
Abstract
Circulating CRC markers might become useful tools for the massive screening of people since they satisfy a high degree of accuracy, noninvasiveness, reproducibility, and economy. Within circulating biomarkers, we will focus on the detection of autoantibodies in serum from cancer patients and their target tumor-associated antigens (TAAs). Although the sensitivity, specificity, and predictive value of individual TAAs present low scores, the combination of multiple TAAs shows improved values to discriminate between CRC patients and controls. In this review, we outline the methodologies used to identify circulating autoantibodies and their target proteins and discuss the relevance of CRC autoantibodies for diagnosis at, particularly, early stages. An overview of the reported biomarkers is given, showing the large complexity of the autoantibody response in cancer. Different strategies to improve CRC diagnostic tests by combining autoantibodies from different studies will be discussed. Association of autoantibodies to prognosis, recurrence, and the survival of patients will be introduced. We conclude that there is a great potential for the use of autoantibodies as diagnostic CRC biomarkers in the near future.
Access provided by Autonomous University of Puebla. Download reference work entry PDF
Similar content being viewed by others


Keywords
- Colorectal cancer
- Humoral response
- Autoantibody
- Cancer immune response
- Biomarker
- Circulating biomarker
- Protein microarrays
- SEREX
- SERPA
- Phage display
Key Facts of Circulating Biomarkers
-
Tumor-associated antigens (TAAs) are self-proteins altered during tumor formation and progression.
-
Cancer humoral responses take place against TAAs.
-
Autoantibodies against TAAs can be used in colorectal cancer patients as circulating biomarkers.
-
Immunoproteomics provide useful approaches for identifying autoantibodies and their reactive TAA targets.
-
Circulating autoantibodies provide an effective, reliable, and reproducible tool in cancer patients for diagnosis, prognosis, recurrence, and therapy monitoring.
-
Targets of circulating autoantibodies might be novel therapeutic targets for intervention.
Definitions of Words and Terms
Biomarker
A biological marker usually a protein or glycoprotein that can be used as an indicator of a biological or pathological state or condition.
TAA
Tumor-associated antigen which consists of an altered self-protein able to induce an immune response in cancer patients.
Autoantibody
Immunoglobulin G produced in response to self-proteins altered during tumor formation or progression which can be used as a cancer biomarker.
ELISA
Enzyme-linked immunosorbent assay which permits to detect the presence of an immune response to a specific TAA in a biological fluid (i.e., serum or plasma) by means of an enzyme immunoassay.
SEREX
Acronym of serological analysis of recombinant cDNA expression libraries (SEREX) which is a technology based on the detection of tumor-associated antigens within recombinantly expressed tumor cDNA phage libraries by autologous antibodies.
SERPA
Acronym of serological proteome analysis where proteins from tumoral samples are resolved by 2D gels to identify TAAs by immunostaining with sera from cancer patients and controls. TAAs are then identified in a subsequent step by LC–MS.
LC–MS
Liquid chromatography coupled to mass spectrometry. High-performance liquid chromatography (LC) coupled to mass spectrometry (MS) to separate a peptide mixture in a liquid phase according to hydrophobic interactions provided by the C18 alkyl chains of the stationary phase of the chromatography column.
Protein Microarray
A chip containing thousands to tens of thousands of different proteins attached to a support surface of glass slide or nitrocellulose membrane.
Phage Display
This technique is used for the high-throughput screening of protein interactions and consists of libraries of millions of different M13 or T7 phages displaying in their surface proteins fused to the N- or C-terminal end, respectively, of proteins of the capsid of the phage.
ROC Curve
Receiver-operating characteristic (ROC) curves illustrate the performance of a diagnostic test. It is obtained by plotting the fraction of true-positive cases among the actual number of positive patients versus the fraction of false-positive controls out of the total actual negative controls at various threshold settings.
PTM
Posttranslational modification. After translation, proteins can be modified by the attachment of functional groups, including phosphorylation, acetylation, carbohydrate, lipid, etc.
CEA
Carcinoembryonic antigen is a recommended clinical biomarker for colorectal cancer recurrence but not for screening.
Introduction
Colorectal cancer (CRC) is the major cause of cancer-associated mortality in developed countries (Duffy et al. 2007). CRC is mainly diagnosed at late stages, when patients have developed clinical symptoms and the tumor has already spread to adjacent lymph nodes or colonized other organs. At that point, 61 % of CRC tumors are so advanced that the 5-year survival rate of patients is only about 11 %, with 5-year survival rates of 91.1 % and 69.8 % for localized and regional stages, respectively (Surveillance, Epidemiology, and End Results Program (2011), National Cancer Institute, USA). If we could shift the diagnosis of CRC patients to early cancer stages, their final outcome would substantially improve.
A suitable biomarker for a screening test should demonstrate a high degree of accuracy, reproducibility, economy, and, more importantly, acceptance by the population. Actually, the current markers for CRC, CEA, CA19.9, and CA125 are not recommended for clinical screening (Levin et al. 2003; Locker et al. 2006; Duffy et al. 2007), because they can be altered in other diseases different from CRC and even in nonpathological states. Moreover, circulating CEA is only recommended to monitor therapy in advanced CRC and for prognostic information (Locker et al. 2006; Duffy et al. 2007).
Circulating biomarker molecules might consist of altered tumoral proteins, which are leaked to the blood. However, they are usually found at very low concentrations and exposed to degradation (Villanueva et al. 2006), making them questionable as diagnostic biomarkers and their actual discovery a challenge (Villanueva et al. 2006; Barderas et al. 2010; Casal and Barderas 2010). However, some cancer proteins are able to induce a humoral response in cancer patients, providing an effective, reliable, and noninvasive tool for cancer screening and preclinical diagnosis (Fig. 1; Anderson and LaBaer 2005; Murphy et al. 2012b). The molecular mechanisms of this humoral response to cancer proteins are rather uncharacterized. It might be due to the alterations of self-proteins during tumor formation and progression, punctual mutations, truncations, aberrant glycosylations, overexpression, or aberrant degradation (Anderson and LaBaer 2005; Murphy et al. 2012b).

Production of cancer autoantibodies against tumoral self-proteins. During tumor formation and progression, altered self-proteins and cellular debris are released from colorectal neoplastic tissue, which results in B-cell proliferation and antibody production when antigen-presenting cells present these proteins to the immune response
Novel high-throughput proteomic approaches have accelerated the identification of circulating serum autoantibodies and their respective target proteins as potential cancer biomarkers. Antibodies are highly stable molecules with a long tradition of being used in immunoassays, thus facilitating their standardization. In the last years, the apparition of new techniques has delivered multiple studies related to the use of autoantibodies for cancer detection. In this review, we describe the features of circulating serum autoantibody biomarkers for CRC and outline the proteomic strategies employed to identify them since the first reports by Old’s laboratory (Fig. 2). We will review the validation and optimization experiments required for use in clinical practice. CRC autoantibodies and their target proteins should facilitate the screening for early diagnosis, prognosis, and monitoring of therapy in CRC patients, as well as for the identification of novel therapeutic targets.

Proteomic strategies used to identify CRC circulating autoantibodies. *, These strategies require a further step of identification of the TAA via sequencing (SEREX and phage microarrays) or LC–MS (SERPA and natural protein microarrays)
Strategies to Identify CRC Circulating Autoantibodies in Cancer Patients
The first reports identifying autoantibodies and cancer autoantigens were published in the late 1970s and early 1980s using an approach called autologous serological typing (Carey et al. 1976; Dippold et al. 1980; Fig. 2), where circulating autoantibodies were identified using tumoral cells in culture and sera from the same patients. Although researchers identified several melanoma tumor-associated antigens (TAAs), the heterogeneity of cancer patient’s response complicated the antigen identification (Carey et al. 1976; Dippold et al. 1980).
SEREX and SERPA
From those early studies, different low- or medium-throughput technologies such as serological analysis of recombinant cDNA expression libraries (SEREX ) or serological proteome analysis (SERPA) have been applied for the discovery of autoantibodies and their target TAAs as cancer diagnostic markers with different successes (Fig. 2; Sahin et al. 1995; Casal and Barderas 2010). SEREX involves the identification of TAAs using phage display expression libraries containing cDNA from tumoral tissues or cells, transferred onto membranes for screening with cancer sera. In SERPA, proteins from tumoral samples are separated in 2D gels to identify TAAs after transference to membranes for immunostaining with cancer sera and healthy controls (Klade et al. 2001).
A variant of SEREX , where phages are printed on a microarray format, has been also tested for autoantibody discovery in a high-throughput version (Wang et al. 2005; Chatterjee et al. 2006; Babel et al. 2011; Chatterjee et al. 2012). Here, TAA discovery takes advantage on the multiplexing analysis of both T7 phages and microarrays (Fig. 2). cDNA from a specific tumor is inserted in the genome of T7 phages, and cancer peptides and proteins are displayed on the surface of the capsid of the phage at the C-terminal end of 10B protein. Alternatively, a random peptide library can also be used instead of tumoral cDNA libraries (Mintz et al. 2003; Jiang et al. 2010). Then, several rounds of biopanning using sera from healthy individuals and cancer patients permit the enrichment of T7 phage libraries on phages displaying cancer-specific peptides or proteins. Finally, thousands of T7 phages are printed onto nitrocellulose microarrays to be screened with sera from cancer patients and controls, and TAAs can be (theoretically) identified after sequencing the cDNA inserted in the genome of the phage.
Although these techniques have permitted the identification of multiple TAAs, they present several problems. SEREX is technically demanding, is labor intensive, and presents several important bottlenecks in the construction of the phage libraries and the identification of the proteins displayed in the phages. In many occasions, it is impossible to identify peptides homologous to known proteins. This might be due to problems in the insertion of the cDNAs correctly in-frame or the cloning of noncoding regions. Despite this, SEREX has been probably the most commonly used technology for TAA discovery. SERPA is biased toward the identification of highly abundant proteins.
Recombinant Protein Microarrays
The low throughput of SEREX and SERPA and their limitations have pushed the field to look for alternatives in the screening of the humoral response against cancer (Fig. 2; Anderson and LaBaer 2005; Casal and Barderas 2010). To this end, high-content protein microarrays have been prepared with thousands of different probes printed on the surface of a chip. They have been used to identify substrates of kinases and small-molecule interactors and detect protein–protein interactions and also to track autoantibodies (Zhu et al. 2001; Zhu and Snyder 2001; Liotta et al. 2003). Protein microarrays provide the best tool to identify circulating cancer autoantibodies with high sensitivity and specificity in order to get the highest coverage of cancer patients. Recombinant proteins printed in the microarray are known beforehand, which facilitates their identification (Zhu and Snyder 2001). Proteins are expressed and purified before being printed at similar concentrations on the chip (Zhu et al. 2001), which increases the chance to discover circulating autoantibodies directed to low-abundant proteins (Babel et al. 2009; Barderas et al. 2010; Casal and Barderas 2010).
There are protein microarrays printed with proteins expressed in insect cells, which contain posttranslational modifications (PTMs) similar to those found in mammalian cells (Hudson et al. 2007; Babel et al. 2009). Commercially available protein arrays containing 9,500 human recombinant proteins expressed in Sf9 insect cells (ProtoArrayTM, Invitrogen) have been used to identify TAAs in different cancer types (Hudson et al. 2007; Babel et al. 2009; Orenes-Pinero et al. 2010). However, in some cases, protein arrays express only fragments of many proteins, which change their conformation and folding and affect antibody recognition. Other alternatives consist of microarrays printed with the hEx1 human cDNA library, containing about 10,000 unique human proteins expressed in bacteria (Bussow et al. 1998, 2000). However, proteins expressed in bacteria lack most of the PTMs.
Importantly, a protein microarray technology is still being developed, and improvements are reported annually (Zhu and Snyder 2001; Liotta et al. 2003; Ramachandran et al. 2004; Chatterjee et al. 2006; Kung and Snyder 2006; Oishi et al. 2006; Hudson et al. 2007; Ran et al. 2008; Babel et al. 2009; Orenes-Pinero et al. 2010; Babel et al. 2011; Pedersen et al. 2011; Takulapalli et al. 2012; Festa et al. 2013). From the first commercially available human protein microarray consisting of 5,000 proteins in 2006, the number of proteins contained in the microarrays has grown up to 9,500 human full-length recombinant proteins in only one chip. Considering the advances performed in DNA microarrays, where microarrays covering the whole genome are available (Kronick 2004), we expect that the features in protein microarrays will keep growing up in the next years. Additionally, there is an alternative called NAPPA, where cDNAs codifying for the different proteins are printed on the surface of the chips and translated into proteins with rabbit reticulocyte lysates or a mammalian cancer cell milieu (Festa et al. 2013). Still, NAPPA technology has not been widely used, and its value remains to be demonstrated at a large scale in multiple laboratories.
Natural Protein Microarrays
In this case, protein extracts isolated from cancer cell lines are fractionated by liquid chromatography before being printed in the microarray slides (Qiu et al. 2004). The main advantage of this approach consists of the preservation of naturally occurring PTMs and other alterations observed in cancer proteins, which include alternative splicing, chimeras, or aberrant glycosylated proteins. This facilitates the display of relevant critical epitopes of the cancer patient’s autoantibodies. Even though natural protein microarrays present advantages, the spots identified are complex mixtures of proteins, which complicate the actual identification of the target protein by mass spectrometry (Qiu et al. 2004). This technology only has been applied few times for the identification of cancer autoantibodies, including colon cancer (Nam et al. 2003; Qiu et al. 2004).
Glycopeptide Microarrays
This approach relies on the fact that malignant transformation of cells is accompanied by alterations in O-glycosylations able to induce a humoral immune response in cancer patients (Wandall et al. 2010). Glycopeptide microarrays are printed with peptides and O-glycosylated recombinant fragments purified by HPLC, which are obtained in vitro using recombinant ppGalNAc-transferases, Drosophila Core-1 b3GalT, human Core3 b3GnT6, and human ST6GalNAc-I (Wandall et al. 2010; Pedersen et al. 2011). These microarrays have been used to characterize humoral immune responses directed against aberrant glycosylated epitopes in colorectal cancer (Wandall et al. 2010).
Application of Circulating Autoantibodies to Colon Cancer Diagnostic Biomarker Discovery
SEREX and Phage Microarrays
The first report describing autoantibodies in CRC was published in 1998 by Old’s group using SEREX (Scanlan et al. 1998). The authors identified 48 antigens, representing a broad spectrum of cellular components, with 31 clones as products of known genes and 17 representing novel gene products. The authors identified six clones that reacted exclusively with sera from colon cancer patients, with a frequency of recognition between 10 % and 27 %, indicating the great heterogeneity in the humoral immune response. Twenty out of 29 serum samples from colon cancer patients detected 1 or more of these clones.
The same group published 4 years later another SEREX study. Thirteen out of 77 antigens tested – p53, MAGEA3, SSX2, NY-ESO-1, HDAC5, MBD2, TRIP4, NYCO-45, KNSL6, HIP1R, Seb4D, KIAA1416, and LMNA – were serologically reactive with sera from CRC patients (Scanlan et al. 2002). In this study, 34 out of 74 colon cancer patients (46 %) were detected to have 1 or more of these 13 antigens. Several phage-displayed cDNA clones recognized in CRC patients showed no homology to sequences deposited in databases. This fact is a common problem of SEREX, which is observed in most of the published reports. The displayed peptide could mimic a conformational epitope or there could be other reasons (see above). Line and coworkers identified eight different serum-reactive cDNA clones with three potential targets C21ORF2, EPRS, and NAP1L1 not recognized by the sera of healthy individuals, but with frequencies of recognition between 4 and 17 % for the sera of CRC patients (Line et al. 2002). Ishikawa et al., also by SEREX, performed a screening of colon cancer cell lines with microsatellite instability (MSI) using the serum from a patient with MSI. The authors found 64 genes immunoreactive in colon cancer, with one of the identified antigens (CDX2) carrying a frameshift mutation in the repetitive G sequences (microsatellite) of its coding region, indicating that the immune response was also raised against TAAs generated by MSI (Ishikawa et al. 2003). Interestingly, the authors found galectin-4 among the immunoreactive clones as previously described (Scanlan et al. 1998). The frequency of recognition of this protein was low and not colon cancer specific, since it was also observed in renal, pancreatic, and esophageal cancer patient’s sera and in 8 out of 38 healthy individuals (Ishikawa et al. 2003). Also by SEREX , it was found that HDAC3 autoantibodies were increased in CRC patients in comparison to controls. The authors found that the C-terminal region of HDAC3, with no homology to HDAC1 and HDAC2, contained the B-cell epitopes recognized by circulating autoantibodies (Shebzukhov et al. 2005).
In 2008, Ran et al. found six phages by SEREX technology able to classify CRC patients from healthy controls (Ran et al. 2008), with two of them showing partial homology to LGR6 and C6orf192 and the other four without obvious homology to any protein. Song et al. in 2011 identified BCP-20 (FBXO39) as a potential autoantigen, but with a prevalence of only 6 % in colon cancer patients (Song et al. 2011). Another study performed that year with a 12-mer random peptide library displayed in phages permitted the identification of a peptide motif (LYSNTL) with homology to ARD1A (Jiang et al. 2010). The authors found that the full-length recombinant protein elicited autoantibodies only in 14.7 % of CRC patients and 2 % of healthy controls (Jiang et al. 2010). Overexpression of ARD1A in CRC tumors was associated to poor prognosis for disease-free survival and shorter overall survival (Jiang et al. 2010). Chang et al. identified a five-phage classifier able to discriminate between CRC patients and controls. Phages were homologous to the uncharacterized protein hcg2038983, to TAS2R39, and to a peptide of the C-terminal constant region of the heavy chain of human IgG, which seems quite unusual, with two phages displaying peptides with no homology to any known protein (Chang et al. 2011). In 2011, Babel et al. by combining phage display and protein microarrays identified 43 phages showing higher immunoreactivity with CRC sera than control sera (Babel et al. 2011). Six phages displayed peptides with some homology to STK4/MST1, SULF1, NHSL1, SREBF2, GRN, and GTF2i amino acid sequences. Interestingly, the replacement of two phages for their corresponding recombinant proteins (STK4/MST1, SULF1) significantly improved the diagnostic ability of the predictor panel from an AUC of 0.78 % to 0.86 % and sensitivity from 72 % to 82.6 % while maintaining the specificity higher than 70 % (Babel et al. 2011).
SERPA and Natural Protein Microarrays
Hanash laboratory applied natural protein arrays printed with LoVo cells to the study of the humoral response in colon cancer (Nam et al. 2003). They identified the ubiquitin C-terminal hydrolase L3 (UCH-L3) as a protein target of autoantibodies in 19 out of 43 CRC patients’ sera, without reactivity in lung cancer patients or healthy individuals’ sera (Nam et al. 2003). The authors verified the reactivity of the UCH-L3 protein by immunostaining of 2D gels run with LoVo cell extracts. In 2007, SERPA was used on HCT116 colon cancer cell line. The authors only found HSP60 as a target, with increased autoantibody levels in 13 out of 25 CRC patients in comparison to 1 out of 15 healthy controls for a 52 % sensitivity and 93.3 % specificity (He et al. 2007). SERPA as well as natural protein microarrays presents the same disadvantage: the immunoreactive proteins need to be identified a posteriori by LC–MS. This is complicated by the complexity of the samples, due to the presence of several proteins in the 2D spots. This fact probably explains the low number of reports identifying immunoreactive proteins by these two approaches.
ELISA Approaches
The first report using ELISA plates coated with TAAs to identify colon cancer autoantibodies was published in 2005. The authors used Fas as a target of autoantibodies based on previous observations claiming that apoptosis and the expression of this receptor were altered during carcinogenesis in the human colon. Anti-Fas titers were significantly higher in patients with colorectal adenomas than in healthy controls and even higher in patients with adenocarcinomas (Reipert et al. 2005). Moreover, autoantibodies to Fas/CD95 showed also some specificity for early CRC detection using sera from 38 healthy controls, 38 patients with colorectal adenomas, and 21 patients with colorectal adenocarcinoma in an ELISA-based array (Reipert et al. 2005).
Kocer et al. found autoantibodies directed to MUC5AC in 45 % of polyp patients, 60 % of CRC patients, and 27.3 % of healthy individuals (Kocer et al. 2006). MUC5AC is a secreted mucin aberrantly expressed in colorectal polyps and carcinomas. In 2007, Chen et al. investigated the humoral immune response of Calnuc by ELISA (Chen et al. 2007). They found autoantibodies against Calnuc in 11.5 % of CRC patients’ sera and only in 1.2 % of control sera (Chen et al. 2007). After testing Calnuc in combination with other previously described TAAs, c-myc, p53, G2/mitotic-specific cyclin-B1 (CCNB1), and G1/S-specific cyclin-D1 (CCND1), the ability to diagnose CRC increased up to 65.4 % sensitivity and 93.9 % specificity (Chen et al. 2007). In 2009, Liu et al. tested by ELISA five TAAs previously identified in other cancers with 46 CRC patients’ sera and 58 healthy individual sera (Liu et al. 2009). Imp1, p62, Koc, p53, and c-myc, tested by ELISA, were permitted to discriminate CRC patients from controls with a sensitivity of 60.9 % and a specificity of 89.7 % (Liu et al. 2009). In 2011, two other different reports identified a humoral immune response against two proteins overexpressed in CRC tissue in comparison to normal mucosa – RPH3AL and SEC61β (Chen et al. 2011; Fan et al. 2011). Recently, Liu et al. in 2013 found autoantibodies against the IGF2 mRNA-binding protein (IMP2/p62) by ELISA in 23.4 % of CRC patients’ sera in comparison to 4.8 % of patients’ sera having colonic adenomas and 2.9 % of healthy individuals (Liu et al. 2013).
Protein and Glycopeptide Microarrays
Regarding protein microarrays, Babel et al. used a commercial ProtoArray v4.0 containing 8,000 human recombinant proteins with sera of metastatic CRC patients and controls (Babel et al. 2009). The authors found 43 protein targets of autoantibodies composing a diagnostic protein signature (Babel et al. 2009). Six TAAs (PIM1, MAPKAPK3, STK4, SRC, FGFR4, and ACVR2B) were validated with a different cohort of CRC sera. An AUC of 0.85 % with a specificity and sensitivity of 73.9 % and 83.3 %, respectively, was obtained with the combination of only two markers – ACVR2B and MAPKAPK3 (Babel et al. 2009). In addition, the same authors reanalyzed these recombinant TAAs (Babel et al. 2009) together with six validated phages displaying CRC-specific peptide TAAs (Babel et al. 2011) using a different set of 96 sera. Remarkably, a CRC predictor panel composed of three phages displaying peptides with homology to GRN, NHSL1, and SREBF2 and four full-length recombinant TAAs – PIM1, MAPKAPK3, FGFR4, and ACVR2B – correctly predicted the presence of cancer with a highly improved accuracy showing an AUC of 94 % for a sensitivity and specificity of 89.1 % and 90.0 %, respectively (Barderas et al. 2012). Later, Casal’s group tested a variant version of the commercial ProtoArray v4.1 protein microarrays with a different nitrocellulose surface than ProtoArray v4.0, but containing the same number of human recombinant proteins (Barderas et al. 2013). After identifying 24 novel TAAs, the authors validated 3 TAAs (EDIL3, GTF2B, and HCK) with a different cohort of sera that correctly discriminate 50 CRC samples from 49 healthy individuals in combination with p53 for an AUC of 0.75 % and sensitivity and specificity of 82 % and 56 %, respectively (Barderas et al. 2013). In 2010, Kijanka et al. used high-density protein microarrays printed with the hEx1 human cDNA library with 43 CRC patient sera and 40 control sera. The authors identified a protein signature of 18 antigens associated to cancer and 4 to the absence of cancer, including p53, HMGB1, TRIM28, TCF3, LASS5, and ZNF346 (Kijanka et al. 2010), which was not further validated by other immunological approaches.
Interestingly, our group also proved that some TAAs can be used as potential therapeutic targets. In particular, blocking of FGFR4 with different small-molecule inhibitors and specific antibodies resulted in a significant reduction of tumor growth (Pelaez-Garcia et al. 2013). The silencing of this receptor in colon cancer cells reverted the epithelial to mesenchymal transition (EMT), producing a decrease in the tumorigenic properties of colon cancer cells (Pelaez-Garcia et al. 2013).
A glycopeptide array approach was also tested in 2011 for identifying aberrant glycopeptides as targets of an immune response in CRC patients (Pedersen et al. 2011). This approach allowed for the identification of a set of aberrant glycopeptides derived from MUC1 and MUC4 (Pedersen et al. 2011). The authors validated the immunogenicity of these glycopeptides using monoclonal antibodies and provided evidence of the expression of these aberrant glycosylated proteins in colon cancer cell lines (Pedersen et al. 2011). In addition, the same group also reported that MUC1-STn and MUC1-Core3 circulating IgG autoantibodies were able to discriminate CRC cases from controls with 8.2 % and 13.4 % sensitivity, respectively, at 95 % specificity in a cohort of 97 postmenopausal women with colorectal cancer and 97 postmenopausal women without any history of cancer (Pedersen et al. 2014).
Shortfalls and Limitations of Autoantibody Research
Interestingly, the use of protein microarrays provides the best results in terms of specificity, sensitivity, and predictive value of the identified TAAs for the discrimination of CRC patients and controls (Table 1). However, the poor matching between the identified TAAs by using different approaches, except for STK4 and MAPKAPK3, with galectin-4, p53, c-myc, p62, Koc, and Imp1 as cancer TAAs but nonspecific for CRC is worrisome. This poor coincidence is probably a consequence of multiple variables including the different sensitivities of the techniques and platforms, different repertoires of proteins printed in the microarrays, different expression systems for protein production, different tags fused to the proteins, or different PTMs in the proteins printed in microarrays with respect to the cancer forms.
Another problem is the absence of validation assays in many TAA discovery reports. Although an important number of autoantibodies have been identified in colon cancer patients, some reports used a small number of sera and nonmatching samples between CRC patient sera and control sera with regard to sex and age, and there is an absence of validation data using different sera cohorts. In this sense, only autoantibodies to MST1/STK4 and p53 have been reported in at least three different studies using different cohorts of serum samples (Table 1; Soussi 2000; Babel et al. 2009, 2011; Barderas et al. 2012), with p53 autoantibody frequency ranging between 20 % and 40 % in all cancer patients (Soussi 2000).
Early Responses
Early detection is critical in cancer diagnostics. Although different reports have claimed the utility of this approach for the early diagnosis of CRC (Barderas et al. 2010), few reports have tested autoantibody panels for early diagnosis. A predictor panel composed of a panel of 7 CRC-specific TAAs achieved an AUC of 90 % and a sensitivity of 88.2 % and specificity of 82.6 % for early stages (Duke’s stage A and B) (Barderas et al. 2012). It has also been reported that humoral responses to p53 can appear in normal-risk individuals between 1.0 and 3.8 years before clinical diagnosis of CRC (Pedersen et al. 2013).
However, little is known about the exact moment when autoantibodies appear, their kinetics, and their evolution according to the progression of the disease. The answer to these questions is almost impossible to be performed using human samples due to the enormous difficulties to get preclinical samples. Even more, when the lesions are removed and the patient treated, the evolution of the autoantibodies cannot be monitored. In this sense, by using colon cancer murine models based on the use of azoxymethane/dextran sodium sulfate, our group recently demonstrated that (i) the autoantibodies were produced very early in the disease, even before the tumoral lesions might be detected by other techniques (histology), (ii) the autoantibody levels increased according to the progression of the disease, and (iii) the presence of autoantibodies was always associated to the presence of malignant lesions, since autoantibodies were not observed in those animals presenting nonmalignant lesions (Fig. 3; Barderas et al. 2013). A subset of human CRC TAAs, HCK, MST1/STK4, EDIL3, GTF2B, SRC, NY-ESO-1, MAPKAPK3, and p53 – identified through protein microarray or SEREX approaches (Soussi 2000; Scanlan et al. 2002; Babel et al. 2009, 2011; Barderas et al. 2013) – was tested using sera from these CRC murine models, which develop a cancer humoral response similar to that observed in human CRC patients (Barderas et al. 2013). This subset of human TAAs was able to discriminate mice carrying malignant lesions from controls (Barderas et al. 2013), showing a great potential to be included in CRC diagnostic tests. Indeed, we demonstrated the exquisite sensitivity of the humoral immune response to colon cancer for preclinical cancer diagnosis. Moreover, as autoantibody levels increase according to the progression of the disease, it might open its application to detect recurrences.


Murine models mimic the humoral immune response observed in human CRC patients. (a) The protocol used to obtain chemically induced colitis-associated CRC in mice is outlined. AOM was injected intraperitoneally at a concentration of 10 mg/kg and DSS (2.5 %) supplied in drinking water. (b) Hematoxylin and eosin staining of distal colon tissue at the end of the protocol (day 63). Images are shown at 200× magnification. (c) Murine autoantibody levels to CRC-specific TAAs – p53 and MST1/STK4 – at day 21 and day 63 can be observed since day 21 and prior to the detection of neoplastic colon lesions by histological staining. (d) Autoantibody levels to p53 of four out of nine randomly selected AOM-/DSS-treated mice together with two out of nine vehicle-treated mice at indicated times to follow their levels according to the progression of the disease (Adapted from Barderas et al. 2013)
Optimization of a Diagnostic Test Based on Circulating Autoantibodies
Despite promising results, very few commercial kits using this technology have reached the market (Lam et al. 2011). Circulating autoantibodies need to be validated in a clinical context using independent and large patient’s cohorts to verify their usefulness in terms of reproducibility, detection limit, and predictive value. Biomarker development consists of several phases including preclinical studies, clinical assay development and optimization, retrospective studies of stored specimens, prospective screening studies, and multicentered randomized clinical trials (Pepe et al. 2001). In addition, as mentioned above, other sources of optimization to improve the diagnostic ability of an autoantibody panel would include (i) the expression system to produce the TAA, (ii) the platform used for testing, and (iii) the number of TAAs to be included in the diagnostic test (Fig. 4).

Road map to identify and develop CRC diagnostic and prognostic tests. Steps considered as needed or mandatory are highlighted
Optimization of the TAA Expression and Purification
Peptides and proteins produced for the validation of the candidate biomarkers represent an important source of variability. Validation of the TAAs should be performed with purified proteins of the highest purity and quality, containing PTMs similar to the actual tumor target (Casal and Barderas 2010). Impurities, particularly after expression in Escherichia coli, might result in false-positive detection of cancer-specific autoantibodies due to the presence of bacteria in the intestinal microbiota. However, minor contaminants from insect cells have also been reported to be reactive to immunoglobulins from cancer patients, which might affect the detection of cancer-specific autoantibodies (Schmetzer et al. 2005). The presence of different tags (GST, MBP, or 6xHis) fused to the selected TAAs in the N- or C-terminal end of the protein should also be tested since it could affect the binding of the autoantibodies to the protein. Moreover, optimization should include producing each antigen in E. coli, insect, and mammalian cells for testing autoantibody recognition. However, no comparative studies have been yet performed.
Optimization of the Platform for TAA Testing
The choice of the platform and/or the solid support used to probe the protein can also affect the final result. It has been recently published that autoantigens perform differently if the protein is coated (ELISA), immobilized on membranes (WB or protein microarrays), or highly unfolded as is the case for the hEx1 proteins (Murphy et al. 2012a). As each cancer patient can develop a different immune response to protein regions presented as discontinuous, conformational, or linear epitopes, diagnostic tests should contain multiple TAAs for a full coverage of cancer patients (Lam et al. 2011; Murphy et al. 2012a). The use of multiplexed immunoassays (magnetic beads or multiplexed ELISA) (Ling et al. 2007) or low-density protein microarrays (Liu et al. 2009) could be useful for the development of diagnostic tests.
Identification of the Minimum Number of TAAs to Be Included in a CRC Predictor Panel: Correlation Analyses
In order to overcome the heterogeneous response among patients, combinations of biomarkers are needed. Disease heterogeneity implies the use of biomarker panels. To avoid the overlap in reactivity with sera and to select the minimum number of TAAs for CRC diagnosis, it is convenient to calculate the redundancy of the markers and discern whether the information provided by each TAA is redundant or not, with the rest of TAAs to correctly classify patients and controls. Low correlation values indicate that reactivity between markers does not show association and they classify different subsets of CRC patients, whereas high correlation indicates that they are recognized by the same set of patients (Barderas et al. 2012). Globally, the final autoantibody panel would benefit if we find a combined classifier with their TAAs showing the lowest correlation to cover the largest number of different CRC patients (Barderas et al. 2012). On the other hand, we could test together those combinations of TAAs identifying the same groups of patients to simplify the diagnostic test (Barderas et al. 2012).
To identify redundancy, CRC-specific TAAs were tested with the same collection of sera, the ELISA values were plotted, and Pearson and Spearman correlation coefficients were performed for all combinations of TAAs (Barderas et al. 2012). An example of this correlation analysis was done for a panel of TAAs identified by protein and phage display protein microarray using 90 sera from CRC patient and healthy individual sera (Table 2; Babel et al. 2009, 2011; Barderas et al. 2012). Here, biomarker candidates discovered through the same platform showed correlation values higher than 0.4, indicating the necessity to combine TAAs with low correlation in the diagnostic TAA panel (Barderas et al. 2012). In summary, optimization steps are required for specificity and sensitivity improvements to get a reliable CRC diagnostic test (Fig. 4).
Potential Applications to Prognosis, Other Diseases, or Conditions
Prognostic markers indicate a survival outcome of patients. Although multiple reports have claimed that autoantibodies are useful not only for diagnosis but also for prognosis, survival, and recurrence, there are almost no reports showing correlation of circulating autoantibodies and prognosis of colorectal cancer. Babel et al. showed a protein signature able to differentiate between CRC patients suffering metastasis to liver and metastasis to lung (Babel et al. 2009). In addition, we also found, based on results with 96 colorectal cancer serum samples, that higher levels of circulating autoantibodies to MST1/STK4 were associated to a better prognosis (personal observations). In two other studies, sera from 55 patients with colon adenocarcinoma, 26 patients with benign surgical diseases, and 40 healthy individuals by ELISA were examined for autoantibodies to dsDNA and tropomyosin and its association to prognosis (Syrigos et al. 1999, 2000). The authors found that the presence of autoantibodies to both proteins was associated to a better outcome of the disease. Finally, in another recent study, an association of high levels of autoantibodies to the aberrant glycopeptide MUC4TR5 with a risk of death in CRC patients has been observed (Pedersen et al. 2013).
In any case, circulating autoantibodies have shown a strong prognostic power in other types of cancer. Using 120 cancer samples from benign ductal carcinoma in situ or invasive breast cancer, a signature of 5 TAAs (RBP-J kappa, HMGN1, PSRC, CIRBP, and ECHDC1) was able to discriminate patients with an AUC = 0.794. Higher autoantibody titers of RBP-J kappa were found in patients with a higher histotype grade and higher recurrence-free survival for those patients who were positive for the five-antibody signature (Mange et al. 2012). Similar results were found in pancreatic cancer (Bracci et al. 2012) and glioblastoma multiforme (Pallasch et al. 2005), where survival rates were higher in patients with higher levels of autoantibodies to CTDSP1 and NR2E3 and GLEA2 and PHF3 autoantigens, respectively. Finally, in prostate cancer patients, higher levels of autoantibodies to TARDBP, TLN1, PARK7, CALD1, and PISP1 were able to discriminate benign prostate hyperplasia patients from prostate cancer patients (O’Rourke et al. 2012).
More reports are necessary to clarify this question, since other reports claim that p53 autoantibodies negatively correlate with the survival of cancer patients (Lubin et al. 1995; Tang et al. 2001). Then, the correlation to overall patient survival of autoantibody levels against a specific TAA might depend on its nature and function in the tumoral tissue.
In addition, circulating biomarkers might be used to monitor the disease and detect recurrences by quantifying changes in autoantibody levels during treatment. However, there are few reports showing their usefulness to detect recurrences. In CRC, a decrease in the immune response to p53 in 78 % of p53-seropositive patients after surgery has been reported (Ochiai et al. 2012). In addition, a rapid and durable loss of p53 antibodies has also been observed in breast cancer after resection of tumors and treatment with neoadjuvant chemotherapy (Anderson et al. 2008). In ovarian cancer, Tainsky lab detected recurrences at 9.07 months prior to clinical recurrence (Chatterjee et al. 2012). They demonstrated that 3 out of 56 antigens displayed in phages and printed in microarrays correctly classified recurrent and nonrecurrent ovarian cancer patients (Chatterjee et al. 2012).
Perspectives and Concluding Remarks
The detection of CRC circulating autoantibodies holds great promise for CRC diagnosis. The combination of different TAAs has allowed the obtainment of assays with very high specificity and sensitivity. Their low invasiveness would facilitate their inclusion in a routine health check in contrast to other invasive screening methods, like colonoscopy. Moreover, the use for early clinical diagnosis would significantly improve the overall survival of CRC patients and save considerable amounts of money to the National Health Systems. However, there are some questions that still remain open. The first and most critical, which are the factors that induce this autoantibody response to cancer proteins? Second, which is the actual TAA repertoire generated for each tumor? The dispersion of identified TAAs among the different technologies and the little reproducibility among laboratories complicate the definition of clear guidelines for TAA acceptance. In this regard, the demonstration of a similar response in mouse cancer models supposes a strong support and provides a good tool for the testing of putative TAAs. Third and finally, validation is a fundamental issue that still continues to be neglected in many reports. A final support for the technology will come only from final clinical assays and application. In this regard, an initial product has been recently commercialized for lung cancer (Lam et al. 2011).
In addition to diagnostic power, the screening of the autoantibody response of CRC patients might be useful for prognostic or therapy monitoring purposes, but further studies are required (Fig. 5). The identification of new therapeutic targets for intervention is also a main strength of this strategy (Barderas et al. 2010; Dudas et al. 2010; Pelaez-Garcia et al. 2013).

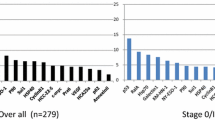
Medical needs covered by CRC autoantibodies according to the stage of the disease
Summary Points
-
Colorectal cancer autoantibodies and their target proteins constitute a very promising alternative as a diagnostic test for the massive screening of populations to identify colon cancer-carrier individuals.
-
A description of the methodologies used to identify circulating autoantibodies is given.
-
Autoantibodies directed against tumor-associated antigens (TAAs) emerge early in the disease and follow its progression.
-
Multiple reports have described a wide number of TAAs due to the heterogeneous response in cancer.
-
Colorectal cancer murine models show similar humoral responses to that observed in humans, allowing its study in a more reproducible way and the possibility to compare predictive values of different autoantibodies identified elsewhere.
-
Autoantibodies might be effective for identifying recurrence and monitoring therapy.
-
Autoantibodies and their target proteins are useful for identifying potential therapeutic targets.
Abbreviations
- CEA:
-
Carcinoembryonic antigen
- CRC:
-
Colorectal cancer
- EMT:
-
Epithelial to mesenchymal transition
- HPLC:
-
High-performance liquid chromatography
- MSI:
-
Microsatellite instability
- NAPPA:
-
Nucleic acid programmable protein arrays
- PTMs:
-
Posttranslational modifications
- SEREX:
-
Serological analysis of recombinant cDNA expression libraries
- SERPA:
-
Serological proteome analysis
- TAA:
-
Tumor-associated antigen
References
Anderson KS, LaBaer J. The sentinel within: exploiting the immune system for cancer biomarkers. J Proteome Res. 2005;4:1123–33.
Anderson KS, Ramachandran N, Wong J, et al. Application of protein microarrays for multiplexed detection of antibodies to tumor antigens in breast cancer. J Proteome Res. 2008;7:1490–9.
Babel I, Barderas R, Diaz-Uriarte R, et al. Identification of tumor-associated autoantigens for the diagnosis of colorectal cancer in serum using high density protein microarrays. Mol Cell Proteomics. 2009;8:2382–95.
Babel I, Barderas R, Diaz-Uriarte R, et al. Identification of MST1/STK4 and SULF1 proteins as autoantibody targets for the diagnosis of colorectal cancer by using phage microarrays. Mol Cell Proteomics. 2011;10:M110 001784.
Barderas R, Babel I, Casal JI. Colorectal cancer proteomics, molecular characterization and biomarker discovery. Proteomics Clin Appl. 2010;4:159–78.
Barderas R, Babel I, Diaz-Uriarte R, et al. An optimized predictor panel for colorectal cancer diagnosis based on the combination of tumor-associated antigens obtained from protein and phage microarrays. J Proteomics. 2012;75:4647–55.
Barderas R, Villar-Vazquez R, Fernandez-Acenero MJ, et al. Sporadic colon cancer murine models demonstrate the value of autoantibody detection for preclinical cancer diagnosis. Sci Rep. 2013;3:2938.
Bracci PM, Zhou M, Young S, et al. Serum autoantibodies to pancreatic cancer antigens as biomarkers of pancreatic cancer in a San Francisco Bay Area case–control study. Cancer. 2012;118:5384–94.
Bussow K, Cahill D, Nietfeld W, et al. A method for global protein expression and antibody screening on high-density filters of an arrayed cDNA library. Nucleic Acids Res. 1998;26:5007–8.
Bussow K, Nordhoff E, Lubbert C, et al. A human cDNA library for high-throughput protein expression screening. Genomics. 2000;65:1–8.
Carey TE, Takahashi T, Resnick LA, et al. Cell surface antigens of human malignant melanoma: mixed hemadsorption assays for humoral immunity to cultured autologous melanoma cells. Proc Natl Acad Sci U S A. 1976;73:3278–82.
Casal JI, Barderas R. Identification of cancer autoantigens in serum: toward diagnostic/prognostic testing? Mol Diagn Ther. 2010;14:149–54.
Chang WJ, Wu LL, Cao FA, et al. Development of autoantibody signatures as biomarkers for early detection of colorectal carcinoma. Clin Cancer Res. 2011;17:5715–24.
Chatterjee M, Mohapatra S, Ionan A, et al. Diagnostic markers of ovarian cancer by high-throughput antigen cloning and detection on arrays. Cancer Res. 2006;66:1181–90.
Chatterjee M, Dyson G, Levin NK, et al. Tumor autoantibodies as biomarkers for predicting ovarian cancer recurrence. Cancer Biomark. 2012;11:59–73.
Chen Y, Lin P, Qiu SM, et al. Autoantibodies to Ca2+ binding protein Calnuc is a potential marker in colon cancer detection. Int J Oncol. 2007;30:1137–44.
Chen JS, Kuo YB, Chou YP, et al. Detection of autoantibodies against Rabphilin-3A-like protein as a potential biomarker in patient’s sera of colorectal cancer. Clin Chim Acta. 2011;412:1417–22.
Dippold WG, Lloyd KO, Li LT, et al. Cell surface antigens of human malignant melanoma: definition of six antigenic systems with mouse monoclonal antibodies. Proc Natl Acad Sci U S A. 1980;77:6114–8.
Dudas SP, Chatterjee M, Tainsky MA. Usage of cancer associated autoantibodies in the detection of disease. Cancer Biomark. 2010;6:257–70.
Duffy MJ, van Dalen A, Haglund C, et al. Tumour markers in colorectal cancer: European Group on Tumour Markers (EGTM) guidelines for clinical use. Eur J Cancer. 2007;43:1348–60.
Fan CW, Chan CC, Chen KT, et al. Identification of SEC61 beta and its autoantibody as biomarkers for colorectal cancer. Clin Chim Acta. 2011;412:887–93.
Festa F, Rollins SM, Vattem K, et al. Robust microarray production of freshly expressed proteins in a human milieu. Proteomics Clin Appl. 2013;7:372–7.
He YJ, Wu YZ, Mou ZR, et al. Proteomics-based identification of HSP60 as a tumor-associated antigen in colorectal cancer. Proteomics Clin Appl. 2007;1:336–42.
Hudson ME, Pozdnyakova I, Haines K, et al. Identification of differentially expressed proteins in ovarian cancer using high-density protein microarrays. Proc Natl Acad Sci U S A. 2007;104:17494–9.
Ishikawa T, Fujita T, Suzuki Y, et al. Tumor-specific immunological recognition of frameshift-mutated peptides in colon cancer with microsatellite instability. Cancer Res. 2003;63:5564–72.
Jiang BH, Ren TT, Dong B, et al. Peptide mimic isolated by autoantibody reveals human arrest defective 1 overexpression is associated with poor prognosis for colon cancer patients. Am J Pathol. 2010;177:1095–103.
Kijanka G, Hector S, Kay EW, et al. Human IgG antibody profiles differentiate between symptomatic patients with and without colorectal cancer. Gut. 2010;59:69–78.
Klade CS, Voss T, Krystek E, et al. Identification of tumor antigens in renal cell carcinoma by serological proteome analysis. Proteomics. 2001;1:890–8.
Kocer B, McKolanis J, Soran A. Humoral immune response to MUC5AC in patients with colorectal polyps and colorectal carcinoma. BMC Gastroenterol. 2006;6.
Kronick MN. Creation of the whole human genome microarray. Expert Rev Proteomics. 2004;1:19–28.
Kung LA, Snyder M. Proteome chips for whole-organism assays. Nat Rev Mol Cell Biol. 2006;7:617–22.
Lam S, Boyle P, Healey GF, et al. Early CDT-lung: an immunobiomarker test as an aid to early detection of lung cancer. Cancer Prev Res (Phila). 2011;4:1126–34.
Levin B, Brooks D, Smith RA, et al. Emerging technologies in screening for colorectal cancer: CT colonography, immunochemical fecal occult blood tests, and stool screening using molecular markers. CA Cancer J Clin. 2003;53:44–55.
Line A, Slucka Z, Stengrevics A, et al. Characterisation of tumour-associated antigens in colon cancer. Cancer Immunol Immunother. 2002;51:574–82.
Ling MM, Ricks C, Lea P. Multiplexing molecular diagnostics and immunoassays using emerging microarray technologies. Expert Rev Mol Diagn. 2007;7:87–98.
Liotta LA, Espina V, Mehta AI, et al. Protein microarrays: meeting analytical challenges for clinical applications. Cancer Cell. 2003;3:317–25.
Liu W, Wang P, Li Z, et al. Evaluation of tumour-associated antigen (TAA) miniarray in immunodiagnosis of colon cancer. Scand J Immunol. 2009;69:57–63.
Liu W, Li Z, Xu W, et al. Humoral autoimmune response to IGF2 mRNA-binding protein (IMP2/p62) and its tissue-specific expression in colon cancer. Scand J Immunol. 2013;77:255–60.
Locker GY, Hamilton S, Harris J, et al. ASCO 2006 update of recommendations for the use of tumor markers in gastrointestinal cancer. J Clin Oncol. 2006;24:5313–27.
Lubin R, Schlichtholz B, Teillaud JL, et al. p53 antibodies in patients with various types of cancer: assay, identification, and characterization. Clin Cancer Res. 1995;1:1463–9.
Mange A, Lacombe J, Bascoul-Mollevi C, et al. Serum autoantibody signature of ductal carcinoma in situ progression to invasive breast cancer. Clin Cancer Res. 2012;18:1992–2000.
Mintz PJ, Kim J, Do KA, et al. Fingerprinting the circulating repertoire of antibodies from cancer patients. Nat Biotechnol. 2003;21:57–63.
Murphy MA, O’Connell DJ, O’Kane SL, et al. Epitope presentation is an important determinant of the utility of antigens identified from protein arrays in the development of autoantibody diagnostic assays. J Proteomics. 2012a;75:4668–75.
Murphy MA, O’Leary JJ, Cahill DJ. Assessment of the humoral immune response to cancer. J Proteomics. 2012b;75:4573–9.
Nam MJ, Madoz-Gurpide J, Wang H, et al. Molecular profiling of the immune response in colon cancer using protein microarrays: occurrence of autoantibodies to ubiquitin C-terminal hydrolase L3. Proteomics. 2003;3:2108–15.
O’Rourke DJ, DiJohnson DA, Caiazzo RJ, et al. Autoantibody signatures as biomarkers to distinguish prostate cancer from benign prostatic hyperplasia in patients with increased serum prostate specific antigen. Clin Chim Acta. 2012;413:561–7.
Ochiai H, Ohishi T, Osumi K, et al. Reevaluation of serum p53 antibody as a tumor marker in colorectal cancer patients. Surg Today. 2012;42:164–8.
Oishi Y, Yunomura S, Kawahashi Y, et al. Escherichia coli proteome chips for detecting protein-protein interactions. Proteomics. 2006;6:6433–6.
Orenes-Pinero E, Barderas R, Rico D, et al. Serum and tissue profiling in bladder cancer combining protein and tissue arrays. J Proteome Res. 2010;9:164–73.
Pallasch CP, Struss AK, Munnia A, et al. Autoantibodies against GLEA2 and PHF3 in glioblastoma: tumor-associated autoantibodies correlated with prolonged survival. Int J Cancer. 2005;117:456–9.
Pedersen JW, Blixt O, Bennett EP, et al. Seromic profiling of colorectal cancer patients with novel glycopeptide microarray. Int J Cancer. 2011;128:1860–71.
Pedersen JW, Gentry-Maharaj A, Fourkala EO, et al. Early detection of cancer in the general population: a blinded case–control study of p53 autoantibodies in colorectal cancer. Br J Cancer. 2013;108:107–14.
Pedersen JW, Gentry-Maharaj A, Nostdal A, et al. Cancer associated auto-antibodies to MUC1 and MUC4 – A blinded case control study of colorectal cancer in UK Collaborative Trial of Ovarian Cancer Screening (UKCTOCS). Int J Cancer. 2014;134:2180–88.
Pelaez-Garcia A, Barderas R, Torres S, et al. FGFR4 role in epithelial-mesenchymal transition and its therapeutic value in colorectal cancer. PLoS One. 2013;8:e63695.
Pepe MS, Etzioni R, Feng Z, et al. Phases of biomarker development for early detection of cancer. J Natl Cancer Inst. 2001;93:1054–61.
Qiu J, Madoz-Gurpide J, Misek DE, et al. Development of natural protein microarrays for diagnosing cancer based on an antibody response to tumor antigens. J Proteome Res. 2004;3:261–7.
Ramachandran N, Hainsworth E, Bhullar B, et al. Self-assembling protein microarrays. Science. 2004;305:86–90.
Ran YL, Hu H, Zhou Z, et al. Profiling tumor-associated autoantibodies for the detection of colon cancer. Clin Cancer Res. 2008;14:2696–700.
Reipert BM, Tanneberger S, Pannetta A, et al. Increase in autoantibodies against Fas (CD95) during carcinogenesis in the human colon: a hope for the immunoprevention of cancer? Cancer Immunol Immunother. 2005;54:1038–42.
Sahin U, Tureci O, Schmitt H, et al. Human neoplasms elicit multiple specific immune responses in the autologous host. Proc Natl Acad Sci U S A. 1995;92:11810–3.
Scanlan MJ, Chen YT, Williamson B, et al. Characterization of human colon cancer antigens recognized by autologous antibodies. Int J Cancer. 1998;76:652–8.
Scanlan MJ, Welt S, Gordon CM, et al. Cancer-related serological recognition of human colon cancer: identification of potential diagnostic and immunotherapeutic targets. Cancer Res. 2002;62:4041–7.
Schmetzer O, Moldenhauer G, Riesenberg R, et al. Quality of recombinant protein determines the amount of autoreactivity detected against the tumor-associated epithelial cell adhesion molecule antigen: low frequency of antibodies against the natural protein. J Immunol. 2005;174:942–52.
Shebzukhov YV, Koroleva EP, Khlgatian SV, et al. Antibody response to a non-conserved C-terminal part of human histone deacetylase 3 in colon cancer patients. Int J Cancer. 2005;117:800–6.
Song MH, Ha JC, Lee SM, et al. Identification of BCP-20 (FBXO39) as a cancer/testis antigen from colon cancer patients by SEREX. Biochem Biophys Res Commun. 2011;408:195–201.
Soussi T. p53 antibodies in the sera of patients with various types of cancer: a review. Cancer Res. 2000;60:1777–88.
Syrigos KN, Charalampopoulos A, Pliarchopoulou K, et al. Prognostic significance of autoantibodies against tropomyosin in patients with colorectal adenocarcinoma. Hybridoma. 1999;18:543–6.
Syrigos KN, Charalambopoulos A, Pliarchopoulou K, et al. The prognostic significance of autoantibodies against dsDNA in patients with colorectal adenocarcinoma. Anticancer Res. 2000;20:4351–3.
Takulapalli BR, Qiu J, Magee DM, et al. High density diffusion-free nanowell arrays. J Proteome Res. 2012;11:4382–91.
Tang RP, Ko MC, Wang JY, et al. Humoral response to P53 in human colorectal tumors: a prospective study of 1,209 patients. Int J Cancer. 2001;94:859–63.
Villanueva J, Shaffer DR, Philip J, et al. Differential exoprotease activities confer tumor-specific serum peptidome patterns. J Clin Invest. 2006;116:271–84.
Wandall HH, Blixt O, Tarp MA, et al. Cancer biomarkers defined by autoantibody signatures to aberrant O-glycopeptide epitopes. Cancer Res. 2010;70:1306–13.
Wang X, Yu J, Sreekumar A, et al. Autoantibody signatures in prostate cancer. N Engl J Med. 2005;353:1224–35.
Zhu H, Snyder M. Protein arrays and microarrays. Curr Opin Chem Biol. 2001;5:40–5.
Zhu H, Bilgin M, Bangham R, et al. Global analysis of protein activities using proteome chips. Science. 2001;293:2101–5.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer Science+Business Media Dordrecht
About this entry
Cite this entry
Barderas, R., Villar-Vázquez, R., Casal, J.I. (2015). Colorectal Cancer Circulating Biomarkers. In: Preedy, V., Patel, V. (eds) Biomarkers in Cancer. Biomarkers in Disease: Methods, Discoveries and Applications. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-7681-4_29
Download citation
DOI: https://doi.org/10.1007/978-94-007-7681-4_29
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-7680-7
Online ISBN: 978-94-007-7681-4
eBook Packages: Biomedical and Life SciencesReference Module Biomedical and Life Sciences