Biddulphia
-
-
Suborder:Rhizosoleniineae
Frustule long, cylindrical; many intercalary bands present
-
Rhizosoleniaceae e.g. Guinardia, Rhizosolenia
-
-
-
Order-Pennales
Structure of the valve is arranged in reference to a central line (pennate) or in reference to a point not on the valve surface (trellisoid); raphe or a hyaline field in the axial are present
-
Suborder: Araphidineae
Hyaline field present in the axial area of the valve
-
Fragilariaceae e.g. Asterionella, Climacosphenia, Fragilaria, Grammatophora, Rhabdonema, Licmophora, Striatella, Subsilicea, Synedra, Tabellaria
-
-
Suborder: Raphidiodineae
Rudimentary raphe present at ends of valve
-
Eunotiaceae e.g. Eunotia
-
-
Suborder: Biraphidineae
Fully developed raphe present on both valves
-
Naviculaceae e.g. Amphipleura, Berkeleya, Caloneis, Diploneis, Frustulia, Gyrosigma, Navicula, Pinnularia, Pleurosigma, Stauroneis
-
Cymbellaceae e.g. Cymbella, Gomphoneis, Gomphonema, Phaeodactylum
-
Entomoneidaceae e.g. Entomoneis
-
Bacillariiaceae e.g. Bacillaria, Cylindrotheca, Hantzschia, Nitzschia
-
Surirellaceae e.g. Campylodiscus, Cymatopleura, Surirella
-
-
Suborder: Monoraphidineae
Raphe present on one valve; hyaline field present on the araphid valve
-
Achanthaceae e.g. Achnanthes, Cocconeis
-
-
Lee (2008) proposed a different classification system of diatoms based on following characteristics and divided class Bacillariophyceae in to two orders.
S. no. | Characteristics | Order-Biddulphiales or centrals | Order-Bacillariales or pennales |
|---|---|---|---|
1 | Ornamentation | Radial or gonoid | Pennate or trellisoid |
2 | Chloroplasts | Many | One or two |
3 | Raphe | No raphe | Possibly present with gliding |
4 | Spermatozoids | Motile with a single tinsel flagellum | No flagellated spermatozoids |
5 | Sexual reproduction | Oogamous type | By conjugation |
Although a recent account of the diatoms recognizes many more classes and orders: there the diatoms (as division Bacillariophyceae) are subdivided into three classes, Fragilariophyceae (pennate diatoms without a raphe system), Bacillariophyceae (pennate diatoms with a raphe system) and Coscinodiscophyceae (the centric diatoms) (Hoek et al. 2009). Recently Williams (2007) and Williams and Kociolek (2011) gave detailed history of diatom’s classification in which they have discussed almost all the proposed classifications for diatoms till now.
3 Occurrence
Diatoms are extremely abundant, cosmopolitan which comprises the main component of aquatic systems, especially marine waters. They are widespread and can be present in terrestrial systems as well as in aquatic habitats as free living photosynthetic autotrophs, colorless heterotrophs, or photosynthetic symbionts. Diatoms are enormously successful organisms as judged by their adaptability, distribution, biomass and relative antiquity to survive in extreme conditions such as hot springs in Kuril and Sakhalin Island or extreme low pH lakes in Western Pomerania (NW Poland) (Nikulina and Kociolek 2011; Witkowski et al. 2011). Diatoms are largely responsible for the very high primary productivity in temperate or cold parts of the oceans where pennales mostly represent benthic forms and centrales are planktonics, which forms the basic food of aquatic animals. Many diatoms grow as epiphytes on the thalli of other algae or stems and leaves and other parts of the higher plants. Diatoms are also found as facultative heterotrophs (e.g. Nitzschia), or as soil diatoms, as endosymbionts diatoms (Hoek et al. 2009). Diatoms are greatly influenced by water salinity and therefore marine and freshwater forms of diatoms are strikingly different (Potapova 2011).
3.1 Freshwater and Marine Diatoms
-
Common marine centric diatoms: Thalassiosira, Chaetoceros, Skeletonema, Triceratium, Asteromphalus, Rhizosolenia, Biddulphia,
-
Common freshwater centric diatoms: Cyclotella, Stephanodiscus, Melosira
-
Common marine pennate diatoms: Licmophora
-
Common freshwater or marine pennate diatoms: Navicula, Nitzschia, Pseudo-nitzschia, Synedra, Cocconeis, Gyrosigma, Rhopalodia
-
Common primarily freshwater pennate diatoms: Cymbella, Epithemia, Pinnularia, Gomphonema, Stauroneis, Cymatopleura, Fragilaria, Tabellaria, Asterionella
3.2 Epizoic and Epiphytic Diatoms
Diatoms are found attached to a number of diverse animal groups such as ciliates, copepods, cladocera, barnacles, hydroids, krill, bryozoans, whales and porpoises, diving birds etc. Some of the common epizoic diatoms are Cocconeis, Bennettella, Epipellis, Melosira, Falcula etc. All types of aquatic vegetation (algae or higher plants, attached or free floating) can provide habitat for epiphytic diatoms. Some of the common epiphytic diatoms are Hyalodiscus, Licmophora, Cocconeis, Campyloneis, Leminicola, Achnanthes, Gomphonema, Fragilaria etc. (Tiffany 2011).
4 Morphology and Ultrastructure
Diatoms are unicellular, colonial or filamentous in organization. Two major series may be distinguished on the basis of their symmetry and other features: the pennate and centric diatoms (Alexopoulos and Bold 1971). With a few exceptions, such as Phaeodactylum tricornutum and endosymbiotic species, all diatoms are recognized by the possession of distinctive, essentially bipartite, variously perforated and ornamented, silica cell walls (frustules) that enclose the eukaryotic protoplast (Cox 2011). Diatoms are essentially unicellular, although chains of cells and colonial aggregation may occur. The vegetative cell of mostly diatoms can be regarded as having either bilateral or radial symmetry. Any diatoms cell with bilateral symmetry (pennate diatoms) can be positioned along three axes (Fig. 1): the apical axis, the pervalvar axis and the transapical axis (Bold and Wynne 1985). Diatom cells always divide along with the apical axis or valvar plane (Hoek et al. 2009).

Axes and planes of symmetry in a pennate diatoms. (a) Apical axis; (b) Transapical axis; (c) Pervalvar axis (Adopted from Bold and Wynne 1985)
4.1 Frustule and Raphe System
The key distinguishing feature of the diatoms is their ability to secrete an external wall, (frustule), which is composed of quartzite (95 %) or hydrated amorphous silica along with small amounts of aluminum, magnesium, iron, and titanium (Prescott 1969; Lee 2008). Frustule of diatoms composed of overlapping halves like that of a close fitting container with one larger valve (epivalve) and the other slightly smaller valve (hypovalve). Girdle is located between these two valves, which can be subdivided into two overlapping portion: the epicingulum and the hypocingulum. The epivalve and epicingulum makes epitheca and hypovalve and hypocingulum makes hypotheca (Bold and Wynne 1985). Under microscope, diatom’s shell can be oriented either in valve view or girdle view (Fig. 2). Simple girdle consist of two cingula only whereas additional pieces of intercalary bands also found in compound girdle (Bold and Wynne 1985). There may be one, two, or more additional intercalary bands besides the girdle that are inserted between the two wall sections (Prescott 1969). The additions of these intercalary bands also help in growth of diatoms along the pervalvar axis. A certain regular patterns in the siliceous material of the frustule leave the wall or frustule ornamented into four types: Centric and radial, Trellisoid, Gonoid and Pennate as shown in Fig. 2 (Lee 2008):

The basic patterns of ornamentation in the Bacillariophyceae. (a) Centric and radial e.g. Coscinodiscus (b) Trellisoid, e.g. Eunotia (c) Gonoid, e.g. Triceratium (d) Pennate e.g. Navicula (Adopted from Lee 2008)
Raphe system (a longitudinal slit in the theca) found in many pennate diatoms is an opening or fissure running along the apical axis (Bold and Wynne 1985). It is supposed that raphe is involved in diatom’s locomotion because raphe less diatoms always move sluggishly. Raphe-bearing diatoms are also known as raphid diatoms. Each raphe also divides the cell in two equal parts by the central nodule which terminates at polar nodule (Lee 2008). In cross section the raphe is sometime shaped like a “V” because slitting of the raphe as in Pinnularia (Hoek et al. 2009). Electron microscopical studies suggest that raphe system is also associated with several other cytoplasmic structures. A pseudoraphe, an unornamented area running down the center of the valve is present in those pennate diatoms, where raphe is absent (Fig. 3). Apart from the raphe, there are two other types of wall perforations found in diatoms (1) the simple pore or hole, and (2) the more complex loculus or areola (Lee 2008).

(a) Valve view of Pinnularia with a raphe system, (b) Girdle view of Pinnularia (c) Tabellaria with a pseudoraphe system (Adopted from Lee 2008)
Areolae are complex structures and have silicified layer, a sieve membrane which is also known as pore membrane or velum or cribrum or rica also found at one end (inside/outside) of the loculus (Lee 2008; Graham and Wilcox 2000). This membrane resembles honeycomb. Some processes or extensions found on colony forming diatom’s valve to maintain contact between themselves. These extensions have different names such as cornutate, strutted, spinulae, awns or setae (Lee 2008).
4.2 Extracellular Mucilage
In some diatoms, extracellular mucilage or slime pores (Mucilage secreting pores) are also reported. The extracellular mucilage aggregates are of five types namely tubes, pad, stalks, fibrils and adhering films (Fig. 4). According to some phycologists mucilaginous secretions help in diatom’s movement and attachment.

4.3 Protoplast
Protoplast is present within the cell wall and in a photosynthetic diatom, it comprises of a nucleus, mitochondria, endoplasmic reticulum, Golgi bodies, chloroplast with or without pyrenoids. One or two large central vacuoles are often present. The position of nucleus varies in both centrales and pennales diatoms.
The nucleus may often be present in the central portion of the vacuole interrupted by a broad band of cytoplasm in some pennales or araphid diatoms (Smith 1955) and raphid diatoms or it may be positioned against the girdle in centrales (Pickett-Heaps et al. 1988). In most of the raphid diatoms nucleus is always positioned at one side of the cell or oscillates across the cell with subsequent mitosis (Mann 1982a, b).
Numerous Golgi bodies surrounded by vesicles are present in protoplast. Golgi bodies in some centric diatoms are closely associated with endoplasmic reticulum and mitochondria and forms the Golgi-Endoplasmic Reticulum-Mitochondria complex (Medlin et al. 2000). Mitochondria are dispersed throughout the protoplast.
4.4 Plastids and Storage Food
Diatoms chloroplast has the same structure as in other typical members of the Heterokontophyta. Chloroplast remains surrounded by two membrane and outer membrane of chloroplast endoplasmic reticulum being continuous with the outer membrane of the nuclear membrane. Pyrenoids remain centrally placed in chloroplast. Chloroplasts are either brownish or greenish in colour and contains chlorophyll a, c 1 and c 2 along with fucoxanthin (carotenoid) which give golden brown colour to diatoms. Chlorophyll b is never found in diatoms. Some diatoms (apochlorotic diatoms) live on decaying marine vegetation and the mucilages of seaweeds (Lee 2008). These diatoms have no visual plastids and thus become facultative heterotrophs (Hoek et al. 2009).
Chrysolaminarin (β-1,3-linked glucan) is the main storage product of diatoms, which is located in vesicles in the diatoms (Hoek et al. 2009). Diatoms also contain unique 4α-methyl sterols, such as 4-desmethylsterol and cholesterol (Granum and Myklestad 2001; Lee 2008).
5 Motility
Diatoms do not move in a continuous straight line but in a series of curves and zigzags. One of the most spectacular patterns of motility is seen in Bacillaria paxillifera, a colony of pennate cells that continuously line up and then synchronously slide apart with precise rhythmic patterns (Bold and Wynne 1985). A number of hypothetical mechanisms have been proposed to explain diatom motility (Graham and Wilcox 2000). Pennate diatoms with a raphe system and centric diatoms with labiate processes can glide over the surface of a substrate which leaves a mucilaginous trail in their wake (Fig. 5). Labiate processes projects inward, terminating a flattened tube with a longitudinal slit surrounded by two liplike edges (Bold and Wynne 1985). These processes may be one to many. Raphe in pennate diatoms determine path of gliding which can be straight (e.g. Navicula), curved (e.g. Amphora) or curved with two different radii (e.g. Nitzschia). Some diatoms also exhibit alternate movement (backward and forward) at an interval of a minute. The contact of raphe with surface also helps in diatom’s gliding. If girdle contacts the substrate, a mucilaginous tether from the raphe near the central nodule attaches to the substratum and the cell pulls itself onto a valve containing a raphe using the tether (Lee 2008). Bundles of actin microfilaments parallel to raphe are found in gliding pennate diatoms. Labiate processes of some centric diatoms have a pore (slit like opening in rimoportula tube which is an elongate tube through the cell wall) in the center and secrete mucilage which helps in movement of centric diatoms (Hoek et al. 2009). According to Pickett-Heaps et al. (1988) raphe of pennate diatoms evolved from labiate processes of some centric diatoms.

6 Formation of the Cell Wall
Diatoms cell wall is an amorphous and non-crystal structure, consisting of polymerized silicic acid, along with protein, polysaccharides and lipids (Hoek et al. 2009). During cell division of diatoms which is a normal method of reproduction, the valve of the parent cell becomes the epitheca of the two daughter cells. Each daughter cell then produces a new hypotheca (Lee 2008). Therefore, one daughter cell is of the same size as the parent cell, and the other is smaller. To overcome this uptake of silica is confined prior to the separation of the two daughter cells and there is a great requirement of silicon, if cell division is to take place. Silicon metabolism in diatoms has been the focus of research interest (Bold and Wynne 1985). Solid silica dissociates to produce silicic acid (97 % of dissolved silicon of oceans and 23 % of freshwater) under water:
Most of the Si(OH)4 is present as ionized silicic acid
At pH less than 9 this silicic acid gets auto-polymerized to form amorphous silica, main form of silicon in diatom cell walls. With the help of a transport protein, silica is taken up into the diatom cell by active transport system. Epitheca is pushed away from hypotheca before cell or nuclear division. The silicalemma or membrane of the silica deposition vesicles formed by the fusion of translucent vesicles produced by golgi body beneath invagination of the plasmalemma during cell division (Li et al. 1989). Packaging of silicon is accomplished by golgi into vesicles with the help of microfilaments in the cytoplasm (Lee 2008). These vesicles determine the final form of the silicified frustule. In the form of 30–50 nm diameter spheres, silica remain deposited as amorphous form in diatoms (Crawford et al. 2001). Silaffin peptide determine the size of silica sphere and controls the frustule ornamentation. Frustulins (a glycoprotein) is also associated with the silica deposition vesicle. After complete deposition of silica inner membrane of the silicalemma becomes the plasmalemma of the daughter cell (Lee 2008). According to Bidle and Azam (1999) this silica usually dissolves after death of diatom cell.
7 Reproduction
The diatoms have a complex life cycle, as in other unicellular microalgae, diatoms also has two main distinct but interconnected phases-vegetative phase and a sexual phase. Vegetative phase includes mitosis and increase in cell number, whereas sexual phase includes meiosis and genetic recombination. Diatoms have a peculiar property in their life cycle, i.e. the diatoms cell progressively reduce in size after each mitotic division (Round et al. 1990). In such diatoms, the sexual phase is the only way to avoid death and reduction in number. Although, some diatom species evolve by enlargement in size to escape extreme miniaturization (Chepurnov et al. 2004).
7.1 Vegetative Phase
7.1.1 Cell Division
This is the commonest method of reproduction and usually cell division occurs at night. When a cell has to undergo division, protoplast expands, losing the connections between the overlapping thecae. The nuclear division takes place mitotically along with the division of cytoplasm in a longitudinal axis. Before the division of cytoplasm the cell organelles also duplicates. Sometimes spindles are formed by the centrosomes during nuclear division. After division each half is present either in epitheca or hypotheca, with opposite sides naked. A new hypotheca forms in each half of divided cell, thus reducing the size of one cell (Figs. 6 and 7).

Cell division in a pennate diatom

Process of cell division in diatoms showing continuous reduction in cell size
7.1.2 Resting Spore Formation
Under stress conditions the cell wall of some diatoms such as Ditylum, become thick, thus forming dormant or resting spores which can survive unfavourable period and divide on the onset of favourable period. During resting spore formation the protoplasm of the parental frustule darkens, contracts and develops a very heavy siliceous wall of its own. The cytoplasmic volume of these resting cells decreases due to the loss of vacuoles and their contents along with reduction in numbers of mitochondria and formation of large lipid bodies. Resting spores show sharp reduction in photosynthesis and respiration process. After formation the resting spore sinks to bottom (Bold and Wynne 1985). During germination, the resting spore produces a number of fine protoplasmic strands, which radiate in all directions (Lee 2008). Compared to auxospores formation (sometime once in more than 2 years) resting spores formation is more frequent in diatoms. Light, temperature, and salinity are comparatively stable parameter in sea thus nutrient depletion is main stress shock for resting spores formation which normally occurs after diatoms bloom (Lee 2008). In freshwater there is frequent environmental changes than marine waters which makes it more suitable for resting spore formation. Compared to pennate diatoms resting spore formation is far more common in centric diatoms.
7.2 Sexual Reproduction or Auxospores Formation
Compared to resting spores formation of auxospores is quite rare which is formed by the fusion of gametes. According to Edlund and Stoermer (1997) sexual reproduction or auxospore formation occur only in two conditions: (1) if cell reach one third of their maximum size and (2) during suitable environmental condition such as temperature, light, nutrients, trace metals, organic growth factors, and osmolarity (Lee 2008). Contrary to other algal groups diatoms undergo sexual reproduction for restoration of cell size. In centric and gonoid diatoms only male gametes bear flagella (motile) and female gamete act as egg cell (non-motile). On the other hand in pennate and trellisoid diatoms gametes are flagella-less. The fusion of gametes results into the zygote which is also known as “Auxospore”. It is larger than the vegetative cells and is surrounded by a two piece silicified wall called “perizonium”, which can be smooth or ornamented. The perizonium is secreted by the zygotic protoplast and it replace the thin zygotic membrane, when a zygote grows. When perizonium covers the auxospores which is a growth spore, it photosynthesis and grow rapidly to attain the size greater than the vegetative cell from which it has been formed. After this frustule is secreted, a new diatom is formed. Auxospores develop into three types’ namely isodiametric auxospores (e.g. Melosira, Coscinodiscus, Stephanopyxis), properizonialFootnote 1 auxospores (e.g. Chaetoceros) and perizonial auxospores (e.g. Navicula, Pseudo-nitzschia). Isodiametric and properizonial auxospores are common in centric diatoms whereas perizonial auxospores are formed in pennate diatoms (Lee 2008). Isodiametric auxospores are normally attached to the parent valves during maturation. The immature properizonial auxospore splits and bands (also known as properizonium) are produced from an asymmetrical auxospore. On the other hand in perizonial auxospores immature globular auxospores has scales embedded in a primary wall and the primary auxospores wall divide into two equal halves. Auxospore formation vary considerably in Centrales and Pennales diatoms as discussed below.
7.2.1 Auxospore Formation in Pennales
Mostly the pennate diatoms are monoecious (homothallic) and rarely dioecious (heterothallis) for e.g. Rhabdonema adriaticum and Navicula halophila. In pennales auxospores are either formed mostly by union between iso and anisogametes, or oogamy and rarely by autogamy or parthenogenesis.
-
Fusion of iso or anisogametes: The fusion of isogametes or anisogametes either forms one auxospore from two cells or two auxospores from two cells
-
A.
Formation of single auxospores from fusion of two cells: It is common in Surirella saxonica, Cocconeis species. During conjugation two diatoms (sister or non-sister cells) come close and lie next to each other either by side to side or end to end (Fig. 8). Once they are close, get eneveloped by common mucilage secreted by them. The first meiotic division takes place in diploid nuclei of both cells, but there is immediate degeneration of one sister nuclei in each cell. There may be degeneration of one nucleus after each division (Cocconeis pediculus) or three haploid nuclei degenerate after the completion of meiosis. In each sister cell (Geitler 1927; Karsten 1899), resulting into one haploid nuclei in each cell. Protoplast of which finally behaves as a haploid gamete. It escapes from the frustule. Since, the liberated gametes are identical are called Isogammetes. The two isogametes show amoeboid movement, escape from the parent frustule and fuse immediately in between the empty parent cells, forming a diploid zygote. The zygote enlarge, (more in longitudinal plane) and functions as an auxospores. Later, it secretes a fresh frustule within the perizonium, thus producing a new vegetative cell resembling parent cell both in size and construction.
Fig. 8 
Two fusing diatoms lying close to each other in a common mucilaginous envelope
-
B.
Formation of two auxospores from fusion of two cells: In Pennale diatoms, this is the commonest method, e.g. Cymbella lanceolata. During this, the two diminutive diatoms of same species come together and secrete a mucilaginous envelop around them. The diploid nucleus of each cell divides meiotically and forms four haploid nuclei, two of which degenerate. The protoplast of sustaining cells now divides into two equal or unequal daughters either longitudinally or transversely, which functions as a gamete. Each gamete thus contains one large haploid nucleus (functional) and one degenerated haploid nucleus. The two gametes formed by a cell may be of equal or unequal in size depending on whether the division is equal or unequal in parent protoplast. The two gametes produced by a cell normally unite with the gamete produced from the other cell and many a times physiological anisogamy is present (Smith 1955). The gametic union occurs by either of the following ways:
-
(i)
Both the gametes are amoeboid, alike and are isogametes. They unite midway between the parent frustule and the two zygote are formed outside the parent frustule and this is the most common method e.g. Navicula radiosa
-
(ii)
Either of the gamete is active or amoeboid and the other one is passive or immobile. This type of condition is observed either in two gametes of same cell, or one parent cell produces two mobile gametes and other parent cell produce two immobile gametes (e.g. Navicula halophila). This condition represents physiological anisogamy.
The mobile gametes, move out of the parent frustule and creep inside the other parent cell to fuse with the passive gametes of other cell and produces two zygotes in one cell. They remain dormant for some time and later enlarge parallel to the longitudinal axes of the parent frustule to form an auxospores. The auxospores is considerable larger than the parent cell and is enclosed by a membrane called perizonium, secreted by the either auxospores or it may be a ruminant of zygotic membrane. A new frustule is secreted inside the perizonium.
-
(iii)
In the third method, the two gametes are morphologically alike and have functional disparity. One of the gamete produced in parent frustule is smaller and active (amoeboid), while the other one is bigger and passive. Such type of gametes represents physiological anisogamy as the division of protoplast to form gametes is asymmetrical resulting into two gametes of unequal size with different functions. The amoeboid gamete from each parent frustule will come out, creep into the other parent frustule and will fuse with its passive counterpart e.g. Cymbella lanceolata, Gomphonema parvulum. By this method each cell have one zygote.
-
(i)
-
A.
-
Auxospore formation by oogamy (von Stosch 1958): Such type of auxospores formation is reported in dioecious diatoms example Rhabdonema adriaticum. One bigger diatom cell acts as female egg cell or oogonium. The diploid nucleus of oogonium undergoes meiosis and produces four haploid nuclei. Later the protoplast cleaves into a large functional ovum with two nuclei out of which one degenerates on maturity. The functional ovum occupies the upper third part of the female cell. Spermatogonia are formed in distinct male cells, which are smaller and numerous in number. Each spermatogonium nucleus undergoes meiosis, followed by cleavage of parent protoplast into two globular, naked, non-flagellated microgametes. Out of two microgametes one is fertile and other aborts. On liberation, the spermatogonium, carried away by water current to the oogonium and both of them get adhered because of mucilaginous pads. Only the male nucleus enters the oogonium for fertilization. The diploid zygote thus formed matures into an auxospores as usual.
-
Auxospore formation by autogamy: During autogamy, the diatom (e.g. Amphora normani) secretes thick mucilage, undergoes meiosis and forms four haploid nuclei, two of which degenerate. Two remaining haploid nuclei fuse immediately forming a diploid nucleus (autogamy). The protoplast with new diploid nucleus comes out from the old frustule and develops into a new auxospore.
-
Auxospore formation by parthenogenesis: During the process auxospore formation does not involve sexual reproduction and meiosis is replaced by mitotic division resulting into a diploid daughter cells. It has been reported in Rhabdonema adriaticum, Navicula, Nitzschia, Cocconeis placentula (Geitler 1973). During parthenogenesis the protoplast of a quiescent cell comes out and directly grows into an auxospore without any nuclear change or in many cases somatic division takes place in parent cell producing two daughters, each growing into an auxospore (Fig. 9).
Fig. 9 
Auxospore formation by parthenogenesis or apogamy in Cocconeis placentula var. lineata. Nuclear division in B and C are mitotic (Adopted from Reddy 2001)


7.2.2 Auxospore Formation in the Centrales
In centric diatoms sexual reproduction and auxospore formation are commonly occur by oogamy or autogamy. Isogamy and anisogamy are not common in centrales. A single auxospore is formed by a single individual which may be an old cell or a newly divided cell.
-
Auxospore formation by oogamy: The sexual mode of auxospore formation is common in many centric diatoms, where different diploid cells act as male or female gametangia. The male diatom cell or antheridia, may be a modified vegetative cell (Cyclotella species) or may be formed by division of a vegetative cell (Melosira species) (Fig. 10). Each antheridium or spermatogonium produces numerous sperm cells which vary from specie to species. There number varies from 4, 8, 16, 32, 64 or it may even reach 128 which is a rare phenomenon. Cell which act as an antheridium undergoes meiosis and produces haploid spermatozoids, which were earlier considered to be asexual in nature and was termed microspores. These spermatozoids are motile and have a single tinsel flagellum. Both uniflagellate and biflagellate sperms has been observed experimentally in culture of Cyclotella meneghiniana and C. cryptica by Schultz and Trainor (1970). The female gamete or egg is produced singly in an oogonium. The female cell undergoes meiosis and produces four haploid nuclei of which three nuclei disintegrates and one surviving nucleus along with undivided protoplast function as an egg. The spermatozoids get released from antheridium by the separation of frustule and theca. They swim and search egg. Both spermatozoid and egg cells get attached by mucilaginous fibres resulting into a zygote. After fertilization the zygote (diploid) gets released from the parent frustule and develops into an auxospore. The auxospore increases in size, its diploid nucleus divides into two by mitosis and only one persist. The auxospore with diploid surviving nucleus secretes a frustule inside the perizonium which called as “First ling Cell”. This is a new diatom individual.
Fig. 10 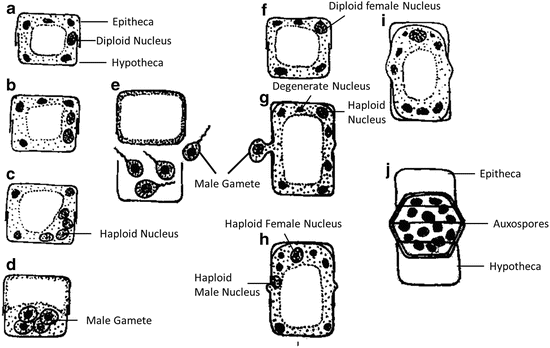
Auxospore formation by oogamy in the distinct male and female centric diatom Melosira varlans (Adopted from Reddy 2001)
-
Auxospore formation by autogamy: During autogamy, the protoplast of the diploid cell secretes a mucilaginous covering, forcing apart the theca, and nucleus undergoes meiosis resulting into four haploid nuclei. Out of four newly formed nuclei only two survive and fuse with each other thus resulting into a diploid nuclei. This is known as autogamy. After the formation of diploid nucleus, the protoplast escape from the parent frustule, and develops into an auxospore by forming a fresh frustule within the perizonium (Fig. 11).
Fig. 11 
Auxospore formation by autogamy in the centric diatom Cyclotella meneghiniana (Adopted from Reddy 2001)


8 Defense Against Predation
Some diatoms (e.g. Phaeodactylum, Skeletonema) release specific chemicals to defend themselves from many predators such as copepods, sea urchins, polychaetes, and ascidians. These chemicals include highly unsaturated fatty acids such as eicosanoic acid, which is only toxic after the unsaturated fatty acid it’s released into the environment and conversion into an aldehyde (Lee 2008). Diatoms spine which help to kept diatoms at upper layer of oceans or lakes also discourage grazing by herbivorous zooplankton (Hoek et al. 2009).
9 Economic Importance
9.1 Diatomaceous Earth
In some parts where diatoms are largely responsible for the very high primary productivity and are found in abundance, the death of diatoms and there sinking to the bottom of these areas results in the accumulation of diatom oozes. Normally the frustules of diatoms are dissolved by bacterial degradation but under certain circumstances, the frustules remain intact and accumulate at the bottom of the water. These large fossil deposits from past geological period (but not older than Cretaceous era) are known as “diatomite” or “diatomaceous earth” or “kieselguhr” (Lee 2008; Hoek et al. 2009). This diatomaceous earth is being used by several industries such as:
-
Toothpaste
-
Metal polishes
-
As an absorbent for liquid nitroglycerin to make dynamite that could be transported with comparative safety
-
Filtration of liquids, especially sugar refineries
-
As insulation of boilers, blast furnaces, and other places where a high temperature is maintained
9.2 Diatoms as Indicators
Diatoms are also being used as indicators of water quality and pollution tolerance (Bold and Wynne 1985). Many heavy metals have inhibitory effect on diatoms such as Cu, Zi and Ge can inhibit silicon metabolism whereas Hg, Cd, and Pb inhibit cell division. On the other hand Cr, Ni, and Se have inhibitory effect only at very high concentration (>1 μM). Due to high sensitivity copper is normally being used to control diatom blooms. Concentrations of germanium dioxide (GeO2) above 1.5 mg liter -1 will specifically suppress the growth of diatoms except those which have little or no silicified wall such as Phaeodactylum tricornutum (Lee 2008).
9.3 Health Effects
Some diatoms show shellfish poisoning such as Nitzschia, Pseudonitzschia, and Amphora (Lee 2008). Memory loss (amnesia), abdominal cramps, vomiting, disorientation, and even death can be seen in human and other animals after subsequent ingestion of shellfish (Lee 2008).
10 Adaptation of Diatoms
Main problem which is faced by heavy silica cell walled diatoms during photosynthesis is to remain in the uppermost layers of lakes or oceans to get enough light (Morris 1971). To overcome this problem several marine diatoms such as Ditylum reduce their density and become more buoyant by excluding heavy ions from their cell sap. Some diatoms increased their surface area by setae (Chaetoceros) or by cells shaped as discs (Coscinodiscus), ribbons (Fragilaria) or elongate forms (Rhizosolenia) (Lee 2008). On the other hand many diatoms bear long spines, horns, setae or other protrusions such as in Thalassiosira that result in a greater surface area relative to the volume (Bold and Wynne 1985; Hoek et al. 2009). Some diatoms (e.g. Ditylum) change rate of fat deposition with light period to remain floating.
Diatoms living at great water depth which receive only blue-green light have a “switch on mechanism” to capture maximum light and increased fixation of carbon dioxide (Lee 2008). Many diatoms such as Rhizosolenia and Hemiaulus also remain as symbionts with nitrogen-fixing bacteria and cyanobacteria. Rhizosolenia can sink (negatively buoyant) after nitrogen deprivation but can move up (positive buoyant) after taking up nutrients (Lee 2008).
11 Phylogeny
In terms of evolutionary diversification, the diatoms have been widely successful (Graham and Wilcox 2000). First diatoms were discovered at the beginning of eighteenth century but lack of advance microscopic technology it took almost one century for their detailed study (Mann and Evans 2007). Because of the siliceous nature of the diatom cell walls, well preserved fossil are available. The centrales have been reported from the Jurassic and the Pennales from early Tertiary (Kumar and Singh 1971). The group of diatom is one of the rare groups in which no organism regarded previously as diatoms have been not to be diatoms. However there are some unconventional representatives too such as shell-less endosymbionts of dinoflagellates (Mann and Evans 2007). Well before the molecular techniques, transmission electron microscopy and basic biochemical techniques had shown long before that diatoms belong to heterokonts, together with Phaeophyceae, Crysophyceae, Xanthophyceae, Eustigmatophyceae, Oomycetes and some other classes of autotrophic and heterotrophic protists (Mann and Evans 2007). Molecular sequences information revealed two lineage or clades. Clade 1 consist of Centric diatoms, many of which may possess peripheral rings of tube-like structure (labiate processes or rimoportulae. Clade 2 includes centric diatoms having central labiate processes or fultoportula and pennate diatoms (Graham and Wilcox 2000). Fossil records suggest that centric diatoms evolved before pennate diatoms (Round et al. 1990).
Notes
- 1.
The term perizonial derived from perizonium. Perizonium is an inner silicified membrane within which the initial cell is formed. In the great majority of centric diatoms only one membrane or fertilization membrane or perizonium remain (Bold and Wynne 1985).
References
Agardh CA (1824) Systema algarum. Lund, Burling, 312pp
Alexopoulos CJ, Bold HC (1971) Algae and fungi, 4th edn. The Macmillan Company, Collier-Macmillan Limited, Toronto/London
Bidle KD, Azam F (1999) Accelerated dissolution of diatom silica by marine bacterial assemblages. Nature 397:508–512
Bold HC, Wynne MJ (1985) Introduction to algae: structure and reproduction, 2nd edn. Prentice-Hall, Englewood Cliffs
Chapman VJ (1962) The algae. Macmillan and Co Ltd., New York, St Martin’s Press London
Chapman VJ, Chapman DJ (1981) The algae, 2nd edn. Macmillan Press Limited, London/Basingstoke
Chepurnov VA, Mann DG, Sabbe K, Vyverman W (2004) Experimental studies on sexual reproduction in diatoms. Int Rev Cytol 237:91–154
Cox EJ (2011) Morphology, cell wall, cytology, ultrastructure and morphogenetic studies. In: Seckbach J, Kociolek JP (eds) Cellular origin, life in extreme habitats and astrobiology volume 19: the diatom world, Springer Publishing House, London, pp 21–45
Crawford SA, Higgins MJ, Mulvaney P, Wetherbee R (2001) Nanostructure of the diatom frustule as revealed by atomic force and scanning electron microscopy. J Phycol 37:543–554
Edlund MB, Stoermer EF (1997) Ecological, evolutionary, and systematic significance of diatom life histories. J Phycol 33:897–918
Eichler AW (1883) Syllabus der Vorlesungen über Phanerogamenkunde, Berlin: Borntraeger 3rd edn
Engler A, Prantl K (1912) Die naturlichen Pflanzenfamilien, 23 vols. W. Engelmann, Leipzig
Fritsch FE (1935) The structure and reproduction of algae, vol I. Cambridge University Press, London
Geitler L (1927) Rhodospora sordida, nov. gen. et n. sp., eine neue “Bangiacee” des Süsswassers. Österreichische Botanisches Zeitschrift 76:25–28
Geitler L (1973) Auxosporenbildung und Systematik bei pennaten Diatomeen und die Cytologie von Cocconeis-Sippen. O¨sterreich. Bot Z 122:299–321
Graham LE, Wilcox LW (2000) Algae. Prentice Hall, Upper Saddle River 07458
Granum E, Myklestad SM (2001) Mobilization of β-1,3-glucan and biosynthesis of amino acids induced by NH4 + addition to N-limited cells of the marine diatom Skeletonema costatum (Bacillariophyceae). J Phycol 37:772–782
Guillou L, Chretiennot-Dinet M-J, Medlin LK, Claustre H, Loiseaux-de Goer S, Vaulot D (1999) Bolidomonas: a new genus with two species belonging to a new algal class, the Bolidophyceae (Heterokonta). J Phycol 35:368–381
Harvey WH (1836) Algae. In: Mackay JT (ed) Flora hibenica, vol 2. Curray, Dublin, pp 157–256
Hoagland KD, Rosowski JR, Gretz MR, Romer SC (1993) Diatom extracellular polymeric substances: function, fine structure, chemistry, and physiology. J Phycol 29:537–566
Hoek C, Mann DG, Jahns HM (2009) Algae. An introduction to phycology. Cambridge University Press, Daryaganj
Karsten G (1899) Die Diatomeen der Kieler Bucht. Wiss Meeresunters 4:17–205
Kumar HD, Singh HN (1971) A textbook on algae. East west press Pvt Ltd, New Delhi
Kützing FT (1843) Phycologia generalis oder Anatomie, Physiologie und Systemkunde der Tange. Mit 80 farbig gedruckten Tafeln, gezeichnet und gravirt vom Verfasser. pp. [part 1]: [i]-xxxii, [1]-142, [part 2:] 143–458, 1, err.], pls 1–80. Leipzig: F.A. Brockhaus
Lee RE (2008) Phycology, 4th edn. Cambridge University Press, London, pp 561
Leeuwenhoek C (1703) Part of a letter from Antony von Leeuwenhoek, concerning green weeds growing in water, and some animicula found about them. Philos Trans 23(283):1304–1311 (reprinted in leeuwenhoek A (1996). Letter no. 239 of 25 December, 1702, In: Palm LC (ed) The collected letters of Antoni von Leeuwenhoek, vol 14. Sweets and Zeitlinger, Lisse, pp 158–179
Li C-W, Chu S, Lee M (1989) Characterizing the silica deposition vesicle of diatoms. Protoplasma 151:156–163
Linnaeus C (1767) Systema naturae Tom. I. Pars II. Editio duodecima, reformata. Holmiae. (Laurentii Salvii), 533–1327
Mann DG (1982a) Auxospore formation in Licmophora (Bacillariophyta). Plant Syst Evol 139:289–294
Mann DG (1982b) Structure, life history and systematics of Rhoicosphenia (Bacillariophyta). II. Auxospore formation and perizonium structure of Rh. curvata. J Phycol 18:264–274
Mann DG, Evans KM (2007) Molecular genetics and the neglected art of diatomics. In: Brodie J, Lewis J (eds) From unravelling the algae the past, present, and future of algal systematics. CRC Press, Taylor & Francis Group, Boca Raton, pp 231–265
Medlin L, Kooistra W, Schmid A-M (2000) A review of the evolution of the diatoms – a total approach using molecules, morphology and geology. In: Witkowski A, Sieminska W (eds) The origin and early evolution of the diatoms: fossil, molecular and biogeographical approaches. Szafer Institute of Botany, Polish Academy Sciences, Crakow, pp 13–35
Morris I (1971) An introduction to the algae, 2nd edn. Hutchinson University Library, London, pp 178–202
Nagy SS (2011) Collection, cleaning, mounting, and photographing diatoms. In: Seckbach J, Kociolek JP (eds) Cellular origin, life in extreme habitats and astrobiology volume 19: the diatom world, Springer Publishing House, London, pp 1–18
Nikulina TV, Kociolek JP (2011) Diatoms from hot spring from Kuril and Sakhalin Islands (far east, Russia). In: Seckbach J, Kociolek JP (eds) Cellular origin, life in extreme habitats and astrobiology volume 19: the diatom world, Springer Publishing House, London, pp 333–363
Pascher A (1931) SystematischeUbersicht uber die mit Flagellaten in Zusamennhang stehenden Algentriehen und versuch einer Einreihung dieser Algenstame in die stamme des Pflanzen – reiches. Beibefte Botanischen Centralblatt 48:317–332
Petit P (1877a) An essay on the classification of diatoms. Mon Microsc J 18:10–14, 65–77
Petit P (1877b) Liste des Diatomees et des Desmidees observes dans les enviros de Paris, preceeded d’un essai de classification des Diatomees. Bulletin de la Societe Botanique de France 23–24:34–46
Pfitzer E (1871) Untersuchungen uber Bau und Entwickelung der Bacillariaceen (Diatomaceen). Bonn, Adolf Marcus [as Heft 2 of J. Hanstein, Botanische Abhandlungen aus dem Gebiet der Morphologie und Physiologie. 1872]
Pickett-Heaps JD, Wetherbee R, Hill DRA (1988) Cell division and morphogenesis of the labiate process in the centric diatom Ditylum brightwellii. Protoplasma 143:139–149
Pickett–Heaps JD, Hill DRA, Wetherbee R (1986) Cellular movement in the centric diatom Odontella sinensis. J Phycol 22: 334–339
Potapova M (2011) Patterns of diatom distribution in relation to salinity. In: Seckbach J, Kociolek JP (eds) Cellular origin, life in extreme habitats and astrobiology volume 19: the diatom world, Springer Publishing House, London, pp 313–332
Prescott GW (1969) The algae: a review. Michigan State University. Thomas Nelson and Sons Ltd. 597, Melbourne
Reddy SM (2001) University botany I: (algae, fungi, bryophyta and pteridophyta), 1st edn. New Age International (P) Limited, Publisher, Daryaganj
Round FE, Crawford RM, Mann DG (1990) The diatoms. Cambridge University Press, Cambridge, 747 pp
Schultz ME, Trainor FR (1970) Production of male gametes and auxospores in a polymorphic clone of the centric diatom Cyclotella. Can J Bot 48:947–951
Schütt F (1896) Bacillariales. In: Engler A, Prantl K (eds) Die Natürlichen Pflanzenfamilien, 1(1b): 31–153. W. Englemann, Leipzig
Smith HL (1872) Conspectus of the families and genera of the Diatomaceae. Lens 1:1–19, 72–93, 154–157
Smith GM (1955) Cryptogamic botany, vol 1, Algae and fungi. McGraw-Hill Book Company, New York
Tiffany MI (2011) Epizoic and epiphytic diatoms. In: Seckbach J, Kociolek JP (eds) Cellular origin, life in extreme habitats and astrobiology volume 19: the diatom world, Springer Publishing House, London, pp 195–209
Van Heurck HF (1878) Le microscope: la construction, son maniement et son application a ‘L’ anatomie vegetale et aux diatomees, Troisieme edition. E. ramlot, Bruxelles
von Stosch HA (1958) Kann die oogame Araphidee Rhabdonema adriaticum als Bindeglied zwischen den beiden grosssen Diatomeengruppen angesehen werden? Ber Dtsch Bot Ges 71:241–249
Williams DM (2007) Classification and diatoms systematics: the past, the present and the future. In: Brodie J, Lewis J (eds) Unravelling the algae. CRC Press, Taylor & Francis Group, Boca Raton, pp 57–91
Williams DM, Kociolek JP (2011) An overview of diatom classification with some prospects for the future. In: Seckbach J, Kociolek JP (eds) Cellular origin, life in extreme habitats and astrobiology volume 19: the diatom world, Springer Publishing House, London, pp 47–91
Witkowski A, Radziejewska T, Wawrzyniak-Wydrowska B, Lange-Bertalot H, Bak M, Gelbrecht J (2011) Living on the pH edge: diatom assemblage of low-pH lakes in Western Pomerania (NW Poland). In: Seckbach J, Kociolek JP (eds) Cellular origin, life in extreme habitats and astrobiology volume 19: the diatom world, Springer Publishing House, London, pp 365–384
Acknowledgment
The authors are grateful for the support provided by Mrs. Meena Malhotra, Department of Botany. Maitreyi College, University of Delhi for sketches.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Kumar, S., Baweja, P., Sahoo, D. (2015). Diatoms: Yellow or Golden Brown Algae. In: Sahoo, D., Seckbach, J. (eds) The Algae World. Cellular Origin, Life in Extreme Habitats and Astrobiology, vol 26. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-7321-8_8
Download citation
DOI: https://doi.org/10.1007/978-94-017-7321-8_8
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-7320-1
Online ISBN: 978-94-017-7321-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)