
Abstract
Artificial reproduction, being a specific human intervention in the process of reproduction, is a key step in aquaculture of percid fishes. This group of fish, exhibits specific traits, considered amenable to artificial reproductive protocols. For example, this is the only extensively studied group of freshwater teleosts where application of human chorionic gonadotropin (hCG) and gonadoliberine analogues (GnRHa) alone, promotes final oocyte maturation (FOM) and spawning without any other hormonal therapy, whereas in other species (cyprinids, catfishes or salmonids) anti-dopaminergic treatment is also needed. Another characteristic trait is that percid females can release their eggs spontaneously in the tank, regardless of the presence of males. This makes artificial spawning of these fish relatively difficult. In the present chapter endocrine regulation as well as reproductive protocols applied to this group of fish are reviewed extensively, however, the focus of this review is on the final gamete maturation, spermiation and ovulation processes are the steps considered from artificial reproduction perspectives. The published data revealed that scientific activity was focused mainly on the problem of synchronization of ovulation and the effectiveness of different hormonal therapies. This evolved into the development of several specific protocols and methods (e.g. percid-specific pre-ovulatory maturational stages of oocytes), which allowed improvement of that in these species. It was also established, that hCG or GnRHa applied alone are the most effective spawning agents in wild or pond-reared percids. However, there is still a considerable lack of data considering the effectiveness of these protocols in controlled reproduction of domesticated broodstocks. Apart from that, there are many other aspects to be investigated. Such as hormonal regulation of final gamete maturation and spawning, verification of some reproductive protocols as possible gamete quality determinants and gamete management protocols (prior to and following fertilization), which were relatively scarcely studied.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
1 Introduction
Spawning, either natural or artificial, is a key step in fish lifecycle. In the wild the act of spawning closes the development of the organism into adult specimen and in this way ensures the survival of the species. In captivity spawning is a necessary element securing constant production. All the commercially important percid fishes exhibit some differences considering the reproductive features, where the structure of the eggs laid (in the form of ‘ribbon’ or as a batch of single eggs) is the most distinctive one (for details see Chap. 2). However, there are also many similarities allowing consideration of these species as a one specific group representing comparable pattern of gonadal development as well as gametes maturation and spawning (see also Chaps. 2 and 3).
Artificial reproduction may be defined as a human intervention in the process of reproduction (Woynarovich and Horváth 1980). It takes into account a number of techniques aimed at producing high quality offspring suitable for further culture process or restocking. Generally, as considering artificial reproduction human intervention was concentrated on the final phases of the maturation of spawners (final oocyte maturation, ovulation and spermiation) and the methods of control over these processes as well as gametes management and other procedures (in vitro fertilization, egg treatment, incubation) up to hatching. This concerns, among others, manipulation of environmental conditions, hormonal therapies, gametes handling and other protocols which were proved to be crucial for successful reproduction.
From an aquaculture perspective, successful reproduction is one of the crucial steps in the culture process. Its effectiveness directly affects the production capacity and in this way may influence the production profitability. In all cultivated percids the reproduction of captive broodstock is feasible. Nevertheless, the artificial reproduction of wild or pond reared fish (which are exposed to natural food source and natural photo-thermal conditions) is still a major concern. Therefore, this chapter extensively described the procedures of artificial reproduction of wild (or pond reared) fish as well the one of breeders held in intensive conditions.
2 Endocrine Regulation of Spermiation
Endocrine regulation of sexual maturation and spermatogenesis in male percids is generally similar to that of other teleosts. As in most animal species spermatogenesis of fish is also controlled by the hypothalamic-pituitary-gonad axis. The gonadotropin releasing hormone (GnRH) produced in the hypothalamus stimulates the release of gonadotropins (GtH) of the pituitary which in turn stimulate the steroid (androgen) production in the testis (Vizziano et al. 2008). While the function of androgens has been studied to some extent in percids, very little information is available on the male-specific actions of either GnRH or GtH-s in these species. GnRH is a decapeptide, a neurohormone that is mainly localized in the anteroventral preoptic region of the telencephalon (Peter 1983) which does not seem to exert male- or female-specific action. Its production is controlled by an antagonist, the dopamine, which is a unique feature in teleosts. Inhibition of GnRH action by dopamine is not observed in all teleosts, and particularly little information is available regarding this process in males. Fish gonadotropins are present in two forms characterized as either GtH I and GtH II being homologous with mammalian FSH and LH (Vizziano et al. 2008) or as FSH and LH proper (Schulz et al. 2010). Unlike in mammals where LH regulates sex steroid release by Leydig cells and FSH is responsible for the regulation of Sertoli cell activities in males, in fish the roles of the two forms are not clearly separated. Receptors of FSH display a preference for FSH, however, they can be activated by LH, too, although at higher concentrations. LH receptors seem to be more LH-specific in fish where cross-activation by FSH is possible only in non-physiologically high concentrations (Bogerd et al. 2005). Nevertheless, FSH is known to stimulate steroidogenesis in fish, suggesting that FSH receptors are present in the Leydig cells. The expression of FSH and LH receptor mRNA in Leydig cells was demonstrated for the first time in fish (and vertebrates) in the African catfish (Clarias gariepinus) by García-López et al. (2009). Generally, FSH concentrations are high during the onset of spermatogenesis (proliferation of spermatogonia, meiosis and early spermiogenesis) while LH levels start to increase close to spawning (spermiogenesis and spermiation) what was described extensively in Chap. 3.
The unique study on the gonadotropins of percid males has shown an increased expression of FSHβ and LHβ (the hormone specific β-subunit of gonadotropins) mRNA in pikeperch (Sander lucioperca) during early and mid-spermatogenesis compared to pre-spermatogenesis (Hermelink et al. 2011). The increase of LHβ mRNA expression was more pronounced than that of the FSHβ mRNA which corresponds with the findings in other teleost species. The main steroid hormones found in the testis include estrogens such as 17β-estradiol (E2), androgens such as testosterone (T) and 11-ketotestosterone (11-KT) and progestins such as 17α,20β-dihydroxy-4-pregnen-3-one (abbreviated as DHP or 17,20β-P) and 17α,20β,21-trihydroxy-4-pregnen-3-one (20β-S) (Schulz et al. 2010). The major sites of sex steroid synthesis in the testis are the Leydig cells that can be found in the interstitium between individual tubules or lobules. These are large polygonal cells characterized by extensive agranular endoplasmatic reticulum and mitochondria with tubular christae. These cells appear before or during sex differentiation depending on the species. These cells are believed to be homologous with the steroid-producing Leydig cells of higher vertebrates (Billard et al. 1982).
The function of estrogens in the testis is still largely unclear but it is suggested that it plays a role in spermatogonial renewal and multiplication (Vizziano et al. 2008). They have also been reported to play a major role in gene expression in the testis (Schulz et al. 2010). In percid males E2 levels were rarely studied. As a notable exception, E2 concentration in males of yellow perch (Perca flavescens) was found to be low throughout the year with a five to tenfold increase in November–December to 50–100 pg/ml while decreasing just before the spawning season (Ciereszko et al. 1998). In contrast, in Eurasian perch (Perca fluviatilis) males E2 concentrations were found to be high during the spawning season (probably sustaining spermiation) and decreased significantly after the end of spawning (Rougeot et al. 2004). This corresponds with the findings of Hermelink et al. (2011) who reported that E2 levels remained at high and stable levels during pre-, early and mid-spermatogenesis in pikeperch.
Androgen receptors are expressed in the testicular somatic cells (primarily Sertoli and interstitial cells) but not in the germ cells (Schulz et al. 2010). They effectively support the entire process of spermatogenesis from spermatogonial multiplication to spermiation. Although both can be found in females, too, levels of 11-KT were found to be ten times higher in pikeperch males than in females (Teletchea et al. 2009a). Seasonal changes in plasma T and 11-KT levels have been relatively well described in percids and follow the same general pattern of decreasing abruptly following spawning and beginning to increase roughly 4–5 months before the subsequent spawning (described in greater details in Chap. 3).
Progestins have been shown to be present in elevated concentrations throughout the spermiation process in many fish species (Schulz et al. 2010). They play a particularly important role in sperm maturation and release, increasing pH of sperm ducts and probably exert their action through a membrane receptor of spermatozoa (Vizziano et al. 2008). In percid males the presence and concentrations of progestings are rarely measured. Interestingly, in the ruffe (Gymnocephalus cernuus) metabolites of 20β-S were found to act as pheromones that stimulated swimming activity and social interactions in conspecific males. When males were exposed to the urine of females injected with 20β-S they started swimming more actively and inspecting each other. In contrast, direct exposure to either progestins or to the urine of females injected with DHP caused no noticeable change in the behavior of males (Sorensen et al. 2004). In the pikeperch, concentrations of DHP stayed stable and low (below 100 pg/ml) during pre-, early and mid-spermatogenesis, suggesting that they only increase during active spermiation (as they do in other teleosts) which was not studied, yet (Hermelink et al. 2011).
3 Endocrine Regulation of Final Oocyte Maturation (FOM) and Ovulation
In teleosts process of oocyte maturation and ovulation is preceded by the oocyte growth (vitellogenesis) which involves incorporation of yolk proteins and lipids into oocytes (Tyler and Sumpter 1996; Devlin and Nagahama 2002; Patino and Sullivan 2002). During the vitellogenesis process oocytes are arrested in the first meiotic prophase (Nagahama 1994). The process is up-regulated by hypothalamic-pituitary-gonadal axis, where the major role plays follicle-stimulating hormone (FSH). These processes are extensively described in Chap. 3.
After the complete vitellogenesis and just before ovulation the final oocyte maturation (FOM) process begins. During the FOM, the first meiotic division is completed and the second is started, which proceeds until the metaphase II stage (Nagahama and Yagashita 2008). The process is accompanied by a number of morphological changes involving, for example, migration and decomposition of the germinal vesicle, formation of yolk and coalescence of oil droplets into one large droplet (Migaud et al. 2003; Żarski et al. 2011a, 2012a, c). In percids the most characteristic feature during FOM was the oil droplet coalescence process. It was found that the small oil droplets coalesce into one big oil droplet, what was accompanied together with the GV migration (Żarski et al. 2011a, 2012a, c). During this phase the most important role plays second gonadotropic hormone which is luteinising hormone (LH) secreted by the pituitary gland during FOM (e.g., Kagawa et al. 1998; Patino et al. 2001; Patino and Sulivan 2002). However, Fontaine et al. (2003) suggest also that in the whole process E2 and T may have been involved. Basically, FOM is triggered by the LH which stimulates theca cell layers of the ovarian follicle for production of 17α-hydroxyprogesterone which traverses the basal lamina and is then converted to the maturation inducing steroid (MIS) by the granulosa cells of the ovarian follicle (e.g., Nagahama 1994; Nagahama and Yamashita 2008). The MIS was proven to bind with the specific receptors of oocyte plasma membrane in postvitellogenic oocytes of teleosts (Nagahama and Yamashita 2008). In the case of freshwater percids, like in most fish species, the role of MIS the most probably plays 17α,20β-dihydroxy-4-pregnen-3-one (DHP) (Goetz and Theofan 1979; Goetz et al. 1989; Barry et al. 1995; Fontaine et al. 2003; Nagahama and Yamashita 2008; see also Chap. 3). The MIS promote the formation and activation of the maturation promoting factor (MPF) which is the complex of cdc2 kinase and cyclin-B (for details see: Yamashita et al. 1992; Nagahama and Yamashita 2008). The MPF is the final inducer of the oocyte maturation process (Nagahama and Yamashita 2008). The data on the MPF in freshwater percids are still missing, however already published data suggest that the pathways and mechanisms of this process is very similar to those described for other teleosts or even amphibians (extensively revised by Nagahama and Yamashita 2008).
In some of the fish species, such as cyprinids, maturation and ovulation are also controlled by the dopamine (DA), which acts as a LH inhibitory factor (Peter and Yu 1997; Mylonas et al. 2010). It is well reflected in the cases when GnRH analogues are administered for induction of maturation and ovulation in vivo, since elevated level of gonadotropins (as a result of GnRH administration) in the blood stream affect production of DA and in this way decreases effectiveness of such hormonal therapy (e.g., Peter and Yu 1997; Mylonas et al. 2010). In consequence, administration of GnRH alone does not affect maturation and ovulation. In such cases dopamine antagonist (e.g., metoclopramide, pimozide) administration is needed to affect ovulation (Krejszeff et al. 2009, 2010; Targońska et al. 2010; Kujawa et al. 2011). However, in the case of percids such additional treatment is not needed and administration of GnRH alone affects ovulation successfully (e.g., Kouril et al. 1997; Kucharczyk et al. 1998; Ronyai and Lengyel 2010; Żarski et al. 2013a). It may be then concluded that inhibitory effect of DA does not occur in the freshwater percids or it is very weak. Nevertheless, as considering endocrine regulation of FOM and ovulation of those species it seems that this aspect may be omitted.
Ovulation process includes expulsion of the matured oocyte from the ovarian follicles (Goetz 1983; Goetz and Garczynski 1997). Generally, this process is up-regulated by the LH secretion what affects consequently production of MIS (as described above and in Chap. 3). However, it is still not clear which factor directly affect process of ovulation. It was found that DHP plays an essential role in this process in yellow perch (Goetz et al. 1989; Goetz and Garczynski 1997). For the act of follicular rupture during ovulation the most probably proteinases are responsible (Nagahama and Yamashita 2008). However, the biochemical pathways between DHP (17,20β-P) and activation of proteinases remains unknown, although the arachidonic acid and prostaglandins were reported to be involved in this process (Bradley and Goetz 1994; Goetz and Garczynski 1997; Patino et al. 2003). It was reported, that indomethacin, a prostaglandin endoperoxide synthase inhibitor, has been responsible for inhibition of ovulation in yellow perch. It may be then suggested that cyclooxygenase metabolites are responsible in some way in regulation of ovulation (Goetz and Theofan 1979; Goetz and Garczynski 1997).
4 Artificial Spawning
4.1 Spontaneous Captive Spawning
One of the easiest methods of fish artificial spawning is so called ‘semi controlled’ (‘semi artificial’) spawning in tanks, small ponds or cages placed in the pond or lake (Steffens et al. 1996; Kucharczyk et al. 2007; Zakęś and Demska-Zakęś 2009). This method was widely used by inexperienced farmers, who wanted to produce larvae of those species. In general, the spontaneous spawning, regardless if performed in ponds, cages or tanks, is a simple method where spawners of both sexes are kept in a closed environment allowing them spawning without strict human control and stimulated only by the environmental conditions (e.g. Kucharczyk et al. 2007; Policar et al. 2008). The big advantage of this method is that fish are not disturbed by human, what limits the stress and they spawn whenever they are really ready to reproduce. The big disadvantages of such reproduction method are, very often, low fertilization rate, no possibility of controlled fertilization (for example specific crossbreeding, genome manipulation etc.) and limited possibilities to predict the moment of spawning. It may be improved by the application of hormonal treatment (e.g. Kucharczyk et al. 2000, 2007), but it still remains rather difficult to control the reproduction, although fish will spawn more synchronously.
In the case of yellow and Eurasian perch the spontaneous spawning was reported to be used many times (Hinshaw 2006; Ronyai and Lengyel 2010; Żarski et al. 2011a). This is caused by the fact that these species lay eggs in the form of ribbon (described in details in Chap. 2) which may be then easily removed and incubated in a regular hatchery devices. In this case, the fertilization rate should not be a problem if fish are kept in a small tank where spermiating male is able to produce a ‘sperm cloud’ which allows the spermatozoa surround and fertilize the ribbon effectively (Kucharczyk et al. 2000). However, there are no possibilities to perform selective breeding and/or other specific techniques such as fertilization of eggs with cryopreserved sperm or genome manipulation.
Application of this method of reproduction in the case of walleye and pikeperch is more complicated due to the fact that these species lay eggs as a batch of hundred thousands of sticky eggs strongly attached to any kind of substrate. Therefore, it is necessary to provide the nests in the spawning area. The nests are usually made of different natural (e.g. juniper branches, conifer branches, roots, sedges, alder, rice grass, wood wool, Fig. 4.1) or synthetic materials (Wojda et al. 1994; Skrzypczak et al. 1998; Kucharczyk et al. 2007). After the spawning the eggs together with the substrate were moved to the hatchery and incubated (Fig. 4.2). The incubation of the eggs attached to the substrate is quite problematic. The unfertilized or not developing eggs cannot be removed what usually leading to the fungal infection significantly reducing the incubation effectiveness. The substrate itself, when natural materials are used, is also a very good medium for fungal infection. Therefore, some improvement may be achieved by using the artificial substrate, although the problem of providing good water exchange around all the eggs still exists. Of course, this method of propagation may also be applied with or without hormonal treatment of spawners. However, this method is still much less effective than in the case of yellow and Eurasian perch, since fertilization rate, and consequently survival rate, are usually much lower than the methods involving controlled fertilization.

A male pikeperch above the nest made of juniper branches with the eggs laid on it (Photo: D. Żarski)

A juniper branch with the eggs of pikeperch laid on it. The abundance of white eggs indicates a low fertility and the whitish cloudy areas the beginning of fungal infection (Photo: D. Żarski)
Actually, the captive spontaneous spawning is the method which may be recommended to inexperienced farmers who has limited knowledge and skills in controlled reproduction. However, it has to be stressed that this method may generate low and unpredictable spawning effectiveness. While this method may be successfully applied in commercial farming of yellow and Eurasian perch, commercial aquaculture of pikeperch and walleye should not rely solely on this kind of propagation method.
4.2 Controlled Reproduction
Controlled reproduction involves the procedures of stimulation of spermiation and ovulation in already matured fish, gametes collection and in vitro fertilization. For culture purposes it is becoming a more and more required way for obtaining eggs and larvae. The biggest advantage of this method of propagation is the possibility of selective breeding of specimens with required pheno- and genotypes and to having control over the entire procedure of reproduction (Thorgaard 1995; Zohar and Mylonas 2001; Mylonas et al. 2010). It includes, among others, timing of fertilization, duration of incubation and timing of hatching. The latter, in the case of percids, may be crucial for larviculture effectiveness where size heterogeneity of larvae, caused by the different hatching times, may increase intensity of cannibalism and, in consequence, production outcome (e.g. Baras et al. 2003; Kestemont et al. 2003; Kooten et al. 2010), as extensively described in Chaps. 9, 10 and 11
4.2.1 Sex Recognition
In all percids there is no clear sexual dimorphism. It was already reported that during the culture process some sex-dependent growth heterogeneity may be observed (for more details see Chap. 2). However, fish at the same size beyond the spawning season usually looks very similar and it is very hard to distinguish males from females. Only just before or during the spawning season the fish gender may be usually recognized as the males are spermiating after gentle pressure of their abdomen and females display distended abdominal part of body. However, it may still happen (especially prior to spawning season) that the sex be indistinguishable. In that case catheterization (Fig. 4.3) may be a useful method for gender verification (Ross 1984).

Catheterization of the Eurasian perch female. Close to the genital pore oocytes may be seen in the lumen of the catheter (Photo: S. Krejszeff)
4.2.2 Determination of Maturity Stage of Females
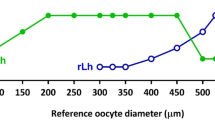
The effectiveness of hormonal stimulation as well as latency time following hormonal treatment in females are strictly dependent on the maturity stage (Brzuska 1979, 1988). Until recently, in freshwater aquaculture, the germinal vesicle (GV) position in preovulatory oocytes was taken into consideration for determining the maturation stage of females. In effect four stages were distinguished: central position of GV (stage 1), early migration of GV (stage 2), late migration of GV (stage 3), peripheral position of GV or oocytes after the GV breakdown, i.e. GVBD (stage 4). However, the application of this method in freshwater percids in many cases created many ambiguities and consequently revealed weakness of this classification in this group of fish (Żarski et al. 2011a, 2012a). The largest ambiguity concerned the last stage (stage 4), where some authors to this stage qualified oocytes with the GV at a peripheral position (Kucharczyk et al. 1996, 1998, 2001; Targońska et al. 2014; Żarski et al. 2011b) and others oocytes which already undergone GVBD (Barry et al. 1995; Malison et al. 1998; Ronyai 2007; Zakęś and Demska-Zakęś 2009). Therefore, a verification of the final oocyte maturation (FOM, as described earlier in Sect. 4.3) process was made in order to distinguish a percid-specific features of preovulatory oocytes. Actually, six preovulatory oocyte maturation stages were distinguished, first in Eurasian perch (Żarski et al. 2011a):
-
Stage I: the GV was situated in the oocyte centre, oil droplets were poorly visible (Figs. 4.4a and 4.5a);
Fig. 4.4 
Oocyte maturation stages for percids on the example of Eurasian perch (according to Żarski et al. 2011a); (a) stage I, (b) stage II, (c) stage III, (d) stage IV, (e) stage V, (f) stage VI; for details see the text; bar: 0.5 mm
Fig. 4.5 
Histological cross-sections of oocytes and ovulated eggs of Eurasian perch at different maturational stages (according to Żarski et al. 2012c): (a) stage I, (b) stage II, (c) stage III, (d) stage IV, (e) stage V, (f) early phase of stage VI, (g) late phase (after proteolysis) of stage VI, (h) stage VII (ovulated eggs). 1 germinal vesicle, 2 oil droplets, 3 zona radiata externa, 4 yolk vesicles and globules, 5 homogenous (after proteolysis) yolk before hydration, 6 yolk after hydration, 7 cortical alveoli, 8 zona radiata externa without visible microvillosities. Bars represent 0.5 mm
-
Stage II: beginning of GV migration, beginning of coagulation of clearly visible oil droplets (Figs. 4.4b and 4.5b);
-
Stage III: migrating of GV (above half of the oocyte diameter), oil droplets were clearly visible (Figs. 4.4c and 4.5c);
-
Stage IV: the GV is at the oocyte periphery, a large forming oil droplet was clearly visible; the droplet diameter was greater than the GV diameter and it reached the size of about one third of the oocyte diameter; also smaller droplets were visible (Figs. 4.4d and 4.5d);
-
Stage V: the GV is situated at the oocyte edge, clearly visible one large (size of about half the oocyte diameter) oil droplet (Figs. 4.4e and 4.5e);
-
Stage VI: oocyte samples taken for analysis were macroscopically transparent; no visible GV after they were placed in Serra’s solution (following GVBD), oocytes at the pre-ovulation stage (Figs. 4.4f and 4.5f, g);


It may be found, that for the classification, not only the position of germinal vesicle but also the oil droplets coalescence rate were considered. This phenomenon, next investigated histologically (Żarski et al. 2012c, Fig. 4.5), seems to be very specific to percid fishes as this classification was later successfully transferred also to pikeperch (Żarski et al. 2012a). Therefore, it may be suggested that this classification may be used (with some minor modifications) in all percids allowing higher synchronization and more accurate prediction of ovulation.
4.2.3 Stimulation of Spermiation
Regarding hormonal induction of spermiation in percids, little information is available that is specific to this taxon. Percids are known to release an ample amount of sperm during the reproductive season. For example, non-treated Eurasian perch was found to release approximately 13–20 mL of sperm per kg of body weight during the spawning season (Kucharczyk et al. 1996, 1998; Alavi et al. 2010). Therefore hormonal induction of males is only a supplementary measure to enhance milt production. Spermiation can be induced by a change in environmental conditions, such as temperature increase during transfer from one habitat to another as it was described for the walleye (Dąbrowski et al. 2000). Human chorionic gonadotropin (hCG), carp pituitary extract (CPE) or a combination of the two were used to stimulate spermiation in the pikeperch and Volga pikeperch (Sander volgensis) but none of them were applied male-specifically (Bokor et al. 2007, 2008). Żarski et al. (2013a) reported, that neither gonadotropins (i.e. hCG, CPE or combination of hCG and pregnant mare serum gonadotropin) nor neurohormone (gonadoliberine analog) affected spermiation rate and spermatozoa motility in pikeperch. In the yellow perch monthly injections of luteinizing hormone releasing hormone analog (LHRHa) starting in January increased production of milt in February and March (Dabrowski et al. 1994). Males of the Eurasian perch produced significantly higher volumes of semen following injection with either 2 mg per kg of body weight of carp pituitary extract or 0.5 pellet of the synthetic GnRH analog Ovopel as compared to the untreated control (Kucharczyk et al. 2001). Also combined injection of FSH (25 μg kg−1) and LH (25 μg kg−1) administered together with a dopamine antagonist (metoclopramide at a dose of 2.5 mg kg−1) significantly increased sperm production in this species (Kucharczyk et al. 1996). During out-of-season spawning trials of Eurasian perch, smaller fish (individual weight: 30–70 g) displayed a higher percentage of spermiating individuals than did larger fish (200–400 g) following injection with various doses of Ovopel, however, it was unclear whether this was an effect of hormonal treatment or that of the pre-spawning chilling period as these conditions were not investigated independently (Szczerbowski et al. 2009). Thus, administration of hormonal products to percid males can enhance spermiation parameters but it is not an inevitable procedure. However, it is very important to note that the sperm volume and quality may be dependent on the timing of its collection, with the highest parameters being recorded during the spawning season (Alavi et al. 2010). However, there is still missing data about the effectiveness of different hormonal treatment protocols and the latency time following hormonal treatment on the quantity and quality of sperm in percids, what has already been proven to have huge impact on the effectiveness of stimulation of spermiation in other fish species (Król et al. 2009; Cejko et al. 2012, 2013).
4.2.4 Stimulation of Ovulation
There are numerous differences in the methods and effectiveness of artificial reproduction of cultured and wild females of percid fishes. In the cultured stocks the most important aspect is to provide proper feeding regime and promote vitellogenesis with the use of proper photothermal manipulations prior to spawning (see Chaps. 3, 20 and 21 for details). These factors directly affect gamete maturation process and consequently eggs quality (Ciereszko et al. 1997, 1998; Sulistyo et al. 1998; Henrotte et al. 2008, 2010; Abdulfatah et al. 2011). Induction of FOM and ovulation in cultured females is possible with the application of photothermal manipulations only, even out-of the spawning season (Müller-Bellecke and Zienert 2008). So, there is no need for any hormonal therapy to obtain the eggs in contrast to wild fish, where hormonal stimulation is usually required. In the case of cultured stocks reproductive procedures are adjusted to the local conditions (within particular fish farm) and in accordance with specific production cycle.
Stimulation of FOM and ovulation under the controlled conditions is conducted mainly by the photo-thermal manipulations (Kolkovski and Dąbrowski 1998; Zohar and Mylonas 2001; Müller-Bellecke and Zienert 2008) and hormonal treatment (Brzuska 2005; Krejszeff et al. 2009, 2010; Mylonas et al. 2010). Additionally, some authors suggest that pheromonal stimulation can play a significant role in these processes (Stacey 2003; Żarski 2012). In the case of percids, among the environmental cues, induction of FOM (which includes morphological changes in oocytes after the completed vitellogenesis as well as germinal vesicle [GV] migration and its breakdown [GVBD]; for more details see Sects. 4.3 and 4.4.2.2) as well as ovulation are mainly controlled by the temperature (Dąbrowski et al. 1996). Photoperiod in those processes plays a minor role (Kayes and Calbert 1979; Dąbrowski et al. 1996; Ciereszko et al. 1997). Thus, in the practice of artificially induced reproduction more attention is paid to the thermal manipulations (Dąbrowski et al. 1994; Kucharczyk et al. 2007; Żarski et al. 2013a). Basically, in all percids similar thermal conditions are applied (Tables 4.1, 4.2, 4.3 and 4.4). The optimal temperature for artificial reproduction ranges between 10 and 15 °C (Kayes and Calbert 1979; Dąbrowski et al. 1994; Kucharczyk et al. 1996, 1998; Kouril et al. 1997; Malison et al. 1998; Zakęś and Demska-Zakęś 2009; Żarski et al. 2011b, 2013a; Křišťan et al. 2012a). More recently it was shown that thermal manipulations could be a very useful tool in synchronization of ovulation in pikeperch (Żarski et al. 2013a). However, in order to evaluate possible reaction to such manipulation, and consequently apply it successfully in artificial reproduction, more studies are required to include species- and population-specific differences.
In the practice of hormonal stimulation two hormonal therapies are generally used. First, is to stimulate releasing of endogenous gonadotropins (GtH) from pituitary gland with the application of preparations containing gonadoliberine (GnRH, for more details see Sects. 4.2 and 4.3). The second one is to inject fish with GtH containing preparations (e.g. carp pituitary extract [CPE], human chorionic gonadotropin [hCG]) directly affecting Leydig cells in the testis or follicle cells in ovaries. This results in production of maturation inducing steroid (MIS) promoting final maturation of gametes and, in consequence, spermiation or ovulation (for more details see Sects. 4.2 and 4.3).
In the case of wild females, the necessity of hormonal stimulation during the spawning season mainly depends on the maturity stage of females (Kucharczyk et al. 2007). Fish at the latest maturation stage VI (after the GVBD; according to the classification of Żarski et al. 2011a, 2012a) would most likely ovulate without the hormonal treatment. However, the moment of ovulation is then very hard to predict. Fish at less advanced maturational stages very often may not ovulate (Kucharczyk et al. 2001; Żarski et al. 2011a, 2013a) or ovulation is extended up to few weeks and its effectiveness is usually very low (Dąbrowski et al. 1994; Kucharczyk et al. 1996, 1998; Ronyai and Lengyel 2010). Reproductive effectiveness in such cases may also depend on other factors such as temperature (Dąbrowski et al. 1994) or possible pheromonal stimulation which was proven to promote GVBD in pikeperch (Barry et al. 1995) or FOM in Eurasian perch (Żarski 2012). Such differences may also stem from handling stress affecting reproductive effectiveness negatively (Schreck et al. 2001; Wang et al. 2006). It is then almost not possible to predict the timing of ovulation and therefore perform in vitro fertilization. Also, the effectiveness of artificial reproduction of wild fish may be dependent on the time of fish capture. Kestemont et al. (1999) reported lower egg quality in late spawning season (when fish are the most advanced in maturation and hormonal treatment may be not necessary) compared with those obtained earlier during the spawning period. That is why many authors have focused on the development of hormonal treatment protocols allowing higher synchronization and predictability of ovulation in females of percids.
In percids the most commonly applied hormonal preparations are human Chorionic Gonadotropin (hCG) and GnRH analogues (GnRHa) while carp pituitary extract (CPH) was used less frequently (see Tables 4.1, 4.2, 4.3 and 4.4). Although, application of GnRHa in the case of percids was reported to be effective without dopamine antagonist (DA) injection, contrarily to some other fish species such as the cyprinids (Kucharczyk et al. 2005, 2008), in many cases administration of DA was reported (see Tables 4.1, 4.2, 4.3 and 4.4). Interestingly, in some cases such preparations (GnRHa+DA) were reported to affect ovulation rate and egg quality negatively (Zakęś and Demska-Zakęś 2009), whereas GnRHa administered alone revealed usually satisfied results (Kouril et al. 1997; Schlumberger and Proteau 1996; Ronyai and Lengyel 2010). However, it was reported in Walleye (Barry et al. 1995; Malison et al. 1998) and pikeperch (Żarski et al. 2013a) that the effectiveness of GnRHa could be also dependent on the maturity stage of the females where reproductive success was higher when more advanced in maturation fish were spawned.
Induction of ovulation is very often dependent on the dose of the hormone applied. In percids the most commonly applied preparation, which is hCG, was proved to be effective within a relatively wide range of doses, from 200 to 5700 IU per kg of females body weight. The most commonly applied dose ranged between 400 and 500 IU kg−1 (Tables 4.1, 4.2, 4.3 and 4.4).
The most effective doses of GnRHa (administered with 10 mg kg−1 of pimozide which is a DA) in yellow perch ranged between 110 (administered in two doses of 10 and 100 μg kg−1 respectively; pimozide was administered with the first dose) and 300 μg kg−1 (Dąbrowski et al. 1994). Similarly, injection of Eurasian perch with 125 μg kg−1 resulted in higher spawning and fertilization rate as compared to groups treated with 25 μg kg−1 (Kouril et al. 1997). In walleye a single GnRHa dose of 100 μg kg−1 was effective to cause GVBD (Barry et al. 1995). The effective dose of mammalian GnRHa (administered together with DA – metoclopramide) in Eurasian perch was 22 μg kg−1 (Kucharczyk et al. 2001) and in pikeperch 40 μg kg−1 (Żarski et al. 2013a). Malison et al. (1998) reported that doubled injection of GnRHa (35 and 100 μg kg−1) caused ovulation only in walleye females with already migrating GV, as compared to the fish with oocyte exhibiting GV in the central position (less advanced in maturation). Such differences in effectiveness of GnRHa application could stem from difference in both fish maturation stages (Żarski et al. 2013a) and activeness of GnRHa forms (amino acid sequence), as already reported in other fish species (Podhorec and Kouril 2009; Targońska et al. 2010).
Application of CPH was very often performed in two doses, with the first one (priming dose) constituting 10–50 % of the second (resolving) one. In general, the average applied total doses ranged between 3 and 5 mg kg−1 (Tables 4.1, 4.2, 4.3 and 4.4). However, in pikeperch (Müller et al. 2004) and walleye up to 15 mg kg−1 and 26.8 mg kg−1 (in two equal doses) (Lessman 1978) were used, respectively. Even such high doses seemed to be not harmful to the fish. Nevertheless, the data on the application of this spawning agent are quite limited, as compared to the other hormonal preparations.
As mentioned above, when considering reproduction of wild females of percids, the procedures which may be undertaken (photothermal manipulation, hormonal and/or pheromonal stimulation) are strictly dependent on the maturational stages of females. And the biggest problem is that wild females during the spawning season, even caught from the same population and at the same time, are usually at different maturity stages (from I to VI as described by Żarski et al. 2011a, 2012a). It is worth mentioning that stimulation with hCG at a maturation stage I and II (according to the classification given by Żarski et al. 2011a, 2012b, c) resulted in lower ovulation and fertilization rates compared with the fish hormonally stimulated at stages III–VI. Thus, it may be recommended to reproduce wild percids females at a stage III or later.
4.2.5 Gamete Collection
Collection of gametes in percids does not create any difficulties as it is in sturgeon females where special surgical techniques are necessary for obtaining the eggs from the genital pore (Mims et al. 2004; Pourasadi et al. 2009) or in catfish males where testes have to be dissected to extract a sufficient amount of sperm (Legendre et al. 1996; Bokor et al. 2010). For the artificial spawning purposes collection of sperm and eggs in percids is proceeded through simple stripping (gentle massaging of the abdomen parts of the fish body) (Rinchard et al. 2011; Kucharczyk et al. 2007; Zakęś and Demska-Zakęś 2009; Żarski et al. 2011a, 2012a).
In males there is a need for taking care to avoid urine contamination of stripped semen when the sperm is collected into the syringes (or any other container) because spermatozoa might have been activated by the urine (Bokor et al. 2007). This can lead to low motility rate of spermatozoa, even after short period of storage (Satterfield and Flickinger 1995a) and, consequently, affect fertilization rate negatively. A possible method is to use a catheter for sperm collection (Bokor et al. 2007; Grozea et al. 2008; Korbuly et al. 2010). It was previously reported that such method allows avoiding urine contamination in salmonids (Glogowski et al. 2000). Another way is to strip the sperm shortly before using it for fertilization (Kucharczyk et al. 2007). More details are provided in the Chap. 5.
Collection of eggs is very simple when fish is taken for stripping shortly after ovulation (Fig. 4.6). However, it exist huge differences among percids regarding the type of ovulated eggs. Eggs of pikeperch and walleye are released as a batch of separated eggs, whereas Eurasian and yellow perch eggs are included within a large, cylindrical gelatinous strand called ribbon (Probst et al. 2009; Formicki et al. 2009, see also Chap. 2). This creates some differences in the in vitro fertilization procedures. Eggs obtained from few pikeperch or walleye females are usually mixed in one bowl and are fertilized according to the “dry” method which involves mixing of eggs and sperm before activation with activating solution. In the case of both perch species it is recommended to fertilize the eggs from each female separately in order to avoid covering one egg ribbon by another, as already reported in Eurasian perch by Żarski et al. (2012d).

Eggs stripping from Eurasian perch (on the left) and pikeperch (on the right). In the case of Eurasian perch eggs in the form of ‘ribbon’ may be noticed (Photo: S. Krejszeff)
One of the biggest obstacles in percids reproduction is to get the ‘dry’ eggs. This stems from the fact that females are able to release eggs in the tank spontaneously (Żarski et al. 2011a, 2012a). In order to get the dry eggs a frequent control of ovulation, sometimes even every 1 h (Křišťan et al. 2013), is necessary. This, in turn, causes huge stress to the fish what can presumably affect the egg quality and health status of the female (e.g. Lessman 1978). Therefore, in some cases in Eurasian perch the genital pore of the fish was sewn up to prevent the spontaneous release of eggs in the tank (Kucharczyk et al. 1996, 1998, 2001). This method, with the high attention paid to the sterility of the procedure performed, was also successfully applied to pikeperch (Żarski, unpublished data, Fig. 4.7). Recently, it was found that the eggs of pikeperch may be ‘stored’ in the ovaries of the female for at least 12 h following ovulation at a temperature of 15 °C without negative effect on eggs viability (Mohagheghi Samarin, Blecha, Policar, unpublished data). Although, the ‘operation’ of the sewing up the genital pore of the fish may be quite laborious, this method is very promising alternative when the ‘dry’ eggs are absolutely essential from the breeding point of view.

A genital pore of Eurasian perch female being sewn up with the sterile surgical thread (on the left) and female pikeperch with sewn up genital pore (on the right) (Photo: S. Krejszeff)
4.2.6 In Vitro Fertilization
The in vitro fertilization is very important part of finfish controlled reproduction. This process involves exposure of the gametes (eggs and sperm) in a medium (so called ‘activating solution’ [AS]) which is able to activate sperm and eggs at the same time (Żarski et al. 2015b). After the contact with water (or any other activating solution) eggs acquiring the ability to be fertilized (Coward et al. 2002; Minin and Ozerova 2008). This ability retains for a certain period, which for many fish species is unknown. Żarski et al. (2012d) found, that eggs of Eurasian perch eggs remain active up to 2.5 min post-activation with hatchery water. However, the sperm after activation was motile only up to 37 s. And the spermatozoa motility rate was highly decreasing in time. Alavi et al. (2007) reported that only 7 % of spermatozoa are motile after 45 s post activation, whereas almost 92 % motility is recorded after 15 s post activation. Taking into account that eggs of cyprinids (Żarski et al. 2015b, 2014) and pikeperch (Żarski et al. 2013b) remain active for about 1 min following activation, eggs of Eurasian perch retain the fertilizability for a relatively long period of time. Interestingly, it was reported, that application of the ‘dry method’ for fertilization in this species (eggs and sperm mixed first and then exposed to AS; e.g. Dąbrowski et al. 1994) affected lower fertilization rate in comparison to the method where eggs were activated first and after 15 s sperm was added (Żarski et al. 2012d). This phenomenon was the most probably related with the fact that eggs within the ribbon upon activation are covered one by another. This enables to penetrate all the eggs by spermatozoa, which very suddenly lose their motility (see Chap. 5) and ability to successfully fertilize the eggs. Therefore for procedure of in vitro fertilization in Eurasian perch it may be recommended to activate the eggs first and next, after 15–30 s, add the sufficient amount of sperm (Fig. 4.8). This should allow spreading the whole ribbon within the activating solution and exposing all the eggs to the active spermatozoa, when added. Moore (2003) also reported that repeated addition of sperm to the eggs (at 0, 30 and 60 s post-activation of eggs) improved fertilization effectiveness in walleye, although mechanism is unclear since there is no data on the duration of eggs and sperm activation. Considering the fact that eggs of pikeperch lose their fertilizability very suddenly (within first minute following activation; Żarski et al. 2013b), it may be suggested that the observations made by Moore (2003) in walleye stemmed from other reasons. Nevertheless, in the view of the recently published data and considering the high variability in the fertilization rate usually observed in percid fishes there is still need for development of novel techniques of eggs fertilization (Satterfield and Flickinger 1995b; Rinchard et al. 2005; Zakęś and Demska-Zakęś 2009; Żarski et al. 2011a, 2012d).

A procedure of in vitro fertilization of eggs of Eurasian perch (according to Żarski et al. 2012d). Eggs are first activated with ‘hatchery water’ in order to spread the egg-ribbon in the water (in order to avoid of covering the eggs one by another) (on the left) and after 15–30 s sperm is added (on the right) (Photo: S. Krejszeff)
For the purposes of in vitro fertilization as an activating solution (AS) the most commonly ‘hatchery water’ was used. However, composition of such AS is not standardized and parameters (e.g. osmolality, ionic composition) are different for different hatcheries. That is why more often other AS-s are used instead of hatchery water, which are very often precisely formulated and standardized. Different activating solutions have been tested only in Eurasian perch so far. It was proven that application of the Woynarovich solution (3 g of urea and 4 g of NaCl in 1 L of distilled water; Woynarovich and Woynarovich 1980) may improve the fertilization as compared to hatchery water. It stems from the fact that in Woynarovich solution eggs retain the ability to be fertilized for over 3 min while motility of spermatozoa was observed up to 89 s post activation (Żarski et al. 2012d). Prolonged motility of percids spermatozoa after the application of specifically formulated AS-s was reported earlier (for details see Chap. 5). However, the possibility of application of those AS-s for in vitro fertilization remains unknown. Especially, when it was already reported that AS which successfully activated sperm motility was not suitable for fertilization (Saad and Billard 1987; Żarski et al. 2014). Considering the fact that application of AS-s other than hatchery water (such as Woynarovich solution in Eurasian perch) may bring huge benefits for the fish farmers increasing fertilization effectiveness, more work is needed aiming at the development of the activating solutions specifically dedicated to the particular species.
One of the most important aspect of in vitro fertilization is to use proper amount of sperm per particular amount of eggs, what is called in the literature as a “sperm:egg ratio” (e.g. Linhart et al. 2006). Rinchard et al. (2005) reported, that maximization of fertilizing ability in walleye may be secured with the minimal sperm:egg ratio of 25,000: 1. On the other hand, Casselman et al. (2006) reported that only 5000 of spermatozoa per egg affected very high (over 90 %) survival rate of embryos in this species. Such contrary results may stem from the fact that different methods for estimation of sperm concentration were used (spermatocrit method by Casselman et al. (2006) and counting chambers method by Rinchard et al. (2005)). Most recently, Křišťan et al. (2012b) found that 100,000 spermatozoa per egg may be considered as a secure sperm:egg ratio in pikeperch. Mean pikeperch sperm concentration may be considered at a level of 20 × 109 spermatozoa per mL (see e.g., Zakęś and Demska-Zakęś 2009; Bokor et al. 2008). Whereas, mean number of eggs may be considered at a level of 1000 eggs g−1, because recorded number of “dry eggs” (before activation) per gram ranged approximately between 800 and 1300 (e.g., Bokor et al. 2008, Żarski, unpublished data). Considering the above mentioned it may be recommended to use 0.5 mL of very good quality sperm for each 100 g of eggs in pikeperch and in walleye. Until now, no published data about the effect of sperm:egg ratio on fertilization effectiveness in Eurasian and yellow perch are available.
4.2.7 Eggs Treatment, Incubation and Hatching
After in vitro fertilization eggs need to be incubated under optimal conditions. In Eurasian and yellow perch incubation of eggs is usually performed without any problem as far as oxygenated water flow is provided washing all the eggs within the whole ribbon. For that purpose small floating cages or flow-through chambers have been successfully used (Kucharczyk et al. 1996; Żarski et al. 2011b, Fig. 4.9). However, pikeperch and walleye eggs are getting to be sticky after their contact with water. Thus, prior to incubation in regular hatchery devices (such as Weiss or Zuger jars), an unsticking procedure must be applied. For that purpose talc with sodium chloride (Schlumpberger and Schmidt 1980; Steffens et al. 1996; Kucharczyk et al. 2007), enzymes (Krise et al. 1986, 1988; Zakęś and Demska-Zakęś 2009) and tannic acid (sometimes preceded by immersion in Woynarovich solution; Bokor et al. 2008) have been used (Demska-Zakęś et al. 2005; Rinchard et al. 2005, 2011; Kucharczyk et al. 2007). In percid fishes the latter compound is the most commonly used method, due to, among others, the low cost of its application. However, opinions about its application for commercial purposes were very often inconsistent (Demska-Zakęś et al. 2005; Kucharczyk et al. 2007; Zakęś and Demska-Zakęś 2009). A procedure involving 2–5 min baths in a tannic acid solution a few min following fertilization, with concentrations ranging between 0.4 and 1.5 g L−1, was usually applied (Czesny et al. 2005; Demska-Zakęś et al. 2005; Rinchard et al. 2005, 2011; Kucharczyk et al. 2007). Demska-Zakęś et al. (2005) suggested that this procedure affect low survival rate of embryos or eggs did not lose its adhesive properties. However, the most recent studies indicated that the problem of low effectiveness of this method was related with the time of application of tannic acid following eggs activation (Żarski et al. 2015a). It was proven that in the eggs of pikeperch following activation an extreme chorion deformation, caused by the cortical reaction, occurred (see also Chap. 6). This deformation lasted up to 10 min post activation (Żarski et al. 2012b), whereas tannic acid was usually applied shortly after activation (up to 5 min post activation) (Czesny and Dąbrowski 1998; Czesny et al. 2005; Demska-Zakęś et al. 2005; Rinchard et al. 2005, 2011; Kucharczyk et al. 2007). According to the findings of Żarski et al. 2015a the best moment of application of tannic acid (at a dose of 0.75 g L−1) was 30 min post fertilization coinciding with the end of the egg swelling process (Żarski et al. 2012b, 2015a). Such procedure allowed applying tannic acid for removing the pikeperch egg stickiness in commercial hatcheries successfully (Żarski, unpublished data, Fig. 4.10).

An incubation of the Eurasian perch egg-ribbons on the wooden frame covered with a net (mesh size 0.5 mm) from the bottom (on the left) and in the floating plastic ‘cages’ where bottom was replaced with the net (on the right) (Photo: S. Krejszeff)

A commercial-size incubation of eggs treated with tannic acid (0.75 g L−1, 30 min following eggs activation) according to the method described by Żarski et al. 2015a (Photo: D. Żarski)
Generally, all percids have similar thermal requirements as considering egg incubation. Median thermal tolerance limits (on the base of normal hatched larvae following incubation at a constant temperature from fertilization to hatching) for walleye, Eurasian and yellow perch is actually almost the same and ranging from 6.0–6.8 °C to 19.2–20.9 °C. Only pikeperch eggs require slightly higher thermal regimes ranging from 9.0–10.0 °C to 21.5–24.0 °C (after Hokanson 1977). Koenst and Smith (1976) reported that optimal temperature for incubation of walleye eggs ranged between 9 and 15 °C. For pikeperch eggs incubation temperatures between 12 and 16 °C were recommended (Kokurewicz 1969; Kucharczyk et al. 2007; Zakęś and Demska-Zakęś 2009). Saat and Veersalu (1996) reported that optimal temperatures for Eurasian perch ranged between 8 and 18 °C, while for yellow perch Hinshaw (2006) suggested temperatures from 5 to 15 °C. However, on the base of the literature data Teletchea et al. (2009b) indicated that for walleye, Eurasian and yellow perch thermal regime for incubation should be fixed at 12.5 °C, and 14 °C for pikeperch. At such temperatures incubation of Eurasian and yellow perch lasted 165 degree-days, while eggs of walleye and pikeperch had to be incubated for 140 and 115 degree-days, respectively (Teletchea et al. 2009b).
Hatching of percids larvae is usually an asynchronous event. In some cases it may last even several days among the same egg batch or egg-ribbon. It is important to emphasize that even very low quality larvae (highly malformed) were observed to hatch spontaneously (Żarski et al. 2011b, see also Chap. 6 for more details). It was already reported that early hatched Eurasian perch larvae were characterized by a higher survival rate than those hatched 2 days later. Also, it was proven that joint rearing of larvae hatched at different times may affect survival and growth rate of the larvae significantly (Kestemont et al. 2003, see also Chaps. 9, 10 and 11). Although it is not clear whether the larvae hatching earlier or later are of different biological properties, a special attention must be given to the extension of hatching time, and possible separation of early and late hatched larvae can be recommended before the initial larval rearing stage.
5 Conclusions
Based on published data, it may be concluded that percids represent a quite unique group of fish exhibiting specific traits from the perspective of artificial reproduction. For example, one of the most characteristic features is that in this group of fish, hormonal stimulation does not have to be supported by the anti-dopaminergic treatment, which is necessary in other freshwater fishes (cyprinids, catfishes and salmonids). Despite the efforts undertaken in the development of protocols allowing full control over reproduction (mostly synchronization of ovulation and hormonal therapy) in percids, there are still many aspects to be investigated. There is a considerable lack of data regarding hormonal regulation of the final gamete maturation process as well as stress-related effects on these processes. This knowledge may help to understand the variability of gamete quality, still widely observed in these species. Also, there are many ambiguities related to gamete management, both prior to and following fertilization (e.g. short-term storage, removing the adhesiveness of eggs), which may facilitate and thus improve this important step of artificial reproduction. Therefore, future research on the artificial reproduction of percids should employ modern scientific tools (e.g. proteomics, transcriptomics, etc.) possibly allowing to answer many unanswered questions.
References
Abdulfatah A, Fontaine P, Kestemont P, Gardeur JN, Marie M (2011) Effects of photothermal kinetics and amplitude of photoperiod decrease on the induction of the reproduction cycle in female Eurasian perch Perca fluviatilis. Aquaculture 322:169–176
Alavi SMH, Rodina M, Policar T, Kozak P, Pšenicka M, Linhart O (2007) Semen of Perca fluviatilis L.: sperm volume and density, seminal plasma indices and effects of dilution ratio, ions and osmolality on sperm motility. Theriogenology 68:276–283
Alavi SMH, Rodina M, Hatef A, Stejskal V, Policar T, Hamačkova J, Linhart O (2010) Sperm motility and monthly variations of semen characteristics in Perca fluviatilis (Teleostei: Percidae). Czech J Anim Sci 55:174–182
Baras E, Kestemont P, Mélard C (2003) Effect of stocking density on the dynamics of cannibalism in sibling larvae of Perca fluviatilis under controlled conditions. Aquaculture 219:241–255
Barry TP, Malison JA, Lapp AF, Procarione LS (1995) Effects of selected hormones and male cohorts onfinal oocyte maturation, ovulation, and steroid production in walleye (Stizostedion vitreum). Aquaculture 138:331–347
Billard R, Fostier A, Weil C, Breton B (1982) Endocrine control of spermatogenesis in teleost fish. Can J Fish Aquat Sci 39:65–79
Bogerd J, Granneman JCM, Schulz RW, Vischer HF (2005) Fish FSH receptors bind LH: how to make the human FSH receptor to be more fishy? Gen Comp Endocrinol 142:34–43
Bokor Z, Müller T, Bercsėnyi M, Horváth L, Urbányi B, Horváth A (2007) Cryopreservation of sperm of two European percid species, the pikeperch (Sander lucioperca) and the Volga pikeperch (S. volgensis). Acta Biol Hung 58(2):199–207
Bokor Z, Horvath A, Horvath L, Urbanyi B (2008) Cryopreservation of pike perch sperm in hatchery conditions. Isr J Aquacult-Bamidgeh 60(3):166–169
Bokor Z, Urbanyi B, Horvath L, Horvath A (2010) Commercial-scale cryopreservation of wels catfish (Silurus glanis) semen. Aquac Res 41:1549–1551
Bradley JA, Goetz FW (1994) The inhibitory effect of indomethacin, nordihydroguaiaretic acid, and pyrrolidinedithiocarbamate on ovulation and prostaglandin synthesis in yellow perch (Perca flavens) follicle incubates. Prostaglandins 48:11–20
Brzuska E (1979) The in vivo method of estimating the stages of oocytes maturation in carp Cyprinus carpio L. Acta Hydrobiol 21:423–433
Brzuska E (1988) Further investigation of in vivo estimation of the maturation stages of carp (Cyprinus carpio L.) females. Acta Hydrobiol 30:421–428
Brzuska E (2005) Artificial spawning of carp (cyprinus carpio L.): differences between females of polish strain 6 and Hungarian strain W treated with carp pituitary homogenate, ovopel or dagin. Aquac Res 36:1015–1025
Casselman SJ, Schulte-Hostedde AI, Montgomerie R (2006) Sperm quality influences male fertilization success in walleye (Sander vitreus). Can J Fish Aquat Sci 63:2119–2125
Cejko BI, Kowalski RK, Żarski D, Dryl K, Targońska K, Chwaluczyk R, Kucharczyk D, Glogowski J (2012) The influence of the length of time after hormonal treatment with [(d-ala 6, pro 9 NEt)-mGnRH + metoclopramide] i.e. ovopel on barbel Barbus barbus (L.) milt quality and quantity indicators. J Appl Ichthyol 28:249–253
Cejko BI, Żarski D, Krejszeff S, Kucharczyk D, Kowalski RK (2013) Effect of hormonal stimulation of the Crucian Carp Carassius carassius (L.) on milt volume, number of sperm and its motility. Isr J Aquacult Bamidgeh (65: 912.)
Ciereszko RE, Dąbrowski K, Ciereszko A (1997) Effects of temperature and photoperiod on reproduction of female yellow perch Perca flavescens: plasma concentrations of steroid hormones, spontaneous and induced ovulation, and quality of eggs. J World Aquacult Soc 28:344–356
Ciereszko RE, Dabrowski K, Ciereszko A, Ottobre JS (1998) Plasma concentrations of steroid hormones in male yellow perch, Perca flavescens: the effect of age and photothermal manipulation. Environ Biol Fish 51:97–105
Coward K, Bromage NR, Hibbitt O, Parrington J (2002) Gamete physiology, fertilization and egg activation in teleost fish. Rev Fish Biol Fish 12:33–58
Craig JF (2000) Percid fishes systematics. Ecology and exploitation. Blackwell Science, Dunscore
Czesny S, Dabrowski K (1998) The effect of egg fatty acid concentrations on embryo viability in wild and domesticated walleye (Stizostedion vitreum). Aquat Living Resour 11:371–378
Czesny S, Rinchard J, Dabrowski K (2005) Intrapopulation variation in egg lipid and fatty acid composition and embryo viability in a naturally spawning walleye population from an inland reservoir. N Am J Fish Manag 25:122–129
Dąbrowski K, Ciereszko A, Ramseyer L, Culver D, Kestemont P (1994) Effects of hormonal treatment on induced spermiation and ovulation in the yellow perch (Perca flavescens). Aquaculture 120(1–2):171–180
Dąbrowski K, Ciereszko RE, Ciereszko A, Toth GP, Christ SA, El-Saidy D, Ottobre JS (1996) Reproductive physiology of yellow perch (Perca flavescens): environmental and endocrinological cues. J Appl Ichthyol 12:139–148
Dąbrowski K, Czesny S, Kolkovski S, Lynch WE Jr (2000) Intensive culture of walleye larvae produced out of season and during regular season spaning. N Am J Aquac 62:219–224
Demska-Zakes K, Zakes Z, Roszuk J (2005) The use of tannic acid to remove adhesiveness from pikeperch, Sander lucioperca, eggs. Aquac Res 36:1458–1464
Devlin RH, Nagahama Y (2002) Sex determination and sex differentiation in fish: an overview of genetic, physiological, and environmental influences. Aquaculture 208:191–364
Fontaine P, Migaud H, Mandiki R, Gardeur JN, Kestemont P, Fostier A (2003) Involvement of sex steroids in final stages of oogenesis in Eurasian perch, Perca fluviatilis. Fish Physiol Biochem 28:331–332
Formicki K, Smaruj I, Szulc J, Winnicki A (2009) Microtubular network of the gelatinous egg envelope within the egg ribbon of European perch, Perca fluviatilis L. Acta Ichthyol Piscat 39:147–151
Garcìa-López Á, Bogerd J, Granneman JCM, van Dijk W, Trant JM, Taranger GL, Schulz RW (2009) Leydig cells express follicle-stimulating hormone receptors in African catfish. Endocrinology 150:357–365
Glogowski J, Kwasnik M, Piros B, Dabrowski K, Goryczko K, Dobosz S, Kuzminski H, Ciereszko A (2000) Characterization of rainbow trout milt collected with a catheter: semen parameters and cryopreservation success. Aquac Res 31:289–296
Goetz FW (1983) Hormonal control of oocyte final maturation and ovulation in fishes. In: Hoar WS, Randall DJ, Donaldson EM (eds) Fish physiology, vol IXB. Academic, New York, pp 117–170
Goetz FW, Garczynski M (1997) The ovarian regulation of ovulation in teleost fish. Fish Physiol Biochem 17:33–38
Goetz FW, Theofan G (1979) In vitro stimulation of germinal vesicle breakdown and ovulation of yellow perch (Perca flavescens) oocytes. Effects of 17a-hydroxy-20ß-dihydroprogesterone and prostaglandins. Gen Comp Endocrinol 37:273–285
Goetz FW, Duman P, Berndtson A, Janowsky EG (1989) The role of prostaglandins in the control of ovulation in yellow perch, Perca flavescens. Fish Physiol Biochem 7(1–6):163–168
Grozea A, Telea A, Korbuly B (2008) Preliminary study regarding the efficiency of different hormones on pikeperch spermination. Faculty of animal science and biotechnologies. Timisoara 41(2):59–64
Hearn MC (1980) Ovulation of pond-reared walleyes in response to various injection levels of human chorionic gonadotropin. Prog Fish Cult 42:228–230
Henrotte É, Overton JL, Kestemont P (2008) Effects of dietary n-3 and n-6 fatty acids levels on egg and larval quality of eurasian perch. Cybium 32(2 Suppl):271–272
Henrotte E, Mandiki RSNM et al (2010) Egg and larval quality, and egg fatty acid composition of eurasian perch breeders (Perca fluviatilis) fed different dietary DHA/EPA/AA ratios. Aquac Res 41(9):53–61
Hermelink B, Wuertz S, Trubiroha A, Rennert B, Kloas W, Schulz C (2011) Influence of temperature on puberty and maturation of pikeperch, Sander lucioperca. Gen Comp Endocrinol 172:282–292
Hinshaw JM (2006) Species profile: yellow perch. Southern Regional Aquaculture Center publication no 7204. North Carolina State University. Stoneville, MS, USA
Hokanson KEF (1977) Temperature requirements of some percids and adaptations to the seasonal temperature cycle. J Fish Res Board Can 34:1524–1550
Kagawa H, Tanaka H, Okuzawa K, Kobayashi M (1998) GTH II but not GTH I induces final maturation and the development of maturational competence of oocytes of red seabream in vitro. Gen Comp Endocrinol 112:80–88
Kayes T (1977) Reproductive biology and artificial propagation methods for adult perch. In: Soderberg RW (ed) Perch fingerling production for aquaculture, Advisory Report #421. University of Wisconsin Sea Grant Program, Madison, pp 6–21
Kayes TB, Calhert HE (1979) Effect of photoperiod and temperature on the spawning of yellow perch (Perca flavescens). Proc World Maricult Soc 10:306–316
Kestemont P, Cooremans J, Abi-Ayad A, Mélard C (1999) Cathepsin L in eggs and larvae of perch perca fluviatilis: variations with developmental stage and spawning period. Fish Physiol Biochem 21(1):59–64
Kestemont P, Jourdan S, Houbart M, Me’lard C, Paspatis M, Fontaine P, Cuvier A, Kentouri M, Baras E (2003) Size heterogeneity, cannibalism and competition in cultured predatory fish larvae: biotic and abiotic influences. Aquaculture 227:333–356
Koenst WM, Smith LL Jr (1976) Thermal requirements of the early life history stages of walleye, Stizostedion vitreum vitreum, and sauger, Stizostedion canadense. J Fish Res Board Can 33:1130–1138
Kokurewicz B (1969) The influence of temperature on the embryonic development of the perches, Perca fluviatilis L. and Lucioperca lucioperca (L.). Zool Pol 19:47–67
Kolkovski S, Dąbrowski K (1998) Off-season spawning of yellow perch. Prog Fish Cult 60:133–136
Kooten T, Andersson J, Byström P, Persson L, Roos AM (2010) Size at hatching determines population dynamics and response to harvesting in cannibalistic fish. Can J Fish Aquat Sci 67:401–416
Korbuly B, Grozea A, Cean A, Bănăţean-Dunea I, Păcală N (2010) Spawning latency period in hormonal induced reproduction of pikeperch (Sander lucioperca). SPASB 43:32–35
Kouril J, Linhart O, Relot P (1997) Induced spawning of perch by means of a GnRH analogue. Aquac Int 5:375–377
Krejszeff S, Targońska K, Żarski D, Kucharczyk D (2009) Domestication affects spawning of the ide (Leuciscus idus)-preliminary study. Aquaculture 295:145–147
Krejszeff S, Targońska K, Żarski D, Kucharczyk D (2010) Artificial reproduction of two different spawn-forms of the chub. Reprod Biol 10:67–74
Krise WF (1988) Optimum protease exposure time for removing adhesiveness of walleye eggs. Prog Fish Cult 50:126–127
Krise WF, Bulkowski- Cummings L, Shellman AD, Kraus KA, Gould RW (1986) Increased walleye egg hatch and larval survival after protease treatment of eggs. Prog Fish Cult 48:95–100
Křišťan J, Stejskal V, Policar T (2012a) Comparison of reproduction characteristics and broodstock mortality in farmed and wild eurasian perch (Perca fluviatilis L.) females during spawning season under controlled conditions. Turk J Fish Aquat Sci 12:191–197
Křišťan J, Żarski D, Palińska K, Krejszeff S, Nowosad J, Alavi SMH, Policar T, Kucharczyk D (2012b) The effect of sperm to egg ratio on fertilization success in artificial insemination of pikeperch Sander lucioperca (L.). Domestication in Finfish Aquaculture, Department of Lake and River Fisheries of University of Warmia and Mazury, Olsztyn, 23–25 Oct, p 70.
Křišťan J, Alavi SMH, Stejskal V, Policar T (2013) Hormonal induction of ovulation in pikeperch (Sander lucioperca L.) using human chorionic gonadotropin (hCG) and mammalian GnRH analogue. Aquac Int 21:811–818
Król J, Kowalski RK, Hliwa P, Dietrich GJ, Stabiński R, Ciereszko A (2009) The effects of commercial preparations containing two different GnRH analogues and dopamine antagonists on spermiation and sperm characteristics in the European smelt Osmerus eperlanus (L.). Aquaculture 286:328–331
Kucharczyk D, Kujawa R, Mamcarz A, Skrzypczak A, Wyszomirska E (1996) Induced spawning in perch. Perca fluviatilis L. using carp pituitary extract and HCG. Aquac Res 27(11):847–852
Kucharczyk D, Kujawa R, Mamcarz A, Skrzypczak A, Wyszomirska E (1998) Induced spawning in perch, Perca fluviatilis L., using FSH + LH with pimozide or metoclopramide. Aquac Res 29:131–136
Kucharczyk D, Mamcarz A, Kujawa R, Wyszomirska E, Skrzypczak A (2000) Spontaneous and artificial spawning of perch (Perca fluviatilis L.) as studied on individuals collected from two populations. Folia Univ Agric Stetin 214(27):135–146
Kucharczyk D, Szczerbowski A, Łuczynski MJ, Kujawa R, Mamcarz A, Wyszomirska E, Szabó T, Ratajski S (2001) Artificial spawning of Eurasian perch, Perca fluviatilis L. Using ovopel. Arch Pol Fish 9(1):39–49
Kucharczyk D, Kujawa R, Mamcarz A, Targońska-Dietrich K, Wyszomirska E, Glogowski J, Babiak I, Szabó T (2005) Induced spawning in bream (Abramis brama L.) using pellets containing GnRH. Czech J Anim Sci 50:89–95
Kucharczyk D, Kestemont P, Mamcarz A (2007) Artificial reproduction of pikeperch. Mercurius, Olsztyn, 80 p
Kucharczyk D, Targońska K, Hliwa P, Gomułka P, Kwiatkowski M, Krejszeff S, Perkowski J (2008) Reproductive parameters of common carp (Cyprinus carpio L) spawners during natural season and out-of-season spawning. Reprod Biol 8:285–289
Kujawa R, Kucharczyk D, Mamcarz A, Zarski D, Targońska K (2011) Artificial spawning of common tench Tinca tinca (Linnaeus, 1758), obtained from wild and domestic stocks. Aquac Int 19:513–521
Legendre M, Linhart O, Billard R (1996) Spawning and management of gametes, fertilized eggs and embryos in Siluroidei. Aquat Living Resour 9:59–80
Lessman CA (1978) Effects of gonadotropin mixtures and two steroids on inducing ovulation in walleyes. Prog Fish-Cult 40:3–5
Linhart O, Rodina M, Kocour M, Gela D (2006) Insemination, fertilization and gamete management in tench, Tinca tinca (L.). Aquac Int 14:61–73
Malison JA, Procarione LS, Kayes TB, Hansen JF, Held JA (1998) Industion of out-of-season spawning in walleye (Stizostedion vitreum). Aquaculture 163:151–161
Migaud H, Mandiki R, Gardeur JN, Kestemont P, Bromage N, Fontaine P (2003) Influence of photoperiod regimes on the eurasian perch gonad genesis and spawning. Fish Physiol Biochem 28:395–397
Mims SD, Onders RJ, Gomelsky B, Shelton WL (2004) Effectiveness of the minimally invasive surgical technique (MIST) for removal of ovulated eggs from first-time and second-time mist-spawned paddlefish. N Am J Aquac 66:70–72
Minin AA, Ozerova SG (2008) Spontaneous activation of fish eggs is abolished by protease inhibitors. Russ J Dev Biol 38:293–296
Moore AA (2003) Manipulation of fertilization procedures to improve hatchery walleye egg fertility and survival. N Am J Aquac 65:56–59
Müller T, Taller J, Nyitrai G, Kucska B, Cernak I, Bercsenyi M (2004) Hybrid of pikeperch, Sander lucioperca L. and Volga perch, S. viologens (Gamelin). Aquac Res 35(9):915–916
Müller-Bellecke A, Zienert S (2008) Out-of-season spawning of pike perch (Sander lucioperca L.) without the need for hormonal treatments. Aquac Res 39:1279–1285
Mylonas CC, Fostier A, Zanuy S (2010) Broodstock management and hormonal manipulations of fish reproduction. Gen Comp Endocrinol 165:516–534
Nagahama Y (1994) Endocrine regulation of gametogenesis in fish. Int J Dev Biol 38:217–229
Nagahama Y, Yamashita M (2008) Regulation of oocyte maturation in fish. Dev Growth Differ 50:195–219
Patino R, Sullivan CV (2002) Ovarian follicle growth, maturation, and ovulation in teleost fish. Fish Physiol Biochem 26:57–70
Patino R, Yoshizaki G, Thomas P, Kagawa H (2001) Gonadotropic control of ovarian follicle maturation: the two-stage concept and its mechanisms. Comp Biochem Physiol B129:427–439
Patino R, Yoshizaki G, Bolamba D, Thomas P (2003) Role of arachidonic acid and protein kinase c during maturation-inducing hormone-dependent meiotic resumption and ovulation in ovarian follicles of atlantic croaker. Biol Rep 68:516–523
Peter RE (1983) The brain and neurohormones in teleost reproduction. In: Randall D, Hoar WS, Donaldson E (eds) Reproduction endocrine tissues and hormones, vol 9, Part A of fish physiology. Academic, New York, pp 97–135
Peter RE, Yu KL (1997) Neuroendocrine regulation of ovulation in fishes: basic and applied aspects. Rev Fish Biol Fish 7:173–197
Podhorec P, Kouril J (2009) Induction of final oocyte maturation in Cyprinidae fish by hypothalamic factors: a review. Vet Med 54:97–110
Policar T, Toner D, Alavi SMH, Linhart O (2008) Chapter 4: Reproduction and spawning. In: Toner D, Rougeot C (eds) Farming of Eurasian Perch volume 1–Juvenile Production, Aquaculture explained special publication no 24, Dun Laoghaire
Pourasadi M, Falahatkar B, Takami GA (2009) Minimally invasive surgical technique for egg collection from the Persian sturgeon, Acipenser persicus. Aquac Int 17:317–321
Probst WN, Stoll S, Hofmann H, Fischer P, Eckmann R (2009) Spawning site selection by eurasian perch (Perca fluviatilis L.) in relation to temperature and wave exposure. Ecol Freshw Fish 18(1):1–7
Rinchard J, Dabrowski K, Tassell JJ, Stein RA (2005) Optimization of fertilization success in Sander vitreus is influenced by the sperm : egg ratio and ova storage. J Fish Biol 67:1157–1161
Rinchard J, WareK DK, Tassell JJ, Marschall EA, Stein RA (2011) Egg thiamine concentration affects embryo survival in Lake Erie walleye. Environ Biol Fish 90:53–60
Rónyai A (2007) Induced out-of-season and seasonal tank spawning and stripping of pike perch (Sander lucioperca L.). Aquac Int 38:1144–1151
Rónyai A, Lengyel SA (2010) Effects of hormonal treatments on induced tank spawning of eurasian perch (Perca fluviatilis L.). Aquac Int 41(9):345–347
Ross RM (1984) Catheterization: a non-harmful method of sex identification for sexually monomorphic fishes. Prog Fish-Cult 46:151–152
Rougeot C, Nicayenzi F, Mandiki SNM, Rurangwa E, Kestemont P, Mélard C (2004) Comparative study of the reproductive characteristics of XY male and hormonally sex-reversed XX male eurasian perch, Perca fluviatilis. Theriogenology 62:790–800
Saad A, Billard R (1987) Composition et emploi d’un dilueur d’insemination chez la carpe, Cyprinus carpio. Aquaculture 66:329–345
Saat T, Veersalu A (1996) The rate of early development in perch Perca fluviatilis L. and ruffe Gymnocephalus cernuus (L.) at different temperatures. Ann Bot Fenn 33:693–698
Satterfield JR Jr, Flickinger SA (1995a) Factors influencing storage potential of preserved walleye semen. Prog Fish-Cult 57:175–181
Satterfield JR Jr, Flickinger SA (1995b) Field collection and short-term storage of walleye semen. Prog Fish-Cult 57:182–187
Schlumberger O, Proteau JP (1996) Reproduction of pikeperch (Stizostedion lucioperca) in captivity. J Appl Ichthyol 12:149–152
Schlumpberger W, Schmidt K (1980) Vorläufiger Stand der Technologie zur Aufzucht von vorgestreckten Zander (Stizostedion lucioperca). Z Binnenfisch DDR 27:284–286
Schreck CB, Contreras-Sanchez W, Fitzpatrick MS (2001) Effects of stress on fish reproduction, gamete quality, and progeny. Aquaculture 197:3–24
Schulz RW, Renato de França L, Lareyre J-J, LeGac F, Chiarini-Garcia C, Nobrega RE, Miura T (2010) Spermatogenesis in fish. Gen Comp Endocrinol 165:390–411
Skrzypczak A, Kucharczyk D, Mamcarz A, Kujawa R, Furgala-Selezniow G (1998) A new breeding technique of pikeperch (Stizostedion lucioperca) in net cages. Czech J Anim Sci 43:398–398
Sorensen P, Murphy C, Loomis K, Maniak P, Thomas P (2004) Evidence that 4-pregnen-17,20beta,21-triol-3-one functions as a maturation-inducing hormone and pheromonal precursor in the percid fish, Gymnocephalus cernuus. Gen Comp Endocrinol 139(1):1–11
Stacey N (2003) Hormones, pheromones and reproductive behavior. Fish Physiol Biochem 28:229–235
Steffens W, Geldhauser F, Gerstner P, Hilge V (1996) German experiences in the propagation and rearing of fingerlink pikeperch (Stizostedion lucioperca). Ann Zool Fenn 33:627–634
Sulistyo I, Rinchard J, Fontaine P, Gardeur J, Capdeville B, Kestemont P (1998) Reproductive cycle and plasma levels of sex steroids in female eurasian perch Perca fluviatilis. Aquat Living Resour 11(2):101–110
Szczerbowski A, Kucharczyk D, Mamcarz A, Łuczyński M, Targońska K, Kujawa R (2009) Artificial off-season spawning of eurasian perch Perca fluviatilis L. Arch Pol Fish 17(2):95–98
Targońska K, Kucharczyk D, Kujawa R, Mamcarz A, Żarski D (2010) Controlled reproduction of asp, Aspius aspius (L.) using luteinizing hormone releasing hormone (LHRH) analogues with dopamine inhibitors. Aquaculture 306:407–410
Targońska K, Szczerbowski A, Żarski D, Łuczyński M, Szkudlarek M, Gomułka P, Kucharczyk D (2014) Comparison of different spawning agents in artificial out-of-season spawning of Eurasian perch, Perca fluviatilis L. Aquac Res 45:765–767
Teletchea F, Gardeur J-N, Psenicka M, Kaspar V, Le Dore Y, Linhart O, Fontaine P (2009a) Effects of four factors on the quality of male reproductive cycle in pikeperch Sander lucioperca. Aquaculture 291(3–4):217–223
Teletchea F, Gardeur JN, Kamler E, Fontaine P (2009b) The relationship of oocyte diameter and incubation temperature to incubation time in temperate freshwater fish species. J Fish Biol 74:652–668
Thorgaard GH (1995) Biotechnological approaches to broodstock management. In: Bromage NR, Roberts RJ (eds) Broodstock management and egg and larval quality. Blackwell Science, Oxford, pp 76–93
Tyler CR, Sumpter JP (1996) Oocyte growth and development in teleosts. Rev Fish Biol Fish 6:287–318
Vizziano D, Fostier A, Loir M, LeGac F (2008) Testis development, its hormonal regulation and spermiation induction in teleost fish. In: Alavi SMH, Cosson JJ, Coward K, Rafiee G (eds) Fish spermatology. Alpha Science, Oxford, pp 103–139
Wang N, Gardeur JN, Henrotte E, Marie M, Kestemont P, Fontaine P (2006) Determinism of the induction of the reproductive cycle in female Eurasian perch, Perca fluviatilis: identification of environmental cues and permissive factors. Aquaculture 261:706–714
Wojda R, Sliwinski J, Ciesla M (1994) Results of studies on natural spawning in pikeperch, Stizostedion lucioperca (L.). Rocz Nauk PZW 7:71–80 (In Polish with English summary)
Woynarovich E, Horváth L (1980) The artificial propagation of warmwater finfishes – a manual for extension. FAO Fisheries Technical Paper, Rome, Italy. (201), 183 p
Woynarovich E, Woynarovich A (1980) Modified technology for elimination of stickiness of common carp Cyprinus carpio eggs. Aquacult Hung 2:19–21
Yamashita M, Fukada S, Yoshikuni M, Bulet P, Hirai T, Yamaguchi A, Lou YH, Zhao Z, Nagahama Y (1992) Purification and characterization of maturation-promoting factor in fish. Dev Biol 149:8–15
Zakęś Z, Demska-Zakęś K (2005) Artificial spawning of pikeperch (Sander lucioperca (L.)) stimulated with human chorionic gonadotropin (hCG) and mammalian GnRH analogue with a dopamine inhibitor. Arch Pol Fish 13:63–75
Zakęś Z, Demska–Zakęś K (2009) Controlled reproduction of pikeperch Sander lucioperca (L.): a review. Arch Pol Fish 17:153–170
Żarski D (2012) First evidence of pheromonal stimulation of maturation in Eurasian perch, Perca fluviatilis L., females. TrJFAS 12:771–776
Żarski D, Bokor Z, Kotrik L, Urbányi B, Horváth A, Targońska K, Krejszeff S, Palińska K, Kucharczyk D (2011a) A new classification of a preovulatory oocyte maturation stage suitable for the synchronization of ovulation in controlled reproduction of Eurasian perch, Perca fluviatilis L. Reprod Biol 3:194–209
Żarski D, Palińska K, Targońska K, Bokor Z, Kotrik L, Krejszeff S, Kupren K, Horvath A, Urbanyi B, Kucharczyk D (2011b) Oocyte quality indicators in Eurasian perch, Perca fluviatilis L., during reproduction under controlled conditions. Aquaculture 313:84–91
Żarski D, Kucharczyk D, Targońska K, Palińska K, Kupren K, Fontaine P, Kestemont P (2012a) A new classification of pre-ovulatory oocyte maturation stages in pikeperch, Sander lucioperca (L.), and its application during artificial reproduction. Aquac Res 43:713–721
Żarski D, Krejszeff S, Palińska K, Targońska K, Kupren K, Fontaine P, Kestemont P, Kucharczyk D (2012b) Cortical reaction as an egg quality indicator in artificial reproduction of pikeperch, Sander lucioperca. Reprod Fertil Dev 24:843–850
Żarski D, Krejszeff S, Horváth Á, Bokor Z, Palińska K, Szentes K, Łuczyńska J, Targońska K, Kupren K, Urbányi B, Kucharczyk D (2012c) Dynamics of composition and morphology in oocytes of Eurasian perch, Perca fluviatilis L., during induced spawning. Aquaculture 364–365:103–110
Żarski D, Horvath A, Kotrik L, Targońska K, Palińska K, Krejszeff S, Bokor Z, Urbanyi B, Kucharczyk D (2012d) Effect of different activating solutions on the fertilization ability of Eurasian perch, Perca fluviatilis L., eggs. J Appl Ichthyol 28:967–972
Żarski D, Targońska K, Kaszubowski R, Kestemont P, Fontaine P, Krejszeff S, Kupren K, Kucharczyk D (2013a) Effect of different commercial spawning agents and thermal regime on the effectiveness of pikeperch, Sander lucioperca (L.), reproduction under controlled conditions. Aquac Int 21:819–828
Żarski D, Křišťan J, Palińska-Żarska K, Krejszeff S, Policar T, Kucharczyk D (2013b) The period of pikeperch, Sander lucioperca (L.), eggs ability to fertilization and its dependence on egg quality. Diversification in Inland Finfish Aquaculture II (DIFA II), 24–26 Sept, University of South Bohemia in České Budějovice, Faculty of Fisheries and Protection of Waters, Vodnany
Żarski D, Horváth A, Bernáth G, Palińska-Żarska K, Krejszeff S, Müller T, Kucharczyk D (2014) Application of different activating solutions to in vitro fertilization of crucian carp, Carassius carassius (L.), eggs. Aquac Int 22:173–184
Żarski D, Targońska K, Palińska K, Kucharczyk D, Krejszeff S, Kupren K, Fontaine P, Kestemont P (2015a) The application of tannic acid to the elimination of egg stickiness at varied moments of the egg swelling process in pikeperch, Sander lucioperca (L.). Aquac Res 46:324–334
Żarski D, Cejko BI, Krejszeff S, Palińska-Zarska K, Horváth Á, Sarosiek B, Judycka S, Kowalski RK, Łaczyńska B, Kucharczyk D (2015b) The effect of osmolality on egg fertilization in common carp, Cyprinus carpio Linnaeus, 1758. J Appl Ichthyol 31:159–163
Zohar Y, Mylonas CC (2001) Endocrine manipulations of spawning in cultured fish: from hormones to genes. Aquaculture 197:99–136
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Żarski, D., Horváth, A., Held, J.A., Kucharczyk, D. (2015). Artificial Reproduction of Percid Fishes. In: Kestemont, P., Dabrowski, K., Summerfelt, R. (eds) Biology and Culture of Percid Fishes. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-7227-3_4
Download citation
DOI: https://doi.org/10.1007/978-94-017-7227-3_4
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-7226-6
Online ISBN: 978-94-017-7227-3
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)