
Abstract
The application of stem cells has always attracted great interest in the field of peripheral nerve regeneration. In recent years, the rapid development of neural tissue engineering makes it possible to use stem cell transplantation to repair peripheral nerve injury. Seed/support cell or cellular source from stem cell has been known as one of the components for neural tissue engineering. The tissue-engineered nerve grafts (TENGs) support the regeneration of longer peripheral nerve gaps than scaffold alone. A number of TENGs have been used experimentally to bridge long peripheral nerve gaps in various animal models, where the desired outcome is peripheral nerve regeneration and functional recovery. Stem cells may improve the local microenvironment in nerve injury sites, providing necessary conditions for axonal regeneration. Nowadays, the types of stem cells and their application tend to diversify. Stem cells are more effective in providing necessary factors that promote peripheral nerve regeneration. So far, the application of stem cells for peripheral nerve regeneration is limited mainly because of the low survival rate of transplanted stem cells due to host immune rejection and changes in the local microenvironment. Here, we summarize the latest research progress and application strategies of stem cells in peripheral nerve regeneration. To push the translation of stem cell application for peripheral nerve regeneration into the clinic, we anticipate that a TENG with a close proximity to the regenerative microenvironment of the peripheral nervous system (PNS) will be developed.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Peripheral nerve injury is a global clinical problem and causes a devastating impact on patients’ quality of life (Noble et al. 1998; Robinson 2000; Taylor et al. 2008; Asplund et al. 2009). In the USA, about 360,000 people suffer from upper extremity paralytic syndromes annually, and 44,000 upper extremity procedures involved the nervous system during the period of 1989–1991 (Kelsey et al. 1997); over 300,000 cases of peripheral nerve injury occur annually in Europe (Mohanna et al. 2003). Although the peripheral nervous system (PNS) has a greater capacity of axonal regeneration than the central nervous system (CNS) after injury, spontaneous repair of peripheral nerve is nearly always unsatisfied with poor functional recovery. Various types of medical therapy have been carried out for several hundred years with the intention of improving outcomes (Artico et al. 1996; Battiston et al. 2009). Peripheral nerve repair consequently represents a unique challenge and opportunity to clinical and translational neurosciences. Various grafts between the nerve stumps are required to bridge the gap and support axonal regeneration in a substantial nerve gap of peripheral nerve injury. An autologous nerve graft, which is usually a functionally less important nerve segment from another site of the body (Johnson and Soucacos 2008), is accepted as the gold standard therapy for peripheral nerve injury. However, there are inherent disadvantages, including the limited supply of donor nerves, a second surgery, donor site morbidity, and a mismatch between the donor nerve and the recipient site (Ortiguela et al. 1987; Mackinnon and Hudson 1992). These collectively have encouraged the development of alternatives to autologous nerve grafts.
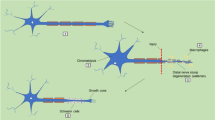
As is known, the microenvironment surrounding an injury site in PNS is often more permissive to axonal regeneration as compared to that in the CNS (Gu et al. 2011). Although peripheral nerve regeneration is ultimately determined by quality and speed of axonal outgrowth (Malin et al. 2009), Schwann cells (SCs) also play a critical role in the establishment of the regenerative microenvironment. It is the special importance of SCs in PNS that is responsible for the effectiveness of using SCs as support cells for the generation of TENGs (Johnson et al. 2005). Myelination of axons seems to be the most basic function of SCs because myelin sheath, as a unique component of the nervous system, can increase axonal conduction, especially salutatory conduction, thus allowing fast and efficient salutatory propagation of action potentials along the nerve (Honkanen et al. 2007; Salzer et al. 2008; Rumsey et al. 2009). On the other hand, the development of myelinated nerve fibers in PNS depends on complex interactions between SCs and axons. Axons can in turn promote the deposition of the basal lamina by SCs, which is required for the ensheathment of axons and the subsequent differentiation of SCs, and the maturation of fully myelinating SCs depends on contact and signaling from axons (Muir 2010; Wanner et al. 2006). Based on the insights into the interactions between SCs and axons, in this study, a unique scheme for incorporating biochemical cues into neural scaffold was adopted to establish an optimal regenerative microenvironment.
With progress in regenerative medicine, especially when tissue engineering and a subfield of neural tissue engineering has emerged, various biological and artificial nerve grafts, which are generally named tissue-engineered nerve grafts (TENGs), have been produced in attempts to supplement or even substitute for autologous nerve grafts. The typical TENGs involve both physiochemical and biological cues, which are provided by a biomaterial-based structure, as well as a multitude of cellular or molecular components. In recent years, many excellent review articles discuss the clinical applications and future directions of TENGs (Johnson and Soucacos 2008; Gu et al. 2011; Schmidt and Leach 2003; Chalfoun et al. 2006; Johnson et al. 2008; Seidlits et al. 2008; Deumens et al. 2010; Jiang et al. 2010; Khaing and Schmidt 2012; Rajaram et al. 2012; Zochodne 2012).
The application of various stem cells has always attracted much interest in the field of peripheral nerve regeneration. In recent years, the rapid development of neural tissue engineering makes it possible for the use of stem cell transplantation to repair peripheral nerve injury. Seed/support cell which usually is, or source from stem cell has been known as one of the components for neural tissue engineering. The TENGs can support longer peripheral nerve regeneration than the scaffold alone. A number of TENGs have been used experimentally to bridge long peripheral nerve gaps in various animal models, where the desired outcome is peripheral nerve regeneration and functional recovery. The stem cell may improve the local microenvironment in nerve injury sites, providing necessary conditions for axonal regeneration. Nowadays, the type of stem cell and their application tend to diversify. Stem cells are more effective to provide the necessary factors that promote peripheral nerve regeneration. So far, the application of stem cell for peripheral nerve regeneration is limited mainly because of the less survival rate of transplanted stem cell due to host immune rejection and changes in the local microenvironment. This chapter summarizes the latest research progress and application strategy of stem cells for peripheral nerve regeneration. To push the translation of stem cell application for peripheral nerve regeneration into the clinic, we anticipate that a TENG with a close proximity to the regenerative microenvironment of the PNS will be developed.
1.1 Types and Sources of Stem Cell for Peripheral Nerve Regeneration
1.1.1 Mesenchymal Stem Cells (MSCs)
The application of stem cells from different sources in the field of neural tissue engineering has attracted much interest, the bone marrow mesenchymal stem cells (also named bone marrow stromal cells, BMSCs) being undoubtedly the most important. MSCs are featured by their abilities to: (1) differentiate into mature cells and populate the resident tissue, having a therapeutic potential for regenerative medicine; (2) secrete growth factors or other soluble mediators; and (3) serve as a vehicle for protein drug delivery, namely perform gene therapy (Horwitz and Dominici 2008). They localize in the stromal compartment of the bone marrow, where they support hematopoiesis and differentiate into mesenchymal lineages (Johnson and Dorshkind 1986; Deryugina and Muller-Sieburg 1993; Bianco et al. 2001; Abdallah and Kassem 2008; Phinney and Prockop 2007; Franchi et al. 2012). Unorthodox plasticity of MSCs, however, has recently been described in that they have ability to cross oligolineage boundaries, in other words, to transdifferentiate into nonmesenchymal cells. Several in vitro studies have reported that MSCs can be induced to differentiate into neural lineages including neurons, astrocytes, oligodendrocytes, microglia, and SCs-like cells (Chen et al. 2006; Lu et al. 2008; Munoz-Elias et al. 2003; Sanchez-Ramos et al. 2000; Suzuki et al. 2004; Wislet-Gendebien et al. 2005; Woodbury et al. 2000), and specific approaches to induce transdifferentiation of rodent or human MSCs toward neural lineage cells have been established (Dezawa et al. 2001, 2004; Mimura et al. 2004). On the other hand, in vivo studies have also reported that after implanted into the brain, MSCs generate neural phenotypes specific to the injury site (Kocsis et al. 2002; Kopen et al. 1999; Lu et al. 2006). These results motivate explorations into the possibility of using MSCs as an alternative to SCs for peripheral nerve repair (Dezawa 2006). The MSCs are easily obtained through the aspiration of the bone marrow and expanded in a large scale by in vitro culture; BMSCs have found increasing applications in cell-based therapies for various diseases, including neural injury and disorders (Horwitz et al. 2002; Fickert et al. 2003; Ortiz et al. 2003; Kunter et al. 2006; Lee et al. 2006; Minguell and Erices 2006; Ringden et al. 2006). Despite the indispensable value of SCs for the construction of TENGs, autologous SCs are difficult to obtain in large number, and allogeneic SCs are involved in immunological rejections. Therefore, BMSCs have become a promising alternative to SCs for using as support cells within TENGs, showing considerable success in experimental studies (Hu et al. 2013; Ding et al. 2010; Yang et al. 2011).
MSCs pre-labeled with bromodeoxyuridine (BrdU) were injected into the distal stump of transected rat sciatic nerves. Dual immunofluorescence labeling showed that BrdU-reactive MSCs survived in the injected area for at least 33 days after implantation, and almost 5 % of BrdU cells exhibited Schwann cell-like phenotype (S-100 immunoreactivity). Walking track test at 18 and 33 days after implantation indicated that MSC implantation promoted functional recovery of injured nerves (Cuevas et al. 2002). GFP-labeled undifferentiated MSCs were seeded in a Matrigel-containing chitosan NGC to bridge a 5-mm rat sciatic nerve gap. After 6 weeks, the growth and myelination of axons and the sciatic functional index were significantly improved as compared to MSC-free NGC. Moreover, confocal microscopy confirmed that implanted MSCs adopted the Schwann cell-like phenotype (Zhang et al. 2005). Transdifferentiated MSCs, which had been transformed into Schwann cell-like cells under induction of a cocktail of cytokines, were implanted into devitalized muscle conduits for bridging a 20-mm rat sciatic nerve gap. The examinations at 6 weeks after implantation indicated that the transplanted MSCs were able to support peripheral nerve regeneration to a certain extent. Although the enhancement of nerve regeneration by MSC-containing NGCs was inferior to that by Schwann cell-containing NGCs or by autologous nerve grafts, such deficit in regenerative outcomes could be compensated by combining MSCs with muscle-derived allografts (Keilhoff et al. 2006a, b). Differentiated MSCs were suspended in Matrigel and transferred into hollow fibers, which were implanted across a 10-mm rat sciatic nerve gap. After 6 months, significant improvements in motor nerve conduction velocity and sciatic functional index were observed in the differentiated MSC-implanted animals, and GFP labeling clearly showed the trace of implanted MSCs within the regenerated nerves (Mimura et al. 2004). In a rhesus monkey model, an acellular allogeneic nerve graft seeded with autologous MSCs was implanted into a 10-mm radial nerve gap. After 8 weeks, nerve regeneration and functional restoration were observed, and the implanted MSCs exhibited Schwann cell-like phenotype as evidenced by double immunostaining for S-100 and BrdU (Wang et al. 2008).
The aforementioned observations lead to a transdifferentiation mechanism of MSCs, which assumes that the actions of MSCs on nerve regeneration, at least in part, stem from their ability to replace damaged neural cells via cellular differentiation; however, the mechanisms are also believed to be due to spontaneous fusion of MSCs with host cells rather than real transdifferentiation (Weimann et al. 2003a, b). The exact mechanisms behind the enhanced nerve regeneration in the presence of undifferentiated MSCs remain to be elucidated. The undifferentiated MSCs may contribute to nerve regeneration possibly by secreting growth factors and depositing basal lamina components (Chen et al. 2007; Wang et al. 2009). The production of neurotrophic molecules by MSCs can delay cell death and restore neural tissues (Borlongan et al. 2004; Chopp and Li 2002; Chopp et al. 2000; Crigler et al. 2006; Gu et al. 2010; Hofstetter et al. 2002; Munoz et al. 2005). Searching for undescribed mediators generated by MSCs will probably reveal a new array of important signaling secreted molecules (Horwitz and Dominici 2008). These results, together with the finding that local differentiation of MSCs is an uncommon event, suggest a new general paradigm for the neural activities of MSCs. Systemically infused MSCs exert a therapeutic effect primarily through the release of growth factors or other soluble mediators, which act on local even distant target tissues. Rather than serving as stem cells to replace neural cells and repair nerve tissues, MSCs behave as a small molecular factory secreting growth factors or other bioactive molecules to stimulate the reconstruction of nerve tissues or to establish a favorable microenvironment for nerve regeneration (Chopp and Li 2002; Caplan and Dennis 2006; Chen et al. 2002; Liu and Hwang 2005; Neuhuber et al. 2005). Some researchers propose that MSCs promote peripheral nerve regeneration not only via their direct release of growth factors, but also through indirect modulation of cellular behaviors of SCs (Wang et al. 2009). To sum up, the molecular mechanisms that are responsible for the favorable effects of MSCs on peripheral nerve regeneration seem to involve many aspects, including cell replacement, growth factor production, ECM molecule synthesis, microenvironment construction, and immune modulation, and they need further exploration.
Adipose tissue has also been identified as a niche for multipotent stem cells that have a phenotypic profile comparable to that of BMSCs and can differentiate into a myelinating Schwann cell-like phenotype in culture with lineage-specific stimuli (Kingham et al. 2007; Xu et al. 2008). In consequence, adipose-derived mesenchymal stem cells (AMSCs), also named adipose-derived stem cells (ADSCs), are potentially valuable because of their capability of multilineage differentiation in a manner resembling that of BMSCs. Importantly, AMSCs are superior to BMSCs in some aspects, such as the convenient harvesting of AMSCs through liposuction, a much less invasive method than bone marrow aspiration, and the greater availability of adipose tissue than bone marrow (Rider et al. 2008). To apply AMSCs for neural tissue engineering, many experimental studies in diverse animal models have been accomplished, in which different neural scaffolds containing either undifferentiated or differentiated AMSCs have bridged peripheral nerve gaps of different lengths (Yang et al. 2011; Erba et al. 2010; Zhang et al. 2010; di Summa et al. 2011; Scholz et al. 2011; Sun et al. 2011; Wei et al. 2011; Gu et al. 2012; Orbay et al. 2012; Shen et al. 2012; Tomita et al. 2012, 2013; Carriel et al. 2013; Mohammadi et al. 2013; Suganuma et al. 2013). All these studies indicate the favorable effects of AMSCs on peripheral nerve reconstruction and open a new approach for the use of support cells for constructing TENGs.
1.1.2 Embryonic Stem Cells (ESCs)
After peripheral nerve injury, many neurons die of insufficient nutrition. Because the number of neurons cannot be easily expanded by in vitro culture, there is a big difficulty to apply primary cultured neurons in nerve tissue engineering. ESCs have a great potential to proliferate unlimitedly and differentiate into neurons under various protocols; hence, they become a good candidate of support cells in cell-based therapies for neural injuries and disorders.
ESCs are undifferentiated, pluripotent cells derived from the inner cell mass of blastocyst-stage embryos and possess a nearly unlimited capacity for self-renewal and an ability to virtually differentiate into any kind of cell type in the body (Jakob 1984). The differentiation of ESCs can be modulated by the stimulation of growth factors (Schuldiner et al. 2000). Retinoic acid (RA) and nerve growth factor have been found to be potent enhancers of neuronal differentiation, eliciting extensive outgrowth of processes and expression of neuron-specific molecules (Schuldiner et al. 2001).
Extensive research has focused on the implantation of ESCs for treating the CNS disorders, while the potential of ESC-based therapy for the PNS injuries is largely unknown. Based on the assumption that implantation of neuronal cells derived from ESCs into denervated muscle would replace lost neurons or nonneuronal cells and prevent muscle atrophy in a peripheral nerve injury model, ESCs were differentiated into cholinergic motor neuron progenitors and labeled with florescence, followed by injection into gastrocnemius muscle of rats after denervation by ipilateral sciatic nerve transection. The observation showed that motor neuron progenitors prevented muscle atrophy after denervation for a brief time (Craff et al. 2007). In another study, ESC-derived neural progenitor cells were implanted into a 10-mm rat sciatic nerve gap, resulting in substantial axonal regrowth and nerve repair. The implanted cells survived until 3 months and differentiated into myelinating cells. Nerve stumps showed nearly normal diameter with longitudinally oriented, densely packed Schwann cell-like cell arrangement. Electrophysiological recordings confirmed that functional activity recovered across the nerve gap (Cui et al. 2008). As reported by Yohn et al. (2008), ESC-derived motor neurons could form functional synapses with denervated host muscle after implantation into transected tibial nerves, thereby attenuating the denervation-induced muscle atrophy.
Furthermore, it is believed that neuronal cells derived from ESCs provide new choices of support cells for incorporation to neural scaffolds, promoting peripheral nerve regeneration.
1.1.3 Neural Stem Cells (NSCs)
NSCs are multipotent cells that reside within paramedian generative zones present along the entire neuraxis throughout all stages of neural development and also during adult life (Gokhan and Mehler 2001). NSCs have the potential to differentiate into three major cellular elements of the nervous system, including neurons, astrocytes, and oligodendrocytes, and they can proliferate unlimitedly and undergo rapid cellular expansion in response to nerve injuries. The properties of NSCs, including multipotential differentiation, strong plasticity, high immigration ability, easily isolation and culture in vitro, and low immunogenicity, make NSCs an attractive source of support cells for the construction of TENGs (Alessandri et al. 2004). Chitosan NGCs seeded with NSCs were used as a tissue-engineered nerve graft to bridge 10-mm facial nerve gaps in rabbits. Nerve regeneration at 12 weeks after implantation was similar to that by autologous nerve grafting (Guo and Dong 2009). By GFP labeling, Hsu et al. (2009) found that 85 % of seeded NSCs were successfully aligned on the micropatterned poly (D,L-lactic acid) (PLA) NGC within 72 h, and the cells expressed the genes that are related to the production of neurotrophic factors and thus facilitated nerve repair and functional recovery in a 10-mm rat sciatic nerve injury model during a period of 6 weeks. Genetically modified NSCs can also serve as a source of neurotrophic factors. NSCs engineered to overexpress glial cell line-derived neurotrophic factor, which is known to protect motoneurons, were implanted to chronically denervated distal tibial nerve. There was a better regeneration of peroneal axons into tibial nerve with a reduction of chondroitin sulfate proteoglycan immunoreactivity in the ECM (Heine et al. 2004). Implantation of NSC overexpressing glial cell line-derived neurotrophic factor could significantly increase the nerve action potential amplitude, axonal area, and axonal number, as well as the labeling for S-100, NF, and beta III tubulin (Shi et al. 2009). After NSCs transfected with neurotrophin-3 or its receptor were incorporated into a PLGA NGC followed by 14-day culture, the viable NSCs were widely distributed within the NGC. This construct permitted the NSCs to differentiate toward neurons and to exhibit synaptic activities (Xiong et al. 2009).
1.1.4 Olfactory Ensheathing Cells (OECs)
Olfactory ensheathing cells (OECs) are neural crest cells which allow growth and regrowth of the primary olfactory neurons. Indeed, the primary olfactory system is characterized by its ability to give rise to new neurons even in adult animals. This particular ability is partly due to the presence of OECs which create a favorable microenvironment for neurogenesis (Guerout et al. 2014).
OECs are a unique type of glial cells that wrap olfactory axons and support their continual regeneration from the olfactory epithelium to the bulb (Su et al. 2013). OECs develop from a peripheral origin, the olfactory placode, and retain the ability to self-renew and differentiate, and are considered as peripheral nerve progenitor cells (Tohill and Terenghi 2004; Fairless and Barnett 2005). OECs that share both Schwann cell and astrocytic characteristics have been shown to promote axonal regeneration after transplantation. The tissue-engineered poly (lactic-co-glycolic acid) (PLGA) seeded with OECs was verified to improve peripheral nerve regeneration in a long sciatic nerve defect (Tan et al. 2013).
OECs have been studied in the context of enhancing repair of peripheral nerve by direct transplantation in different peripheral nerve lesion models for enhancement of axonal nerve regeneration by providing a scaffold for the regenerating axons as well as trophic factors and directional cues (Deumens et al. 2006). OECs are known to provide trophic factors conducive to axonal regeneration and survival. They may promote endogenous SCs mobilization possibly by a trophic influence (Cao et al. 2007; Au et al. 2007). Experimental studies performed in rodents show that transplantation of OECs into injured nerve or implantation of OEC-seeded conduits leads to an enhancement in axonal regeneration and improved functional outcome under some experimental conditions. Axonal dieback of the proximal nerve stump is reduced in the OEC transplanted nerves suggesting that the OECs provided early trophic support leading to earlier onset of regeneration. This could be critical for allowing the regenerating axons to navigate across the injury site before impeding scar tissue develops. However, OECs share many properties with SCs such as their production of neurotrophic factors and extracellular matrix (ECM) molecules as well as their ability to form peripheral myelin. Transplanted identified eGFP-expressing OECs integrate into the nerve injury site and remyelinate the regenerated axons, suggesting direct participation of OECs in the repair process (Radtke and Kocsis 2012).
OECs-containing silicone tubes were noted to support an improved axonal regeneration in 50 or 79 % of rats with a 15- or 12-mm sciatic nerve injury gap (Verdu et al. 1999). Another case showed that bridging of 15-mm nerve gap in rat sciatic nerve injury model with muscle-stuffed vein seeded with OECs as a substitute for autologous nerve graft. Neurophysiological recovery, as assessed by electrophysiological analysis, was faster in the constructed biological nerve conduit compared to that of autologous nerve graft (Lokanathan et al. 2014). Although the therapeutic potential of OECs in peripheral nerve repair is yet far from conclusive, there have been later studies reporting on the treatment of peripheral nerve injury by direct injection of OECs to the injured site (Andrews and Stelzner 2004; Dombrowski et al. 2006; Radtke et al. 2009; Guerout et al. 2011).
1.1.5 Induced Pluripotent Stem (iPS) Cells
Cell source is a major issue for tissue engineering and regenerative medicine. An exciting breakthrough in stem cell biology is that adult somatic cells (e.g., skin fibroblasts) can be reprogrammed into induced pluripotent stem cells (iPSCs) by the activation of a limited number of genes (transgenes) such as Oct3/4, Sox2, c-Myc, and KLF4 (Takahashi et al. 2007; Park et al. 2008) or Oct3/4, Sox2, Nanog, and Lin28 (Yu et al. 2007). The iPSCs derived from somatic cells make it possible for patient-specific cell therapies, which bypass immune rejection issue and ethical concerns of deriving and using ESCs as a cell source. The unlimited expansion potential of iPSCs also makes them a valuable cell source for tissue engineering. However, to use iPSCs as a cell source, many important issues remain to be addressed, such as the differences among various iPSC lines in differentiation and expansion and the appropriate differentiation stage of the cells for specific tissue engineering applications. In general, most of iPSC lines, as ESCs, can differentiate into neural crest stem cells (NCSCs), although the differentiation efficiency was different. NCSCs can differentiate into cell types of all three germ layers and represent a valuable model system to investigate the differentiation and therapeutic potential of stem cells (Rao and Anderson 1997; Morrison et al. 1999; Crane and Trainor 2006; Lee et al. 2007; Sauka-Spengler and Bronner-Fraser 2008). The adult cell sources are limited by the number of cells that can be obtained and complicated by the need to sacrifice additional nerves and tissues. Moreover, there is a lack of efficiency and consistency in cell isolation and expansion, which causes variability in therapeutic efficacy. In contrast, NCSCs derived from iPSCs can be immune compatible, expandable, and well characterized as a valuable cell source for the regeneration of peripheral nerve and other tissues (Wang et al. 2011).
Stem cells from sources other than the bone marrow are now getting more attention. The gliogenic secondary neurospheres derived from iPS cells have the ability to differentiate into SCs. The iPS cells were added to a PLC-based NGC, followed by implantation across a sciatic nerve gap in mice, showing regeneration of peripheral nerves and functional recovery (Uemura et al. 2012). iPSCs and their derivatives are valuable cell sources for tissue engineering.
1.1.6 Skin-Derived Precursors (SKPs)
Skin-derived precursors (SKPs) are stem cells found in the dermis (Fernandes et al. 2004; Blazejewska et al. 2009). These stem cells persist into adulthood, as they can be isolated from adult skin (Toma et al. 2001). In their endogenous environment, the dermis, they instruct hair follicle growth and contribute to maintaining the dermis and repairing it after injury. One niche where SKPs are found is at the base of hair follicles, in the dermal papilla (DP) and surrounding the follicle in the dermal sheath (DS) (Fernandes et al. 2004). The DP is the control center for hair growth (Jahoda et al. 1984), as such SKPs found in the DP instruct hair growth. SKPs can also migrate out of the DP. Upon injury to the dermis, SKPs migrate to the site of injury and differentiate into dermal fibroblasts, thereby replenishing the interfollicular dermis.
The skin dermis contains neural crest-related precursor cells, and the SKPs can be cultured to differentiate into neural crest cell types with the characteristics of neurons and SCs in the PNS (Fernandes et al. 2004; McKenzie et al. 2006). Both rodent and human SKPs are differentiated into SCs when transplanted into the brains of shiverer mice (McKenzie et al. 2006), which have a genetic deficiency in MBP and hypomyelination of the CNS (Dupouey et al. 1979). Both whisker pad SKPs and dorsal back SKPs differentiate into SCs with similar efficiency (Jinno et al. 2010). Of note, dorsal trunk SKPs are somite derived but can still differentiate into SCs, which are neural crest derived during development (Jinno et al. 2010).
SCs differentiated from SKPs express typical Schwann cell markers and can myelinate DRG axons in vitro. When transplanted into rodent models of peripheral nerve injury, SKP-SCs are able to myelinate axons in vivo and aid in injury repair (McKenzie et al. 2006; Walsh et al. 2009, 2010). The first experiment using SKP-SCs transplantation into the PNS used a crush model. Here, the sciatic nerve of a mouse was crushed with forceps (but not transected), and SKP-SCs were immediately transplanted distal to the crush site. SKP-SCs were able to myelinate the regenerating axons (McKenzie et al. 2006). SKP-SCs can also be used to repair transected nerves (Shakhbazau et al. 2014). In an acute model of peripheral injury, SKP-SCs were transplanted into an acellular nerve graft and used to bridge a 12-mm gap in the sciatic nerve. SKP-SCs promoted axon regeneration, myelination, and electrophysiological recovery (Walsh et al. 2009).
SKP-SCs were also able to regenerate chronic peripheral nerve injury. More motor neurons regenerated into the chronically denervated nerve with SKP-SCs transplantation compared with media controls, and these regenerated axons were larger. The compound muscle action potential (CMAP) amplitude in the gastrocnemius muscle and its muscle weight were larger with SKP-SCs transplantation, suggesting better muscle reinnervation. The reparative ability of SKP-SCs approached that of nerves that were immediately sutured and were not chronically denervated (Walsh et al. 2010). SKP-SCs therapy also improves behavioral recovery after acute, chronic, and nerve graft repair beyond the current standard of microsurgical nerve repair (Khuong et al. 2014).
The SKPs with neurotropic function show a full capacity of differentiating into SCs and promoting axon regeneration in vivo (Chen et al. 2012). In one study, SKPs were injected into neural scaffolds (NGCs) that had been prepared with L-lactide–trimethylene carbonate (L-lac/TMC) copolymer or type I collagen, respectively, to generate a TENG, which was then used to bridge a 16-mm sciatic nerve gap in rats. The results of the study confirmed the beneficial effects of SKPs on nerve regeneration (Marchesi et al. 2007). In another study, porcine SKPs were found to induce prominent nerve regeneration in porcine peripheral nerve injury sites after SKPs were added to a collagen/fibrin NGC for bridging a 10-mm femoral nerve gap in pigs (Park et al. 2012). More studies demonstrated further evidence for the effectiveness of using SKPs as support cells in TENGs (Walsh et al. 2009; Chen et al. 2012).
1.2 Application Strategy of Stem Cells for Peripheral Nerve Regeneration
1.2.1 Transplantation of Stem Cells Combined with Scaffolds for Repairing Peripheral Nerve Injury
It is reported in a recent study that evaluated the long-term safety of using support cells-containing TENGs to repair a 50-mm-long median nerve gap in monkeys in terms of the data from blood test, immunological and tumor marker detection, and histopathological examination of organs and glands (Hu et al. 2013). They also directly transplant MSCs combined with chitosan/PLGA scaffold to successfully repair the 50- and 60-mm sciatic nerve gap in dogs (Ding et al. 2010; Xue et al. 2012), and they developed a new design of TENGs by introducing bone marrow mesenchymal stem cells (MSCs) of rats, as support cells, into a silk fibroin (SF)-based scaffold, which was composed of an SF nerve guidance conduit and oriented SF filaments as the conduit lumen filler. The biomaterial SF had been tested to possess good biocompatibility and noncytoxicity with MSCs before the TENG was implanted to bridge a 10-mm-long gap in rat sciatic nerve. Functional and histological assessments showed that at 12 weeks after nerve grafting, TENGs yielded an improved outcome of nerve regeneration and functional recovery, which was better than that achieved by SF scaffolds and close to that achieved by autologous nerve grafts. During 1–4 weeks after nerve grafting, MSCs contained in the TENG significantly accelerated axonal growth, displaying a positive reaction to S-100 (a Schwann cell marker). During 1–3 weeks after nerve grafting, MSCs contained in the TENG led to gene expression upregulation of S100 and several growth factors (brain-derived neurotrophic factor, ciliary neurotrophic factor, and basic fibroblast growth factor). The cell behaviors and neurotrophic functions of MSCs might be responsible for their promoting effects on peripheral nerve regeneration (Yang et al. 2011).
Several papers showed the xenogeneic or allogenic acellular nerve grafts implanted with MSCs promote nerve regeneration effectively in rats or even monkeys (Wang et al. 2008; Zhang et al. 2010; Hu et al. 2007; Jia et al. 2012). So far, most studies directly implant stem cells together with biomaterials for repairing peripheral nerve injury. Other types of stem cells are also mostly implanted in the same way as MSCs mentioned above (Guerout et al. 2014; Lokanathan et al. 2014; Wang et al. 2011; Uemura et al. 2012; Walsh et al. 2009; Ikeda et al. 2014).
1.2.2 Joint Use of Stem Cells, Cytokines, and Scaffolds for Repairing Peripheral Nerve Injury
Upon injury to peripheral nerves, the local presence of growth factors at the injury sites plays a vital and complex role in modulating phenotypic changes of a variety of neural and nonneural cells. Although the endogenous growth factors secreted by neural cells in the distal nerve stump can support axon regeneration, the supportive action may not be sustained indefinitely due to an obvious decline with time in cellular production of growth factors, and hence, the continuous supply of growth factors is critically required, which is mainly dependent on the addition of exogenous growth factors. To date, the most commonly used growth factors belong to two classes: (1) neurotrophins, including NGF, brain-derived neurotrophic factor (BDNF), and neurotrophin-3 (NT-3); and (2) growth factors with neurotrophic actions, including glial cell line-derived neurotrophic factor (GDNF), ciliary neurotrophic factor (CNTF), and fibroblast growth factors (FGFs). The details about their application for neural tissue engineering are available in review papers (Gu et al. 2011; Jiang et al. 2010).
The cell-based delivery of growth factors has been developed. A silicone-based NGC seeded with genetically modified SCs overexpressing fibroblast growth factor-2 (FGF-2) was used to bridge a 15-mm sciatic nerve gap in adult rats. Different FGF isoforms overexpressed by implanted SCs improved both lengths and number of regenerating myelinated axons over different time periods post-grafting (Timmer et al. 2003; Haastert et al. 2008). Likewise, the design of GDNF-transduced Schwann cell grafts for enhancing regeneration of erectile nerves represents the same attempt of cell-based delivery of neurotrophic factors (May et al. 2008, 2013). In addition, a combination of iPSc-derived neurospheres and basic fibroblast growth factor (bFGF)-containing gelatin microspheres was incorporated into a neural scaffold (a synthetic polymer-based, double-layered NGC), and the constructed TENG was used to bridge 5-mm-long sciatic nerve gaps in mice, achieving regenerative outcomes to some degree (Ikeda et al. 2014). The above-mentioned are typical examples of the combined use of support cells and growth factors as biochemical cues within TENGs.
MSCs not only enhance functional outcomes of nerve repair through production of various growth factors, but also act as gene delivery vehicles of growth factors that are released to nerve injury sites for augmenting axonal growth (Lu et al. 2009). For example, it has been reported that transduction of MSCs to overexpress a certain factor, for example, brain-derived neurotrophic factor, could result in significant increase in the extent and diversity of axonal growth in the host nerves (Chen et al. 2005; Lu et al. 2005). Although gene transfer to SCs, MSCs or other stem cells, even to nerve tissues usually facilitates a continuous release of growth factors to encourage peripheral nerve regeneration, it has also found that lentiviral vector-mediated overexpression of glial cell line-derived neurotrophic factor causes trapping of regenerating axons and failure of appropriate target reinnervation during nerve regeneration in a rat sciatic nerve injury model (Tannemaat et al. 2008). It seems that the possible side effects are not negligible in gene-based therapy.
1.2.3 Construction of Tissue-Engineered Nerve Grafts In Vitro for Repairing Peripheral Nerve Injury
TENGs are considered as a promising alternative to autologous nerve grafts used for peripheral nerve repair. The differences between these two types of nerve grafts are mainly in the regenerative microenvironment established by them. To construct ideal TENGs, it is therefore required to develop a better way to introduce biochemical cues into a neural scaffold, as compared to single or combined use of support cells and growth factors.
Tang et al. (2012) have reported that a coculture system of dorsal root ganglia (DRGs) and SCs could give rise to an in vitro cultured nerve equivalent that was likely to mimic the native nerve microenvironment. Their TENGs consisted of an in vitro cultured nerve equivalent residing in a SF-based scaffold were used to bridge a 10-mm sciatic nerve defect in rats. At 12 weeks after nerve grafting, a series of measurements were performed to evaluate the regenerative capacity of these TENGs. The recovery in the motor function of the injured hindlimb in TENG group, as indexed by the SFI value, was close to that in autograft group without significant difference between each other, and prevailed over that in scaffold group. The restoration of electrophysiological properties for 3 grafted groups was reflected in detectable CAMP data, which represented an important measure for the conduction function of peripheral nerves. The comparison in the CMAP amplitude between 3 grafted groups provided further evidence that functional recovery in TENG group was more close to that in autograft group than that in scaffold group. Histological analysis showed that either the regenerated nerve or target gastrocnemius muscle achieved the similar reconstruction, both in qualitative and quantitative aspects, between TENG and autograft groups, and these similar results were significantly better than those in scaffold group. Their findings suggested that more axons might successfully grow through our developed TENG to reach the distal stumps for reinnervation of target muscle. In other words, the incorporation of a nerve equivalent into SF-based scaffold led to an enhanced repair capacity for peripheral nerve injuries. Successful myelination of PNS depends on induction of major protein components of myelin including PMP22, and myelin stability is also sensitive to PMP22 levels (Wrabetz et al. 2006). The high expressions of N-cadherin and PMP22 meant that in their developed nerve equivalent, as in the autologous nerve, dynamic interactions between axons and SCs contributed to the establishment of an ideal microenvironment for nerve regeneration via the increased expression of several bioactive molecules. The introduction of an in vitro cultured nerve equivalent into a scaffold might contribute to establishing a native-like microenvironment for nerve regeneration.
This culture system in vitro is also the same applies in candidate stem cells of the TENG for repairing peripheral nerve injury. The stem cells cocultured with DRGs or seeded onto the biomaterial scaffold in vitro for a period of time provide chance for cells in TENG to communicate with each other or establish a native-like microenvironment effectively.
The rotary bioreactors could influence major cellular events such as differentiation, proliferation, viability, and cell cycle. Introduced by the National Aeronautics and Space Administration (NASA) of the USA in 1987, simulate microgravity (SMG) cultures seem to be ideal for overcoming some drawbacks associated with static culturing systems (Vunjak-Novakovic et al. 1999; Goodwin et al. 1993). For instance, SMG conditions allow the cells to well proliferate in a rotary cell culture system (RCCS) of microgravity environment but at low shear stress and low turbulence environment. SMG culture conditions can provide appropriate microenvironments which have proven advantageous for intercellular communication on tissue-specific cell assembly, cell adhesion, signal transduction, glandular structures, and function (Meyers et al. 2005; Goodwin et al. 1993). In addition, when cells are maintained in a 3D growth environment, they tend to aggregate. SMG culture can enhance cell–cell interactions and supply such 3D growth microenvironment. So, the aggregating growth of cells is an important effect of SMG on some cell lines (Unsworth and Lelkes 1998). SMG promoted porcine liver cells to grow into 3D cell aggregation, which displayed that SMG culture system was suitable for long-term and expanding cell culture (Dabos et al. 2001). The dynamic flow of RCCS might improve nutrient supply and increase metabolic waste removal for the cells in the interior cellular spheres. Thus, comparison with the static culture, rotating simulated microgravity culture environment can show better cellular vitality and function for some cultivated cells. There is also specific application in the construction of TENG in vitro recently (Luo et al. 2014).
Nowadays, for the construction of neural tissue engineering, it needs to be addressed how to obtain adequate and unified standardized conditions to promote the peripheral nerve regeneration. A series of problems should be solved before wide clinical application of the RCCS in tissue engineering technology.
1.2.4 Construction of Acellular Tissue-Engineered Nerve Grafts In Vitro for Repairing Peripheral Nerve Injury
The field of stem cells and regenerative medicine offers considerable promise as a means of delivering new treatments for a wide range of diseases. In order to maximize the effectiveness of cell-based therapies—whether stimulating expansion of endogenous cells or transplanting cells into patients—it is essential to understand the niche signals that regulate stem cell behavior. One of those signals is from the ECM. New technologies have offered insights into how stem cells sense signals from the ECM and how they respond to these signals at the molecular level, which ultimately regulate their fate (Watt and Huck 2013).
To date, however, cellular and molecular therapies directed at peripheral nerve repair have not yet gone beyond the laboratory stage, and their translation to the clinic has been beset with numerous challenges, such as the type and quantity of cells or factors, their delivery, cell viability or factor activity, cell phenotypic stability, timing of treatment, regulatory issues, and high costs (McAllister et al. 2008; Burdick et al. 2013). Therefore, alternate approaches are being developed to substitute an inclusion of support cells or growth factors in a nerve graft.
The ECM is composed of diverse molecules, including proteins, glycoproteins, and glycosaminoglycans, produced by the resident cells in tissues or organs. The composition and structure of ECM are dependent on the phenotype of the resident cells and the function of the tissues or organs. In turn, ECM affects the phenotype and behavior of the resident cells (Bissell et al. 1982; Boudreau et al. 1995; Ingber 1991). It is clear that ECM contains sufficient biological cues to regulate cell phenotype and function in tissues or organs. Based on this knowledge, the mimicking of the native ECM of peripheral nerves within a nerve scaffold seems to be a promising strategy to replace cellular or molecular components added to the scaffold. As an early attempt, nerve scaffolds were prepared with purified individual ECM components, such as collagen, fibrin, laminin, fibronectin, and hyaluronan (Gu et al. 2011; Schmidt and Leach 2003; Deumens et al. 2010; Jiang et al. 2010; Khaing and Schmidt 2012). Some of these scaffolds (e.g., NeuraGen®, NeuroMatrix™, Neuroflex™, NeuraWrap™, and NeuroMend™) have been commercially available and approved by FDA (Kehoe et al. 2012; Meek and Coert 2008). Unfortunately, individual ECM components fail to create an extracellular environment similar to that in vivo within a scaffold in a comprehensive manner because various ECM components have different functions and any individual component cannot substitute for the complete ECM (Ravindran et al. 2012). Indeed, an ECM scaffold can be engineered by using acellular biomaterials, which have been considered a feasible alternative to cellular and/or molecular therapy (Khaing and Schmidt 2012; Burdick et al. 2013). Allogeneic and xenogeneic nerve (or nonnerve) tissues are treated with chemical or thermal decellularization to produce a tissue-derived ECM, which represents one of acellular biomaterials suitable for preparing nerve scaffolds. The resulting tissue-derived ECM scaffolds, commonly called acellular nerve grafts, have been well studied, and some of them are of considerable commercial interest with a FDA-approved product on the market (e.g., Avance®) (Kehoe et al. 2012). As compared to scaffolds prepared with individual ECM components, tissue-derived ECM scaffolds (also called acellular nerve grafts) have a better ability to retain the basic structure of native nerves and promote peripheral nerve regeneration (Whitlock et al. 2009). Mounting evidence indicates that although tissue-derived ECM scaffolds recapitulate biochemical and biophysical cues intrinsic to tissues (Gilbert et al. 2006; Lu et al. 2011; Badylak et al. 2009; Hoshiba et al. 2010; Wolchok and Tresco 2010), they may suffer from several drawbacks, including tissue scarcity, host responses, pathogen transfer, insufficient mechanical properties, and uncontrollable degradation kinetics (Badylak et al. 2009; Cheng et al. 2009; Liao et al. 2010; Skora et al. 2012). In contrast, cultured cell-derived ECM scaffolds have recently attracted attention. They are similar to or even better than tissue-derived ECM scaffolds because the former excludes pathogen transfer when cultured and expanded under pathogen-free conditions and maintains the desired geometry and flexibility when reconstituted with common biomaterials either of synthetic or natural origin (Wolchok and Tresco 2010; Lu et al. 2011; Cheng et al. 2009; Narayanan et al. 2009; Volpato et al. 2013). In consequence, cultured cell-derived ECM scaffolds have been used in some fields of tissue engineering (Wolchok and Tresco 2010; Liao et al. 2010; Lu et al. 2011; Choi et al. 2010). The application of this type of ECM scaffolds, however, has not been fully studied in nerve tissue engineering.
As discussed in detail elsewhere (Williams 2008, 2014), these paradigms move away from the search for biomaterials and structures that passively allow cells to express new ECM; instead, these materials have to be actively involved in the delivery of cues to cells. Indeed, it should be borne in mind that a tissue engineering template should replicate, as far as possible, the niche of those target cells. ECM plays a prominent role in establishing and maintaining an ideal microenvironment for tissue regeneration, and ECM scaffolds are used as a feasible alternative to cellular and molecular therapy in the fields of tissue engineering. Because of their advantages over tissue-derived ECM scaffolds, cultured cell-derived ECM scaffolds are beginning to attract attention, but they have been scarcely studied for peripheral nerve repair.
1.3 Outlook on Stem Cells for Peripheral Nerve Regeneration
The research of peripheral nerve regeneration dates back to many years ago, and the past century has witnessed the accelerated development in peripheral nerve repair strategies, especially a significant progress from early artificial tubular NGCs to current TENGs, but clinical applications of state-of-the-art approaches are still limited and the relevant functional outcomes are not completely satisfactory, largely depending on many factors including the size and location of injured nerves as well as the age of patients. As has been repeatedly indicated by previous literature (Fields et al. 1989), only a clear and thorough understanding of the fundamental events, which occur after nerve injury and during nerve regeneration, could result in a medical breakthrough for peripheral nerve repair. The challenges and possibilities facing surgical management of peripheral nerve injuries are likely to be summarized as two main aspects: (1) survival of the damaged neurons and establishment of the microenvironment that facilitates the neurite outgrowth; (2) accurate pathfinding that guides axons to their targets for appropriate reinnervation. Obviously, peripheral nerve regeneration is determined by the large quantity and high speed of axonal outgrowth, the remyelination of axons by SCs, and the maturity of regenerating nerve fibers. Incorporation of support cells and/or growth factors has proven effective for modulating the above cellular behaviors as evidenced by a wide range of animal experiments, but it has not come to clinical use due to multiple barriers.
Target reinnervation is another pivotal cellular event that predetermines functional recovery following peripheral nerve repair. When a nerve is transected with a damage of basal lamina tubes, axonal sprouts are not restrained to their original basal lamina tubes and axons become unable to grow faithfully along original pathways to their target regions. The mismatch of regenerated axons to their targets may contribute, at least in part, to unsatisfactory functional recovery after nerve grafting. Further elucidation of the pathfinding mechanisms of axons is necessary for developing more efficient methods that will enable nerve regeneration to better mimic natural process of neurogenesis. To conclude, nerve regeneration, even in the simpler PNS (as compared to CNS), is also a quite complex phenomenon that is, to date, still far from being fully understood. However, with the advancement of tissue engineering and regenerative medicine and with an accumulated knowledge in the neuroscience field, one can expect that TENGs with a close similarity to native nerve tissues, both in structure and function, will be eventually developed.
The heterogeneity of the stem cell population could also pose a problem, which may be exacerbated if cell manipulation and growth in culture introduce additional genomic variants, which may affect normal functioning. Culture systems can add heterogeneity to the phenotype and genotype, thus complicating selection criteria for transplantation (Bara et al. 2014). Incorrect or incomplete differentiation can also be a concern, as in the case of improperly differentiated endothelium from transplanted PSCs, which can lead to maladapted fibrosis and affect organ function (Ginsberg et al. 2012). Although these potential risks can be partly investigated and addressed by transplantation of cells into suitable immunosuppressed animals, these models may only approximate the human disease and often do not account for the intact human immune system.
Despite the gold standard for peripheral nerve gap repair, autologous nerve grafts fail to achieve an entirely satisfactory restoration of function after they are implanted. Intended to supplement and replace autologous nerve grafts, TENGs should be able to compete with or even surpass autologous nerve grafts in the outcomes of nerve regeneration and functional recovery. Therefore, although the past several decades have witnessed great advance from the earliest nerve tube to the state-of-the-art TENG, neural tissue engineering needs further significant progress toward the development of ideal TENGs and their translation to clinical applications.
It has to be recognized that many materials have been tested and used for TENGs without clear resolution of the optimal structure, a fact by itself which indicates that there is much to learn about their performance. We have delineated here the experimental use of various inorganic and indeed metallic materials, which go beyond the framework of traditional biomaterials, for neural template fabrication. We have also addressed the favorable features of some nanostructured neural scaffolds due to their topographical resemblance of natural ECM architecture. This is a key issue in light of the need for the template to replicate the niche of the target cells. It is unlikely that conventional materials, including most synthetic polymers, will meet the strict requirements of this cell niche concept; the use of decellularized natural tissues and various forms of biopolymers, including hydrogel forms of both proteins and polysaccharides, are clearly very important here. We have described the incorporation of SKPs and AMSCs (as support cells) into neural templates in the construction of TENGs and illustrate newly developed delivery systems for growth factors within them. Certainly, there are still other prospective cues that can be integrated within TENGs, such as molecular inflammatory mediators (Kiefer et al. 2001; Wang et al. 2012; Camara-Lemarroy et al. 2010; Tang et al. 2013), bioactive peptides (Schense et al. 2000; Cheng et al. 2014), and antioxidant reagents (Shen et al. 2007; Wilson et al. 2007). These extra cues are ready to demonstrate their promising applications in neural tissue engineering.
Although the promise of cell therapy for tissue regeneration is exciting, troublesome details persist, such as cell selection, delivery, viability, and phenotypic stability, in addition to timing of treatments, regulatory issues, and high costs (McAllister et al. 2008; Mummery et al. 2010). Nowadays, TENGs used in the clinic, however, are limited to those composed of a neural scaffold alone without any biochemical components due to the presence of various barriers. The construction of an effective TENG should be considered as a complex scientific and engineering problem that involves multifaceted interactions between a diverse array of physicochemical and biological cues, which have been and are still being elucidated within the constantly updated knowledge of peripheral nerve injury and regeneration. The various cues have distinctive effects on the performance of TENGs, but it is necessary to understand and implement the orchestration of the different cue-induced effects. A considerable number of comparative studies must be conducted to decipher. For example, which are more prominent cues, whether and how different cues are interrelated to and interfere with each other. Obviously, the research on these issues will benefit from an improved insight into the molecular events and mechanisms that underlie peripheral nerve injury and regeneration (Navarro et al. 2007; Raimondo et al. 2011; Napoli et al. 2012; Fricker et al. 2013). So far, not only animal models that have TENGs achieved good results, but clinical trials with TENGs to treat human patients with peripheral nerve injury have also met with a certain degree of success (Kehoe et al. 2012; Meek and Coert 2008; Lin et al. 2013; Rinkel et al. 2013; Zhang et al. 2013). Especially, many commercial available products of neural scaffolds have been used in the clinic with promising outcomes (Gu et al. 2011; Kehoe et al. 2012; Meek and Coert 2008).
Although it may be possible to promote the activity of endogenous stem cells to enhance tissue repair, it is becoming increasingly clear that as much attention needs to be paid to the environment in which the cells reside as to the nature of the cells themselves (Watt and Hogan 2000).
The physical niche for any somatic stem cell is composed of two basic components: an acellular ECM and local cellular constituents. The molecular components of the matrix have a profound effect on the biology of stem cells in regulating their quiescence, proliferation, symmetric and asymmetric divisions, and fate (Watt and Huck 2013); conversely, stem cells typically produce constituents of their own matrix. The cells found in the niche, such as differentiated cells of the tissue, interstitial mesenchymal cells, and cellular components of the vasculature, may influence stem cell functionality by direct contact or by locally secreted paracrine factors. Despite considerable success of acellular nerve grafts in supporting peripheral nerve regeneration (Khaing and Schmidt 2012; Krekoski et al. 2001; Kim et al. 2004), they are still associated with several drawbacks, which inspire the emergence of cultured cell-derived ECM scaffolds. The developed nerve scaffolds might have not these drawbacks because (1) the isolated SCs can be in vitro expanded, and even commercially available SC cell lines are also used to obtain ECM; (2) SCs can be cultured in a pathogen-free medium; and (3) the joint use of SC-derived ECM and natural biomaterials allows our developed scaffolds to have robust mechanical properties and exhibit tailored degradation profiles. The safety examination indicated that adverse effects, including adverse immune responses, were not observed in animals after bridging rat sciatic nerve gap with different scaffolds, which provided a necessary basis for the translation of our developed scaffolds to the clinic. Targeted activation of endogenous stem cells to repair the tissues in which they reside requires an understanding of the molecular pathways that normally control stem cell function and how those signals might have changed in the setting of injury or disease, perhaps rendering the stem cells less responsive to extrinsic cues.
A major challenge for the development of targeted therapeutics to enhance endogenous stem cell function is the modeling, in vitro, of the environment that sustains the stem cells in a state of reduced responsiveness in vivo. Typically, plating stem cells in culture induces them to begin dividing and differentiating, thus altering the cellular state that would be the therapeutic target (Dimmeler et al. 2014). To push the translation of stem cell application for peripheral nerve regeneration into the clinic, we anticipate that the most appropriate stem cells and other biological cues with a close proximity to the regenerative microenvironment of the PNS will be developed.
References
Abdallah BM, Kassem M. Human mesenchymal stem cells: from basic biology to clinical applications. Gene Ther. 2008;15(2):109–16.
Alessandri G, Emanueli C, Madeddu P. Genetically engineered stem cell therapy for tissue regeneration. Ann NY Acad Sci. 2004;1015:271–84.
Andrews MR, Stelzner DJ. Modification of the regenerative response of dorsal column axons by olfactory ensheathing cells or peripheral axotomy in adult rat. Exp Neurol. 2004;190(2):311–27.
Artico M, et al. Birthday of peripheral nervous system surgery: the contribution of Gabriele Ferrara (1543–1627). Neurosurgery. 1996;39(2):380–2 (discussion 382–3).
Asplund M, et al. Incidence of traumatic peripheral nerve injuries and amputations in Sweden between 1998 and 2006. Neuroepidemiology. 2009;32(3):217–28.
Au E, et al. SPARC from olfactory ensheathing cells stimulates Schwann cells to promote neurite outgrowth and enhances spinal cord repair. J Neurosci. 2007;27(27):7208–21.
Badylak SF, Freytes DO, Gilbert TW. Extracellular matrix as a biological scaffold material: structure and function. Acta Biomater. 2009;5(1):1–13.
Bara JJ, et al. Concise review: bone marrow-derived mesenchymal stem cells change phenotype following in vitro culture: implications for basic research and the clinic. Stem Cells. 2014;32(7):1713–23.
Battiston B, et al. Chapter 1: Peripheral nerve repair and regeneration research: a historical note. Int Rev Neurobiol. 2009;87:1–7.
Bianco P, et al. Bone marrow stromal stem cells: nature, biology, and potential applications. Stem Cells. 2001;19(3):180–92.
Bissell MJ, Hall HG, Parry G. How does the extracellular matrix direct gene expression? J Theor Biol. 1982;99(1):31–68.
Blazejewska EA, et al. Corneal limbal microenvironment can induce transdifferentiation of hair follicle stem cells into corneal epithelial-like cells. Stem Cells. 2009;27(3):642–52.
Borlongan CV, et al. Bone marrow grafts restore cerebral blood flow and blood brain barrier in stroke rats. Brain Res. 2004;1010(1–2):108–16.
Boudreau N, Myers C, Bissell MJ. From laminin to lamin: regulation of tissue-specific gene expression by the ECM. Trends Cell Biol. 1995;5(1):1–4.
Burdick JA, et al. Acellular biomaterials: an evolving alternative to cell-based therapies. Sci Transl Med. 2013;5(176):176ps4.
Camara-Lemarroy CR, Guzman-de la Garza FJ, Fernandez-Garza NE. Molecular inflammatory mediators in peripheral nerve degeneration and regeneration. NeuroImmunoModulation. 2010;17(5):314–24.
Cao L, et al. Olfactory ensheathing cells promote migration of Schwann cells by secreted nerve growth factor. Glia. 2007;55(9):897–904.
Caplan AI, Dennis JE. Mesenchymal stem cells as trophic mediators. J Cell Biochem. 2006;98(5):1076–84.
Carriel V, et al. Combination of fibrin-agarose hydrogels and adipose-derived mesenchymal stem cells for peripheral nerve regeneration. J Neural Eng. 2013;10(2):026022.
Chalfoun CT, Wirth GA, Evans GR. Tissue engineered nerve constructs: where do we stand? J Cell Mol Med. 2006;10(2):309–17.
Chen X, et al. Ischemic rat brain extracts induce human marrow stromal cell growth factor production. Neuropathology. 2002;22(4):275–9.
Chen Q, et al. Protective effects of bone marrow stromal cell transplantation in injured rodent brain: synthesis of neurotrophic factors. J Neurosci Res. 2005;80(5):611–9.
Chen X, et al. Study of in vivo differentiation of rat bone marrow stromal cells into schwann cell-like cells. Microsurgery. 2006;26(2):111–5.
Chen CJ, et al. Transplantation of bone marrow stromal cells for peripheral nerve repair. Exp Neurol. 2007;204(1):443–53.
Chen Z, et al. Skin-derived precursors as a source of progenitors for cutaneous nerve regeneration. Stem Cells. 2012;30(10):2261–70.
Cheng NC, et al. Chondrogenic differentiation of adipose-derived adult stem cells by a porous scaffold derived from native articular cartilage extracellular matrix. Tissue Eng Part A. 2009a;15(2):231–41.
Cheng HW, et al. Decellularization of chondrocyte-encapsulated collagen microspheres: a three-dimensional model to study the effects of acellular matrix on stem cell fate. Tissue Eng Part C Methods. 2009b;15(4):697–706.
Cheng Q, et al. Neurotrophic and neuroprotective actions of achyranthes bidentata polypeptides on cultured dorsal root ganglia of rats and on crushed common peroneal nerve of rabbits. Neurosci Lett. 2014;562:7–12.
Choi KH, et al. The chondrogenic differentiation of mesenchymal stem cells on an extracellular matrix scaffold derived from porcine chondrocytes. Biomaterials. 2010;31(20):5355–65.
Chopp M, Li Y. Treatment of neural injury with marrow stromal cells. Lancet Neurol. 2002;1(2):92–100.
Chopp M, et al. Spinal cord injury in rat: treatment with bone marrow stromal cell transplantation. NeuroReport. 2000;11(13):3001–5.
Craff MN, et al. Embryonic stem cell-derived motor neurons preserve muscle after peripheral nerve injury. Plast Reconstr Surg. 2007;119(1):235–45.
Crane JF, Trainor PA. Neural crest stem and progenitor cells. Annu Rev Cell Dev Biol. 2006;22:267–86.
Crigler L, et al. Human mesenchymal stem cell subpopulations express a variety of neuro-regulatory molecules and promote neuronal cell survival and neuritogenesis. Exp Neurol. 2006;198(1):54–64.
Cuevas P, et al. Peripheral nerve regeneration by bone marrow stromal cells. Neurol Res. 2002;24(7):634–8.
Cui L, et al. Transplantation of embryonic stem cells improves nerve repair and functional recovery after severe sciatic nerve axotomy in rats. Stem Cells. 2008;26(5):1356–65.
Dabos KJ, et al. The simulated microgravity environment maintains key metabolic functions and promotes aggregation of primary porcine hepatocytes. Biochim Biophys Acta. 2001;1526(2):119–30.
Deryugina EI, Muller-Sieburg CE. Stromal cells in long-term cultures: keys to the elucidation of hematopoietic development? Crit Rev Immunol. 1993;13(2):115–50.
Deumens R, et al. Neurite outgrowth promoting effects of enriched and mixed OEC/ONF cultures. Neurosci Lett. 2006;397(1–2):20–4.
Deumens R, et al. Repairing injured peripheral nerves: bridging the gap. Prog Neurobiol. 2010;92(3):245–76.
Dezawa M. Insights into autotransplantation: the unexpected discovery of specific induction systems in bone marrow stromal cells. Cell Mol Life Sci. 2006;63(23):2764–72.
Dezawa M, et al. Sciatic nerve regeneration in rats induced by transplantation of in vitro differentiated bone-marrow stromal cells. Eur J Neurosci. 2001;14(11):1771–6.
Dezawa M, et al. Specific induction of neuronal cells from bone marrow stromal cells and application for autologous transplantation. J Clin Invest. 2004;113(12):1701–10.
di Summa PG, et al. Long-term in vivo regeneration of peripheral nerves through bioengineered nerve grafts. Neuroscience. 2011;181:278–91.
Dimmeler S, et al. Translational strategies and challenges in regenerative medicine. Nat Med. 2014;20(8):814–21.
Ding F, et al. Use of tissue-engineered nerve grafts consisting of a chitosan/poly(lactic-co-glycolic acid)-based scaffold included with bone marrow mesenchymal cells for bridging 50-mm dog sciatic nerve gaps. Tissue Eng Part A. 2010;16(12):3779–90.
Dombrowski MA, et al. Myelination and nodal formation of regenerated peripheral nerve fibers following transplantation of acutely prepared olfactory ensheathing cells. Brain Res. 2006;1125(1):1–8.
Dupouey P, et al. Immunochemical studies of myelin basic protein in shiverer mouse devoid of major dense line of myelin. Neurosci Lett. 1979;12(1):113–8.
Erba P, et al. Regeneration potential and survival of transplanted undifferentiated adipose tissue-derived stem cells in peripheral nerve conduits. J Plast Reconstr Aesthet Surg. 2010;63(12):e811–7.
Fairless R, Barnett SC. Olfactory ensheathing cells: their role in central nervous system repair. Int J Biochem Cell Biol. 2005;37(4):693–9.
Fernandes KJ, et al. A dermal niche for multipotent adult skin-derived precursor cells. Nat Cell Biol. 2004;6(11):1082–93.
Fickert S, Fiedler J, Brenner RE. Identification, quantification and isolation of mesenchymal progenitor cells from osteoarthritic synovium by fluorescence automated cell sorting. Osteoarthritis Cartilage. 2003;11(11):790–800.
Fields RD, et al. Nerve regeneration through artificial tubular implants. Prog Neurobiol. 1989;33(2):87–134.
Franchi S, et al. Intravenous neural stem cells abolish nociceptive hypersensitivity and trigger nerve regeneration in experimental neuropathy. Pain. 2012;153(4):850–61.
Fricker FR, et al. Axonal neuregulin 1 is a rate limiting but not essential factor for nerve remyelination. Brain. 2013;136(Pt 7):2279–97.
Gilbert TW, Sellaro TL, Badylak SF. Decellularization of tissues and organs. Biomaterials. 2006;27(19):3675–83.
Ginsberg M, et al. Efficient direct reprogramming of mature amniotic cells into endothelial cells by ETS factors and TGFbeta suppression. Cell. 2012;151(3):559–75.
Gokhan S, Mehler MF. Basic and clinical neuroscience applications of embryonic stem cells. Anat Rec. 2001;265(3):142–56.
Goodwin TJ, et al. Reduced shear stress: a major component in the ability of mammalian tissues to form three-dimensional assemblies in simulated microgravity. J Cell Biochem. 1993a;51(3):301–11.
Goodwin TJ, et al. Rotating-wall vessel coculture of small intestine as a prelude to tissue modeling: aspects of simulated microgravity. Proc Soc Exp Biol Med. 1993b;202(2):181–92.
Gu Y, et al. Neurotrophic actions of bone marrow stromal cells on primary culture of dorsal root ganglion tissues and neurons. J Mol Neurosci. 2010;40(3):332–41.
Gu X, et al. Construction of tissue engineered nerve grafts and their application in peripheral nerve regeneration. Prog Neurobiol. 2011;93(2):204–30.
Gu JH, et al. Transplantation of adipose derived stem cells for peripheral nerve regeneration in sciatic nerve defects of the rat. Curr Stem Cell Res Ther. 2012;7(5):347–55.
Guerout N, et al. Transplantation of olfactory ensheathing cells promotes axonal regeneration and functional recovery of peripheral nerve lesion in rats. Muscle Nerve. 2011;43(4):543–51.
Guerout N, et al. Transplantation of olfactory ensheathing cells to evaluate functional recovery after peripheral nerve injury. J Vis Exp. 2014;84:e50590.
Guo BF, Dong MM. Application of neural stem cells in tissue-engineered artificial nerve. Otolaryngol Head Neck Surg. 2009;140(2):159–64.
Haastert K, et al. The effects of FGF-2 gene therapy combined with voluntary exercise on axonal regeneration across peripheral nerve gaps. Neurosci Lett. 2008;443(3):179–83.
Heine W, et al. Transplanted neural stem cells promote axonal regeneration through chronically denervated peripheral nerves. Exp Neurol. 2004;189(2):231–40.
Hofstetter CP, et al. Marrow stromal cells form guiding strands in the injured spinal cord and promote recovery. Proc Natl Acad Sci USA. 2002;99(4):2199–204.
Honkanen H, et al. Isolation, purification and expansion of myelination-competent, neonatal mouse Schwann cells. Eur J Neurosci. 2007;26(4):953–64.
Horwitz EM, Dominici M. How do mesenchymal stromal cells exert their therapeutic benefit? Cytotherapy. 2008;10(8):771–4.
Horwitz EM, et al. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: implications for cell therapy of bone. Proc Natl Acad Sci USA. 2002;99(13):8932–7.
Hoshiba T, et al. Decellularized matrices for tissue engineering. Expert Opin Biol Ther. 2010;10(12):1717–28.
Hsu SH, Su CH, Chiu IM. A novel approach to align adult neural stem cells on micropatterned conduits for peripheral nerve regeneration: a feasibility study. Artif Organs. 2009;33(1):26–35.
Hu J, et al. Repair of extended peripheral nerve lesions in rhesus monkeys using acellular allogenic nerve grafts implanted with autologous mesenchymal stem cells. Exp Neurol. 2007;204(2):658–66.
Hu N, et al. Long-term outcome of the repair of 50 mm long median nerve defects in rhesus monkeys with marrow mesenchymal stem cells-containing, chitosan-based tissue engineered nerve grafts. Biomaterials. 2013;34(1):100–11.
Ikeda M, et al. Acceleration of peripheral nerve regeneration using nerve conduits in combination with induced pluripotent stem cell technology and a basic fibroblast growth factor drug delivery system. J Biomed Mater Res A. 2014;102(5):1370–8.
Ingber D. Extracellular matrix and cell shape: potential control points for inhibition of angiogenesis. J Cell Biochem. 1991;47(3):236–41.
Jahoda CA, Horne KA, Oliver RF. Induction of hair growth by implantation of cultured dermal papilla cells. Nature. 1984;311(5986):560–2.
Jakob H. Stem cells and embryo-derived cell lines: tools for study of gene expression. Cell Differ. 1984;15(2–4):77–80.
Jia H, et al. Sciatic nerve repair by acellular nerve xenografts implanted with BMSCs in rats xenograft combined with BMSCs. Synapse. 2012;66(3):256–69.
Jiang X, et al. Current applications and future perspectives of artificial nerve conduits. Exp Neurol. 2010;223(1):86–101.
Jinno H, et al. Convergent genesis of an adult neural crest-like dermal stem cell from distinct developmental origins. Stem Cells. 2010;28(11):2027–40.
Johnson A, Dorshkind K. Stromal cells in myeloid and lymphoid long-term bone marrow cultures can support multiple hemopoietic lineages and modulate their production of hemopoietic growth factors. Blood. 1986;68(6):1348–54.
Johnson EO, Soucacos PN. Nerve repair: experimental and clinical evaluation of biodegradable artificial nerve guides. Injury. 2008;39(Suppl 3):S30–6.
Johnson EO, Zoubos AB, Soucacos PN. Regeneration and repair of peripheral nerves. Injury. 2005;36(Suppl 4):S24–9.
Johnson EO, Charchanti A, Soucacos PN. Nerve repair: experimental and clinical evaluation of neurotrophic factors in peripheral nerve regeneration. Injury. 2008;39(Suppl 3):S37–42.
Kehoe S, Zhang XF, Boyd D. FDA approved guidance conduits and wraps for peripheral nerve injury: a review of materials and efficacy. Injury. 2012;43(5):553–72.
Keilhoff G, et al. Peripheral nerve tissue engineering: autologous Schwann cells vs. transdifferentiated mesenchymal stem cells. Tissue Eng. 2006a;12(6):1451–65.
Keilhoff G, et al. Transdifferentiated mesenchymal stem cells as alternative therapy in supporting nerve regeneration and myelination. Cell Mol Neurobiol. 2006b;26(7–8):1235–52.
Kelsey JL, et al. Upper extremity disorders: frequency, impact, and cost. New York, NY: Churchill Livingstone; 1997.
Khaing ZZ, Schmidt CE. Advances in natural biomaterials for nerve tissue repair. Neurosci Lett. 2012;519(2):103–14.
Khuong HT, et al. Skin derived precursor Schwann cells improve behavioral recovery for acute and delayed nerve repair. Exp Neurol. 2014;254:168–79.
Kiefer R, et al. The role of macrophages in immune-mediated damage to the peripheral nervous system. Prog Neurobiol. 2001;64(2):109–27.
Kim BS, Yoo JJ, Atala A. Peripheral nerve regeneration using acellular nerve grafts. J Biomed Mater Res A. 2004;68(2):201–9.
Kingham PJ, et al. Adipose-derived stem cells differentiate into a Schwann cell phenotype and promote neurite outgrowth in vitro. Exp Neurol. 2007;207(2):267–74.
Kocsis JD, et al. Cell transplantation of peripheral-myelin-forming cells to repair the injured spinal cord. J Rehabil Res Dev. 2002;39(2):287–98.
Kopen GC, Prockop DJ, Phinney DG. Marrow stromal cells migrate throughout forebrain and cerebellum, and they differentiate into astrocytes after injection into neonatal mouse brains. Proc Natl Acad Sci USA. 1999;96(19):10711–6.
Krekoski CA, et al. Axonal regeneration into acellular nerve grafts is enhanced by degradation of chondroitin sulfate proteoglycan. J Neurosci. 2001;21(16):6206–13.
Kunter U, et al. Transplanted mesenchymal stem cells accelerate glomerular healing in experimental glomerulonephritis. J Am Soc Nephrol. 2006;17(8):2202–12.
Lee RH, et al. Multipotent stromal cells from human marrow home to and promote repair of pancreatic islets and renal glomeruli in diabetic NOD/scid mice. Proc Natl Acad Sci USA. 2006;103(46):17438–43.
Lee G, et al. Isolation and directed differentiation of neural crest stem cells derived from human embryonic stem cells. Nat Biotechnol. 2007;25(12):1468–75.
Liao J, et al. Bioactive polymer/extracellular matrix scaffolds fabricated with a flow perfusion bioreactor for cartilage tissue engineering. Biomaterials. 2010;31(34):8911–20.
Lin MY, Manzano G, Gupta R. Nerve allografts and conduits in peripheral nerve repair. Hand Clin. 2013;29(3):331–48.
Liu CH, Hwang SM. Cytokine interactions in mesenchymal stem cells from cord blood. Cytokine. 2005;32(6):270–9.
Lokanathan Y, et al. Olfactory ensheathing cells seeded muscle-stuffed vein as nerve conduit for peripheral nerve repair: a nerve conduction study. J Biosci Bioeng. 2014;118(2):231–4.
Lu P, Jones LL, Tuszynski MH. BDNF-expressing marrow stromal cells support extensive axonal growth at sites of spinal cord injury. Exp Neurol. 2005;191(2):344–60.
Lu J, et al. Adult bone marrow cells differentiate into neural phenotypes and improve functional recovery in rats following traumatic brain injury. Neurosci Lett. 2006;398(1–2):12–7.
Lu L, et al. Morphological and functional characterization of predifferentiation of myelinating glia-like cells from human bone marrow stromal cells through activation of F3/Notch signaling in mouse retina. Stem Cells. 2008;26(2):580–90.
Lu Z, et al. Overexpression of CNTF in Mesenchymal Stem Cells reduces demyelination and induces clinical recovery in experimental autoimmune encephalomyelitis mice. J Neuroimmunol. 2009;206(1–2):58–69.
Lu H, et al. Autologous extracellular matrix scaffolds for tissue engineering. Biomaterials. 2011a;32(10):2489–99.
Lu H, et al. Cultured cell-derived extracellular matrix scaffolds for tissue engineering. Biomaterials. 2011b;32(36):9658–66.
Luo H, et al. Tissue-engineered nerve constructs under a microgravity system for peripheral nerve regeneration. Tissue Eng Part A. 2014;21:267.
Mackinnon SE, Hudson AR. Clinical application of peripheral nerve transplantation. Plast Reconstr Surg. 1992;90(4):695–9.
Malin D, et al. The extracellular-matrix protein matrilin 2 participates in peripheral nerve regeneration. J Cell Sci. 2009;122(Pt 7):995–1004.
Marchesi C, et al. Skin-derived stem cells transplanted into resorbable guides provide functional nerve regeneration after sciatic nerve resection. Glia. 2007;55(4):425–38.
May F, et al. GDNF-transduced Schwann cell grafts enhance regeneration of erectile nerves. Eur Urol. 2008;54(5):1179–87.
May F, et al. Schwann cell-mediated delivery of glial cell line-derived neurotrophic factor restores erectile function after cavernous nerve injury. Int J Urol. 2013;20(3):344–8.
McAllister TN, et al. Cell-based therapeutics from an economic perspective: primed for a commercial success or a research sinkhole? Regen Med. 2008;3(6):925–37.
McKenzie IA, et al. Skin-derived precursors generate myelinating Schwann cells for the injured and dysmyelinated nervous system. J Neurosci. 2006;26(24):6651–60.
Meek MF, Coert JH. US Food and Drug Administration/Conformit Europe-approved absorbable nerve conduits for clinical repair of peripheral and cranial nerves. Ann Plast Surg. 2008;60(4):466–72.
Meyers VE, et al. RhoA and cytoskeletal disruption mediate reduced osteoblastogenesis and enhanced adipogenesis of human mesenchymal stem cells in modeled microgravity. J Bone Miner Res. 2005;20(10):1858–66.
Mimura T, et al. Peripheral nerve regeneration by transplantation of bone marrow stromal cell-derived Schwann cells in adult rats. J Neurosurg. 2004;101(5):806–12.
Minguell JJ, Erices A. Mesenchymal stem cells and the treatment of cardiac disease. Exp Biol Med (Maywood). 2006;231(1):39–49.
Mohammadi R, Azizi S, Amini K. Effects of undifferentiated cultured omental adipose-derived stem cells on peripheral nerve regeneration. J Surg Res. 2013;180(2):e91–7.
Mohanna PN, et al. A composite poly-hydroxybutyrate-glial growth factor conduit for long nerve gap repairs. J Anat. 2003;203(6):553–65.
Morrison SJ, et al. Prospective identification, isolation by flow cytometry, and in vivo self-renewal of multipotent mammalian neural crest stem cells. Cell. 1999;96(5):737–49.
Muir D. The potentiation of peripheral nerve sheaths in regeneration and repair. Exp Neurol. 2010;223(1):102–11.
Mummery CL, Davis RP, Krieger JE. Challenges in using stem cells for cardiac repair. Sci Transl Med. 2010;2(27):27ps17.
Munoz JR, et al. Human stem/progenitor cells from bone marrow promote neurogenesis of endogenous neural stem cells in the hippocampus of mice. Proc Natl Acad Sci USA. 2005;102(50):18171–6.
Munoz-Elias G, Woodbury D, Black IB. Marrow stromal cells, mitosis, and neuronal differentiation: stem cell and precursor functions. Stem Cells. 2003;21(4):437–48.
Napoli I, et al. A central role for the ERK-signaling pathway in controlling Schwann cell plasticity and peripheral nerve regeneration in vivo. Neuron. 2012;73(4):729–42.
Narayanan K, et al. Three-dimensional reconstituted extracellular matrix scaffolds for tissue engineering. Biomaterials. 2009;30(26):4309–17.
Navarro X, Vivo M, Valero-Cabre A. Neural plasticity after peripheral nerve injury and regeneration. Prog Neurobiol. 2007;82(4):163–201.
Neuhuber B, et al. Axon growth and recovery of function supported by human bone marrow stromal cells in the injured spinal cord exhibit donor variations. Brain Res. 2005;1035(1):73–85.
Noble J, et al. Analysis of upper and lower extremity peripheral nerve injuries in a population of patients with multiple injuries. J Trauma. 1998;45(1):116–22.
Orbay H, et al. Differentiated and undifferentiated adipose-derived stem cells improve function in rats with peripheral nerve gaps. J Plast Reconstr Aesthet Surg. 2012;65(5):657–64.
Ortiguela ME, Wood MB, Cahill DR. Anatomy of the sural nerve complex. J Hand Surg Am. 1987;12(6):1119–23.
Ortiz LA, et al. Mesenchymal stem cell engraftment in lung is enhanced in response to bleomycin exposure and ameliorates its fibrotic effects. Proc Natl Acad Sci USA. 2003;100(14):8407–11.
Park IH, et al. Reprogramming of human somatic cells to pluripotency with defined factors. Nature. 2008;451(7175):141–6.
Park BW, et al. Peripheral nerve regeneration using autologous porcine skin-derived mesenchymal stem cells. J Tissue Eng Regen Med. 2012;6(2):113–24.
Phinney DG, Prockop DJ. Concise review: mesenchymal stem/multipotent stromal cells: the state of transdifferentiation and modes of tissue repair–current views. Stem Cells. 2007;25(11):2896–902.
Radtke C, Kocsis JD. Peripheral nerve injuries and transplantation of olfactory ensheathing cells for axonal regeneration and remyelination: fact or fiction? Int J Mol Sci. 2012;13(10):12911–24.
Radtke C, et al. Transplantation of olfactory ensheathing cells enhances peripheral nerve regeneration after microsurgical nerve repair. Brain Res. 2009;1254:10–7.
Raimondo S, et al. Perspectives in regeneration and tissue engineering of peripheral nerves. Ann Anat. 2011;193(4):334–40.
Rajaram A, Chen XB, Schreyer DJ. Strategic design and recent fabrication techniques for bioengineered tissue scaffolds to improve peripheral nerve regeneration. Tissue Eng Part B Rev. 2012;18(6):454–67.
Rao MS, Anderson DJ. Immortalization and controlled in vitro differentiation of murine multipotent neural crest stem cells. J Neurobiol. 1997;32(7):722–46.
Ravindran S, et al. Biomimetic extracellular matrix-incorporated scaffold induces osteogenic gene expression in human marrow stromal cells. Tissue Eng Part A. 2012;18(3–4):295–309.
Rider DA, et al. Autocrine fibroblast growth factor 2 increases the multipotentiality of human adipose-derived mesenchymal stem cells. Stem Cells. 2008;26(6):1598–608.
Ringden O, et al. Mesenchymal stem cells for treatment of therapy-resistant graft-versus-host disease. Transplantation. 2006;81(10):1390–7.
Rinkel WD, et al. What is evidence based in the reconstruction of digital nerves? A systematic review. J Plast Reconstr Aesthet Surg. 2013;66(2):151–64.
Robinson LR. Traumatic injury to peripheral nerves. Muscle Nerve. 2000;23(6):863–73.
Rumsey JW, et al. Node of Ranvier formation on motoneurons in vitro. Biomaterials. 2009;30(21):3567–72.
Salzer JL, Brophy PJ, Peles E. Molecular domains of myelinated axons in the peripheral nervous system. Glia. 2008;56(14):1532–40.
Sanchez-Ramos J, et al. Adult bone marrow stromal cells differentiate into neural cells in vitro. Exp Neurol. 2000;164(2):247–56.
Sauka-Spengler T, Bronner-Fraser M. A gene regulatory network orchestrates neural crest formation. Nat Rev Mol Cell Biol. 2008;9(7):557–68.
Schense JC, et al. Enzymatic incorporation of bioactive peptides into fibrin matrices enhances neurite extension. Nat Biotechnol. 2000;18(4):415–9.
Schmidt CE, Leach JB. Neural tissue engineering: strategies for repair and regeneration. Annu Rev Biomed Eng. 2003;5:293–347.
Scholz T, et al. Neuronal differentiation of human adipose tissue-derived stem cells for peripheral nerve regeneration in vivo. Arch Surg. 2011;146(6):666–74.
Schuldiner M, et al. Effects of eight growth factors on the differentiation of cells derived from human embryonic stem cells. Proc Natl Acad Sci USA. 2000;97(21):11307–12.
Schuldiner M, et al. Induced neuronal differentiation of human embryonic stem cells. Brain Res. 2001;913(2):201–5.
Seidlits SK, Lee JY, Schmidt CE. Nanostructured scaffolds for neural applications. Nanomedicine (Lond). 2008;3(2):183–99.
Shakhbazau A, et al. Sensory recovery after cell therapy in peripheral nerve repair: effects of naive and skin precursor-derived Schwann cells. J Neurosurg. 2014;121(2):423–31.
Shen Y, et al. Neuroprotective effect of carnosine on necrotic cell death in PC12 cells. Neurosci Lett. 2007;414(2):145–9.
Shen CC, Yang YC, Liu BS. Peripheral nerve repair of transplanted undifferentiated adipose tissue-derived stem cells in a biodegradable reinforced nerve conduit. J Biomed Mater Res A. 2012;100(1):48–63.
Shi Y, et al. Transplantation of neural stem cells overexpressing glia-derived neurotrophic factor promotes facial nerve regeneration. Acta Otolaryngol. 2009;129(8):906–14.
Skora J, et al. Evaluation of the humoral and cellular immune responses after implantation of a PTFE vascular prosthesis. Postepy Hig Med Dosw (Online). 2012;66:469–74.
Su Z, et al. Olfactory ensheathing cells: the primary innate immunocytes in the olfactory pathway to engulf apoptotic olfactory nerve debris. Glia. 2013;61(4):490–503.
Suganuma S, et al. Uncultured adipose-derived regenerative cells promote peripheral nerve regeneration. J Orthop Sci. 2013;18(1):145–51.
Sun F, et al. Combined use of decellularized allogeneic artery conduits with autologous transdifferentiated adipose-derived stem cells for facial nerve regeneration in rats. Biomaterials. 2011;32(32):8118–28.
Suzuki H, et al. Neurospheres induced from bone marrow stromal cells are multipotent for differentiation into neuron, astrocyte, and oligodendrocyte phenotypes. Biochem Biophys Res Commun. 2004;322(3):918–22.
Takahashi K, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131(5):861–72.
Tan CW, et al. Sciatic nerve repair with tissue engineered nerve: olfactory ensheathing cells seeded poly (lactic-co-glygolic acid) conduit in an animal model. Indian J Orthop. 2013;47(6):547–52.
Tang X, et al. Bridging peripheral nerve defects with a tissue engineered nerve graft composed of an in vitro cultured nerve equivalent and a silk fibroin-based scaffold. Biomaterials. 2012;33(15):3860–7.
Tang X, et al. Signaling pathways regulating dose-dependent dual effects of TNF-alpha on primary cultured Schwann cells. Mol Cell Biochem. 2013;378(1–2):237–46.
Tannemaat MR, et al. Differential effects of lentiviral vector-mediated overexpression of nerve growth factor and glial cell line-derived neurotrophic factor on regenerating sensory and motor axons in the transected peripheral nerve. Eur J Neurosci. 2008;28(8):1467–79.
Taylor CA, et al. The incidence of peripheral nerve injury in extremity trauma. Am J Phys Med Rehabil. 2008;87(5):381–5.
Timmer M, et al. Axonal regeneration across long gaps in silicone chambers filled with Schwann cells overexpressing high molecular weight FGF-2. Cell Transplant. 2003;12(3):265–77.
Tohill M, Terenghi G. Stem-cell plasticity and therapy for injuries of the peripheral nervous system. Biotechnol Appl Biochem. 2004;40(Pt 1):17–24.
Toma JG, et al. Isolation of multipotent adult stem cells from the dermis of mammalian skin. Nat Cell Biol. 2001;3(9):778–84.
Tomita K, et al. Differentiated adipose-derived stem cells promote myelination and enhance functional recovery in a rat model of chronic denervation. J Neurosci Res. 2012;90(7):1392–402.
Tomita K, et al. Glial differentiation of human adipose-derived stem cells: implications for cell-based transplantation therapy. Neuroscience. 2013;236:55–65.
Uemura T, et al. Transplantation of induced pluripotent stem cell-derived neurospheres for peripheral nerve repair. Biochem Biophys Res Commun. 2012;419(1):130–5.
Unsworth BR, Lelkes PI. Growing tissues in microgravity. Nat Med. 1998;4(8):901–7.
Verdu E, et al. Olfactory bulb ensheathing cells enhance peripheral nerve regeneration. NeuroReport. 1999;10(5):1097–101.
Volpato FZ, et al. Using extracellular matrix for regenerative medicine in the spinal cord. Biomaterials. 2013;34(21):4945–55.
Vunjak-Novakovic G, et al. Bioreactor cultivation conditions modulate the composition and mechanical properties of tissue-engineered cartilage. J Orthop Res. 1999;17(1):130–8.
Walsh S, et al. Supplementation of acellular nerve grafts with skin derived precursor cells promotes peripheral nerve regeneration. Neuroscience. 2009;164(3):1097–107.
Walsh SK, et al. Skin-derived precursor cells enhance peripheral nerve regeneration following chronic denervation. Exp Neurol. 2010;223(1):221–8.
Wang D, et al. Bridging small-gap peripheral nerve defects using acellular nerve allograft implanted with autologous bone marrow stromal cells in primates. Brain Res. 2008;1188:44–53.
Wang J, et al. Bone marrow mesenchymal stem cells promote cell proliferation and neurotrophic function of Schwann cells in vitro and in vivo. Brain Res. 2009;1262:7–15.
Wang A, et al. Induced pluripotent stem cells for neural tissue engineering. Biomaterials. 2011;32(22):5023–32.
Wang Y, et al. Gene network revealed involvements of Birc2, Birc3 and Tnfrsf1a in anti-apoptosis of injured peripheral nerves. PLoS ONE. 2012;7(9):e43436.
Wanner IB, et al. Role of N-cadherin in Schwann cell precursors of growing nerves. Glia. 2006;54(5):439–59.
Watt FM, Hogan BL. Out of Eden: stem cells and their niches. Science. 2000;287(5457):1427–30.
Watt FM, Huck WT. Role of the extracellular matrix in regulating stem cell fate. Nat Rev Mol Cell Biol. 2013;14(8):467–73.
Wei Y, et al. Chitosan/silk fibroin-based tissue-engineered graft seeded with adipose-derived stem cells enhances nerve regeneration in a rat model. J Mater Sci Mater Med. 2011;22(8):1947–64.
Weimann JM, et al. Contribution of transplanted bone marrow cells to Purkinje neurons in human adult brains. Proc Natl Acad Sci USA. 2003a;100(4):2088–93.
Weimann JM, et al. Stable reprogrammed heterokaryons form spontaneously in Purkinje neurons after bone marrow transplant. Nat Cell Biol. 2003b;5(11):959–66.
Whitlock EL, et al. Processed allografts and type I collagen conduits for repair of peripheral nerve gaps. Muscle Nerve. 2009;39(6):787–99.
Williams DF. On the mechanisms of biocompatibility. Biomaterials. 2008;29(20):2941–53.
Williams DF. Essential biomaterials science. Cambridge: Cambridge University Press; 2014.
Wilson AD, et al. Delayed acetyl-L-carnitine administration and its effect on sensory neuronal rescue after peripheral nerve injury. J Plast Reconstr Aesthet Surg. 2007;60(2):114–8.
Wislet-Gendebien S, et al. Plasticity of cultured mesenchymal stem cells: switch from nestin-positive to excitable neuron-like phenotype. Stem Cells. 2005;23(3):392–402.
Wolchok JC, Tresco PA. The isolation of cell derived extracellular matrix constructs using sacrificial open-cell foams. Biomaterials. 2010;31(36):9595–603.
Woodbury D, et al. Adult rat and human bone marrow stromal cells differentiate into neurons. J Neurosci Res. 2000;61(4):364–70.
Wrabetz L, et al. Different intracellular pathomechanisms produce diverse myelin protein zero neuropathies in transgenic mice. J Neurosci. 2006;26(8):2358–68.
Xiong Y, et al. Synaptic transmission of neural stem cells seeded in 3-dimensional PLGA scaffolds. Biomaterials. 2009;30(22):3711–22.
Xu Y, et al. Myelin-forming ability of Schwann cell-like cells induced from rat adipose-derived stem cells in vitro. Brain Res. 2008;1239:49–55.
Xue C, et al. Joint use of a chitosan/PLGA scaffold and MSCs to bridge an extra large gap in dog sciatic nerve. Neurorehabil Neural Repair. 2012;26(1):96–106.
Yang Y, et al. Repair of rat sciatic nerve gap by a silk fibroin-based scaffold added with bone marrow mesenchymal stem cells. Tissue Eng Part A. 2011;17(17–18):2231–44.
Yohn DC, et al. Transplanted mouse embryonic stem-cell-derived motoneurons form functional motor units and reduce muscle atrophy. J Neurosci. 2008;28(47):12409–18.
Yu J, et al. Induced pluripotent stem cell lines derived from human somatic cells. Science. 2007;318(5858):1917–20.
Zhang P, et al. Bridging small-gap peripheral nerve defects using biodegradable chitin conduits with cultured schwann and bone marrow stromal cells in rats. J Reconstr Microsurg. 2005;21(8):565–71.
Zhang Y, et al. A nerve graft constructed with xenogeneic acellular nerve matrix and autologous adipose-derived mesenchymal stem cells. Biomaterials. 2010;31(20):5312–24.
Zhang P, et al. Biodegradable conduit small gap tubulization for peripheral nerve mutilation: a substitute for traditional epineurial neurorrhaphy. Int J Med Sci. 2013;10(2):171–5.
Zochodne DW. The challenges and beauty of peripheral nerve regrowth. J Peripher Nerv Syst. 2012;17(1):1–18.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Shanghai Jiao Tong University Press, Shanghai and Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Xue, C., Yi, S., Gu, X. (2015). Stem Cell and Peripheral Nerve Regeneration. In: Zhao, R. (eds) Stem Cells: Basics and Clinical Translation. Translational Medicine Research, vol 1. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-7273-0_10
Download citation
DOI: https://doi.org/10.1007/978-94-017-7273-0_10
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-7272-3
Online ISBN: 978-94-017-7273-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)