
Abstract
We developed a conventional imaging method to measure Ca2+ concentration in cytosol (using FuraRed as an indicator) and mitochondria (using Rhod-2 as an indicator), simultaneously, by alternative excitation with specific wave length. After confirming the availability of the method in Hela cells, we applied it to mouse whole-brain slice preparation, which was exposed to oxygen- and glucose-deprived artificial cerebrospinal fluid (ischemic ACSF) for 12 min. The fluorescence (>570 nm) at the cerebral cortex and hippocampus due to FuraRed (excited by 480 ± 10 nm) decreased (indicating the increase in cytosolic Ca2+concentration), while the fluorescence due to Rhod-2 (excited by 560 ± 10 nm) increased (indicating the increase in mitochondrial Ca2+ concentration) during exposure to ischemic conditions. We found the characteristic protective effects of cyclosporine A (10−6 M), a known blocker for mitochondrial permeability transition, and SEA0400 (10−6 M), a blocker for Na+/Ca2+ exchanger, on the abnormal Ca2+ increase in cytosol. We confirmed that the present method will be useful for future pathological and pharmacological studies on ischemia-induced brain damage.
Access provided by Autonomous University of Puebla. Download conference paper PDF
Similar content being viewed by others

Keywords
- Ischemia
- Cytosol
- Mitochondria
- Fluorescence Ca2+ indicator
- Rhod-2
- FuraRed
- Cyclosporine A
- SEA0400
- Na+/Ca2+exchanger
- Mitochondrial permeability transition
Introduction
Brain tissue has been known to require a very large amount of energy. Embolism or infarction of cerebral blood vessels and also accidental hypoxia and hypoglycemia cause encephalopathies, which are sometimes fatal or have severe sequelae depending upon the sites attacked. The mechanisms of acute neuronal cell death due to the energy deficit have been demonstrated to be dependent upon the massive increase in intracellular Ca2+ concentration. The increased Ca2+ subsequently triggers the Ca2+-dependent process to produce superoxides and cascade activation of many kinds of proteases, which cause the severe breakdown of functional molecules inside the cell, cytoskeleton, and plasma membrane [1]. Abnormal Ca2+ concentration increase during ischemia seemed to be one of the most important targets for developing new drugs to protect the brain from the damage due to ischemia. Since we know mitochondria is one of the most important organelles for controlling the intracellular Ca2+ concentration, it seems very important to not only the cytosolic Ca2+ concentration, but also the mitochondrial Ca2+ concentration during and after the ischemia. Nowadays, many kinds of Ca2+ indicators are commercially available. Almost all indicators are known to distribute inside cytosol, but an indicator called Rhod-2, a derivative of rhodamine, has been proven to distribute in mitochondria preferentially [4]. In the present study, we tried to develop a conventional method for simultaneous measurement of cytosolic and mitochondrial Ca2+ concentration and attempted to apply the method for drug evaluation.
Materials and Methods
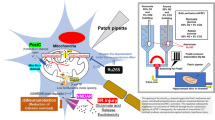
Experimental Procedures for Simultaneous Detection of Cytosolic and Mitochondrial Ca2+ Concentration
The peak excitation wave length of Rhod-2, a mitochondrial Ca2+ indicator, is 560 nm, and the peak fluorescence due to the excitation is observed at around 590 nm. As for an indicator for cytosolic Ca2+ dynamics, we selected FuraRed/AM, which distributes in cytosol [2]. The peak excitation wave length of the dye is around 500 nm, which induces fluorescence with a peak wave length of around 650 nm. We loaded those two types of Ca2+ indicators onto the cells in culture or the mouse brain slices simultaneously. Using a conventional epifluorescence inverted microscope (Olympus IX81) with an automatic excitation filter exchange system, we detected the fluorescence due to those two different indicators with almost simultaneous timing (within 0.1 s of excitation filter exchange) through a dichroic mirror of 570 nm and a long-pass emission filter (>575 nm). The fluorescence images were detected by a highly sensitive digital CCD camera (ORCA ER, Hamamatsu Photonics) and fed into an image analysis system (AQUACOSMOS, Hamamatsu Photonics). The fluorescence signals obtained from two different indicators were analyzed separately and provided as cytosolic and mitochondrial signals respectively.
Procedure for Preparing Cultured Hela Cells and for Loading Ca2+ Indicators
Hela cells are maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10 % fetal bovine serum (FBS) at 37 °C under 5 % CO2 on a glass-bottomed culture dish (IWAKI, 3931-035). Two days later, the cultured cells on the dish were rinsed with the balanced salt solution (BSS), and then exchanged with the BSS containing FuraRed/AM (5 μM) and Rhod-2/AM (5 μM). After 45 min of incubation at room temperature (27 ± 1 °C), the cells were rinsed with fresh BSS and kept for another 30 min at room temperature to allow breakdown of the ester form by the intrinsic esterase. The cells on the glass-bottomed dish were perfused continuously at a rate of 2 mL/min. The fluorescence images of the cells were observed × 40 lens and fed into the above-mentioned image analysis system.
Procedure for Preparing Fresh Mice Brain Slice Preparations and for Loading Ca2+ Indicators
An adult mouse (C57 BK, 20–30 g) was decapitated under deep ether anesthesia. Whole brain was isolated quickly and kept in ice-cold aerobic artificial cerebrospinal fluid (ACSF; bubbled constantly with 95 % O2 and 5 % CO2) for 10 min. Then the 300 μM horizontal brain slices including the hippocampus and cerebral cortex were prepared by a tissue slicer (Dohsaka DTK-1000). The brain slices were placed in the aerated ACSF containing FuraRed/AM (10 μM) and Rhod-2/AM (5 μM) for 60 min, and then in a large volume of fresh ACSF at room temperature for 30 min. The slice preparation was placed on the glass-bottomed recording chamber. The slice was perfused with aerated ACSF at a rate of 2 mL/min at 32 ± 1 °C. The fluorescence images of the slice preparation were detected by × 4 objective lens and fed into the image analysis system.
Protocol for Exposing the Slice Preparation to Ischemia and the Evaluation of Drug Effects
To expose the slice preparation to the ischemic conditions, normal aerobic ACSF was replaced for 12 min by the ischemic ACSF containing 2-deoxy-d-glucose, bubbled with 95 % N2 and 5 % CO2) at 31 ± 1 °C. Drug effects on the slice preparation were evaluated by administration from 10 min before exposure to the end of the experiment.
Drugs Used
SEA0400 was kindly supplied by Taisho Pharmaceutical (Saitama, Tokyo, Japan). Cyclosporine A, ionomycin, rotenone, and histamine were purchased from Sigma-Aldrich (St. Louis, MO, USA). Rhod-2/AM was purchased from Dojindo Molecular Technologies. (Kumamoto, Japan), and FuraRed.AM was purchased from Invitrogen, Molecular probes (Eugene, OR, USA).
Results
Confirmation of Differential Distribution of FuraRed and Rhod-2 in Hela Cells
As shown in Fig. 1a, the cells loaded with FuraRed and Rhod-2 showed different fluorescence images (>570 nm) when excited by 500 and 560 nm, alternately. The fluorescence intensity of the cells obtained by 500-nm excitation was almost homogeneous, while that excited by 560 nm was concentrated in specific filamentous structures in the cell. The Rhod-2 fluorescence images seemed to show the structure of living mitochondria. We applied histamine (10−5 M) to the cells and examined their fluorescence by alternate excitation with 500 and 560 nm. As shown in Fig. 1b, the fluorescence due to 500-nm excitation decreased, which indicated an increase in cytosolic Ca2+ concentration, while the fluorescence due to 560-nm excitation increased, which indicated an increase in mitochondrial Ca2+ concentration.

Fluorescence images and Ca2+ dynamics in Hela cells loaded with both FuraRed/AM and Rhod-2. (A) Differential fluorescence images (>570 nm) of FuraRed and Rhod-2 in the same cells. (B) Pseudo-color images of cytosolic and mitochondrial Ca2+ concentrations. a, b and c indicate before, during, and after histamine (10−5 M) administration. Arrows indicate the direction of color change due to an increase in Ca2+ concentration
We applied ionomycin (10−6 M), a Ca2+ ionophore, for 1 min, and observed a large increase in Ca2+ in both cytosol and mitochondria. In the cells treated with rotenone, a mitochondrial Ca2+ uptake blocker, we were able to observe an increase in cytosolic Ca2+ concentration, while the increase in mitochondria was largely reduced (data not shown).
These results indicate that the present method can be used to detect the specific Ca2+ dynamics in both cytosol and mitochondria simultaneously.
Examination of the Drugs Affecting the Ca2+ Dynamics in Hela Cells
We examined the effects of two types of drugs affecting Ca2+ dynamics. Cyclosporine A (CsA) has been proven to have a strong protective effect on mitochondrial permeability transition [5], which will occur during Ca2+ overloading in mitochondria. Another drug is SEA0400 (SEA), which has been demonstrated to inhibit the activity of the Na+/Ca2+ exchanger in plasma and mitochondrial membrane [3].
The drugs were administered on Hela cells by perfusion 5 min before the exposure to ionomycin (10−6 M) for 1 min, which caused a massive increase in cytosolic and mitochondrial Ca2+. We evaluated the effects of the drugs according to the percentage recovery from the extensive increase in Ca2+ concentration in cytosol and mitochondria 1 min after exposure to ionomycin. As shown in Fig. 2, CsA (0–10−5 M) and SEA (0–10−5 M) showed dose-dependent protective effects on the ionomycin-induced severe Ca2+ increase in both cytosol and mitochondria.

Effects of SEA0400 and cyclosporine A on the ionomycin-induced Ca2+ increase in cytosol and mitochondria in Hela cells. (a) Averaged time courses of the Ca2+ dynamics and the protective effects of SEA0400 (10−6 M) and cyclosporine A on cytosol (upper panel) and mitochondria (lower panel; n = 10, SE indicated). The fluorescence signals in cytosol were reversed to compare them with mitochondria. (b) The dose response relationships to the effects of SEA0400 and cyclosporine A
Application of the Method to Mouse Brain Slices
We applied the method to the mouse brain-slice preparations. Our preliminary experiments showed that the loading of Rhod-2 to the brain-slice preparation was more efficient than FuraRed and that the excitation for FuraRed was more effective at a shorter wave length of 480 nm than in the Hela cell experiments. Thus, we decided the loading concentration for the brain-slice preparation to be 10 μM for FuraRed/AM and 5 μM for Rhod-2/AM. The excitation wave length for FuraRed was fixed at 480 nm.
We confirmed the availability of the method for the brain-slice preparation by administering a high potassium ACSF (50 K; 50 mM, isotonic like normal ACSF by exchanging sodium for potassium). During 5-min administration of 50 K to the slice preparation, we were able to observe the decrease in FuraRed and the increase in Rhod-2 fluorescence, suggesting that our method successfully detected the increase in cytosolic and mitochondrial Ca2+ respectively.
Examination of Ca2+ Dynamics During Ischemia and the Effects of Drugs
Under the above-mentioned experimental conditions, we examined the Ca2+ dynamics in cytosol and mitochondria during exposure to 12 min of ischemia. We were able to observe a characteristic Ca2+ increase in the whole brain during ischemia. Figure 3 shows the changes that occurred in the hippocampal CA1 region. The increase in the cytosolic and mitochondrial Ca2+ concentrations began within 5 min, and developed progressively during exposure to the ischemic conditions. The increase in cytosolic Ca2+ was apparently irreversible, but mitochondrial Ca2+ seemed to be partially reversible. We administered CsA and SEA by pretreatment for 10 min before the onset of ischemia. As shown in Fig. 3, CsA (10−6 M) and SEA (10−6 M) significantly depressed the increase in cytosolic Ca2+ increase during ischemia. CsA seemed to reduce the increase in mitochondrial Ca2+, but the inhibition was not significant.

The time courses of Ca2+ dynamics in cytosol and mitochondria in mouse brain slices because of a 12-min exposure to ischemic conditions, and the effects of SEA0400 and CsA. The time courses of Ca2+ dynamics observed during exposure to ischemia in the hippocampal CA1 region, and the effects of SEA0400 (10−6 M) (a) and CsA (10−6 M) (b). Each graph shows the average time course (n = 5, SE indicated)
Conclusion
In the present study, we established a conventional method of measuring cytosolic and mitochondrial Ca2+ dynamics in culture cells and brain-slice preparation simultaneously. The simultaneous measurement of mitochondria and cytosol using laser confocal microscopy has been reported in isolated pancreatic acinar cells [6]. However, we ventured to develop an image analysis system equipped with an epifluorescence microscope with an automatic excitation filter exchanger, which is more popular in laboratories and easier to handle. We confirmed that the present method worked effectively on the cells in culture. We were able to observe the dramatic rescuing effects of SAE0400 and CsA on cells exposed to ionomycin, which induced a massive increase in cytosolic and mitochondrial Ca2+ concentration and caused acute cell death. The pharmacological profiles of the two drugs seemed to be similar from a Ca2+ dynamics point of view.
Although the dynamics of Ca2+ in cytosol and mitochondria may show different profiles under abnormal conditions in energy production such as ischemia, few studies discussing the difference between the Ca2+ dynamics in the two compartments during ischemia have been published so far. We expected the efficiency of the present method on observing the ischemia-induced Ca2+ abnormality and the effects of drugs on it. We found that the simultaneous observation of Ca2+ dynamics in both cytosol and mitochondria provides important clues in searching for drugs effective for preventing severe brain damage due to Ca2+ overloading. However, we are not satisfied with the present results. The signal observed as cytosolic Ca2+ in the slice preparation was unexpectedly low. We should therefore search for the most relevant experimental conditions suitable for brain-slice preparations in order to increase reliability.
References
Kristian T, Siesjo BK (1998) Calcium in ischemic cell death. Stroke 29(3):705–718
Kurebayashi N, Harkins AB, Baylor SM (1993) Use of FuraRed as an intracellular calcium indicator in frog skeletal muscle fibers. Biophys J 64(6):1934–1960
Matsuda T, Arakawa N, Takuma K, Kishida Y, Kawasaki Y, Sakaue M, Takahashi K, Takahashi T, Suzuki T, Ota T, Hamano-Takahashi A, Onishi M, Tanaka Y, Kameo K, Baba A (2001) SEA0400, a novel and selective inhibitor of the Na+-Ca2+ exchanger, attenuates reperfusion injury in the in vitro and in vivo cerebral ischemic models. J Pharmacol Exp Ther 298:249–256
Minta A, Kao JP, Tsien RY (1989) Fluorescent indicators for cytosolic calcium based on rhodamine and fluorescein chromophores. J Biol Chem 264(14):8171–8178
Uchino H, Elmér E, Uchino K, Lindvall O, Siesjö BK (1995) Cyclosporine A dramatically ameliorates CA1 hippocampal damage following transient forebrain ischemia in the rat. Acta Physiol Scand 155(4):469–471
Voronia S, Sukhomlin T, Johnson PR, Erdemili G, Petersen OH, Tepkin A (2002) Correlation of NADH and Ca2+ signals in mouse pancreatic acinar cells. J Physiol 539:41–52
Conflict of Interest
We declare that we have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer-Verlag Wien
About this paper
Cite this paper
Nishiyama, T. et al. (2013). Simultaneous Measurement of Cytosolic and Mitochondrial Ca2+ During Ischemia in Mice Whole-Brain Slice Preparation and Its Application to Drug Evaluation. In: Katayama, Y., Maeda, T., Kuroiwa, T. (eds) Brain Edema XV. Acta Neurochirurgica Supplement, vol 118. Springer, Vienna. https://doi.org/10.1007/978-3-7091-1434-6_11
Download citation
DOI: https://doi.org/10.1007/978-3-7091-1434-6_11
Published:
Publisher Name: Springer, Vienna
Print ISBN: 978-3-7091-1433-9
Online ISBN: 978-3-7091-1434-6
eBook Packages: MedicineMedicine (R0)