
Abstract
Biofortification to enhance the bioavailability of micronutrients such as iron, zinc, vitamin A, folic acid (vitamin B9), and cyanocobalamin (vitamin B12) in staple food is the necessity of people living under malnourished conditions. Almost half of the world population is suffering from deficiency of iron, zinc, vitamin A, folic acid, etc. due to poor quality of food they are consuming. They have either low access or no access for these components of food. HarvestPlus has been working to enhance the availability of these nutrients through improved varieties of food so as to provide them health protection. Though the level of micronutrients in the staple food is enhanced by conventional breeding or biotechnology, the presence of antinutrients in food causes hurdle in absorption and bioavailability of Fe, Zn, Ca, folates, etc. These antinutrients are phytates, polyphenols and tannins, protease and α-amylase inhibitors, saponins, lectins, and lathyrogens. Various processing techniques are adopted to remove or reduce these antinutrients to enhance bioavailabilty of essential nutrients of food. Processing techniques such as soaking, germination, milling (dehusking), cooking, and autoclaving are effective in removal of antinutrients, especially from food grains.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Biofortification is the breeding of crops to enhance the nutritional value of food. This is done either by conventional breeding or through biotechnology. During the process of biofortification, the nutrients are enhanced by changing the chemical composition of the food by incorporation of gene responsible for biosynthesis of targeted element or nutrient. It differs from ordinary fortification, in which nutrients are added during processing. The rural and poor mass of society gets the advantage of biofortified food as they are not able to afford the commercially fortified food. It is more advantageous especially in developing world for micronutrient deficiencies such as iron, zinc, vitamin A, and folates. As such, biofortification is seen as an upcoming strategy for dealing with deficiencies of micronutrients. Deficiencies of vitamin A, iron, and zinc affect over one-half of the world’s population, especially women and preschool children. The biofortification is feasible without compromising agronomic productivity. The challenge is to get producers and consumers to accept biofortified crops and increase their intake of the target nutrients. The biofortification of staple food crops is a new health approach to control vitamin A, iron, and zinc deficiencies in poor countries.
2 The Need for Biofortification
Biofortification is the development of micronutrient or vitamin-rich staple crops, especially iron, zinc, vitamin A, and folates, using the best breeding practices or modern biotechnology. This approach has multiple advantages because staple foods predominate in the diets of the poor, and the poor people do not have access to other food supplements which provide them all essential nutrients. Nutritionally improved varieties are therefore required to be grown continuously to feed these people. Biofortification provides a feasible means of reaching undernourished populations in relatively remote rural areas. Biofortification and commercial fortification therefore are highly complementary. Acceptance of mineral-packed seeds among farmers is very popular because these trace minerals provide resistance against diseases and other environmental stresses. Biofortification provides direct benefit for better human health and enhances the immune system by controlling the deficiency of essential nutrients. This requires a multidisciplinary research approach and ultimate dissemination of the biofortified seeds.
Deficiencies of various micronutrients such as zinc and iron and vitamins like vitamin A and folic acid are common in the developing world and affect billions of people. These can lead to higher incidence of blindness and a weaker immune system. The poor, particularly the rural poor, tend to subsist on a diet of staple crops such as rice, wheat, and maize, which are low in these micronutrients. Most of these rural people cannot afford enough fruits, vegetables, or meat products that are necessary to obtain healthy levels of these nutrients. As such, increasing the micronutrient levels in staple crops can prevent and reduce the micronutrient deficiencies. In one trial in Mozambique, eating sweet potatoes biofortified with beta-carotene reduced the incidence of vitamin A deficiency in children by 24 % (Van Jaarsveld et al. 2005). Golden Rice is an example of a GM crop developed for its nutritional value. The latest version of Golden Rice contains genes from a common soil bacterium Erwinia and maize and contains increased levels of beta-carotene which can be converted by the body into vitamin A. Golden Rice is developed as a potential new way to address vitamin A deficiency (Dawe et al. 2002; Zimmermann and Qaim 2004; Paine et al. 2005). The biofortified foods may also be useful for increasing micronutrient uptake in high-income countries. Researchers at the University of Warwick have been looking for ways to boost the low selenium levels in British grains and have been working to help develop a grain to be used in making bread biofortified with selenium (Wikipedia 2015).
The approach of biofortification may have advantages over other health interventions such as providing fortified foods after processing, or providing supplements. Biofortification is also fairly cost-effective, and the implementation costs of growing biofortified foods are much lower than supplementation which is comparatively expensive. The dietary diversification might be one of the methods to reduce the prevalence of iron deficiency. Thus, biofortification (the use of traditional plant breeding methods or genetic engineering to improve the available iron content of staple food crops) holds promise for the future (Nestel et al. 2006). It would make it possible to deliver iron, zinc, vitamin A, or any other desired nutrient to those most in need. They often depend on subsistence farming and have limited access to fortified foods. More researches are needed to develop successful crops with adequate bioavailable iron, zinc, vitamin A, and folate (vitamin B9) content.
3 Common Problems of Biofortification
Although there is acceptance for biofortified food, some people are against the genetically modified (GM) crops such as Golden Rice. There are difficulties in acceptance of biofortified foods, if they have different characteristics to their unfortified counterparts. For example, vitamin A-enhanced foods are often dark yellow or orange in color – this, for example, is problematic for many in Africa, where white maize is eaten by humans, whereas yellow maize is associated with animal feed (McClafferty and Islam 2008), or where white-fleshed sweet potato is preferred than orange-fleshed counterpart (Nestel et al. 2006). Some qualities may be relatively simple to breed out of biofortified crops according to consumer demand. Due care must be taken to convince the local farmers and consumers that the crop in question is worth growing and consuming. This can be done through improving the cultivation qualities of the plant, for example, making the orange sweet potato mature earlier than white-fleshed sweet potato so that it can be taken to market earlier. It is possible through public health education, elaborating the benefits of eating biofortified foods. While other micronutrients such as zinc or iron can be added to crops without noticeably changing their taste or appearance, some of the consumers do not think that their food has been altered and do not bother much about the external appearance.
Some of the people did disapprove of the biofortification programs, because biofortification is a strategy that aims to concentrate more nutrients in few staple foods. The lack of access to a diverse and balanced diet is the major cause of malnutrition. The use of biofortification as part of a larger strategy involving diversification of foods in the developing world is necessary to solve the problem of deficiency of certain essential nutrients. The biofortification is accepted as a long-term strategy. The diversity in substantially increasing diet will take many decades, and the biofortification will be an effective strategy to reduce the micronutrient malnutrition.
4 Recompense of Biofortification
The biofortification strategy seeks to take advantage of the consistent daily consumption of large amounts of food staples by all family members, including women and children who are most at risk for micronutrient malnutrition. As a consequence of the predominance of food staples in the diets of the poor, this strategy implicitly targets low-income households. After the one-time investment is made to develop seeds that fortify themselves, recurrent costs are low and germplasm can be shared internationally. The biofortified crops are highly sustainable, and the nutritionally improved varieties will continue to be grown and consumed year after year.
Biofortification provides a truly feasible means of reaching malnourished populations in relatively remote rural areas, delivering naturally fortified foods to people with limited access. Breeding for higher trace mineral density in seeds will not incur a yield penalty. In fact, biofortification may have important effects for increasing farm productivity in developing countries in an environmentally beneficial way. Recent researches have shown that trace mineral-packed plants have high level of resistance against diseases and other environmental stresses. More seedlings survive and initial growth is more rapid.
4.1 Consequences of Micronutrient Malnutrition
More than two billion people worldwide are iron deficient (Stoltzfus 2001). Iron-deficiency anemia is by far the most common micronutrient deficiency in the world. Iron efficiency during childhood and adolescence impairs physical growth, mental development, and learning capacity. In adults, iron-deficiency anemia reduces the capacity to do physical labor. Iron deficiency increases the risk of women dying during delivery or in the postpartum period.
The zinc deficiencies have equally serious consequences for health. For example, meta-analyses of recent randomized controlled trials show that zinc supplementation can reduce morbidity from a number of common childhood infections, especially diarrhea, pneumonia, and possibly malaria, by one-third. In addition, zinc deficiency is an important cause of stunting. Billions of people are also at risk for zinc deficiency. As for anemia, prevalence is highest for South and Southeast Asia and Africa. Because there is no widely accepted method for measuring zinc deficiency, no estimates are available of numbers of people who are zinc deficient.
Globally, approximately three million preschool-age children have visible eye damage owing to vitamin A deficiency. Annually, an estimated 250,000–500,000 preschool children go blind from this deficiency, and about two-thirds of these children die within months of going blind. Even more importantly, the last two decades have brought an awareness that vitamin A is essential for immune function. Estimates of the prevalence of subclinical vitamin A deficiency range between 100 million and 250 million for preschool children.
People with subclinical vitamin A deficiency more often experience anemia, impaired linear growth, and morbidity from common childhood infections such as respiratory and diarrheal diseases, measles, and malaria. Most importantly, a number of randomized controlled trials in developing countries have shown that administration of vitamin A capsules among infants and preschool children reduces mortality rates from all causes by 23 %, and that administration of capsules with vitamin A and beta-carotene among women during childbearing years can reduce maternal mortality related to pregnancy by 40 % and 49 %, respectively. Prevalence of vitamin A deficiencies by region is available only for preschool children. Moreover, similar to iron and zinc, prevalence is highest in South and Southeast Asia and sub-Saharan Africa.
4.2 Underlying Causes of Micronutrient Malnutrition
Billions of people in developing countries suffer from an insidious form of hunger known as micronutrient malnutrition. Even mild levels of micronutrient malnutrition may damage cognitive development, lower disease resistance in children, and reduce the likelihood that mothers survive childbirth. The costs of these deficiencies in terms of lives lost and poor quality of life are staggering (Graham et al. 2001). It is important to identify who the malnourished are, where they are located, and what they eat in order to develop an effective strategy to reduce micronutrient malnutrition. And they are mostly women and children, who reside in developing countries where dietary quality is often poor. In addition, they have higher nutritional requirements due to reproduction and rapid growth resulting in malnutrition.
The primary underlying cause of micronutrient malnutrition is poor quality diets, characterized by high intakes of food staples, but low consumption of animal and fish products, fruits, and vegetables, which are rich sources of bioavailable minerals and vitamins. As such, most of the malnourished are those who cannot afford to purchase high-quality, micronutrient-rich foods or who cannot obtain these foods from their own production.
What is perhaps most alarming, however, is the upward trend in non-staple food prices. Cereal prices have fallen by 40 % since the early 1970s. The Green Revolution can rightly take credit for its crucial contribution to this tremendous achievement. Falling cereal prices have not only led to increased food security in terms of energy, but also allowed greater purchases of non-staples by freeing up cash. Unfortunately, the rate of production of non-staple foods (e.g., fruits, vegetables) has not kept pace with demand, so that these micronutrient-rich food sources have become ever more expensive for the poor (Bouis 2003).
4.3 Role of Iron, Zinc, and Folate in Human Beings
Iron is integral to the structure and function of red blood cells, and its deficiency can result in anemia. Anemia and iron deficiency during pregnancy can cause preterm birth and low birth weight (Allen 2001). In non-anemic mothers, iron supplementation may offer protection against low birth weight (Palma et al. 2008). Iron is also involved in myelination, neurotransmitter function, various cellular and oxidative processes, energy production, and thyroid hormone metabolism. Iron deficiency has been implicated in neurological and cognitive disorders in the mother; these include major depressive disorder, recognized to have health consequences on both the mother and child (Bodnar and Wisner 2005; Leung and Kaplan 2009).
The high prevalence of iron deficiency in developing countries has on people’s well-being and productivity. Physical work capacity is reduced. Iron deficiency in pregnancy contributes to the risk of severe anemia and increased maternal morbidity and mortality (Khan et al. 2006). Iron-deficiency anemia in early pregnancy is associated with a higher risk of preterm delivery (Scholl 2005). Iron supplementation (usually in combination with folic acid) in pregnancy has been reported to reduce the risk of postpartum hemorrhage (Christian et al. 2009a), improve birth weight (Cogswell et al. 2003), and reduce early neonatal (Zeng et al. 2008; Titaley et al. 2010) and childhood mortality (Christian et al. 2009b).
Zinc is integral to DNA synthesis and necessary for the structure and function of regulatory, structural, and enzymatic proteins as well as cell membranes. It is involved in neurological function and proper immune function (Fraker et al. 2000; Huang 1997). Zinc deficiency is also implicated in depressive disorders. Moreover, various studies have implicated zinc deficiency in preterm and low birth weight, although routine supplementation is not recommended unless there is an identified deficiency (Ladipo 2000).
Folate is involved in the metabolism of nucleic acids and amino acids and in neurological functioning. While inadequate folate is implicated in various birth defects and poor pregnancy outcomes, its role in neural tube defects has received the most attention. In various countries, women of childbearing age are advised to take supplements. Folate deficiency is also implicated in depressive disorders. Food fortification policies are in effect in response to the strong evidence of the importance of folic acid intake in the very early stages of pregnancy (Wilson et al. 2003).
The RDA is defined as the average daily dietary intake level that is sufficient to meet the nutrient requirements of nearly all healthy individuals in a particular life-stage and gender group. For pregnant women, the RDA of iron and zinc is 27 and 11 mg/day, respectively, while that of folate as dietary folate equivalents (DFE) is 600 μg/day.
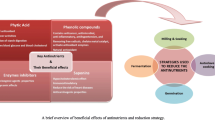
5 Role of Antinutrients in Biofortification
Nutritional iron deficiency occurs when the diet supplies inadequate bioavailable iron to meet the body’s requirements for growth and pregnancy and to replace iron lost from the gastrointestinal tract and in the urine and through menstruation (in women). The causes have been known for over 50 years. The major factor is poverty. Secondly, the agricultural revolution replaced animal foods rich in bioavailable iron by cereals, legumes, and plant-based diets (Cordain 1999). The classical iron balance study carried out by Widdowson and McCance (1942) demonstrated that less iron was absorbed from bread with high bran content than from white bread. The inhibitory role of an antinutrient called phytate was suspected, because phytic acid was shown to inhibit iron absorption (McCance et al. 1943). Later on, a second major class of antinutrient known as polyphenols was discovered (Disler et al. 1975; Gillooly et al. 1983). This large body of experimental work provided the basis for predicting dietary bioavailability of iron (Reddy et al. 2000; Hallberg and Hulthen 2000) and designing efficacious strategies for alleviating nutritional iron-deficiency anemia (Hurrell 1999; Hurrell et al. 2010).
Wheat flour has been fortified with iron in America since the 1940s, and the marked decline in the prevalence of iron-deficiency anemia in infants and young children in the United States is generally attributed to the fortification of foods (Miller et al. 1985; Fomon 2001). Iron is also added to many processed foods, such as breakfast cereals, in Western countries (Yip 2002). Despite a relatively clear understanding of the physiology of food iron absorption, iron deficiency is estimated to affect as many as two billion people (Zimmermann and Hurrell 2007). Developed countries may have a mechanism to provide bioavailable micronutrient, but the poor mass of developing countries depend on staple food such as maize, rice, wheat, or grain legumes. Therefore, there is an urgent need to supply biofortified food rich in these micronutrients. Truly speaking, these biofortified foods contain antinutrients like phytates and polyphenols, which are required to be minimized by processing such as soaking, germination, fermentation, cooking, or autoclaving so as to make the nutrient available to human beings.
In developing countries, screening for iron deficiency is usually based solely on hemoglobin (Hb) measurements. Sensitivity is low because the overlap in Hb concentrations between healthy and iron-deficient individuals is considerable, especially if the cutoff values used to identify anemia are not appropriately adjusted for age, gender, pregnancy, ethnicity, and altitude. Specificity is poor, because a multitude of disorders other than iron deficiency can cause anemia. They include other nutritional deficiencies due to vitamins A, B12 (cyanocobalamin), and B9 (folic acid). Ferrous fumarate, a less reactive yet bioavailable form of iron, is recommended or used in many countries in Central and South America. The use of encapsulation technologies may provide a practical solution to fortification with more reactive bioavailable compounds. Condiments such as salt, soy sauce, fish sauce, and curry may be an alternative for the delivery of fortified iron. It will be essential to keep legislators and program managers informed about the necessity of providing iron in a bioavailable form and in sufficient quantities to meet the needs of women, children, and adolescents at highest risk.
Biofortification holds promise for the future. It would make it possible to deliver iron to those most in need. They often depend on subsistence farming and have limited access to fortified foods. A modest improvement in iron stores in non-anemic Filipino women consuming iron-biofortified rice was reported in one recent trial. However, the meals provided an average of only 1.42 mg/d additional iron (Lynch 2011). More research is needed to develop a crop/genotypes with adequate improvements in bioavailable iron content.
5.1 Processing to Minimize Antinutrients
It is very necessary now to move from food security to nutrition security and improve the quality of foods both in macro- and micronutrients in order to break the transgenerational effects of malnutrition. Enhancing food and nutrition security through innovative diversified agriculture and dietary practices, prevention and control of infection, promotion of food safety, and fortification of staples with appropriate attention on emerging chronic disorders are essential (Krishnaswamy 2001). The low bioavailability of some key micronutrients from foods, such as Fe and Zn, is substantially enhanced with the right food combinations and with appropriate food processing and preparation techniques. Simple appropriate technology for the preservation of micronutrient-rich foods would need further development and promotion for their year-round availability (Tontisirin et al. 2002). As iron deficiency is the most common micronutrient deficiency in the world, iron biofortification is a preventative strategy that alleviates Fe deficiency by improving the amount of absorbable Fe in crops. High Fe-bioavailability maize contains more bioavailable Fe than the low Fe-bioavailability maize (Tako et al. 2013). Maize shows promise for Fe biofortification; therefore, human trials need to determine the efficacy of consuming the high bioavailable Fe maize to reduce Fe deficiency.
International research efforts, including those funded by HarvestPlus, a challenge program of the Consultative Group on International Agricultural Research (CGIAR), are focusing on conventional plant breeding to biofortify staple crops such as maize, rice, cassava, beans, wheat, sweet potatoes, and pearl millet to increase the concentrations of micronutrients that are commonly deficient in specific population groups of developing countries (La Frano et al. 2014). The bioavailability of micronutrients in unfortified staple crops in developing regions is typically low. Reducing the amounts of antinutrients and food processing generally increases the bioavailability of micronutrients. In general, biofortified foods with relatively higher micronutrient density have higher total absorption rates than non-biofortified varieties. There is a need to breed plants with increased micronutrient concentrations in order to decrease the influence of inhibitors and to offset losses from processing.
5.2 Important Antinutrients in Food
The antinutrients have impact on lowering the bioavailability of essential nutrients. There is a need to summarize various antinutrients influencing directly or indirectly the bioavailability of these micronutrients. Most of the cereals and legumes are rich in these micronutrients; therefore, techniques to minimize these antinutrients to enhance the bioavailability of micronutrients are required to be developed or brought to practice. Some of the antinutrients influencing bioavailability of micronutrients are summarized below.
5.2.1 Phytates
Phytic acid, myoinositol 1,2,3,4,5,6-hexakis (dihydrogen phosphate), occurs abundantly in most of the food legumes. It is a chelating agent for cations as well as for phosphorus storage in many seeds. Phytate rapidly accumulates in seeds during the ripening period. Excessive phytic acid in the diet can have a negative effect on mineral balance because of the insoluble complexes it forms with essential minerals (Cu2+, Zn2+, Fe3+, and Ca2+), which cause poor mineral bioavailability (Zhou and Erdman 1995; Urbano et al. 2000). The ability of phytic acid to complex with proteins and particularly with minerals (Ca, Fe, Zn) has been a subject of investigation for several reasons. The interaction between phytate and proteins leads to decreased solubility of proteins and amino acid digestibility. The calcium ions also interact with proteins and phytate and decrease the solubility of proteins. The reduced solubility of proteins as a result of protein phytate complex adversely affects certain functional properties of protein which are dependent of their hydration and solubility. The ability of phytic acid to bind with minerals, proteins, or starch, directly or indirectly, may alter solubility, functionality, digestibility, and absorption of these nutrients. In addition, monogastric animals have a limited ability to hydrolyse phytates and release phosphate for absorption due to a lack of intestinal phytases (Zhou and Erdman 1995; Greiner and Konietzny 1996a; Urbano et al. 2000). Phytic acid is able to make complex with proteins also, decreasing protein solubility. Therefore, phytates have negative impact on enzyme activity, and there is evidence of its negative effects on key digestive enzymes like lipase, α-amylase, pepsin, trypsin, and chymotrypsin (Thompson 1993; Greiner and Konietzny 1996b; Urbano et al. 2000). The binding of phytic acid to these enzymes reduces nutrient digestibility. Phytic acid also binds with starch through phosphate linkages (Lajolo et al. 2004). Efforts made in developing high-iron and zinc-biofortified crops remain unutilized due to reduced bioavailability; therefore, technology to remove phytates from the grains is required to be developed to enhance the bioavailability of these micronutrients (Table 24.1).
Phytate-mineral complexes are well documented. The nutritionally important minerals such as Ca, Mg, Cu, Fe, and others form complexes with phytic acid, resulting in reduced solubility of the metals (Biehl et al. 1995). Low iron availability due to iron-phytate complexes is also of concern. As most of us consume enough minerals in common foods which enable us to meet more than our requirement, and small amounts of these micronutrients might be tied up by phytates. The phytate-associated deficiencies of iron and zinc do occur in some third-world countries where people mostly eat grains.
The bioavailability of iron and zinc can be increased by combination of processing techniques of grain, viz., soaking in water, germination, fermentation, cooking, pressure cooking, etc. As far as food legumes are concerned, they are generally consumed after processing. The purpose behind these processing is to remove or minimize the antinutrients present in the grain. Routine processing such as soaking of grain and discarding the soaking water and then cooking or germination and fermentation reduces phytic acid content of food to an extent of up to 50–70 % (Iyer et al. 1980, 1989; Reddy et al. 1982). Soaking in yogurt, buttermilk, or water combined with lemon juice or vinegar is also much helpful to enhance this breakdown (Table 24.2).
Soaking of seed can eliminate up to 69.6 % of phytate in beans (Iyer et al. 1980), whereas germination can reduce phytate content to an extent of 77.4 % in seeds of various food legumes (Reddy et al. 1982). Highest reduction in phytate content of different food legumes was reported during germination, followed by fermentation, autoclaving, and roasting (Chitra et al. 1996). Germination (24–96 h) of seeds of Vigna unguiculata has been found to eliminate 38–95 % of total phytates, whereas germination followed by autoclaving removed 96–100 % of total phytates (Kalpanadevi and Mohan 2013). They also reported that soaking of seeds in water for 12 h followed by cooking or autoclaving removed 68–71 % of phytates. Soaking of seed in distilled water or 2 % sodium carbonate solution or 1 % citric acid solution followed by cooking eliminates 45.1–73.4 % of phytates in soybean (Sharma et al. 2013). Removal of phytates enhances the bioavailability of iron, zinc, and other minerals. The phytate has some health benefits, including anti-inflammatory effects. In laboratory research, phytate has helped normalize cell growth and stopped the proliferation of cancer cells. They also may help prevent cardiovascular disease and lower a food’s glycemic load. Some myoinositol phosphates, including IP6 from soybean, have been suggested to have beneficial health effects, such as amelioration of heart disease by controlling hypercholesterolemia and atherosclerosis, prevention of kidney stone formation, and a reduced risk of colon cancer (Greiner et al. 2002).
5.2.2 Polyphenols
The polyphenols or tannins are one of the most important antinutrient of food legumes and found in the range of 0.45–20.00 mg g−1 grain. Among the food legumes, urd bean, mung bean, kidney beans, faba beans, and pigeon peas contain higher content of tannins. The tannin content in pulses varies with the color of the seed coat or testa. The white- or light-colored seeds generally have low tannins than dark-colored (red, brown, bronze, black) varieties. The polyphenol content of seed changes during seed maturation, and it reduces generally during maturation of seed due to polymerization of polyphenolic compounds to high molecular weight insoluble polymers such as lignins.
The seed coat is having the major amount of polyphenols, and the process of dehulling or dehusking reduces the polyphenols of seed drastically (Table 24.3). Reducing the polyphenolic compounds from the grain helps in enhancing the bioavailability of minerals such as iron and zinc and protein. Indian diet is composed of several ingredients (rice, wheat, maize, sorghum, dehulled beans, leafy and other vegetables, roots and tubers, eggs, fish, meat, sugar, and oil), and about daily intake of 10 % of tannins is derived from food legumes.
The polyphenolic compounds cause decreased secretion of digestive enzymes, toxicity of absorbed tannins, increased excretion of endogenous protein, and formation of tannin complexes with dietary protein and other food components. The greater tendency of tannins to form complexes with proteins rather than carbohydrates and other food polymers is attributed to the strong hydrogen bond affinity of the carboxyl oxygen of the peptide group. One tannin molecule binds two or more carboxyl oxygen of peptide group with possible formation of cross-links between the protein chains. It is also reported that tannin-protein complexes are responsible for growth depression, low protein digestibility, decreased amino acid availability, and decreased fecal nitrogen (Reddy et al. 1985). Tannins also reduce the bioavailability of iron and form tannin-iron complexes. Low tannin varieties have higher bioavailability of micronutrients (Fe, Zn), and processing such as soaking in water or other solutions removes major part of tannins. Other processing such as germination, soaking before cooking, or autoclaving is a boon to reduce tannins and ultimately higher iron and zinc bioavailability (Table 24.4).
The polyphenols can be removed by processing technique such as dehulling, soaking, cooking, germination, fermentation, etc. However, breeding approach is more effective in breeding varieties with lower tannins. Biofortification is therefore finding valuable importance in lowering down the tannin content of seed. Processing of grain before consumption is equally effective in removing tannins to enhance the bioavailability of iron and zinc. Dehulling eliminates about 74–98 % of tannins in beans (Reddy et al. 1985). Dehulling also helps in improving in vitro protein digestibility and ionizable iron absorption. Soaking of grain in water removes 18–74 % of tannins of legumes, whereas soaking in sodium carbonate solution (2 %) for 12 h causes a reduction of 67.2–90.9 % of tannins. Cooking of grain of legumes reduces 37.5–77.0 % of tannins (Reddy et al. 1985), whereas germination of seed for 24–48 h removes 36–64 % of total tannins of seed (Rao and Deostale 1982). Germination of seeds is very effective in removing tannins (32–76 %), whereas soaking followed by cooking or autoclaving remove 79–84 % of total tannins (Kalpanadevi and Mohan 2013). Soaking of soybean seeds in water or 1 % citric acid solution followed by cooking removed 42.5–60.4 % of tannins (Sharma et al. 2013). It is, therefore, established that soaking, germination, and cooking or autoclaving of seed help in removing major amount of tannins. Removal of tannins from seed ultimately enhances the bioavailability of iron, zinc, and protein. Therefore, biofortified grains must be consumed after conventional processing to enhance the bioavailability of micronutrients (iron and zinc).
5.2.3 Lectins
Lectins are proteinaceous compounds commonly found in leguminous plants. In 1908, it was reported that the seeds of edible species of some common legumes such as lentils, navy beans, and garden peas contained phytohemagglutinins. Extracts of many edible crude legume seeds agglutinated red blood cells, although hemagglutinating activity has been detected in over 800 different plant species, of which over 600 are from the family Leguminosae. The lectins have molecular weight ranging from 100,000 to 150,000. Some of the lectins sometimes exhibit toxicity. Most of the lectins contain 4–10 % carbohydrates. Concanavalin A of jack beans and peanut lectins are devoid of carbohydrates.
The purified lectins from beans are toxic and sometime cause death of young rats. The lectins from immature seeds of soybean, cowpea, lima beans, pigeon pea, and rice beans when injected in young rats produced liver damage and death. However, lectins from mung bean have been reported to be nontoxic. Traditionally, lectins have been measured by their hemagglutinating activity (Grant 1991). However, when possible, enzyme-linked immunosorbent assay (ELISA)-based immunological methods that recognize specific antibodies are being increasingly used due to their higher specificity (Hajos et al. 1996; Muzquiz et al. 2001). Grant et al. (1983) found that among the 15 legumes they studied, chickpea seeds had the lowest hemagglutinin activities and were nontoxic.
Processing such as soaking, germination, cooking, or autoclaving helps in removing most of the lectins (Kalpanadevi and Mohan 2013; Vasishtha 2011; Srivastava and Vasishtha 2013a, b and 2014; Vasishtha et al. 2012 and 2014; Vasishtha et al. 2012). Autoclaving for 5–30 min removes entire lectins of grain, depending on the type of legume. Detoxification of beans can be achieved by ordinary process of cooking (Table 24.5).
Nutritionally, dietary lectins vary considerably in the nature and extent of their antinutritional effects. Lectins can be toxic; they can interfere with hormone balance and deplete nutrient reserves leading to severe growth depression and a high incidence of deaths. However, lectins could be also useful as it stimulates gut function, limits tumor growth, and ameliorates obesity (Pusztai et al. 2004).
5.2.4 Protease Inhibitors
Several substances which have the ability to inhibit the proteolytic activity of certain enzymes are found in food grains. Trypsin inhibitors belong to a broad class of proteins (protease inhibitors) that inhibit proteolytic enzymes. Trypsin and chymotrypsin inhibitors are of much importance. Trypsin inhibitor activity increases as seed maturity progresses. Most of the plant protease inhibitors are destroyed by heat resulting in enhancement of nutritive value of protein. Heating is employed for cooking of grains under moist condition. Moisture content, time, temperature, and pressure are the major factors that influence cooking rates, inactivation of trypsin inhibitor, and subsequent increase in nutritive value. Moist heat has been shown to be effective in destroying trypsin inhibitor activity in food grains especially food legumes. Germination also influences the trypsin inhibitor activity.
Over the last two decades, it was observed that protease inhibitors have been linked to health-promoting properties (Champ 2002) and are considered as natural bioactive substances (Hill 2004). Protease inhibitors may act as anticarcinogenic agents (Clemente et al. 2004). It also reduced the incidence and frequency of colon tumors in dimethylhydrazine-treated rats. However, this effect was not observed with autoclaved Bowman-Birk inhibitors (BBI), suggesting that protease inhibitor activity was necessary for anticarcinogenic activity (Kennedy et al. 2002).
5.2.5 α-Amylase Inhibitors
α-Amylases (α-1,4-glucan-4-glucanohydrolases) are endoamylases that catalyze the hydrolysis of α-D-(1,4) glycosidic linkages, which occur in starch and related compounds. They play a major role in the carbohydrate metabolism of animals and humans by providing them glucose as an energy source and as a building block for synthesis of other sugars. Among α-amylase inhibitors (α-AIs) found in plants, legume α-AIs, especially α-AIs from beans, have received considerable attention (Whitaker 1988). Jaffe et al. (1973) screened 95 legume cultivars for α-AI levels and found that lima beans (Phaseolus lunatus), mung beans (Phaseolus aureus), and horse gram (Dolichos biflorus L.) had the highest levels of inhibitory activity. Although little is known about the screening of αAIs in chickpea, Mulimani et al. (1994) determined the α-AI activity of 28 varieties of chickpea and found variations ranging from 11.6 to 51.4 inhibitory units g−1. Singh et al. (1982) observed that the amylase inhibitor activity on pancreatic amylase of chickpea cultivars ranged from 7.8 to 10.5 units g−1 (desi) and 5.6 to 10.0 units g−1 (kabuli), with substantial variations among these cultivars. A similar variation but of lower magnitude was observed using salivary amylase. A comparison under similar assay conditions indicated that the amylase inhibitor activity had a stronger influence on pancreatic amylase than salivary amylase for both desi and kabuli cultivars. Jaffe et al. (1973) reported that the partially purified kidney bean inhibited the salivary amylase more than the pancreatic amylase. This shows that amylase inhibitors from different legume seeds may exhibit unequal activity against different enzymes.
α-AIs reduce amylase activity and starch digestion in the gut when given orally to humans (Singh et al. 1982). As a result, they lower postprandial increases in circulating glucose and insulin. These inhibitors may therefore prove to be useful in the treatments of obesity or diabetes mellitus.
5.2.6 Saponins
Saponins are secondary plant metabolites present in many of the food grains. It contains a carbohydrate moiety (mono or oligosaccharide) attached to an aglycone, which may be steroidal or triterpenoid in structure. The complexity of the saponin structure depends on the variability of the aglycone structure, the attachment position of the glycosidic moieties, and the nature of these glycosides. Aglycones are generally linked to D-galactose, L-rhamnose, D-glucose, D-xylose, D-mannose, and D-glucuronic acids, some of which may be acetylated (Fenwick et al. 1991). All legumes have triterpene-type saponins.
The chemical composition of soybean (G. max) saponins has been extensively investigated. They are triterpene saponins (known as soyasaponins), of which more than ten types have been isolated (Shibuya et al. 2006). Soyasapogenol B was identified as the aglycone in saponins from different legume species (Price et al. 1987, 1988; Ayet et al. 1996). Tava et al. (1993) found 17.7 and 21.9 g kg−1 of soyasapogenol B in desi and kabuli types, respectively. Ruiz et al. (1996, 1997) showed the presence of soyasaponin VI, a conjugated form of soyasaponin I, in many legumes and was the only saponin detected in the seeds of chickpea. Soyasaponin VI may have an important physiological role in preventing lipid peroxidation of DNA and proteins by free radical attack.
The saponins are now considered beneficial because of their cholesterol-lowering property and antioxidant and cancer protective effect. The saponins undergo degradation during cooking and pressure cooking. Soaking and germination reduce saponins to some extent. Saponins of food legumes undergo certain changes during soaking, germination, cooking, and autoclaving (Srivastava and Vasishtha 2012 and 2014; Vasishtha and Srivastava 2011 and 2014).
Potential benefits from the consumption of food saponins include a reduced risk of cardiovascular disease and some cancers. Dietary saponins have been repeatedly shown to lower plasma cholesterol in animals. However, their hypocholesterolemic effect in humans is more speculative. They may also have anticarcinogenic properties, as suggested by a recent rodent study in which feeding a saponin-containing diet inhibited the development of preneoplastic lesions in the colon (Koratkar and Rao 1997). Kerem et al. (2005) found that the major saponin in chickpea seeds possesses antifungal properties. In addition, some saponins have been reported to inhibit in vivo human immunodeficiency virus (HIV) infectivity (Thompson 1993).
5.2.7 Lathyrogens
Lathyrogens are natural toxins present in seeds of lathyrus. The seeds of lathyrus contain a neurotoxic compound commonly called BOAA (β-N-oxalyl amino alanine) or ODAP (β-N-oxalyl L-α, β-diamino propionic acid). This compound is responsible to cause neurolathyrism in human beings. Neurolathyrism is the condition which involves degeneration of part of the spinal cord, affecting lower limbs and infects disability in locomotion. The central nervous system is affected. Neurological lesions of spinal cord degeneration result in weakness and spastic paralysis of the legs, convulsions, and death in extreme cases (Striepler et al. 1978; Attal et al. 1978). The disease is irreversible, though drugs may give temporary relief from muscular rigidity. Lathyrus is prevalent especially during the period of famines when it is cultivated as an alternative to other crops, and the poor sections of society depend mainly on lathyrus as their staple food for several months. Epidemiological surveys have indicated that young men between the age of 15 and 45 are most severely affected by lathyrism. A survey conducted by ICMR (1964) revealed that a diet consisting of about 25 % lathyrus for a period of 2–6 months produces lathyrism.
The neurotoxin present in lathyrus is BOAA, which exists in alpha and beta isomeric forms. Both forms yield oxalic acid and α, β-diamino propionic acid on hydrolysis. Of the two forms, β-isomer is predominant and accounts for 95–96 % of total BOAA content in different varieties of lathyrus. The BOAA in seed of lathyrus is found in the range of 0.05–0.3 %. Certain varieties are having very low BOAA and considered not harmful from the health point of view if consumed judiciously after proper processing. Soaking of dehusked grain in water for 6–12 h removes almost 1/3rd of neurotoxin (Srivastava and Srivastava 2002). Soaking of dehusked grain in acidic solution of pH 4.0 or alkaline solution of pH 9.2 for 30–60 min at 80–100 °C helps in removing major amount of BOAA (Srivastava and Singh 2013). Heating the dehusked grain at 120–150 °C for 2 h also removes about 40 % of total BOAA (Srivastava and Singh 2013). The BOAA content of less than 0.1 % has been considered to be safer from the neurotoxicity point of view (Siddiq 1995). Firstly, consumption of lathyrus should be advocated after necessary processing so as to minimize the neurotoxin content from the health point of view. Secondly, the genotypes having reasonably low BOAA should be encouraged for human consumption.
6 Conclusion
Biofortification enhances the bioavailability of micronutrients such as iron, zinc, vitamin A, folic acid, and cyanocobalamin in staple foods. Almost half of the world population is suffering from these deficiencies as they have either low access or no access for these quality foods. The presence of antinutrients such as phytates, polyphenols and tannins, protease and α-amylase inhibitors, saponins, lectins, and lathyrogens in food causes hurdle in absorption and bioavailability of Fe, Zn, Ca, folates, etc. Various processing techniques like soaking, germination, milling (dehusking), cooking, and autoclaving are effective in removal of these antinutrients.
References
Allen LH (2001) Biological mechanisms that might underline iron’s effects on fetal growth and preterm birth. J Nutr 131(2S–2):581S–589S
Attal HC, Kulkarni SW, Choubey BS, Palkal ND, Deotale PG (1978) A field study of lathyrism- some critical aspects. Indian J Med Res 67:608–611
Ayet G, Muzquiz M, Burbano C, Robredo LM, Cuadrado C, Price K (1996) Determinación de saponinas en las principales leguminosas cultivadas en España. (Determination of saponins as the main legumes cultivated in Spain). Food Sci Technol Int 2:96–100
Biehl RR, Baker DH, DeLuca HF (1995) 1-α-hydroxylated cholecalciferol compounds act additively with microbial phytase to improve phosphorus, zinc and manganese utilization in chicks fed soy-based diets. J Nutr 125:2407–2416
Bodnar LM, Wisner KL (2005) Nutrition and depression implications for improving mental health among childbearing aged women. Biol Psychiatry 58(9):679–685
Bouis HE (2003) Micronutrient fortification of plants through plant breeding: can it improve nutrition in man at low cost? Proc Nutr Soc 62:403–411
Champ MM (2002) Non-nutrient bioactive substances of pulses. Br J Nutr 88:307–319
Chitra U, Singh U, Venkateswararao P (1996) Phytic acid, in vitro protein digestibility, dietary fiber and minerals of pulses as influenced by processing methods. Plant Food Hum Nutr 49:307–316
Christian P, Khatry SK, Leclerq SC, Dali SM (2009a) Effects of prenatal micronutrient supplementation on complications of labor and delivery and puerperal morbidity in rural Nepal. Int J Gynecol Obstet 106(1):3–7. doi:10.1016/j.ijgo.2009.03.040
Christian P, Stewart CP, LeClerq SC, Wu L, Katz J, West KP Jr, Khatry SK (2009b) Antenatal and postnatal iron supplementation and childhood mortality in rural Nepal a prospective follow-up in a randomized, controlled community trial. Am J Epidemiol 170:1127–1136
Clemente A, Mackenzie DA, Johnson IT, Domoney C (2004) Investigation of legume seed protease inhibitors as potential anticarcinogenic proteins. In: Muzquiz M, Hill GD, Pedrosa MM, Burbano C (eds) Proceedings of the fourth international workshop on antinutritional factors in legume seeds and oilseeds, EAAP Publication No. 110, Wageningen, pp 137–142
Cogswell ME, Parvanta I, Ickes L, Yip R, Brittenham GM (2003) Iron supplementation during pregnancy, anemia, and birth weight a randomized controlled trial. Am J Clin Nutr 78:773–781
Cordain L (1999) Cereal grains humanity’s double-edged sword. World Rev Nutr Diet 84:19–73
Dawe D, Robertson R, Unnevehr L (2002) Golden rice: what role could it play in alleviation of vitamin A deficiency? Food Policy 27(5–6):541–560
Deshpande SS, Cheryan M (1983) Changes in phytic acid, tannins and trypsin inhibitory activity on soaking of dry beans (Phaseolus vulgaris L.). Nutr Rep Int 27(2):371–375
Disler PB, Lynch SR, Charlton RW, Torrance JD, Bothwell TH, Walker RB, Mayet F (1975) The effect of tea on iron absorption. Gut 16:193–200
Fenwick GR, Price KR, Tsukamoto C, Okubo K (1991) Saponins. In: D’Mello JPF, Duffus CM, Duffus JH (eds) Toxic substances in crop plants. Royal Society of Chemistry, Cambridge, MA, pp 285–327
Fomon S (2001) Infant feeding in the 20th century formula and beikost. J Nutr 131:S409–S420
Fraker PJ, King LE, Laakko T, Vollmer TL (2000) The dynamic link between the integrity of the immune system and zinc status. J Nutr 130(5S):1399S–1406S
Gillooly M, Bothwell TH, Torrance JD, MacPhail AP, Derman DP, Bezwoda WR, Mills W, Charlton RW, Mayet F (1983) The effects of organic acids, phytates and polyphenols on the absorption of iron from vegetables. Br J Nutr 49:331–342
Graham R, Welch R, Bouis H (2001) Addressing micronutrient malnutrition through the nutritional quality of staple foods: principles, perspectives, and knowledge gaps. Adv Agron 70:77–142
Grant G (1991) Legumes. In: D’Mello JPF, Duffus CM, Duffus JH (eds) Toxic substances in crop plants. Royal Society of Chemistry, Cambridge, MA, pp 49–67
Grant G, More LJ, McKenzie NH, Stewart JC, Pusztai A (1983) A survey of the nutritional and haemagglutination properties of legume seeds generally available in the UK. Br J Nutr 50:207–214
Greiner R, Konietzny U (1996a) Phytate hydrolysis in black beans by endogenous and exogenous enzymes. In: Bardocz S, Muzquiz M and Pusztai A (eds) Effects of antinutrients on the nutritional value of legume diets. Proceedings of the fourth scientific workshop, official publications of the European communities, Luxembourg, pp 19–26
Greiner R, Konietzny U (1996b) Construction of a bioreactor to produce special breakdown products of phytate. J Biotechnol 48:153–159
Greiner R, Larsson AM, Carlsson NG, Muzquiz M, Burbano C, Cuadrado C, Pedrosa MM, Goyoaga C (2002) Pathway of dephosphorylation of myoinositol hexakisphosphate by phytases of legume seeds. J Agric Food Chem 50:6865–6870
Hajos G, Gelencser E, Pusztai A, Bardocz S (1996) Measurements of the antinutrient contents of legume samples. In: Bardocz S, Gelencser E, Pusztai A (eds) Effects of antinutrients on the nutritional value of legume diets, vol I. Official Publications of the European Communities, Luxembourg, pp 130–134
Hallberg L, Hulthen L (2000) Prediction of dietary iron absorption an algorithm for calculating absorption and bioavailability of dietary iron. Am J Clin Nutr 71:1147–1160
Hill GD (2004) Grain legumes and oilseeds: the way ahead. In: Muzquiz M, Hill GD, Pedrosa MM, Burbano C (eds) Proceedings of the fourth international workshop on antinutritional factors in legume seeds and oilseeds. EAAP Publication No. 110, Wageningen pp 353–364
Huang EP (1997) Metal ions and synaptic transmission think zinc. Proc Natl Acad Sci U S A 94(25):13386–13387
Hurrell R (1999) Iron. In: Hurrell R (ed) The mineral fortification of foods. International Ltd, Leatherhead/Surrey, pp 54–93
Hurrell R, Ranum P, de Pee S, Biebinger R, Hulthen L, Johnson Q, Lynch S (2010) Revised recommendations for iron fortification of wheat flour and an evaluation of the expected impact of current national wheat flour fortification programs. Food Nutr Bull 31S:7–21
ICMR (1964) Lathyrism -a preventable paralysis. Indian Council of Medical Research, New Delhi
Iyer V, Salunkhe DK, Sathe SK, Rockland LB (1980) Quick cooking beans (Phaseolus vulgaris L.) II Phytates, oligosaccharides and anti-enzymes. Plant Foods Hum Nutr 30:45–49
Iyer V, Kadam SS, Salunkhe DK (1989) Cooking. In: Salunkhe DK, Kadam SS (eds) Handbook of world food legumes- nutritional chemistry, processing technology and utilization, vol III. CRC Press, Boca Raton, pp 141–163
Jaffe WG, Moreneo R, Wallis V (1973) Amylase inhibitors in legume seeds. Nutr Rep Int 7:169–174
Kalpanadevi V, Mohan VR (2013) Effect of processing on antinutrients and in vitro protein digestibility of the underutilized legume, Vigna unguiculata (L.) Walp subsp. unguiculata. LWT: Food Sci Technol 51(2):455–461. doi:10.1016/j.lwt.2012.09.030
Kennedy AR, Billings PC, Wan XS, Newberne PM (2002) Effects of Bowman Birk inhibitor on rat colon carcinogenesis. Nutr Cancer 43:174–186
Kerem Z, GermanShashoua H, Yarden O (2005) Microwave assisted extraction of bioactive saponins from chickpea (Cicer arietinum L.). J Sci Food Agric 85:406–412
Khan KS, Wojdyla D, Say L, Gulmezoglu AM, Van Look PF (2006) WHO analysis of causes of maternal death a systematic review. Lancet 367:1066–1074
Koratkar R, Rao AV (1997) Effect of soya bean saponins on azoxymethane-induced preneoplastic lesions in the colon of mice. Nutr Cancer 27:206–209
Krishnaswamy K (2001) Perspectives on nutrition needs for the new millennium for South Asian regions. Biomed Environ Sci 14:66–74
La Frano MR, de Moura FF, Boy E, Lonnerdal B, Burri BJ (2014) Bioavailability of iron, zinc, and provitamin A carotenoids in biofortified staple crops. Nutr Rev 72(5):289–307
Ladipo OA (2000) Nutrition in pregnancy: mineral and vitamin supplements. Am J Clin Nutr 72(1):280S–290S
Lajolo FM, Genovese MI, Pryme IF, Dale M (2004) Beneficial (antiproliferative) effects of different substances. In: Muzquiz M, Hill GD, Pedrosa MM, Burbano C (eds) Proceedings of the fourth international workshop on antinutritional factors in legume seeds and oilseeds, EAAP Pubn #110, Wageningen, pp 123–135
Leung BM, Kaplan BJ (2009) Perinatal depression: prevalence, risks, and the nutrition link- a review of the literature. J Am Diet Assoc 109(9):1566–1575. doi:10.1016/j.jada.2009.06.368
Lynch SR (2011) Why nutritional iron deficiency persists as a worldwide problem. J Nutr. doi:10.3945/jn.110.130609
McCance RA, Edgecombe CN, Widdowson EM (1943) Phytic acid and iron absorption. Lancet 2:126–128
McClafferty B, Islam Y (2008) Fighting the hidden hunger. In TCE, Feb 2008, p 27
Miller V, Swaney S, Deinard A (1985) Impact of the WIC program on the iron status of infants. Paediatrics 75:100–105
Mulimani VH, Rudrappa G, Supriya D (1994) α-Amylase inhibitors in chick pea (Cicer arietinum L.). J Sci Food Agric 64:413–415
Muzquiz M, Burbano C, Cuadrado C, Martin M (2001) Analytical methods for determination of compounds with no nutritive value. In: Jacobsen HJ, Muzquiz M, Hassa A (eds) Handbook on common bean related laboratory methods. Galicia, Spain, pp 11–26
Nestel P, Bouis HE, Meenakshi JV, Pfeiffer W (2006) Biofortification of staple food crops. J Nutr 136:1064–1067
Paine JA, Shipton CA, Chaggar S, Howells RM, Kennedy MJ, Vernon G, Wright SY, Hinchliffe E, Adams JL (2005) A new version of golden rice with increased pro-vitamin A content. Nat Biotechnol 23:482–487
Palma S, Perez-Iglesias R, Prieto D, Pardo R, Llorca J, Delgado-Rodriguez M (2008) Iron but not folic acid supplementation reduces the risk of low birth weight in pregnant women without anaemia- a case control study. J Epidemiol Community Health 62(2):120–124
Price KR, Johnson IT, Fenwick RG (1987) The chemistry and biological significance of saponins in foods and feeding stuff CRC. Crit Rev Food Sci Nutr 26:27–131
Price KR, Eagles J, Fenwick GR (1988) Saponin composition of 13 varieties of legume seed using fast atom bombardment mass spectrometry. J Sci Food Agric 42:183–193
Pusztai A, Bardocz S, MartinCabrejas M (2004) The mode of action of ANFs on the gastrointestinal tract and its microflora. In: Muzquiz M, Hill GD, Pedrosa MM, Burbano C (eds) Proceedings of the fourth international workshop on antinutritional factors in legume seeds and oilseeds, EAAP Publication No. 110, Wageningen, pp 87–100
Rao PU, Deostale YG (1982) Tannin content of pulses varietal differences and effects of germination and cooking. J Sci Food Agric 33:1013–1016
Reddy NR, Sathe SK, Salunkhe DK (1982) Phytates in legumes and cereals. In: Chichester CO (ed) Advances in food research, vol 28. Academic Press, Inc, New York, pp 1–4
Reddy NR, Pierson MD, Sathe SK, Salunkhe DK (1985) Dry bean tannins a review of nutritional implications. J Am Oil Chem Soc 62(3):541–549
Reddy MB, Hurrell RF, Cook JD (2000) Estimation of nonheme-iron bioavailability from meal composition. Am J Clin Nutr 71:937–943
Ruiz RG, Price K, Rose M, Rhodes M, Fenwick R (1996) A preliminary study on the effect of germination on saponin content and composition of lentils and chick peas. Z Lebensm Unters Forsch 203(4):366–369. doi:10.1007/BF01231075
Ruiz RG, Price KR, Fenwick RG, Rhodes MJC (1997) Changes in saponin content and composition of chickpeas and lentils during germination and cooking. In: Bardocz S, Muzquiz M, Pusztai A (eds) Effects of antinutrients on the nutritional value of legume diets, proceedings of the fourth scientific workshop, Official Publication of European Communities, Luxembourg, pp 15–18
Scholl TO (2005) Iron status during pregnancy: setting the stage for mother and infant. Am J Clin Nutr 81:S1218–S1222
Sharma S, Goyal R, Barwal S (2013) Domestic processing effects on physicochemical, nutritional and anti-nutritional attributes in soybean (Glycine max L. Merill). Int Food Res J 20:3203–3209
Shibuya M, Hoshino M, Katsube Y, Hayashi H, Kushiro T, Ebizuka Y (2006) Identification of bamyrin and sophoradiol 2,4-hydroxylase by expressed sequence tag mining and functional expression assay. FEBS J 273:948–959
Siddiq EA (1995) Lathyrus genetic resources in Asia. In: Proceedings of the regional workshop on Lathyrus, Raipur, 27–29 Dec, pp 1–2
Singh U, Kherdekar MS, Jambunathan R (1982) Studies on desi and kabuli chickpea (Cicer arietinum L.) cultivars. The levels of amylase inhibitors, levels of oligosaccharides and in vitro starch digestibility. J Food Sci 47:510–512
Srivastava RP, Singh J (2013) Soaking and dehusking effect on lathyrus (Lathyrus sativus) genotypes. In: Proceedings of the international conference on role of plant biochemistry and biotechnology in food and nutritional security, 11–14 Dec 2013, Sri Venkateswara University, Tirupati
Srivastava RP, Srivastava GK (2002) Nutritional quality of newly developed toxin free lathyrus. Indian J Agric Biochem 15:33–36
Srivastava RP, Vasishtha H (2012) Saponins and lectins of Indian chickpeas (Cicer arietinum) and lentils (Lens culinaris). Indian J Agric Biochem 25:44–47
Srivastava RP, Vasishtha H (2013a) Dehusking and cooking effect on dietary fibre, soluble protein and lectin of lentils (Lens culinaris). Indian J Agric Biochem 26:36–40
Srivastava RP, Vasishtha H (2013b) Dietary fiber, protein and lectin contents of lentils (Lens culinaris) on soaking and cooking. Curr Adv Agric Sci 5(2):238–241
Srivastava RP, Vasishtha H (2014) Changes in protein, sapogenols and lectins of different types of chickpeas (Cicer arietinum) during soaking and germination. Indian J Agric Biochem 27:116–122
Stoltzfus RJ (2001) Defining iron deficiency anemia in public health terms; a time for reflection. J Nutr 131:565S–567S
Striepler M, Cohn DF, Hirano A, Schujaman E (1978) The central nervous system in a case of neurolathyrism. Neurology 27:1176
Tako E, Hoekenga OA, Kochian LV, Glahn RP (2013) High bioavailability iron maize (Zea mays L.) developed through molecular breeding provides more absorbable iron in vitro (Caco-2 model) and in vivo (Gallus gallus). Nutr J 12:3. doi:10.1186/1475-2891-12-3
Tava A, Oleszek W, Jurzysta M, Berardo N, Odoardi M (1993) Alfalfa saponins and sapogenins: isolation and quantification in two different cultivars. Phytochem Anal 4:269–274
Thompson LU (1993) Potential health benefits and problems associated with antinutrients in foods. Food Res Int 26:131–149
Titaley CR, Dibley MJ, Roberts CL, Hall J, Agho K (2010) Iron and folic acid supplements and reduced early neonatal deaths in Indonesia. Bull World Health Organ 88:500–508
Tontisirin K, Nantel G, Bhattacharjee L (2002) Food-based strategies to meet the challenges of micronutrient malnutrition in the developing world. Proc Nutr Soc 61(2):243–250
Urbano G, LopezJurado M, Aranda P, VidalValverde C, Tenorio EJ, Porres E (2000) The role of phytic acid in legumes: antinutrient or beneficial function. J Physiol Biochem 56:283–294
Van Jaarsveld PJ, Faber M, Tanumihardjo SA, Nestel P, Lombard CJ, Benade AJS (2005) β-Carotene-rich orange-fleshed sweet potato improves the vitamin A status of primary school children assessed by the modified-relative-dose-response test. Am J Clin Nutr 81:1080–1087
Vasishtha H (2011) Processing effect on hypocholesterolemic and haemagglutinating characters of important pulses. PhD thesis, CSJM University, Kanpur
Vasishtha H, Srivastava RP (2011) Effect of dehusking and cooking on sapogenols of pigeonpea (Cajanus cajan) genotypes. Indian J Agric Biochem 24:60–64
Vasishtha H, Srivastava RP (2014) Effect of soaking and cooking on dietary fiber, protein and lectins of rajmash (Phaseolus vulgaris) beans. Indian J Agric Biochem 27:219–222
Vasishtha H, Srivastava RP, Verma P (2012) Milling and cooking effect on dietary fiber, protein and lectins of pigeonpea (Cajanus cajan). Curr Adv Agric Sci 4:134–138
Whitaker JR (1988) α-Amylase inhibitors of higher plants and microorganisms. In: Kinsella JE, Soucle WG (eds) Food proteins. Proceedings of the protein. Co-products symposium, American Oil Chemists’ Society, Champaign II, pp 354–380
Widdowson EM, McCance RA (1942) Iron exchanges of adults on white and brown bread diets. Lancet 1:588–591
Wikipedia. Biofortification. http://en.wikipedia.org/wiki/biofortification. Accessed 15 Jun 2015
Wilson RD, Davies G, Desilets V, Reid GJ, Summers A, Wyatt P, Young D (2003) The use of folic acid for the prevention of neural tube defects and other congenital anomalies. J Obstet Gynaecol Can 25(11):959–973
Yip R (2002) Prevention and control of iron deficiency policy and strategy issues. J Nutr 132:S802–S805
Zeng L, Cheng Y, Dang S, Yan H, Dibley MJ, Chang S, Kong L (2008) Impact of micronutrient supplementation during pregnancy on birth weight, duration of gestation, and perinatal mortality in rural western China: double blind cluster randomised controlled trial. Br Med J 337:2001–2005
Zhou JR, Erdman JW (1995) Phytic acid in health and disease. Crit Rev Food Sci Nutr 35:495–508
Zimmermann MB, Hurrell RF (2007) Nutritional iron deficiency. Lancet 370:511–520
Zimmermann R, Qaim M (2004) Potential health benefits of golden rice: a Philippine case study. Food Policy 29(2):147–168
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer India
About this chapter
Cite this chapter
Srivastava, R.P. (2016). Antinutrients Restraining Biofortification. In: Singh, U., Praharaj, C., Singh, S., Singh, N. (eds) Biofortification of Food Crops. Springer, New Delhi. https://doi.org/10.1007/978-81-322-2716-8_24
Download citation
DOI: https://doi.org/10.1007/978-81-322-2716-8_24
Published:
Publisher Name: Springer, New Delhi
Print ISBN: 978-81-322-2714-4
Online ISBN: 978-81-322-2716-8
eBook Packages: Chemistry and Materials ScienceChemistry and Material Science (R0)