
Abstract
The existence and potential importance of the soil seed bank have been recognized by ecologists and evolutionary biologists since the dawn of modern biology, from Darwin (1859) to Mall and Singh (2011) and Hong et al. (2012). The earlier studies of soil seed banks began in 1859 with Darwin, when he observed the emergence of seedlings using soil samples from the bottom of a lake. However, the first paper published as a scientific research report was written by Putersen in 1882, studying the occurrence of seeds at different soil depths (Roberts 1981). Very early ecologists started to investigate the nature and the density of living seeds in the soil and the soil seed bank (Darwin 1859; Chippindale and Milton 1934; Nordhagen 1937; Bannister 1966; Barclay-Estrup and Gimingham 1975), and in modern times to determine the significance of soil seeds in the regeneration of different plant communities (Thompson and Grime 1979; Roberts 1981; Mallik et al. 1984; Simpson et al. 1989; Thompson et al. 1997; Miller and Cummins 2001; Lemenih and Teketay 2006; Tessema et al. 2011b; Mall and Singh 2001; Hong et al. 2012) and the similarity between the soil seed bank and aboveground vegetation (Tessema et al. 2011b). A soil seed bank, which begins at dispersal and ends with the germination or death of the seed (Walck et al. 2005), is a reserve of mature viable seeds located on the soil surface or buried in the soil (Roberts 1981) that provides a memory of past vegetation and represents the structure of future populations (Fisher et al. 2009). Seeds are a crucial and integral part of an ecosystem that show the past history of standing vegetation and its future deviation. An understanding of the population dynamics of buried viable seeds is of practical importance in conservation of different communities and weed management in agriculture (Fenner 1985; Fenner and Thompson 2005). The balance between trees and grasses, however, is often highly disturbed as a consequence of heavy grazing and poor management (Pugnaire and Lazaro 2000). This study aimed to gain a better understanding of soil seed bank dynamics in different ecosystems of the world. All plants establish themselves by the expansion and subsequent fragmentation of vegetative parts such as tillers, rhizomes, or runners by the successful establishment of a soil seed bank or bulbils (Freedman et al. 1982). During the past decade, there has been a rapid increase of the number of studies assessing seed density and species richness and the composition of soil seed banks in a wide range of plant communities (Thompson et al. 1997). In India, the soil seed bank has been estimated in humid tropical forest (Chandrashekara and Ramakrishnan 1993), grasslands, irrigated and dry land agro-ecosystems (Srivastava 2002), tropical dry forest (Khare 2006), jhum cultivation (Saxena and Ramakrishnan 1984; Sahoo 1996), Himalayan moist temperate forest (Viswanth et al. 2006), and wastelands and roadsides (Yadav and Tripathi 1981).
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
History
The existence and potential importance of the soil seed bank have been recognized by ecologists and evolutionary biologists since the dawn of modern biology, from Darwin (1859) to Mall and Singh (2011) and Hong et al. (2012). The earlier studies of soil seed banks began in 1859 with Darwin, when he observed the emergence of seedlings using soil samples from the bottom of a lake. However, the first paper published as a scientific research report was written by Putersen in 1882, studying the occurrence of seeds at different soil depths (Roberts 1981). Very early ecologists started to investigate the nature and the density of living seeds in the soil and the soil seed bank (Darwin 1859; Chippindale and Milton 1934; Nordhagen 1937; Bannister 1966; Barclay-Estrup and Gimingham 1975), and in modern times to determine the significance of soil seeds in the regeneration of different plant communities (Thompson and Grime 1979; Roberts 1981; Mallik et al. 1984; Simpson et al. 1989; Thompson et al. 1997; Miller and Cummins 2001; Lemenih and Teketay 2006; Tessema et al. 2011b; Mall and Singh 2001; Hong et al. 2012) and the similarity between the soil seed bank and aboveground vegetation (Tessema et al. 2011b). A soil seed bank, which begins at dispersal and ends with the germination or death of the seed (Walck et al. 2005), is a reserve of mature viable seeds located on the soil surface or buried in the soil (Roberts 1981) that provides a memory of past vegetation and represents the structure of future populations (Fisher et al. 2009). Seeds are a crucial and integral part of an ecosystem that show the past history of standing vegetation and its future deviation. An understanding of the population dynamics of buried viable seeds is of practical importance in conservation of different communities and weed management in agriculture (Fenner 1985; Fenner and Thompson 2005). The balance between trees and grasses, however, is often highly disturbed as a consequence of heavy grazing and poor management (Pugnaire and Lazaro 2000). This study aimed to gain a better understanding of soil seed bank dynamics in different ecosystems of the world. All plants establish themselves by the expansion and subsequent fragmentation of vegetative parts such as tillers, rhizomes, or runners by the successful establishment of a soil seed bank or bulbils (Freedman et al. 1982). During the past decade, there has been a rapid increase of the number of studies assessing seed density and species richness and the composition of soil seed banks in a wide range of plant communities (Thompson et al. 1997). In India, the soil seed bank has been estimated in humid tropical forest (Chandrashekara and Ramakrishnan1993), grasslands, irrigated and dry land agro-ecosystems (Srivastava 2002), tropical dry forest (Khare 2006), jhum cultivation (Saxena and Ramakrishnan 1984; Sahoo 1996), Himalayan moist temperate forest (Viswanth et al. 2006), and wastelands and roadsides (Yadav and Tripathi 1981).
Dynamics of a Soil Seed Bank
The soil seed bank is defined as seeds at or beneath the soil surface that are capable of germinating. Soil seed banks are important in various ecosystems where grasses, forbs, and weeds account for a large part of the vegetation for both annual and perennial species. The soil seed bank describes the composition, diversity, and density of the seed bank. The study of the soil seed bank is important because the bank influences seed reserves in the soil for current, past, and future vegetation. It is the term for the viable seeds that are present in the topsoil (Roberts 1981). All the viable seeds present in soil or with soil debris constitute the soil seed bank (Simpson et al. 1989). Soil seed banks include all seeds buried in the soil and those on the soil surface. Seed banks are essential to maintaining life and growth in different habitats such as grasslands, agro-ecosystems, savannas, desert, wetland, sand dunes, ecotones, plantations, and forests. The seed bank of an agro-ecosystem would be less diverse than the seed bank of a forest. Studies of soil seed banks are of relatively recent origin considering their importance as a source of diversity and continued occupation. The seed banks are the source of genetic material or evolutionary memory (Harper 1977).
Soil seed banks have importance in all types of vegetation. Seed banks can reflect evolutionary changes in plant communities as a result of changes in land use: they provide knowledge of the size and composition of species and predict future vegetation. A seed bank may be defined as a buildup of viable but ungerminated seeds in or on the soil. It is “an aggregation of ungerminated seeds potentially capable of replacing adult plants that may be annuals, dying a natural or unnatural death, or perennials, susceptible to death by disease, disturbances, or consumption by animals including man” (Baker 1989). The second part of this definition, that is, the potential for replacing adult plants, is essential. If seeds are permanently buried too deeply, they fail to be an effective seed bank; the same is true for some seeds in aerial portions of plants. It is the reservoir of viable seeds or of vegetative propagules that are present in the soil and that is able to recompose natural vegetation. The seeds of pioneer species are commonly present in tropical forest soils, particularly in secondary forests (Cao et al. 2000).
The work of Thompson and Grime (1979) reflects fall and spring germination periods and degree of persistence. They defined four types of soil seed banks for herbaceous species, which fell into transient and persistent categories. Tropical soil seed banks show even more diversified strategies. Garwood (1989) described five soil seed bank strategies from the tropics: transient, persistent, pseudo-persistent, seasonal transient, and delayed transient seed banks: these arise from the more complex reproductive phenology of tropical plants and germination patterns of tropical seeds. A diversity of alternative patterns is possible, such as the pseudo-persistent seed banks found in tropical forests and cold deserts where continued dispersal guarantees the presence of a soil seed bank. Many studies suggest that seed banks persist in disturbance regimes, but persistent seed banks are not able to maintain their populations and regenerate without the environmental changes that are caused by disturbances. Annuals generally have persistent seed banks more often than other life forms; in annuals, the persistent stage is the seed only, but even in harsh environments not all annuals produce a persistent seed bank. In some communities perennial woody species produce large seed banks. Generally, small seeds have greater longevity in the seed banks (Harper 1977). The soil seed bank is the cause of persistence of annual species, but for perennials, there is a bank of vegetative propagules such as tubers, rhizomes, and stolons (Fernandez-Quintanilla et al. 1991). The revegetation of plants in the standing vegetation of any ecosystem after any disturbance to the vegetation often depends on the persistence of seeds in the soil (Bakker et al. 1996). The seed bank types in a plant community often determine how the plant community will react to disturbance. Thus, an understanding of persistent seed banks is the key to many aspects of the practical management of weeds in agricultural fields and in conservation of different ecosystems such as grassland, plantation, natural forests, and deserts. Seed banks are classified as temporary or persistent. Temporary seed banks are composed of seeds with a short life that do not enter dormancy and are dispersed for short periods of time during the year (Garwood1989). Persistent seed banks are composed of seeds that have more than 1 year of age, and reserves of these seeds remain in the soil year after year, buried in the soil. Seeds of species that form seed banks must be viable for long periods of time. Seeds persist in the soil far longer at high altitudes, but this can be because low temperatures prevent seed germination, or perhaps conditions found at high altitudes may favor seed survival. Persistent seed banks are the product of large seed crops produced in particularly favorable years, and the spatial pattern of seeds reflects the spatial pattern of seed production.
A number of environmental conditions influence seed bank dynamics via influence on seed production or other stages of the life cycle. Variations in soil conditions also affect the development and size of seed banks. Different soil parameters according to season affect the germination of seeds from soil seed banks and influence species composition in aboveground vegetation. Annuals generally germinate in a soil seed bank because perennial grasses often propagate vegetatively (O’Connor 1996). This pattern is derived by differences in the phenology of perennial versus annual species resulting from late-season disturbances: 98 % of the native species in a plant community are perennial, so by managing perennial species, native species can be maintained. The seed bank reflects the historical process of the plant life cycle from its establishment in the environment to its distribution in time and space. The seed bank is the store of seeds buried in the soil, composed of seed produced on site and seed moved (dispersed) into the area. The dormant seed banks are seeds awaiting stimuli or the right conditions before germination. Active seed banks are seeds in a temporary stage, requiring only favorable temperature and moisture to germinate. Dispersed seed with simple germination whose stimulus requirements have already been met is seed recruited from the dormant seed bank. Different soil depths that provide favorable soil moisture or humidity for seed germination determine recruitment and plant community structure. Environmental variables as well as the standing vegetation of a site can induce or inhibit seed germination from the soil seed bank (Bueno and Baruch 2011). Canopy species modify understory conditions by altering water and nutrient availability, thus modifying the microclimate, and by determining the quality of the litter that covers the soil (Godefroid et al. 2006). Microenvironmental changes following gap creation allow seed germination, seedling establishment, and sapling recruitment (Cao et al. 2000). Graham and Hopkins (1990) predicted that any forest development tending to increase areas of persistent high light intensity would allow the invasion and reproduction of weeds.
Different fertilizers affect the size (Barberi et al. 1998), diversity (Boguzas et al. 2004), and community structure of a seed bank (Davis et al. 2005a). These changes in diversity are related to the productivity and stability of the ecosystem (Zhang et al. 2004a; Zhang et al. 2004b; Xiang et al. 2006). The increasing amounts of nitrogen and other different fertilizers in the soil may affect the composition and dynamics of the soil seed bank, leading to a decrease in species richness. In most seed bank studies a number of species were detected in the seed banks that were not seen in the vegetation (Fig. 3.1).

Flow chart of soil seed bank dynamics (From Harper 1977)
Vertical Distribution and Longevity of Seeds
Vertical distribution of seed banks shows that majority of seeds in grasslands (and probably no-till agricultural fields) are located in the upper 1 in. (2 cm) of the soil profile; nearly the entire seed bank is in the upper 10 cm. The majority of seeds in cultivated soils are in the upper 15 cm of the soil profile and can be found as deep as the soil is tilled; as the intensity of tillage declines, the seed bank moves closer to the soil surface. Many physical and biological factors affect the vertical distribution of seeds in the soil, as most seeds are found near the soil surface and deeply buried seeds rarely germinate from the soil. Long-lived seeds are characteristic of disturbed habitats. Most long-lived seeds are annuals or biennials; biennials are especially prevalent in soil samples taken from dated archaeological sites. Small seeds tend to have much longer soil lives than large ones; very large seeds have very short soil lives. Longevity depends on species, depth of seed burial, soil type, tillage, crop rotations, etc. Seed longevity increases with depth of burial. Many seeds in the soil decay or are lost because of pathogenic soil microflora and predation. Seedling germination is greater in cultivated fields than undisturbed soils; cultivation reduces the soil seed bank more rapidly than in undisturbed soil. Seedling germination increases with decreasing soil burial depth.
Seed Rain
Seed rain represents an historical record of the past vegetation that grew on or near the area. The population dynamics of aboveground plants can be strongly influenced by vegetative reproduction (via rhizomes, new aboveground shoots, or other organs, e.g., bud bank) (Benson et al. 2004) and by sexual reproduction (seed rain and seed bank) (Bakker et al. 1996). Therefore, studies of the seed rain, seed bank, and bud bank are of crucial importance in understanding the regeneration of plant communities after disturbance events and the consequent increase of plant diversity. Seed dispersal can have an impact on species composition (Matthiessen and Hillebrand 2006), forest diversity (Janzen 1970), and the dynamics of plant communities (Nathan and Ne’eman 2004) and can determine spatial ranges of population regeneration (Levine and Murrell 2003). For seed rain study, seed traps are placed inside and outside the gaps. Late-season seed rain has a greater proportion of perennial species and native species in comparison to early-season seed rain. The term seed rain refers to the process by which seeds enter the seed bank. Seeds either generated and produced on site or carried to the site by a dispersal agent become incorporated into the soil. These patterns arise because species that successfully form persistent seed banks are the species with greatest seed longevity. It is one of the two contrasting strategies plants may employ to realize success in replacing themselves and ensuring maintenance of the species. A plant dispersing its seed is most successful, evolutionarily speaking, if it can place its seed in an environment suitable for germination and growth. Such safe sites, as they are called, are generally rare in natural landscapes. To find them, seeds must be dispersed widely in space (to increase the probability of landing in a rare safe site) or widely in time (to increase the probability the seed will survive long enough for a safe site to materialize). Plants with high seed longevity have evolved the strategy of waiting patiently for the right time. If seeds can remain viable for many years, and new seeds continue to “rain” down, it is easy to see where the term seed bank comes from, as the seeds accumulate over time and form a reserve of seeds in the soil. Seed rain decreases with increasing altitude but seed bank density changes slightly. The establishment of target species will depend on seed dispersal (Bossuyt and Honnay 2008). Thus, seed rain or seed dispersal is important for restoration of an ecosystem.
Dynamics of a Weed Seed Bank in an Agro-Ecosystem
A soil seed bank present in an agro-ecosystem is related to weed studies of the so-called weed seed bank. Weed species resist in several adverse climatic conditions, tolerating high and low temperatures, dry and humid environments, and variations in oxygen supply (Christoffoleti and Caetano 1998). The weed seed bank has been studied more intensely than other seed banks because of its economical importance. In agro-ecosystems, the soil seed bank is closely related to weed studies. Its determination allows building models of population establishment through time, making possible the definition of weed control programs (Martins and Silva 1994). Knowledge of the emergence rate of the different species from a soil seed bank in these areas can be applied to soil and crop management programs, which can result in a rational use of herbicides (Voll et al. 1996). Weed seed banks are reserves of viable seeds present on the surface and in the soil. The seed bank consists of new seeds recently shed by a weed plant as well as older seeds that have persisted in the soil for several years. The weed seed bank is the main source of weeds in agricultural fields. Most weeds start their life cycle from a single seed in the soil. If these weeds escape control, they grow and produce thousands of seeds, depending on the species. These seeds are returned to the soil seed bank and become the source of future weed populations. Therefore, knowledge of seed return and seed bank dynamics can help in future weed management. There are enormous numbers of viable weed seeds in the soil. Although a great number of the buried seeds die within a few years, seeds of some species can remain viable for decades. It has been estimated that only 1–9 % of the viable seeds produced in a given year develop into seedlings; the rest remain viable and will germinate in subsequent years, depending on the depth of their burial. Seeds are dispersed both horizontally and vertically in the soil profile. It has been reported that the majority (approximately 95 %) of the seeds entering the seed bank are from annual weeds; only about 4 % come from perennial weeds. Seed bank input is determined by seed rain from the weed plants. In other words, seed bank input is the number of seeds produced and shed by the plant. Although the horizontal distribution of weed seeds in the seed bank generally follows the direction of crop rows, the type of tillage is the main factor determining the vertical distribution of weed seeds within the soil profile.
The weed seed bank is the reserve of viable weed seeds present on the soil surface and scattered throughout the soil profile. It consists of both new weed seeds recently shed and older seeds that have persisted in the soil from previous years. In practice, a weed seed bank also includes the tubers, bulbs, rhizomes, and other vegetative structures through which some of our most serious perennial weeds propagate themselves. In the following discussion, the term “weed seed bank” is defined as the sum of viable weed seeds and vegetative propagules that are present in the soil and thus contribute to weed pressure in future crops. Agricultural soils can contain thousands of weed seeds and a dozen or more vegetative weed propagules per square foot. The weed seed bank serves as a physical history of the past successes and failures of cropping systems, and knowledge of its content (size and species composition) can help producers both anticipate and ameliorate potential impacts of crop–weed competition on crop yield and quality. Eliminating “deposits” to the weed seed bank, which is also called seed rain, is the best approach to ease future weed management. During a 5-year period in Nebraska, broadleaf and grass weed seed banks were reduced to 5 % of their original density when weeds were not allowed to produce seeds. However, in the sixth year, weeds were not controlled and the seed bank density increased to 90 % of the original level (Burnside et al. 1986). Weed seeds can reach the soil surface and become part of the soil seed bank through several processes. The main source of weed seeds in the seed bank is from local matured weeds that set seed. Agricultural weeds can also enter a field on animals, wind, and water, as well as on machinery during activities such as cultivation and harvesting. Weed seeds can have numerous fates after they are dispersed into a field. Some seeds germinate, emerge, grow, and produce more seeds; others germinate and die, decay in the soil, or fall to predation. The seeds and other propagules of most weeds have evolved mechanisms that render a portion of propagules dormant (alive but not able to germinate) or conditionally dormant (depending on soil moisture, temperature, and light) for varying periods of time after they are shed. This adaptation helps the weed survive in a periodically disturbed, inhospitable, and unpredictable environment.
Weed seeds can change from a state of dormancy to nondormancy, in which they can then germinate over a wide range of environmental conditions. Because dormant weed seeds can create future weed problems, weed scientists think of dormancy as a dispersal mechanism through time. Maintaining excellent weed control for several consecutive seasons can eliminate a large majority of the weed seed bank, but a small percentage of viable, highly dormant seeds persist, which can be difficult to eliminate (Egley 1996). Researchers are seeking more effective means to flush out these dormant seeds through multiple stimuli (Egley 1996). Weed species also differ in the seasonal timing of their germination and emergence. Germination of many species is governed by “growing degree-days” (GDD) or the summation of the number of degrees by which each day’s average temperature exceeds a base temperature. This initial or primary dormancy delays emergence until near the beginning of the next growing season – late spring for warm-season weeds (dormancy broken by the cold period over winter) and fall for winter annual weeds (dormancy broken by a hot period in summer) – when emerging weeds have the greatest likelihood of completing their life cycles and setting the next generation of seed.
Management of Weed Seed Bank
Several factors other than mean daily soil temperature have a major impact on the timing of weed germination and emergence in the field. Adequate soil moisture is critical for germination, and good seed–soil contact is also important in facilitating the moisture uptake that is required to initiate the process. In addition, many weed seeds are also stimulated to germinate by light (even the very brief flash occasioned by daytime soil disturbance), fluctuations in temperature and moisture, or increases in oxygen or nitrate nitrogen (N) levels in the soil. Tillage, which exposes seeds to these stimuli, is therefore a critical determinant of seed germination. The timing of N fertilizer applications can also influence the number of weeds germinating (Menalled and Schnobeck 2011). For example, many weed species can be stimulated by large increases in soluble N after incorporation of a legume cover crop or inhibited by delayed applications of N fertilizer (Menalled and Schnobeck 2011). Although the horizontal distribution of weed seeds in the seed bank generally follows the direction of crop rows, type of tillage is the main factor determining the vertical distribution of weed seeds within the soil profile. In plowed fields, the majority of weed seeds are buried 10.16–15.24 cm (below the surface (Cousens and Moss 1990). Under reduced tillage systems such as chisel plowing, approximately 80–90 % of the weed seeds are distributed in the top 4 in. In no-till fields, the majority of weed seeds remain at or near the soil surface. Clements et al. (1996) have shown that soil texture may influence weed seed distribution in the soil profile under these different tillage systems. Understanding the impact of management practices on the vertical distribution of seeds is important because it can help us predict weed emergence patterns. For example, in most soils small-seeded weeds germinate at very shallow depths (less than 0.5 in.). Large-seeded weeds such as the common sunflower have more seed reserves and may germinate from greater depths.
Effect of Tillage on Weed Seed Bank
In no-tillage fields, the majority of weed seeds remain at or near the soil surface. Soil texture may influence weed seed distribution in the soil profile under these different tillage systems. The horizontal distribution of the weed seeds in the seed bank generally follows the direction of crop rows whereas the vertical distribution is influenced by the type of tillage (Menalled and Schnobeck 2011). The greatest diversity of weed species has been observed on field edges and heathlands. In terms of vertical distribution, reports from several states have shown that the majority of weed seeds in a no-till system were located in the top 2 in. of the soil profile. In an annual plow system, seeds were distributed in the upper 12 in. of the soil profile with 25 % of the seed in the upper 0–3 in.. Under reduced tillage systems such as ridge till and chisel plow, 50 % of the weed seeds were located in the upper 3 in. of the soil profile. The size and composition of the seed bank reflects the past and present weed management systems applied in the field and determines the future weed populations. So, it is important to limit current contributions to the weed seed bank for future weed management (Menalled and Schnobeck 2011). The effect of tillage on the weed seed banks will vary by soil type. Understanding the processes that influence the weed seed bank allow us to manipulate and manage weed seed banks effectively by implementing more informed weed management strategies. One strategy is to shift weed seeds from the dormant to the active part of the seed bank: keeping weed seeds on the soil surface and exposing them to harsh environmental conditions and predation can enhance the mortality of the seeds. Weeds can never be eradicated, only managed. The first step toward improving our weed control practices is to understand how tillage can influence the positioning of weed seeds in the soil. Actual seed longevity in the soil depends on an interaction of many factors, including intrinsic dormancy of the seed population, depth of seed burial, frequency of disturbance, environmental conditions (light, moisture, temperature), and biological processes such as predation, allelopathy, and microbial attack (Davis et al. 2005a; Liebman et al. 2001). Understanding how management practices or soil conditions can modify the residence time of viable seeds can help producers minimize future weed problems. For example, seeds of 20 weed species that were mixed into the top 6 in. of soil persisted longer in untilled soil than in soil tilled four times annually (Mohler 2001), which likely reflects greater germination losses in the disturbed treatment. On the other hand, a single tillage can enhance the longevity of recently shed weed seeds, because buried seeds are usually more persistent compared to those left at the surface where they are exposed to predators, certain pathogens, and wide fluctuations of temperature and moisture. However, soil-borne pathogens may also contribute to attrition of buried seeds, even in large-seeded species such as velvetleaf (Davis and Renner 2006) (Fig. 3.2).

A conceptual model of the movements and fates of seed. Phase I dispersal is movement of seed from parent to surface; phase II dispersal includes subsequent horizontal or vertical movements (From Chambers and MacMohan 1994)
Soil Seed Bank and Aboveground Vegetation
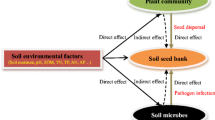
Several studies in the past have addressed similarities between soil seed banks and aboveground vegetation (Leck and Graveline 1979; Henderson et al. 1988; Levassor et al. 1991), although several studies showed poor similarities between species composition of the soil seed bank and the aboveground vegetation (Bakker and Berendse 1999; Lemenih and Teketay 2006). The similarity between seed bank and standing vegetation is expected to decrease with increasing community stability and stress because lack of disturbance creates sites for germination from the seed bank and there is a higher investment in clonal rather than sexual reproduction in stressful conditions (Bekker et al. 1997). Most species typical of stable habitats in contrast do not produce long-lived seeds (Lee 2004). This dissimilarity is the result of frequent occurrence of perennial grasses and woody species in aboveground vegetation (Tessema et al. 2011a) and more annual forbs in the soil seed bank (Solomon et al. 2006; Hopfensperger 2007). Such dissimilarities are caused by species differences in seed dormancy and germination rates (Baskin and Baskin 2004) that result from embryonic dormancy or an impermeable seed coat or both (Baskin and Baskin 2004). The differences in species composition, number of species, and germination success of the soil seed bank down the soil profile might be attributed to differences in soil texture and other soil quality parameters (Hopkins and Graham 1983) under the influence of grazing (Tessema et al. 2011a). Heavy grazing reduces the soil buffer provided by the soil seed bank (Tessema et al. 2011b). The seeds in a tropical soil seed bank were generally near the surface. The similarity between seed bank and vegetation was greater at the site with the more recent history of grazing, and was more dissimilar at the site that had been in public ownership longer. The greatest similarity between seed bank and vegetation was in the ecotonal community, which forms at the interface between prairie and woodland. Similar patterns have been demonstrated for forests. In a study of seed banks in European forests varying in age from young (55–116 years old and established on formerly arable land) to old-growth forest (greater than 250 years), similarity between seed bank and vegetation decreased in the older forests. Species in the seed banks were mainly those typically found along forest edges, in earlier successional stages, or in small disturbances within the forest. This finding makes it clear that minimization of disturbances is imperative for successful management of old-growth forests. Floristic dissimilarity has been observed between soil seed banks and aboveground vegetation generally in various ecosystems of the world (Kellman 1970; Thompson and Grime 1979). Environmental factors that are present around buried viable seeds in soil depths influence their germination and survival (Sakai et al. 2005). Forest and grasslands can be considered as long-term stable ecosystems with low disturbance destroying the vegetation, but in some moist tropical grasslands, a high similarity is found between the seed bank and standing vegetation as a consequence of high seed density, species richness, diversity, and evenness, which may be caused by moderate grazing that creates better soil texture and soil depth rotation. This high similarity may result from the presence of both transient and persistent seed banks, but low similarity is found in moist tropical forests because of high-level germination of weed seed, grasses, and forbs rather than shrubs and woody plants (Mall and Singh, unpublished data). High similarity between the seed bank and aboveground vegetation of any ecosystems of the world produces better recruitment for seed germination.
In tropical forests, there is often little correspondence between the composition of the vegetation and the seed bank (Guevara and Gomez-Pompa 1972; Hall and Swaine 1980; Saulei and Swaine 1988; Hopkins et al. 1990; Teketay and Granstrom 1995), or between the annual seed rain and the seed bank (Uhl and Clark 1983; Saulei and Swaine 1988). The same patterns are common in temperate forests (Livington and Allessio 1968; Enright and Cameron 1988; Matlack and Good 1990; Schiffman and Johnson 1992; Sem and Enright 1996).
Thus, we predict that soil seed banks and weed seed banks can be important tools for conservation: conservation of different ecosystems not only of India but of the world, with the help of vegetative propagation and seed rain, which also provide establishment and recruitment for germination, giving future diversity of standing vegetation and better management of weeds in agro-ecosystems.
Soil Seed Bank and Ecological Significance in Conservation of Different Plant Communities
Conservation ecology is a new paradigm of ecology that not only scientifically contributes to international social movements aiming at maintaining Earth’s biodiversity (Primack 1995) but also is committed to adaptive ecosystem management indispensable to the intergenerational long-term sustainability of mankind (Christensen et al. 1996). Population ecology plays a central role in conservation biology because the decline of local, endemic, or rare species populations and biological invasion by invasive cosmopolitan species both constitute major aspects of local and global biodiversity degradation (Washitani 2001) (Table 3.1).
Soil seed bank studies are of great importance for the understanding of the secondary succession, considered as a necessary first step for the design of ecological restoration plans (Bossuyt and Honnay 2008). The seed bank represents the regeneration potential of the ecosystem and provides the memory of past vegetation, so it can be an important clue for conservation and restoration of plant species. In the near future there is a great role for the soil seed bank in the conservation of plant diversity and vegetation dynamics. At the community scale, seed banks can play a role in local diversity maintenance through temporal storage effects (Sletvold and Rydgren 2007). Because of their implications for population persistence and community resilience, seed banks have also been of interest for conservation research, but some studies conclude that they have little potential for restoration of natural ecosystems (Mitchell et al. 2008). Many studies found lesser species richness and a higher percentage of weed seeds and early-successional species in the seed bank relative to the standing vegetation (Bekker et al. 2000), concluding that seed banks should be a “spillover” of ungerminated seeds rather than an independent driver of population and community dynamics (Bekker et al. 2000). The soil seed bank of an ecosystem serves as an indicator of past and present weed populations: it is the primary source of future seed infestation as the soil seed bank may differ in species composition, richness, and diversity. There has been much work on this topic related to plant diversity conservation for endemic, invasive, and other plant species, whether herbs, shrubs, and trees, of different regions of the world. Restoration should be focused on self-regeneration of native species. The soil seed bank helps in documentation for conservation of different plant communities of the world. Soil seed bank study is more useful these days because it can help in conservation and restoration of vegetation and ecosystems. The soil seed bank is an important component of vegetation dynamics affecting both ecosystem resistance and resilience (Pugnaire and Lazaro 2000). It is a buildup of viable but ungerminated seeds present in or on soil and is essential to maintain life and growth in ecosystems (Baker 1989). It helps in population maintenance and provides information about the size, number, floristic composition, and dynamics of the seeds present in soil (Dalling et al. 1998).
The seed banks contain a generally high proportion of early-successional or invasive nontarget species. The understanding of the dynamics and functions of seed banks has become a great challenge to ecologists of plant communities because this understanding is necessary to determine the role of this community trait in ecosystem functioning and also to improve the integrated management of ecosystems (Luzuriaga et al. 2005). At high grazing pressures, perennial grasses are replaced by annual herbs, which could trigger a vegetation collapse from which recovery to the grassland state is extremely difficult even if grazing pressure is greatly reduced (Rietkerk et al. 1996). The recovery of species that have disappeared from the aboveground vegetation because of heavy grazing, however, can be facilitated by the soil seed banks (Baker 1989). Soil seed bank samples that were collected at the end of the growing season (October–November) after seed production can serve as an indicator of viable seeds that did not germinate in the field during the season. The ecosystem and habitat affect seed bank size. The seed bank of perennial grasses is often quite small because seed production is very small relative to that with annual species. Annual weed species ecosystems have very large seed banks, especially where the land has been grazed: grazing increases seed bank size. Trampling may create conditions in which seeds retain greater viability in the soil. Soil disturbance affects seed bank size: one local or temporary disturbance that allows 1 year of annual weed growth and seed production can have profound impact on the seed bank.
The soil seed bank helps in understanding of the population dynamics of buried viable seeds and is of practical importance in conservation and agriculture (Fenner and Thompson 2005). Few species have seed banks in the tropical forest in general (Garwood 1989), and autochthonous seed banks rarely contribute to the regeneration of deforested tropical forest areas (Teketay and Granstrom 1995). Seeds are available in the soil at the end of the dry season. Collecting the transient seed bank (litter and soil) from the forested area at the end of the dry season and depositing it onto degraded areas seems to be a promising strategy for forest restoration (Sampaio and Scariot, unpublished data). Generally, in plantations that are near harvest their soil seed bank influences the establishment of secondary succession after disturbances. Some of the persistent seeds in the soil and the seed rain (if any) may lead to restoration of the vegetation (Teketay 1998). The soil seed banks of plantations and natural forests are dominated by herbaceous species and mostly exhibit a persistent soil seed bank (Teketay and Granstrom 1995), but few woody species are also observed in a soil seed bank (Teketay and Granstrom 1995; Teketay 1998). A soil seed bank is a key factor for counteracting local extinction of plant species. The ecological implications of a soil seed bank in regeneration of different ecosystems need to be examined by monitoring the dynamics of the bank following cultivation activities (Cao et al. 2000).
In India, much research work has been accomplished in different ecosystems in both dry and moist topics. We compared grasslands and agro-ecosystems, finding a higher seedling emergence from a soil seed bank or weed seed bank in the moist tropics than in the dry tropics (Srivastava 2002).
In this review, in comparison between different ecosystems of temperate and tropical regions we observed that different ecosystems have more seed germination in the tropics than in temperate regions, which may be the result of environmental conditions and high diversity in both soil seed banks and aboveground vegetation. Agro-ecosystems of both regions have great weed seed bank density, so the present study concluded that the weed seed bank should be properly managed to control weeds for better crop yield. We observed that better emergence of seedlings (seed density m−2) provides ideas about the conservation and restoration of grasslands and wetlands, which may be because of the high similarity between aboveground vegetation and the soil seed bank, which comprises grasses, forbs, weeds, shrubs, etc., because of seed rain or seed dispersal. Plantations and natural forest of both tropical and temperate regions have low germination of seeds from the soil seed bank in comparison with other ecosystems but have both high and low similarity, which occurs from the high germination rate of agricultural weeds, annual/perennial grasses, and forbs. Trees provide a low contribution of seed germination from the soil seed bank. Conservation of these ecosystems through the soil seed bank is limited and has less significance in these ecosystems. Thus, conservation and restoration of managed and natural forests should be accomplished by preservation of desirable seeds in situ and ex situ so that their biodiversity may exist in future generations.
References
Amiaud B, Touzard B (2004) The relationship between soil seed bank, aboveground vegetation and disturbances in old embanked marshlands of western France. Flora 199:25–35
Assini S (2001) Seed bank and dynamics of Polygonum hydropiper L. community along Po River (Central-West Italy). Plant Biosyst 135:223–232
Baker HG (1989) Some aspects of the natural history of seed banks. In: Leck MA, Parker VT, Simpson RL (eds) Ecology of soil seed banks. Academic, London, pp 5–19
Bakker JP, Bakker ES, Rosen E, Verweij GL, Bekker R (1996) Soil seed bank composition along a gradient from dry alvar grassland to Juniperus shrubland. J Veg Sci 7:165–176
Bakker JP, Poschlod P, Strystra RJ, Bekker RM, Thompson K (1996) Seed banks and seed dispersal: important topics in restoration ecology. Acta Bot Neerl 45:461–490
Bakker JP, Berendse F (1999) Constraints in the restoration of ecological diversity in grassland and heath land communities. Trends Ecol Evol 14:63–69
Bannister P (1966) Biological flora of British Isles. Erica tetralix. J Ecol 54:795–813
Barberi P, Cozzani A, Macehia M, Bonari E (1998) Size and composition of the weed seed bank under different management systems for continuous maize cropping. Weed Res 38:319–334
Barclay-Estrup P, Gimingham CH (1975) Seed-shedding in heather (Calluna vulgaris (L.) Hull). Trans Bot Soc Edinb 42:275–278
Baskin JM, Baskin CC (2004) A classification system for seed dormancy. Seed Sci Res 14:1–16
Bekker R, Venvveij GL, Smith REN, Reine R, Bakker JP, Schneider S (1997) Soil seed banks in European grasslands: does land use affect regeneration perspectives? J Appl Ecol 34:1293–1310
Bekker RM, Verweij GL, Bakker JP, Fresco LFM (2000) Soil seed bank dynamics in hayfield succession. J Ecol 88:594–607
Benson EJ, Hartnett DC, Mann KH (2004) Belowground bud banks and meristem limitation in tallgrass prairie plant populations. Am J Bot 91:416–421
Benvenuti S (2007) Natural weed seed burial: effect of soil texture, rain and seed characteristics. Seed Sci Res 17:211–219
Boguzas V, Marcinkeviciene A, Kairyte A (2004) Quantitative and qualitative evaluation of weed seed bank in organic farming. Agron Res 1:13–22
Bossuyt B, Honnay O (2008) Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. J Veg Sci 19:875–884
Bueno A, Baruch Z (2011) Soil seed bank and the effect of needle litter layer on seedling emergence in a tropical pine plantation. Int J Trop Biol 59(3): 1071–1079
Burnside OC, Moomaw RS, Roeth FW, Wicks GA, Wilson RC (1986) Weed seed demise in soil in weed-free corn (Zea mays) production across Nebraska. Weed Sci 34:248–251
Cao M, Tang Y, Sheng C, Zhang J (2000) Viable seeds buried in the tropical forest soils of Xishuangbanna, SW China. Seed Sci Res 10:255–264
Chambers JC, MacMohan JA (1994) A day in the life of a seed: movements and fates of seeds and their implications for natural and managed systems. Annu Rev Ecol Syst 25:263–292
Chandrashekara UM, Ramakrishnan PS (1993) Germinable soil seed bank dynamics during the gap phase of a humid tropical forest in the Western Ghats of Kerala, India. J Trop Ecol 9:455–467
Chippindale HG, Milton WE (1934) On the viable seeds present in the soil beneath pastures. J Ecol 22:508–531
Christensen NL, Bartuska AM, Brown JH, Carpenter S, D’Antonio C, Francis R, Franklin J, MacMohan JA, Noss RF, Parsons DJ, Peterson CH, Turner MG, Woodmansee RG (1996) The report of the Ecological Society of America committee on the scientific basis for ecosystem management. Ecol Appl 6:665–691
Christoffoleti PJ, Caetano RSX (1998) Soil seed banks. Sci Agric 55:74–78
Clements DR, Benoit DL, Swanton CJ (1996) Tillage effects on weed seed return and seed bank composition. Weed Sci 44:314–322
Cousens R, Moss SR (1990) A model of the effects of cultivation on the vertical distribution of weed seeds within the soil. Weed Res 30:61–70
Dalling SW, Denslow JS (1998) Soil seed bank composition along a forest chronosequence in a seasonally moist tropical forest in Panama. J Veg Sci 9:669–678
Dalling JW, Swaine MD, Garwood NC (1998) Dispersal patterns and seed bank dynamics of pioneer trees in moist tropical forest. Ecology 79:564–578
Darwin C (1859) The origin of the species by means of natural selection, or the preservation of favoured races in the struggle for life. Murray, London
Davis AS, Renner A (2006) Influence of seed depth and pathogens on fatal germination of velvetleaf (Abutilon theophrasti) and giant foxtail (Setaria faberi). Weed Sci 55:30–35
Davis AS, Cardina J, Forcella F, Johnson GA, Kegode G, Lindquist JL, Lusheri EC, Renner A, Sprague CL, Williams MM (2005a) Environmental factors affecting seed persistence of annual weeds across the U.S. corn belt. Weed Sci 53:860–868
Davis AS, Renner KA, Gross KL (2005b) Weed seed bank and community shifts in a long-term cropping systems experiment. Weed Sci 53:296–306
Decocq G, Valentin B, Toussaint B, Hendoux F, Saguez R, Bardat J (2004) Soil seed bank composition and diversity in a managed deciduous forest. Biodivers Conserv 13:2485–2509
Dougall TAG, Dodd JC (1997) A study of species richness and diversity in seed banks and its use for the environmental mitigation of a proposed holiday village development in a coniferized woodland in south east England. Biodivers Conserv 6:1413–1428
Egley GH (1996) Stimulation of weed seed germination in soil. Rev Weed Sci 2:67–89
Enright NJ, Cameron EK (1988) The soil seed bank of a Kauri (Agathis australis) forest remnant near Auckland, New Zealand. N Z J Bot 26:223–236
Erenler HE, Ashton PA, Gillman MP, Ollerton J (2010) Factors determining species richness of soil seed banks in lowland ancient woodlands. Biodivers Conserv 19:1631–1648
Esmailzadeh O, Hosseini SM, Tabari M (2011) Relationship between soil seed bank and above-ground vegetation of mixed-deciduous temperate forest in Northern Iran. J Agric Sci Technol 13:411–424
Feng W, Pan G, Qiang S, Li R, Wei J (2008) Influence of long term different fertilization on soil seed bank diversity of a paddy soil under rice/rape rotation. Front Biol China 3(3):320–327
Fenner M (1985) Seed ecology. Chapman & Hall, London
Fenner M, Thompson K (2005) The ecology of seeds, 2nd edn. Cambridge University Press, Cambridge
Fernandez-Quintanilla C, Saavedra MS, Garcia TL (1991) Ecologia de las malas hierbas. In: Garcia TL, Fernandez-Quitanilla C (eds) Fundamentos sobre malas hierbas y herbicides. Mundi-Prensa, Madrid, pp 49–69
Fisher JL, Loneragan W, Dixo K, Veneklaas EJ (2009) Soil seed bank compositional change constraints biodiversity in an invaded species-rich woodland. Biol Conserv 142:256–269
Franke AC, Singh S, McRoberts N, Nehra AS, Godara S (2007) Phalaris minor seed bank studies: longevity, seedling emergence and seed production as affected by tillage regime. Weed Res 47:73–83
Freedman B, Hill N, Svoboda J, Henry GHR (1982) Seed banks and seedling occurrence in a high arctic oasis at Alexandra Fjord, Ellesmere Island, Canada. Can J Bot 60:2112–2118
Garcia MA (1995) Relationships between weed community and soil seed bank in a tropical agro-ecosystem. Agric Ecosyst Environ 55:139–146
Garwood NC (1989) Tropical soil seed banks: a review. In: Leck MA, Parker VT, Simpson RL (eds) Ecology of soil seed banks. Academic, San Diego, pp 149–209
Godefroid S, Phartyal S, Koedam N (2006) Depth distribution and composition of seed banks under different tree layers in a managed temperate forest ecosystem. Acta Oecol (Berl) 29:283–292
Graham AW, Hopkins MS (1990) Soil seed banks of adjacent underlogged rain forest types in North Queensland. Aust J Bot 38:261–268
Granstrom A (1988) Seed banks at six open and afforested heathland sites in southern Sweden. J Appl Ecol 25:297–306
Guevara S, Gomez-Pompa A (1972) Seeds from surface soils in a tropical region of Veracruz, Mexico. J Arnold Arboretum 53:312–335
Hall JB, Swaine MD (1980) Seed stocks in Ghanaina forest soils. Biotropica 12:256–263
Harper JK (1977) Plant population biology. Academic, New York
Henderson CB, Petersen KE, Redak RA (1988) Spatial and temporal patterns in the seed bank and vegetation of a desert grassland community. J Ecol 76:717–728
Hong J, Guopeng SL, Zhang Y (2012) Soil seed bank techniques for restoring wetland vegetation diversity in Yeyahu wetland, Beijing. Ecol Eng 42:192–202
Hopfensperger KN (2007) A review of similarity between seed bank and standing vegetation across ecosystems. Oikos 116:1438–1448
Hopkins MS, Graham AW (1983) The species composition of soil seed banks beneath lowland rainforests in North Queensland, Australia. Biotropica 15:90–99
Hopkins MS, Tracey JG, Graham AW (1990) The size and composition of soil seed banks in remnant patches of three structural rainforest types in Queensland. Aust J Ecol 15:43–50
Janzen DH (1970) Herbivores and the number of tree species in tropical forest. Am Nat 104:501–528
Kalamees R, Zobel M (2002) The role of the seed bank in gap regeneration in a calcareous grassland community. Ecology 83:1017–1025
Kalmees R, Pussa K, Zobel K, Zobel M (2012) Restoration potential of the persistent soil seed bank in successional calcareous (alvar) grassland in Estonia. Appl Veg Sci 15:208–218
Kassahun A, Snyman HA, Smit GN (2009) Impact of rangeland degradation on pastoral production systems, livelihoods and perceptions of the Somali pastoralists in Eastern Ethiopia. J Arid Environ 72:1265–1281
Kellman MC (1970) The viable seed content of some forest soil in coastal British Colombia. Can J Bot 48:1383–1385
Kelton JA, Price AJ, Santen EV, Balkcom KS, Arriaga FJ, Shaw JN (2011) Weed seed bank density and composition in a tillage and landscape variability study. Commun Biometry Crop Sci 6(1):21–30
Khare PK (2006) Ecological study of soil seed bank in tropical dry forests of Sagar, Central India. PhD thesis, Dr. Hari Singh Gaur Vishwavidyalya, Sagar, India
Korb JE, Springer JD, Powers SR, Moore MM (2005) Soil seed banks in Pinus ponderosa forests in Arizona: clues to site history and restoration potential. Appl Veg Sci 8:103–112
Landman GB, Kolka RK, Sharitz RR (2007) Soil seed bank analysis of planted and naturally revegetating thermally disturbed wetland forests. Wetlands 27(2):211–223
Leck MA (2003) Seed bank and vegetation development in a created tidal freshwater wetland on the Delaware River, Trenton, New Jersey, USA. Wetlands 23(2):310–343
Leck MA, Graveline KJ (1979) The seed bank of a freshwater tidal marsh. Am J Bot 66:1006–1015
Leckie S, Vellend M, Bell G, Waterway MJ, Lechowicz MJ (2000) The seed bank in an old-growth temperate deciduous forest. Can J Bot 78:181–192
Lee P (2004) The impact of burn intensity from wildfires on seed and vegetative banks and emergent understory in aspen-dominated boreal forests. Can J Bot 82: 1468–1480
Lemenih M, Teketay D (2006) Changes in soil seed bank composition and density following deforestation and subsequent cultivation of a tropical dry Afromontane forest in Ethiopia. Trop Ecol 47:1–12
Levassor C, Ortega M, Peco B (1991) Seed banks dynamics of Mediterranean pastures subjected to mechanical disturbance. J Veg Sci 1:339–344
Levine JM, Murrell DJ (2003) The community-level consequences of seed dispersal patterns. Annu Rev Ecol Evol Syst 34:549–574
Li EH, Liu GH, Li W, Yuan LY, Li SC (2008) The seed-bank of a lakeshore wetland in Lake Honghu: implications for restoration. Plant Ecol 195:69–76
Li SS, Wei SH, Zuo RL, Wei JG, Qiang S (2012) Changes in the weed seed bank over 9 consecutive years of rice-duck farming. Crop Prot 37:42–50
Liebman M, Mohler CL, Staver CP (2001) Ecological management of agricultural weeds. Cambridge University Press, New York
Livington RB, Allessio ML (1968) Buried viable seed in successional fields and forest stands. Harvard Forest, Massachusetts. Bull Torrey Bot Club 95: 58–69
Luzuriaga AL, Escudero A, Olano MJ (2005) Regenerative role of seed banks following an intense soil disturbance. Acta Oecol (Berl) 27:57–66
Mall U, Singh GS (2011) Management and conservation of biodiversity through soil seed bank in moist tropics of India. N Y Sci J 4:30–37
Mallik AU, Hobbs RJ, Legg CJ (1984) Seed dynamics in Calluna–Arctostaphylos heath in north-eastern Scotland. J Ecol 72:855–871
Martins CC, Silva WR (1994) Da estudos de bancos de sementes do solo. Inform Abrates 4:49–56
Matlack GR, Good RE (1990) Spatial heterogeneity in the soil seed bank of a mature Coastal Plain forest. Bull Torrey Bot Club 117:143–152
Matthiessen B, Hillebrand H (2006) Dispersal frequency affects local biomass production by controlling local diversity. Ecol Lett 9:652–662
Matus G, Papp M, Tothmeresz B (2005) Impact of management on vegetation dynamics on seed bank formation of inland dune grassland in Hungary. Flora 200:296–306
Menalled F, Schnobeck M (2011). Manage the weed seed bank: minimize “Deposits” and maximize “Withdrawals”. http://www.extension.org/pages/18527.
Metcalfe DJ, Turner IM (1998) Soil seed bank in Singapore: canopy-gap and litter-gap demanders. J Top Ecol 14:103–108
Miller GR, Cummins RP (2001) Geographic variation in seed-setting by heather (Calluna (L.) Hull) in the Scottish highlands. J Biogeogr 28:1023–1031
Mitchell RJ, Rose RJ, Palmer SCF (2008) Restoration of Calluna vulgaris on grassland-dominated moorlands: the importance of disturbance, grazing and seeding. Biol Conserv 141:2100–2111
Mohler CL (2001) Weed life history: identifying vulnerabilities. In: Liebman M (ed) Ecological management of agricultural weeds. Cambridge University Press, New York, pp 40–98
Mukhongo JN, Kinyamario JI, Chira RM, Musila W (2011) Assessment of soil seed bank from six different vegetation types in Kakamega forest, Western Kenya. Afr J Biotechnol 10(85):14384–14391
Nathan R, Ne’eman G (2004) Spatiotemporal dynamics of recruitment in Aleppo pine (Pinus halepensis Miller). Plant Ecol 171:123–137
Nordhagen R (1937) Studien uber die monotypische Gattung Calluna Salisb. I. Bergens Museums Aarbog. Naturvitenskaplig Rekke 4:1–55
O’Connor TG (1996) Hierarchical control over seedling recruitment of the bunch-grass Themeda triandra in a semi-arid savanna. J Appl Ecol 33:1094–1106
Onaindia M, Amezaga I (2000) Seasonal variation in the seed banks of native woodland and coniferous forest in Northern Spain. For Ecol Manag 126:163–172
Primack RB (1995) A primer of conservation biology. Sinauer, Sunderland
Pugnaire FI, Lazaro R (2000) Seed bank and understory species composition in semi-arid environment: the effect of shrub age and rainfall. Ann Bot 86:807–813
Rietkerk M, Ketner P, Stroosnijder L, Prins HHT (1996) Sahelian rangeland development a catastrophe? J Range Manag 49:512–519
Roberts HA (1981) Seeds banks in soils. Adv Appl Biol 6:1–56
Sahoo UK (1996) Depletion in arable soil seed bank under ‘jhum’ and terrace cultivation in Meghalaya. Seed Res 24(1):20–25
Sakai A, Sato S, Sakai T, Kuramoto S, Tabuchi R (2005) A soil seed bank in mature conifer plantation and establishment of seedlings after clear-cutting in southwest Japan. J For Res 10:295–304
Saulei SM, Swaine MD (1988) Rain forest seed dynamics during succession at Gogol, Papua, New Guinea. J Ecol 76:1133–1152
Saxena KG, Ramakrishnan PS (1984) Herbaceous vegetation development and weed potential in slash and burn agriculture (jhum) in North- Eastern India. Weed Res 24:135–142
Schiffman PM, Johnson WC (1992) Sparse buried seed bank in a Southern Appalachian oak forest: implications for succession. Am Midl Nat 127:258–267
Sem G, Enright NJ (1996) The relationship between seed rain and the soil seed bank in a temperate rainforest stand near Auckland, New Zealand. New Zealand J Bot 34:215–226
Senebeta F, Teketay D (2002) Soil seed banks in plantations and adjacent natural dry Afromontane forests of central and southern Ethiopia. Trop Ecol 43(2):229–242
Senebeta F, Teketay D, Naslund BA (2002) Native woody species regeneration in exotic tree plantations at Munessa-Shashemene forest, southern Ethiopia. New For 24:131–145
Simpson RL, Leck MA, Parker VT (1989) Seed banks: general concepts and methodology issues. In: Leck MA, Parker VT, Simpson RL (eds) Ecology of soil seed banks. Academic, London, pp 3–8
Sjursen H, Brandsæter LO, Seljåsen R (2008) Change in the weed seed bank during the first four years of a five-course crop rotation with organically grown vegetables. In: 16th IFOAM Organic World Congress, Modena
Sletvold N, Rydgren K (2007) Population dynamics in Digitalis purpurea: the interaction of disturbance and seed bank dynamics. J Ecol 95:1346–1359
Solomon TB, Snyman HA, Smit GN (2006) Soil seed bank characteristics in relation to land use systems and distance from water in a semi-arid rangeland of Southern Ethiopia. S Afr J Bot 72:263–271
Srivastava R (2002) Analysis of weed populations and soil seed bank in dryland and irrigated agro-ecosystems in dry tropics. PhD thesis, Banaras Hindu University, Varanasi
Staff H, Jonsson M, Olsen G (1987) Buried germinative seeds in mature beech forests with different herbaceous vegetation and soil types. Holarctic Ecol 10:268–277
Teketay D (1998) Soil seed bank at an abandoned Afromontane arable site. Feddes Rep 109:161–174
Teketay D, Granstrom A (1995) Soil seed banks in dry Afromontane forests of Ethiopia. J Veg Sci 6:777–786
Tessema ZK, De Boer WF, Baars RMT, Prins HHT (2011a) Changes in vegetation structure, herbaceous biomass and soil nutrients in response to grazing in a semi-arid savanna in Ethiopia. J Arid Environ 75:662–670
Tessema ZK, De Boer WF, Baars RMT, Prins HHT (2011b) Influence of grazing on soil seed banks determines the restoration potential of above-ground vegetation in a semi-arid savanna of Ethiopia. Biotropica 2(2):1–4
Thompson K (1986) Small-scale heterogeneity in the seed bank of an acidic grassland. J Ecol 74:733–738
Thompson K, Grime JP (1979) Seasonal variation in the seed bank of herbaceous species on ten contrasting habitats. J Appl Ecol 20:141–156
Thompson K, Bakker JP, Bekker RM (1997) The soil seed banks of North West Europe: methodology, density and longevity. Cambridge University Press, Cambridge
Tu M, Titus JH, Tsuyuzaki S, Moral R (1998) Composition and dynamics of wetland seed banks on Mount St. Helens, Washington, USA. Folia Geobot 32:3–16
Uhl C, Clark K (1983) Seed ecology of selected Amazon Basin successional species. Bot Gaz 144:419–425
Viswanth S, Singh RP, Thapliyal RC (2006) Seed bank dynamics of Buxus wallichiana baillon in a Himalayan moist temperate forest. Trop Ecol 47(1):145–148
Voll E, Gazziero DLP, Karam D (1996) Dinamica de population de Brachiaria plantginea (Link.) Hitch. Sob manejo do solo e de herbicides: 2. Emergencia. Pesq Agropec Bras 30(12):27–35
Walck JL, Baskin JM, Baskin CC, Hidayati SN (2005) Defining transient and persistent seed banks in species with pronounced seasonal dormancy and germination patterns. Seed Sci Res 15:189–196
Wang CH, Tang L, Fei SF, Wang JQ, Gao Y, Wang Q, Chen JK, Li B (2009a) Determinants of seed bank dynamics of two dominant helophytes in a tidal salt marsh. Ecol Eng 35:800–809
Wang J, Ren H, Yang L, Danyan L, Guo Q (2009b) Soil seed banks in four 22-year-old plantations in South China: implications for restoration. For Ecol Manag 258:2000–2006
Wang N, Jiao JY, Jia YF, Zhang X (2011) Soil seed bank composition and distribution on eroded slopes in the hill-gully Loess plateau region (China): influence on natural vegetation colonization. Earth Surf Proc Land 36:1823–1835
Washitani I (2001) Plant conservation ecology for management and restoration of riparian habitats of lowland Japan. Popul Ecol 43:189–195
Weerasinghe UR, Akiko S, Palitha J, Seiki T (2008) The role of the soil seed bank in vegetation recovery on an oceanic island severely damaged by introduced goats. Appl Veg Sci 11(3):355–364
Xiang CG, Zhang PJ, Pan GX, Qiu DS, Chu QH (2006) Changes in diversity, protein content and amino acids composition of earthworms from a paddy soil under long-term different fertilizations in the Tai lake region, China. Acta Ecologica Sinica 26:1667–1674
Yadav AS, Tripathi RS (1981) Population dynamics of the ruderal weed Eupatorium odoratum and its natural regulation. Oikos 36:355–361
Yan RR, Wei ZJ, Xin XP, Liu HM, Yang J, Wuren Q (2012) Effects of the grazing systems on germinable soil seed bank of desert steppe. Sci Cold Arid Reg 4(1):40–45
Zhang QJ, Fu BJ, Chen LD, Zhao WW, Yang QK, Liu GB, Gulinck H (2004a) Dynamics and driving factors of agricultural landscape in the semiarid hilly area of the Loess Plateau, China. Agric Ecosyst Environ 103:535–543
Zhang X, Shao M, Li S, Peng K (2004b) A review of soil and water conservation in China. J Geogr Sci 14:259–274
Zhao L, Cheng J, Wan H (2008) Dynamic analysis of the soil seed bank for grassland in a typical prairie on the Loess Plateau (in Chinese). Bull Soil Water Conserv 28:15–56
Zhao LP, Su JS, Wu GL, Gillet F (2011) Long-term effect of grazing exclusion on above-ground and belowground plant species diversity in a steppe of the Loess Plateau, China. Plant Ecol Evol 144(3):313–320
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer India
About this chapter
Cite this chapter
Mall, U., Singh, G.S. (2014). Soil Seed Bank Dynamics: History and Ecological Significance in Sustainability of Different Ecosystems. In: Fulekar, M., Pathak, B., Kale, R. (eds) Environment and Sustainable Development. Springer, New Delhi. https://doi.org/10.1007/978-81-322-1166-2_3
Download citation
DOI: https://doi.org/10.1007/978-81-322-1166-2_3
Published:
Publisher Name: Springer, New Delhi
Print ISBN: 978-81-322-1165-5
Online ISBN: 978-81-322-1166-2
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)