
Abstract
Plants are often simultaneously exposed to multiple biotic and abiotic stresses resulting in substantial yield loss. Moreover, increase in the frequency of climate extremes is likely to influence the distribution, establishment and epidemiology of plant diseases. Emerging evidences suggest the changing scenario of diseases in chickpea, a grain legume largely grown in rain-fed environments. In this chapter, we have focused on the major and emerging soil-borne diseases in chickpea that are largely influenced by differential temperature and soil moisture stress. Changes in the disease spectrum in chickpea for the past one decade were monitored through extensive surveys. Analysis of disease and weather data indicated shift in the occurrence and distribution of chickpea diseases as well as emergence of new diseases. Dry root rot (Rhizoctonia bataticola) is becoming more intense in tropical humid areas under high temperature and soil moisture stress. Contrary to this, sporadic occurrence of collar rot (Sclerotium rolfsii) has been noticed under high soil moisture levels. Host resistance influenced by soil moisture levels and rise in temperature have also been discussed. Extensive research is required in this domain to develop adaptation and mitigation strategies for sustained food security. Breeding being an essential part of crop improvement needs to keep pace with these emerging diseases.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
4.1 Introduction
Chickpea (Cicer arietinum L.) is one of the essential semiarid tropical legume crops and is either grown in the post-rainy season on stored soil moisture (south Asia and spring-sown Mediterranean) or as a Mediterranean winter crop. In both of these instances, the crop is exposed to terminal drought accompanied with high temperatures. Under the changing scenario of climate, more erratic rainfall patterns and spells of temperature extremes will consequently affect the crop productivity (Graham and Vance 2003). A steady increase in temperature, decrease/increase in relative humidity and moisture stress will not only affect the crop per se but will also change the relative activity and abundance of diseases, natural enemies, and their interaction with the host plants. As a consequence of it, shift in the chickpea diseases have been seen in the major growing regions worldwide.
Chickpea is a short-duration, self-pollinated, diploid (2n = 2x = 16) legume with genome size of 738 Mb (Varshney etal. 2013). It is cultivated in different parts of the world mainly in the Mediterranean, South Asia, North Africa, Middle East and North and Central America. It is a rich and cheap source of vegetarian protein, vitamins and minerals (Jukanti etal. 2012), making it nutritionally more valuable. These valuable aspects of chickpea caused an increase in its global cultivation and the overall production reached 10.0 million metric tons (mt) from 6.6 mt (http://www.cgiar.org/ourresearch/crop-factsheets/chickpea, as on 23rd April 2013) during the last 30 years. South Asia accounts for more than 75% of the total area under chickpea cultivation, and India is the world leader in chickpea production with 7.5 mt (FAOSTAT 2010), followed by Pakistan and Turkey.
Both biotic and abiotic stresses cause significant yield losses (corresponding to 11.2 mt) in chickpea (Ryan 1997). Among the abiotic stresses, drought causes a 40–50% reduction in yield globally (Graham and Vance 2003) and is emerging as a major barrier to its wider cultivation on the drought prone semiarid tropic (SAT) region. With the increasing drought, fungal diseases like dry root rot that thrive in the drought conditions are emerging as major threat to its production. Temperature may have important repercussions on the effectiveness of host plant resistance. Theoretically, following three types of abiotic–biotic interactions can be expected in chickpea:
-
A direct effect on pathogens
-
Indirect effect on pathogens through other community interactions
-
Interaction effects through host physiology, i.e. multiple stress concept
This chapter deals with the soil-borne diseases and pathogens of chickpea being largely impacted by temperature and soil moisture stresses within the production situations. We have highlighted sequential occurrence of chickpea diseases under different weather scenarios in SAT regions, followed by short narratives of emerging diseases and few evidences/data that support these results and finally conclusions and presumption for the future.
4.2 Sequential Occurrence of Soil-Borne Diseases of Chickpea in SAT Environments
The spatial and temporal succession of soil-borne diseases such as Fusarium wilt (Fusarium oxysporum f. sp. ciceris), dry root rot (Rhizoctonia bataticola), collar rot (Sclerotium rolfsii), black root rot (Fusarium solani) and wet root rot (Rhizoctonia solani) of chickpea in SAT is strongly influenced by the prevailing climate variables. An analysis of the weather data and diseases pattern in chickpea in the past one decade indicated a shift in the disease pattern (Sharma 2012). The production of chickpea is largely constrained by Fusarium wilt in crop season with no drastic variations in weather, and, therefore, all the breeding efforts in past were towards developing wilt resistant cultivars. As a result, several wilt resistant cultivars have been deployed and released worldwide (Gowda and Gaur 2004). However, in present scenario frequency of the occurrence of diseases like collar rot and dry root rot has increased due to change in temperature and rainfall. Past and present scenarios of chickpea diseases as influenced by environment are represented in Figs. 4.1 and 4.2. Figure 4.1 signifies the predominance of Fusarium wilt throughout the crop season provided the weather conditions have no drastic variations, and the Fig. 4.2 illustrates the predominance of collar rot at the seedling stage and dry root rot at the time of flowering and podding predisposed by high moisture and drought, respectively. The later scenario has become more common in SAT, thereby demanding the research focus on emerging diseases with respect to their epidemiology, pathogen biology and host × pathogen × environment interactions. In the following section, two examples of chickpea diseases influenced by drought and high soil moisture have been discussed.

Past scenario of chickpea diseases with respect to past weather (*indicates less incidence; ***indicate more incidence of the disease)

Present scenario of chickpea diseases with respect to changed weather (*indicates less incidence; ***indicate more incidence)
4.2.1 High Temperature and Soil Moisture Stress Predisposing Factor for Dry Root Rot
Dry root rot caused by Rhizoctonia bataticola (Macrophomina phaseolina) is an important component of the disease complex that causes root rots and seedling blight in many grain legumes when they are weakened by other stress factors (Hwang etal. 2003). R. bataticola is a soil inhabiting organism and most commonly infects chickpea at post-reproductive stage in dry and warm regions (Sharma and Pande 2013). The disease is of increasing importance in chickpea and has the potential to cause devastation in susceptible cultivars, particularly in the conditions of high temperature and soil moisture stress. Savary etal. (2011) described dry root rot as an acute-emerging disease that occurs irregularly, both temporally and spatially, and may cause massive disruptions in system performances and whose range is expanding to new areas.
The disease has been reported from most chickpea growing areas in India and other countries like Iran, the USA and several countries in Asia and Africa (see details in Sharma etal. 2015) but has become a major threat to chickpea production in recent years due to longer drought spells at the time of flowering and podding (Sharma and Pande 2013). Ghosh etal. (2013) conducted a survey during 2010–2013 in India and indicated widespread and increased incidence of dry root rot in the central and southern states of India. The disease was found irrespective of soil types, cropping system and cultivars used and incidence ranged from 5 to 50% or more in badly infected soils. This noticeable widespread geographic distribution of dry root rot probably makes it a significant disease in chickpea. R. bataticola is more virulent under high temperatures (32 °C) and as a result cause severe damage on chickpeas in the warmer Salinas Valley in California (Raabe 1985).
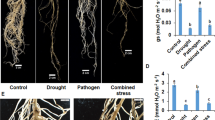
Sharma and Pande (2013) demonstrated the relationship between temperature and soil moisture stress on the development of dry root rot. They conducted series of experiments in controlled environment conditions to understand the effect of temperature and soil moisture alone or in combination on infection, colonisation and development of dry root rot. They concluded that a combination of high temperature (≥30 °C) and soil moisture content (≤60%) are positively correlated with dry root rot incidence/severity in chickpea (Fig. 4.3). Singh and Sharma (2002) also reported that soil moisture deficit favours the severe disease development on pulse crops. Under hot and dry conditions, many economically important crops are predisposed to R. bataticola infection such as soybean (Pearson etal. 1984), sunflower (Nawaz Khan 2007), etc. Sclerotial development was affected by soil moisture levels; more sclerotia were found under relatively low water potentials favouring the disease development (Olaya and Abawi 1996); however the effect was adverse under high water potential.

Effect of temperature and soil moisture (a) on dry root rot severity (1–9 scale) (b) and on incidence of dry root rot (c) correlation between soil moisture content, soil temperature and dry root rot incidence
4.2.2 High Soil Moisture Predisposing Factor for Collar Rot
Collar rot (S. rolfsii) is a soil-borne disease, emerging as a potential threat to chickpea production in the tropics, subtropics and other warm temperate regions. The disease in chickpea is favoured by abundant soil moisture, high soil temperature (25–30 °C) and low organic matter in the soil and has a major impact on plant fitness/growth. The disease can cause 55–95% mortality of chickpea seedlings under favourable environmental conditions. Pathogen’s extensive host range (at least 500 species in 100 families) most commonly in legumes, crucifers and cucurbits further makes this disease challenging. Ghosh etal. (2013) reported collar rot as an emerging disease in chickpea particularly in years when there is heavy rainfall at the seedling stage, leading to high moisture conducive for the disease development.
S. rolfsii grows, survives and attacks plants at or near the soil line. The fungus can overwinter as mycelium in infected tissues or plant debris. Sclerotia serve as the principal overwintering structure and primary inoculum for disease persistence near the soil surface. Sclerotia may exist free in soil or in association with plant debris. Those buried deep in the soil may survive for a year or less, whereas those at surface remain viable and may germinate in response to volatile compounds released from decomposing plant material. Before the pathogen penetrates host tissue, it produces a considerable mass of mycelium on the plant surface, a process which can take 2–10 days. Production of mycelium and the formation of sclerotia rely upon favourable environmental conditions. Hussain etal. (2006) showed that there was a positive correlation between disease severity and inoculum concentrations where seedling mortality increased with an increase in inoculum load. Lack of information about factors that affect the development of collar rot has made control of this disease difficult.
4.3 Host Resistance Influenced by Changes in Temperature and Other Factors
The influence of temperature on expression of resistance or susceptibility to plant pathogens has been shown by various researchers (Ash and Rees 1994; Brake etal. 1995; French and Elder 1996; Ge etal. 1998; Harling etal. 1998; Judelson and Michelmore 1992; Kim and Bockus 2003; Omwega and Roberts 1992; Sydenham etal. 1997). Fusarium wilt (F. oxysporum f. sp. ciceris) in combination with cyst nematode is already on rise in some countries probably due to soil temperature rise (Jimenez-Díaz etal. 1993). Also, the race-specific resistant response of chickpea cultivars to infection by F. oxysporum f. sp. ciceris was reported to be significantly influenced by the increase in temperature (Landa etal. 2006). Artificial inoculation experiments showed that a 3 °C increase, from 24 to 27 °C, in the incubation temperature was sufficient for the reaction of kabuli cv. Ayala and accession PV-1 to race 1A to shift from moderately/highly resistant at constant 24 °C to highly susceptible at 27 °C. A similar but less pronounced effect was found for ‘Ayala’ infected with race 6 (Landa etal. 2006). However, the susceptible reaction of accession JG-62 to races 1A and 6 was not influenced by the temperature increase. This temperature effect has an impact on the use of cultural practices for management of Fusarium wilt of chickpea. High level of resistance of ‘Ayala’ to Fusarium wilt when sown in mid- to late January differed from a moderately susceptible reaction under warmer temperatures when sowing was delayed to late February or early March (Landa etal. 2006) indicating that resistance in this cultivar may be temperature dependent and that warmer temperatures, associated with later sowings, may affect the disease reaction of this cultivar. This study demonstrates the importance of temperature in identifying resistant genotypes and races of the pathogen, as well as choosing sowing dates and using resistant chickpea genotypes for the management of Fusarium wilt in different growing areas.
Temperature can also influence the plant–rhizobacteria interactions related to biocontrol potential. For instance, in chickpea, seed and soil treatment with Pseudomonas fluorescens RGAF 19, P. fluorescens RG 26, Bacillus megaterium RGAF 51 and Paenibacillus macerans RGAF 101 can suppress Fusarium wilt, but the extent of disease suppression by these rhizobacteria is modulated by soil temperature. Pseudomonas fluorescens isolates significantly increased chickpea shoot dry weight at 20 °C and root dry weight at 25 and 30 °C (Landa etal. 2004). All bacterial isolates colonised the chickpea rhizosphere and internal stem tissues at 20, 25 and 30 °C, and there was a positive linear trend between bacterial population size in the rhizosphere and temperature increase. The maximum inhibition of mycelial growth and conidial germination of Fusarium oxysporum f.sp. ciceris race 5 in vitro occurred at a temperature range optimal for bacterial growth and production of inhibitory metabolites. These results demonstrate the need to understand the effects of environmental factors on the biological activities of introduced rhizobacteria of significant importance for plant disease suppression.
4.4 Effect of Drought and Moisture on Plant–Pathogen Interactions at Biochemical and Molecular Level
Plant responses to different stresses are highly complex and involve changes at the cellular, physiological and transcriptome levels. It has been found that plants respond to multiple stresses differently from how they do to individual stresses, activating a specific programme of gene expression relating to the exact environmental conditions encountered. Rather than being additive, the presence of an abiotic stress can have the effect of reducing or enhancing susceptibility to a biotic pest or pathogen, and vice versa. This interaction between biotic and abiotic stresses is orchestrated by hormone signalling pathways that may induce or antagonise one another. Specificity in multiple stress responses is further controlled by a range of molecular mechanisms that act together in a complex regulatory network (Atkinson and Urwin 2012).
The studies on plant responses to abiotic stresses particularly heat and drought at the biochemical and molecular level have advanced considerably in recent years and have shown that abiotic stresses impact responses to pathogens in several crops. The impact of concurrent drought stress and pathogen infection on plants has been recently documented by Pandey etal. (2014). Although no such studies have been reported on pathogens infecting chickpea so far; however it was found in preliminary studies on Rhizoctonia bataticola-chickpea pathosystem that anti-oxidant enzymes like PAL, PPO, POD and phenol increased under moisture stress as compared to high moisture levels in sick soils (Sharma etal. unpublished data).
Genes and signalling pathways involved in resistance to pathogens have been unravelled in some of the model plants (Dangl and Jones 2001, Wan etal. 2002). In Arabidopsis, the Early Responsive to Dehydration 15 gene (ERD15) is rapidly induced in response to drought and pathogen infection (Kariola etal. 2006). The overexpression and silencing of the ERD15 gene not only affected abiotic stress tolerance but also disease resistance. Drought also activates the ABA-responsive signalling pathway and other response to biotic stresses. Changes in endogenous ABA levels affect SA-, JA- and ET-related defence responses (Kariola etal. 2006; Asselbergh etal. 2008; Zavala etal. 2009). Incompatible interaction of Arabidopsis and Pseudomonas syringae is prevented by exogenous ABA treatment (Mohr and Cahill 2007). Several such examples where drought can dramatically affect plant defence responses against pathogens has been reported by Eastburn etal. (2010). Increasing temperatures from 22 to 28 °C reduced the effectiveness of both basal and R gene-mediated resistance in A. thaliana when challenged with virulent and avirulent strains of P. syringae pv. tomato, respectively (Wang etal. 2009). Increased level of symptoms were found to be the result of changes in the defence responses associated with the host–pathogen interaction, rather than just an increase in pathogen growth at the higher temperature. These studies indicate the need to understand resistance mechanism during interplay between biotic and abiotic stresses in chickpea.
4.5 Conclusions
Changes in climate, with its multiple effects on ecosystems, are likely to change the interactions between an infectious propagule, a susceptible host and favourable environmental conditions leading to the development of new epidemics. The lack of long-term data is hampering the ability to document the certainty changes in disease profiles. For instance in chickpea, surveys and recent investigations clearly indicate the emergence of new diseases like dry root rot and collar rot which has got a direct relationship with temperature and soil moisture. Further, the dynamics affecting host–pathogen interactions is leading to the selection of new pathotypes or pathogens. The changes in temperature and moisture have also shown to affect the disease reaction of cultivars by changing their resistance/susceptible reactions. Therefore, there is a need to address host x pathogen interactions in the light of multiple stress factors. Breeding needs to keep pace with these emerging diseases as it is an essential part of crop improvement. Increases in yield per unit of area will continue to depend largely on more efficient control of stresses (biotic) along with increase in yield potential. Integrated crop management is, therefore, the platform for sustainable agriculture, and extensive research is required in this domain to develop adaptation and mitigation strategies for sustained food security.
References
Ash GJ, Rees RG (1994) Effect of post-inoculation temperature and light intensity on expression of resistance to stripe rust in some Australian wheat cultivars. Aust J Agric Res 45:1379–1386
Asselbergh B, De Vleesschauwer D, Hofte M (2008) Global switches and fine-tuning-ABA modulates plant pathogen defense. Mol Plant–Microbe Interact 21:709–719
Atkinson NJ, Urwin PE (2012) The interaction of plant biotic and abiotic stresses: from genes to the field. J Exp Bot 63:3523–3543
Brake VM, Pegg KG, Irwin JAG, Chaseling J (1995) The influence of temperature, inoculum level and race of Fusarium oxysporum f. sp. cubense on the disease reaction of Banana cv. Cavendish. Aust J Agric Res 46:673–685
Dangl JL, Jones JDG (2001) Plant pathogens and integrated defence responses to infection. Nature 411:826–833
Eastburn DM, DeGennaro MM, Delucia EH, Dermody O, McElrone AJ (2010) Elevated atmospheric carbon dioxide and ozone alter soybean diseases at SoyFACE. Glob Chang Biol 16:320–330
FAOSTAT data (2010) http://faostat.fao.org/site/339/default. 2010;aspx. Accessed 12 Nov 2012
French CJ, Elder M (1996) Movement of tomato mosaic virus strains in Gomphrena globosa as related to temperature sensitivity and plant resistance. Can J Bot 74:46–50
Ge YF, Johnson JW, Roberts JJ, Rajaram S (1998) Temperature and resistance gene interactions in the expression of resistance to Blumeria graminis f. sp. tritici. Euphytica 99:103–109
Ghosh R, Sharma M, Telangre R, Pande S (2013) Occurrence and distribution of chickpea diseases in central and southern parts of India. Am J Plant Sci 4:940–944
Gowda CLL, Gaur PM (2004) Global scenario of chickpea research-present status and future thrusts. In: Masood A, Singh BB, Kumar S, Dar V (eds) Pulses in new perspective. Proceedings of the national symposium on crop diversification and natural resource management, Kanpur, 2004, pp 1–22
Graham PH, Vance CP (2003) Legumes: importance and constraints to greater use. Plant Physiol 131:872–877
Harling R, Taylor GS, Matthews P, Arthur AE (1998) The effect of temperature on symptom expression and colonization in resistant and susceptible carnation cultivars infected with Fusarium oxysporum f. sp. dianthi. J Phytopathol 121:103–117
Hussain A, Iqbal SM, Ayub N, Zahid MA (2006) Factors affecting development of collar rot disease in chickpea. Pak J Bot 38(1):211–216
Hwang SF, Gossen BD, Chang KF, Turnbull GD, Howard RJ, Blade SF (2003) Etiology, impact and control of rhizoctonia seedling blight and root rot of chickpea on the Canadian prairies. Can J Plant Sci 83:959–967
Jimenez-Díaz RM, Alcala-Jimenez AR, Hervas A, Trapero-Casas JL (1993) Pathogenic variability and hosts resistance in the Fusarium oxysporum f. sp. ciceris/Cicer arietinum pathosystem. In: Proceedings of the 3rd European seminar Fusarium Mycotoxins, taxonomy, pathogenicity and host resistance. Hodowsla Roslin Aklimatyazacja i Nasiennictwo. Plant Breeding and Acclimatization Institute, Radzikov, Poland,,1993., 87e94
Judelson HS, Michelmore RW (1992) Temperature and genotype interactions in the expression of host resistance in lettuce to downy mildew. Physiol Mol Plant Pathol 40:233–245
Jukanti AK, Gaur PM, Gowda CLL, Chibbar RN (2012) Chickpea: nutritional properties and its benefits. Br J Nutr 108:S11–S26
Kariola T, Brader G, Helenius E, Li J, Heino P, Palva ET (2006) Early responsive to dehydration 15, a negative regulator of abscisic acid responses in Arabidopsis. Plant Physiol 142:1559–1573
Kim YK, Bockus WW (2003) Temperature- sensitive reaction of winter wheat cultivar AGSECO 7853 to Stagonospora nodorum. Plant Dis 87:1125–1128
Landa BB, Navas-Cortes JA, Jimenez-Díaz RM (2004) Influence of temperature on plant-rhizobacteria interactions related to biocontrol potential for suppression of Fusarium wilt of chickpea. Plant Pathol 53:341–352
Landa BB, Navas-Cortes JA, Jimenez-Gasco MM, Katan J, Retig B, Jiménez-Díaz RM (2006) Temperature response of chickpea cultivars to races of Fusarium oxysporum f. sp. ciceris, the causal agent of Fusarium wilt. Plant Dis 90:365–374
Mohr PG, Cahill DM (2007) Suppression by ABA of salicylic acid and lignin accumulation and the expression of multiple genes, in Arabidopsis infected with Pseudomonas syringae pv. tomato. Funct Integr Genomics 7:181–191
Nawaz KS (2007) Macrophomina phaseolina as causal agent for charcoal rot of sunflower. Mycopath 5:111–118
Olaya G, Abawi GS (1996) Effect of water potential on mycelial growth and on production and germination of sclerotia of Macrophomina phaseolina. Plant Dis 80:1347–1350
Omwega CO, Roberts PA (1992) Inheritance of resistance to Meloidogyne spp. in common bean and the genetic basis of its sensitivity to temperature. Theor Appl Genet 83:720–726
Pandey P, Sinha R, Mysore KS, Senthil-Kumar M (2014) Impact of concurrent drought stress and pathogen infection on plants. In: Mahalingam R (ed) Combined stresses in plants: physiological, molecular, and biochemical aspects. Springer, Cham, pp 203–222
Pearson CAS, Schwenk FW, Crowe FJ (1984) Colonization of soybean roots by Macrophomina phaseolina. Plant Dis 68:1086–1088
Raabe RT (1985) Association of soil particles with seeds and three pathogens of chickpea in California. Indian Phytopathol 69:238–239
Ryan J (1997) A global perspective on pigeon pea and chickpea sustainable production systems: present status and future potential. In: Asthana A, Ali M (eds) Recent advances in pulses research. Indian Society for Pulses Research and Development, Kanpur, pp 1–31
Savary S, Nelson A, Sparks AH, Willocquet L, Duveiller E, Mahuku G, Forbes G, Garrett KA, Hodson D, Padgham J, Pande S, Sharma M, Yuen J, Djurle A (2011) International agricultural research tackling the effects of global and climate changes on plant diseases in the developing world. Plant Dis 95:1204–1216
Sharma M (2012) Climate change, emerging plant diseases and their management: recent developments. In: Global meet of Biologists, Hyderabad, December 26–28, pp 43–45
Sharma M, Pande S (2013) Unravelling effects of temperature and soil moisture stress response on development of dry root rot [Rhizoctonia bataticola (Taub.)] butler in chickpea. Am J Plant Sci 4:584–589
Sharma M, Ghosh R, Pande S (2015) Dry root rot (Rhizoctonia bataticola (Taub.) Butler): an emerging disease of chickpea – where do we stand? Arch Phytopathol Plant Protect 48(13–16):797–812
Singh G, Sharma YR (2002) Fungal diseases of pulses. In: Gupta VK, Paul YS (eds) Diseases of field crops. Indus publishing, New Delhi, pp 155–192
Sydenham GM, McSorley R, Dunn RA (1997) Effects of temperature on resistance in Phaseolus vulgaris genotypes and on development of Meloidogyne species. J Nematol 29:90–103
Varshney RK, Mohan SM, Gaur PM, Gangarao NVPR, Pandey MK, Bohra A, Sawargaonkar SL, Chitikineni A, Kimurto PK, Janila P, Saxena KB, Fikre A, Sharma M, Rathore A, Pratap A, Tripathi S, Datta S, Chaturvedi SK, Mallikarjuna N, Anuradha G, Babbar A, Choudhary AK, Mhase MB, Bharadwaj C, Mannur DM, Harer PN, Guo B, Liang X, Nadarajan N, Gowda CL (2013) Achievements and prospects of genomics-assisted breeding in three legume crops of the semi-arid tropics. Biotechnol Adv 3:1120–1134
Wan J, Dunning M, Bent AF (2002) Probing plant-pathogen interactions and downstream defense signalling using microarrays. Funct Integr Genomics 2:259–273
Wang Y, Bao Z, Zhu Y, Hua J (2009) Analysis of temperature modulation of plant defense against biotrophic microbes. Mol Plant–Microbe Interact 22:498–506
Zavala JA, Casteel CL, Nabity PD, Berenbaum MR, DeLucia EH (2009) Role of cysteine proteinase inhibitors in preference of Japanese beetles (Popillia japonica) for soybean (Glycine max) leaves of different ages and grown under elevated CO2. Oecologia 161:35–41
Acknowledgement
The authors are thankful to the Department of Science and Technology, Climate Change Division for partial funding support provided to conduct various research activities reported in this chapter.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer (India) Pvt. Ltd.
About this chapter
Cite this chapter
Sharma, M., Ghosh, R. (2017). Heat and Soil Moisture Stress Differentially Impact Chickpea Plant Infection with Fungal Pathogens. In: Senthil-Kumar, M. (eds) Plant Tolerance to Individual and Concurrent Stresses. Springer, New Delhi. https://doi.org/10.1007/978-81-322-3706-8_4
Download citation
DOI: https://doi.org/10.1007/978-81-322-3706-8_4
Published:
Publisher Name: Springer, New Delhi
Print ISBN: 978-81-322-3704-4
Online ISBN: 978-81-322-3706-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)