
Abstract
Preterm delivery in humans, defined as birth before 90 % gestation, is a leading cause of infant morbidity and mortality worldwide, occurring in approximately 10 % of all pregnancies (McIntire et al. 1999). Infants born prematurely account for the majority of all neonatal deaths. Not surprisingly, preterm infants show various signs of organ immaturity and this may make preterm neonates more sensitive to serious feeding-induced gastrointestinal complications (Siggers et al. 2011). The immature gastrointestinal tract is less able to deal with microbiology, immunology, and nutrition-related challenges of postnatal life as a result of deficiencies in intestinal structural integrity, digestive capacity, and intestinal immunity (Neu 2007). Such deficiencies are associated with increased enteric disease susceptibility in preterm versus term neonates. Thus, investigating means of improving these deficiencies will aid in improving the maturation of the preterm gastrointestinal tract, in reducing gut inflammation, and in optimizing nutrition and health in this compromised population.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Preterm Delivery
Preterm delivery in humans, defined as birth before 90 % gestation, is a leading cause of infant morbidity and mortality worldwide, occurring in approximately 10 % of all pregnancies (McIntire et al. 1999). Infants born prematurely account for the majority of all neonatal deaths. Not surprisingly, preterm infants show various signs of organ immaturity and this may make preterm neonates more sensitive to serious feeding-induced gastrointestinal complications (Siggers et al. 2011). The immature gastrointestinal tract is less able to deal with microbiology, immunology, and nutrition-related challenges of postnatal life as a result of deficiencies in intestinal structural integrity, digestive capacity, and intestinal immunity (Neu 2007). Such deficiencies are associated with increased enteric disease susceptibility in preterm versus term neonates. Thus, investigating means of improving these deficiencies will aid in improving the maturation of the preterm gastrointestinal tract, in reducing gut inflammation, and in optimizing nutrition and health in this compromised population.
2 Animal Models
However, less detailed information is available from human infants partly due to the difficulties in performing well-controlled studies on this vulnerable population of infants. In addition, it is neither ethical nor practical to conduct these experiments with the human fetus or infant.
Often, the more suitable approach is the use of animal models. While some earlier studies have focused on information derived mainly from rodent models (e.g., rats and mice) (Sodhi et al. 2008), it is difficult to conduct experiments with laboratory rodents because of their small body size and immature organs at birth, and this makes the large farm animals (e.g., pigs, cattle, sheep) more attractive models in this field.
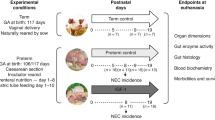
Although no animal model will ever perfectly mimic the human condition, the pig has emerged as a superior non-primate experimental animal model because of similarities in anatomy, development, nutrition, and physiology between the pig and the human (Ball et al. 1996; Clouard et al. 2012). Pigs are also the only widely utilized animal model that is truly omnivorous, and they have strikingly similar nutritional requirements to that of humans (Patterson et al. 2008). The gut in the newborn pig is more mature than in newborn rodents, although less mature than in infants (Sangild 2006). Thus, in pigs, preterm delivery at 90 % gestation is comparable to preterm infants born at approximately 75 % gestation (30 weeks) (Siggers et al. 2011). In contrast to rodent models, the size of the newborn pig easily allows for clinically tissue collections and experimental manipulation of physiologic conditions. Besides, the ontogeny, the physiology of digestion, and associated metabolic processes are very similar between humans and pigs (Patterson et al. 2008; Patrycja and Barbara 2008), which makes the pig an attractive animal model for further studies on intestinal complications related to preterm birth.
3 Determinants in Intestinal Inflammatory Diseases
Preterm infants suffer from numerous devastating intestinal diseases, including necrotizing enterocolitis (NEC). Nutritional, microbial, and immunological dysfunctions may all play a role in disease progression. The lowered digestive and nutrient absorptive function, impaired intestinal epithelial barrier, inappropriate bacterial colonization, and a dysregulated mucosal immune system may add to this increased susceptibility to enteric disease.
3.1 Immature Digestion and Nutrient Absorption in Preterm Neonates
Preterm birth is associated with immature motility, digestive capacity, and nutrient absorption, thus leading to nutrient fermentation, bacterial overgrowth, and mucosal inflammation.
Gastrointestinal motility is limited largely to infants less than 34 weeks gestation (Riezzo et al. 2009). The motility is found considerably less organized in premature infants (Neu 2007), probably, because of the intrinsic immaturity of the enteric neurons. Also in preterm piglets, bowel movements are not well developed during the first days of enteral feeding (Sangild et al. 2002a, b, c). The incomplete innervation and poor motility of the immature gut, thereby, may lead to stasis, nutrient fermentation, inappropriate colonization, and further contribute to the development of enteric disease in preterm infants (Neu 2007; Oste et al. 2005).
Studies in both animals and infants indicate that immature brush-border enzyme activities following preterm delivery may result in maldigestion, excessive nutrient fermentation, intestinal distension, and mucosal ischemia in preterm infants. In preterm infants, the intestine is relatively short (Weaver et al. 1991) and may have a reduced absorptive area, consistent with studies in pigs (Sangild et al. 2000, 2002a, b, c). Besides, preterm-delivered pigs also differ from term neonates in their intestinal cell proliferative and apoptotic responses (Burrin et al. 2000; Bittrich et al. 2004). The impaired ability of the immature intestine to increase cell proliferation, decrease apoptosis, and regulate the mesenteric blood flow (Crissinger et al. 1994; Clark et al. 2005; Dyess et al. 1993) may lead to mucosal atrophy, dysfunction, and necrosis in preterm neonates. In addition to ontogenetic immaturity of enterocyte function, the possible hypoxia, hypothermia, altered endocrine and metabolic status may make preterm neonates more sensitive to serious feeding-induced complications (Sangild 2006).
These deficiencies in intestinal structure and function appear to be one of the most critical problems resulting in feeding intolerance, a commonly encountered problem in neonatal care. Formula-fed preterm neonates are thus at increased risk of developing diseases due to their compromised digestive system. Similar results are observed in preterm newborn pigs. Preterm birth affects the intestinal response to enteral nutrition in newborn piglets. Formula feeding in preterm newborn pigs leads to a diminished intestinal trophic responses relative to colostrum (Bjornvad et al. 2005; Oste et al. 2005; Sangild et al. 2006), marked atrophy of the mucosal surface, and increased permeability (Rouwet et al. 2002). Preterm neonates, thus, may require a period of total parenteral nutrition (TPN) before enteral nutrition is administered (Heird and Gomez 1996; Sangild et al. 2002a, b, c). Nevertheless, parenteral nutrition has significant detrimental side effects, including intestinal atrophy, malfunction, and sepsis leading to increased susceptibility to inflammatory stimuli and the development of intestinal inflammation (Siggers et al. 2011). This is consistent with the observation that TPN increases the risk of NEC in prematurely born piglets.
3.2 Deficient Host-Associated Defense Mechanisms in Preterm Neonates
Relative to term neonates, preterm neonates show immature intestinal barrier that lacks several key protective mechanisms that normally prevent invasion by luminal bacteria. The premature gastrointestinal tract also has increased intestinal permeability (Neu 1996) commonly observed in preterm neonates (Neu 2007), making the immature intestine more permeable to macromolecules, while little is known about the maturation of tight junction proteins such as occludin and claudins, which constitute the major paracellular barrier of the epithelium (Nusrat et al. 2000). The mucus layer forms a physical barrier between the underlying epithelium and the lumen of the gastrointestinal tract (Atumal et al. 2001), protecting the epithelium against noxious agents and pathogenic bacteria. However, the production of mucous was reported to be immature in preterm infants (Claud and Walker 2001; Omari and Davidson 2003; Sangild 2006), leading to a diminished intestinal barrier function, impaired mucosal repair, and lowered degradation of bacterial toxins.
Additionally, impaired functioning of immune defenses in preterms (Sangild et al. 2002a, b, c; Baxter 2010), as well as lower levels of immunoglobulins (e.g., IgA, IgM, IgG) (Lin 2004), gut B and T lymphocytes, makes premature neonates particularly susceptible to enteric inflammation and injury during the early postnatal period (Claud and Walker 2001; Kuitunen and Savilahti 1995). Coupled with the increased intestinal mucosal permeability in preterms (Rouwet et al. 2002; van Elburg et al. 2003), this impairment leads to transmural translocation of microbes or their toxic products into the immature intestinal mucosal barrier in neonates. This may, in turn, further compromise intestinal defense mechanisms and eventually culminate in an inflammatory cascade, leading to NEC (Berman and Moss 2011). Furthermore, the premature gastrointestinal tract also has decreased regenerative capabilities, and the imbalance between epithelial cell injury and repair usually leads to a vicious cycle of maldigestion, impaired mucosal protection, immune activation and results in a greater potential for tissue damage (Siggers et al. 2011).
3.3 Gut Microflora and Inflammatory Responses in Preterm Neonates
Several studies in both human infants and piglets show that assemblages of gut bacteria differ markedly between preterm and term neonates (Schmidt et al. 2008; Cilieborg et al. 2010). While the full-term infants are rapidly colonized with a more diverse microbiota, preterm neonates have a slow bacterial colonization and decreased bacterial diversity present, which may predispose the premature gut to bacterial overgrowth by pathogenic bacteria (Arboleya et al. 2012; Fanaro et al. 2003; Schwiertz et al. 2003), and this difference may be directly related to the degree of intestinal prematurity, the deficient mechanical defense barriers, as well as the environmental factors (Hallstrom et al. 2004). Wang et al (2011) showed that intestinal microbiota had important functions in host energy metabolism, amino acid nutrition, immunity, and health.
Moreover, the nature of the enteral foods may affect initial bacterial colonization patterns in preterm infants (Caicedo et al. 2005; Claud and Walker 2001) and pigs (Shulman 2002; Wang et al. 2011), and the use of parenteral nutrition further delay colonization (Fanaro et al. 2003; Caicedo et al. 2005) in neonates following preterm delivery. Maldigestion and disturbances in intestinal barrier function may lead to bacterial overgrowth and excessive nutrient fermentation, thereby rendering the mucosa more susceptible to bacterial infections and further initiating uncontrolled inflammatory reactions (Siggers et al. 2011).
A disordered enterocyte signaling to bacterial toxins, via the production of various pro-inflammatory cytokines (e.g., IL-1, IL-6, IL-8, TNF-alpha), is thought to be crucial in the development of NEC in the susceptible preterm infants (Hunter et al. 2008). Those pro-inflammatory mediators may further initiate the inflammatory cascade, thus favoring mucosal barrier disruption and adversely affecting mucosal repair. Toll-like receptors are identified among the immunological components of the early mucosal dysfunction. It has been shown that TLRs (2 and 4) are upregulated in intestinal tissue from preterm pigs with NEC (Sangild 2006). In addition, p38 kinase, cyclooxygenase-2, and NF-kB signaling pathways may all be involved in mucosal inflammation (Grishin et al. 2006; Wang et al. 2010). It remains difficult, however, to further understand why the premature newborn is susceptible to NEC as well as other inflammatory bowel diseases, which may need further study in preterm animal models.
4 Nutritional Modulation of Mucosal Defense and Immunology in Preterm Neonates
As previously stated, the immature neonates are prone to bacterial infection and exaggerated immune responses, potentially resulting in irreversible tissue damage. Mother’s milk and colostrum contains numerous bioactive factors, including growth factors, immunoglobulins, anti-inflammatory components, as well as amino acids (Møller et al. 2011; Claud et al. 2003), which help to protect against the development of intestinal diseases. Immunonutrition with specific nutrients may also be effective to modulate the activity of the immune system. Some dietary components, such as amino acids, have already been shown to enhance mucosal barrier function and immunologic responses in animal models and in humans (Stechmiller et al. 2004). Kim et al (2007) reported that provision of amino acids and fatty acids with specific functions may enhance the performance of pregnant and lactating sows by modulating key metabolic pathways by which can enhance conception rates, embryogenesis, blood flow, antioxidant activity, appetite, translation initiation for protein synthesis, immune cell proliferation, and intestinal development. Increasing evidence shows that dietary supplementation of specific amino acids to animals and humans with malnutrition and infectious disease enhances the immune status, thereby reducing morbidity and mortality (Li et al. 2007; Yin et al. 2010).
4.1 Arginine
The arginine, which is nutritionally essential for neonates, is involved in a number of biological and physiological processes. Arginine is crucial for the synthesis of protein and molecules (e.g., nitric oxide (NO), creatine, and polyamines) with enormous physiological importance (Flynn et al. 2002; Rhoads et al. 2004; He et al. 2009; Kim et al. 2007; Li et al. 2007; Wu et al. 2009). Nitric oxide is a vasodilator involved in intestinal permeability, mucosal integrity, and barrier function (Upperman et al. 2005; Wu et al. 2007). Moderate levels of NO are important for regulation of mesenteric blood flow and protect the mucosa from injury. Polyamines are involved in the regulation of gene expression, DNA and protein synthesis, apoptosis, as well as cellular division (Flynn et al. 2002). Moreover, arginine stimulates the secretion of growth hormone and insulin in preterm infants (Vlaardingerbroek et al. 2011), thereby playing an important role in regulating nutrients metabolism (Liu et al. 2008; Yao et al. 2008, 2011; Yin and Tan 2010; Tan et al. 2009, 2011).
Intestinal amino acid metabolism differs between preterm and term birth. A significant nutritional problem in preterm infants is a severe deficiency of arginine (hypoargininemia), which occurs in more than 50 % of the preterm infant population (Wu et al. 2004). Arginine deficiency may result in hyperammonemia as well as intestinal, immunological, and neurological dysfunction (Flynn et al. 2002) and it is often associated with an increased incidence of NEC in preterm infants (Becker et al. 2000). Thus, the knowledge of arginine metabolism and physiological effects is beneficial for optimizing neonatal survival and health in this compromised population.
Wu et al. (1999) reported that the amino acid composition of the fetal pig was similar to that of human fetus. It has been reported that endogenous synthesis of arginine is important for maintaining arginine homeostasis in the neonatal pigs (Flynn and Wu 1996), and the underdevelopment of intestinal arginine synthesis may be primarily responsible for hypoargininemia in preterm neonates (Dekaney et al. 2003). Glutamine and proline are major substrates for intestinal synthesis of citrulline in pigs (Wu et al. 2000a, b, 2009; Wang et al. 2008). However, synthesis of citrulline is low and there is little conversion of citrulline into arginine in enterocytes of preterm neonates owing to the limited expression of the genes for key enzymes (e.g., pyrroline-5-carboxylate synthase, argininosuccinate synthase, and lyase), thereby contributing to hypoargininemia (Wu et al. 2004). Furthermore, the possible increase in whole-body arginine catabolism, as well as absence of perinatal cortisol surge due to premature delivery, may also be responsible for the limited endogenous synthesis of arginine. In preterm piglets, low rates of intestinal arginine synthesis are associated with low plasma arginine concentrations (Urschel et al. 2007). Plasma levels of citrulline, arginine, and glutamine are lower in premature neonates with NEC compared with healthy infants (Wu et al. 2001; Becker et al. 2000); and provision of exogenous arginine prevents hyperammonemia and reduces NEC (Amin et al. 2002). Thus, an enhancement of endogenous arginine synthesis in preterm neonates may be obtained by the promotion of the maturation of intestinal arginine-synthetic enzymes.
Glucocorticoids play a crucial role in advancing the maturation of intestinal arginine synthesis and possibly decrease the incidence of NEC in preterm infants (Bauer et al. 1984). Administration of cortisol is effective to advance the maturation of intestinal arginine synthesis in preterm neonates (Wu et al. 2004). Another promising candidate is glucagon-like peptide-2 (GLP-2), a nutrient-responsive gut hormone, which may exert multiple effects on intestinal mucosa growth in preterm neonates (Estall and Drucker 2005). In neonates, proline is a dietary precursor for arginine and is dependent on intact gut metabolism (Vlaardingerbroek et al. 2011). Preterm infants receiving PN are unable to synthesize sufficient proline de novo (Miller et al. 1995). Future research is needed to define mechanisms for arginine metabolism and develop strategies for arginine deficiency in preterm infants.
Arginine plays an important role in improving intestinal function and regulating nutrient metabolism, but the underlying mechanisms are largely unknown (Liu et al. 2008). He et al. (2009) conducted metabolomic analysis of the response of growing pigs to dietary l-arginine and found that arginine alters the catabolism of fat and amino acids in the whole body, enhances protein synthesis in skeletal muscle, and modulates intestinal microbial metabolism. Tan et al (2011) indicated that Arg differentially regulates expression of fat-metabolic genes and increases mTOR signaling activity in skeletal muscle (Yao et al. 2008) and white adipose tissue, therefore favoring lipogenesis in muscle but lipolysis in adipose tissue.
4.2 Glutamine
Glutamine is the preferred fuel for rapidly proliferating cells including enterocytes (Chauhan et al. 2008). It is essential for many metabolic processes, and supplementation with this amino acid has been demonstrated to improve mucosal integrity and intestinal barrier function in critically ill patients. In vitro, glutamine is required for barrier function (Ewaschuk et al. 2011) and helps recovery from loss of transepithelial resistance and increase of permeability induced by stress in Caco-2 cells (Li et al. 2003). Wang et al (2008) found that early weaning resulted in increased expression of genes related to oxidative stress and immune activation but decreased expression of genes related to macronutrient metabolism and cell proliferation in the gut. Dietary glutamine supplementation increased intestinal expression (120–124 %) of genes that are necessary for cell growth and removal of oxidants, while reducing (34–75 %) expression of genes that promote oxidative stress and immune activation. Functionally, the glutamine treatment enhanced intestinal oxidative-defense capacity, prevented jejunal atrophy, and promoted small intestine growth and body weight gain in weaned piglets. These findings reveal coordinate alterations of gene expression in response to weaning and aid in providing molecular mechanisms for the beneficial effect of dietary glutamine supplementation to improve nutrition status in young mammals.
Glutamine is abundant in mother’s milk but present in much lower levels in formula milk (Agostoni et al. 2000). Thus, neonates, prematurely born infants, would benefit from glutamine addition. However, clinical studies of glutamine supplementation remain inconclusive. Parenteral glutamine appears to be well tolerated and safe in preterm neonates and this amino acid reduces the time to achieve full enteral nutrition (Thompson et al. 2003). Provision of glutamine showed some beneficial effects such as inhibition of whole body protein breakdown (Kadrofske et al. 2006) and activation of immune system in preterm infants (Parimi and Kalhan 2007). However, Tubman and Thompson (2001) reported that no additional benefit of the addition of glutamine to preterm infants was observed in their study. More studies are needed to evaluate the efficacy of this amino acid in neonatal nutrition and to understand the mechanism of glutamine dysfunction-related pathology.
4.3 Methionine and Cysteine
Methionine, an essential amino acid, is also a source through cysteine production for the synthesis of glutathione (GSH). This latter compound plays a crucial role in reducing intestinal oxidative damage and inflammation (Thomas et al. 2008). The rates of transsulfuration of methionine are high in prematurely born low birth weight infants (Maaike et al. 2007a, b). This may reflect high demands for glutathione (GSH) and methionine in parenteral amino acid mixtures for premature babies. It is reported that general cysteine requirement is less than 18 mg/kg per day and that cysteine is probably not a conditionally essential amino acid in the prematurely born infant (Maaike et al. 2007a, b). Regardless of the adequate GSH and protein synthesis, methionine has been implicated in increased homocysteine concentration (Courtney-Martin et al. 2008; Shoveller et al. 2004) in the neonate. Therefore, a balance between methionine and cysteine should be taken in consideration to provide the adequate total sulfur amino acid (SAA) in neonatal nutrition (Courtney-Martin et al. 2010).
4.4 Branched-Chain Amino Acids
The essential branched-chain amino acids (BCAAs), leucine, isoleucine, and valine, are used for incorporation into body protein (Maingay-de Groof et al. 2010; Li et al. 2011), and utilization by intestine is also high. The uptakes of total leucine and valine carbon are relatively large and the oxidation rates of these essential branched-chain amino acids are high in fetuses and neonates (van den Akker et al. 2011). Among the BCAAs, leucine can act as a nutrient signal and stimulates protein synthesis via the activation of translation initiation factors (Vlaardingerbroek et al. 2011). Recent work with young pigs shows that reducing dietary protein intake can improve gut function after weaning but result in inadequate provision of essential amino acids for muscle growth. Yin et al. (2010) reported that supplementing l-leucine to a low-protein diet may maintain the activation of translation initiation factors and adequate protein synthesis in multiple organs of post-weaning pigs. This novel finding provides a molecular basis for designing effective nutritional means to increase the efficiency of nutrient utilization for protein accretion in neonates. Leucine is not only a substrate for protein synthesis of skeletal muscle but also plays as signaling molecules to affect feeding behavior, energy balance, and fuel efficiency (Li et al. 2011). Leucine activates signaling factor of mammalian target of rapamycin (mTOR) to promote protein synthesis in skeletal muscle and in adipose tissue. It is also a major regulator of the mTOR sensitive response of food intake to high protein diet. Meanwhile, leucine regulates blood glucose level by promoting gluconeogenesis and aids in the retention of lean mass in a hypocaloric state. It is beneficial to animal nutrition and clinical application and extrapolation to humans (Li et al. 2011). Studies in newborn pigs suggest that enteral leucine supplementation may have a beneficial effect on neonatal growth as it may enhance protein synthesis in an mTORC1-dependent pathway (Suryawan et al. 2012; Li et al. 2011). The gut has a high demand for leucine and protein synthesis has been found to be limited by deficient leucine intake (Elango et al. 2002). BCAA-enriched parenteral nutrition in preterm neonates might influence functional outcome in the direct postnatal phase. The requirements of the individual BCAAs are almost twice the current recommendations (Maingay-de Groof et al. 2010). To optimize current parenteral and enteral feeding, the optimal BCAA ratio should be determined for both. Further studies are required to better understand the role of BCAAs in the regulation of neonatal growth.
4.5 Threonine
Threonine is an indispensable amino acid that must come from dietary sources. It is critical in the production of mucins in the gut (Schaart et al. 2009). Therefore, this amino acid is of presumably vital nutritional importance to maintain the protective mucus layer and thus the intestinal barrier function. Lack of threonine can result in diarrhea and reduced mucin production, indicating the important role of threonine in the structure and function of the gut (Vlaardingerbroek et al. 2011). However, prolonged dietary excess of threonine fed to neonates, may have negative behavioral consequences and may induce serious metabolic disturbances (Chapman et al. 2009). Dietary threonine imbalance is known to reduce the growth of the small intestine, liver, and skeletal muscle in young animals. Using the pig model, Wang et al. (2007) found that either a deficiency or an excess of dietary threonine impairs protein synthesis in these tissues. This finding provides a mechanism for the low growth performance of animals fed a threonine-imbalanced diet. Currently, neonatal amino acid solutions provide intakes of threonine (111–165 mg/kg per day) are greater than an infant’s enteral intake from breast milk (76 mg/kg per day) (WHO 2007). Chapman et al. (2009) concluded that current parenteral solutions should be revised to incorporate the population-safe requirements of threonine to promote optimum metabolic and neurologic growth in neonates.
4.6 Tryptophan
Tryptophan is an essential amino acid while the concentration is low in plasma and content low in proteins compared with the other essential amino acids (Vlaardingerbroek et al. 2011). Intestinal inflammation, malnutrition, and pro-inflammatory situation may result in tryptophan depletion (Christmas et al. 2011), thus affecting the weight gain and nitrogen balance in neonates. In young pigs, experimental inflammation was associated to a decrease in plasma tryptophan concentrations compared with healthy piglets (Le Floc’h et al. 2008). Furthermore, studies showed that in neonatal piglets, no difference is found in tryptophan requirements when enteral and parenteral feeding are compared (Alegria et al. 1999). Therefore, most parenteral neonatal amino acid solutions contain similar concentrations of tryptophan compared with human breast milk (Cvitkovic et al. 2004). Lastly, the commercially available preterm formula content ranges from 18 to 36 mg/kg per day when infants receive 160 ml/kg per day of milk (Vlaardingerbroek et al. 2011).
4.7 Other Amino Acids
Taurine is considered conditionally essential because needs are not met when intake is low (Verner et al. 2007). It has important roles in intestinal absorption, membrane stability, and visual development in preterm infants. Currently, taurine concentrations in modern parenteral amino acid solutions are more than sufficient to meet recommendations (Verner et al. 2007). Tyrosine is also a conditionally essential amino acid resulting from the insufficient enzymatic activity in preterm infants. Due to poor tyrosine solubility in parenteral nutrition (Roberts et al. 2001), hydroxylation of phenylalanine to tyrosine may be a good way to provide tyrosine when the diet is tyrosine-deficient.
5 Conclusion
Deficiencies in intestinal integrity, barrier function, digestive capacities, and intestinal immunity, make preterm neonates more susceptible to inflammatory diseases. Using preterm pigs, recent evidence suggests that nutritional modulation has great potential to improve neonatal intestinal development, manipulate the gut microbiota, in parallel with direct effects on the mucosal immune system, to prevent the onset of NEC. Some amino acids have been shown to enhance mucosal barrier function, immunologic responses, and NEC resistance in piglets and in neonates. However, it always remains a challenge to translate data generated from animal models to corresponding conditions in humans. Understanding the unique function of amino acids could eventually play a pivotal role in improving future nutritional strategies for premature infants. Clearly, further studies involving amino acids as compounds for prevention and clinical therapies against devastating intestinal diseases are needed, both in parenterally and enterally fed infants. Such work will greatly advance our knowledge with regard to the “optimal” amino acid pattern and it will also be beneficial for designing the next generation of amino acid supplemental solutions to optimize survival and health in preterm neonates.
References
Agostoni C, Carratù B, Boniglia C, Riva E, Sanzini E (2000) Free amino acid content in standard infant formulas: comparison with human milk. J Am Coll Nutr 19:434–438
Alegria A, Barbera R, Farre R, Lagarda MJ, Lopez JC (1999) Amino acid contents of infant formulas. J Food Compos Anal 12:137–146
Amin HJ, Zamora SA, McMillan DD, Fick GH, Butzner JD, Parsons HG et al (2002) Arginine supplementation prevents necrotizing enterocolitis in the premature infant. J Pediatr 140:425–431
Arboleya S, Binetti A, Salazar N, Fernández N, Solís G, Hernández-Barranco A, Margolles A, de Los Reyes-Gavilán CG, Gueimonde M (2012) Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol Ecol 79:763–772
Atumal C, Strugala V, Allen A, Holm L (2001) The adherent gastrointestinal mucus gel layer: thickness and physical state in vivo. Am J Physiol Gastrointest Liver Physiol 280:G922–G929
Ball RO, House JD, Wykes LL, Pencharz PB (1996) A piglet model for neonatal amino acid metabolism during total parenteral nutrition. In: Tumbleson ME, Schook LB (eds) Advances in swine in biomedical research. Plenum Press, New York, pp 713–731
Bauer CR, Morrison JC, Poole WK, Korones SB, Boehm JJ, Rigatto H, Zachman RD (1984) A decreased incidence of necrotizing enterocolitis after prenatal glucocorticoid therapy. Pediatrics 73:682–688
Baxter D (2010) Impaired functioning of immune defenses to infection in premature and term infants and their implications for vaccination. Hum Vaccin 6:494–505
Becker RM, Wu G, Galanko JA, Chen W, Maynor AR, Bose CL, Rhoads JM (2000) Reduced serum amino acid concentrations in infants with necrotizing enterocolitis. J Pediatr 137:785–793
Berman L, Moss RL (2011) Necrotizing enterocolitis: an update. Semin Fetal Neonatal Med 16:145–150
Bittrich S, Philipona C, Hammon HM, Rome V, Guilloteau P, Blum JW (2004) Preterm as compared with full-term neonatal calves are characterized by morphological and functional immaturity of the small intestine. J Dairy Sci 87:1786–1795
Bjornvad CR, Schmidt M, Petersen YM, Jensen SK, Offenberg H, Elnif J, Sangild PT (2005) Preterm birth makes the immature intestine sensitive to feeding-induced intestinal atrophy. Am J Physiol Regul Integr Comp Physiol 289:R1212–R1222
Burrin DG, Stoll B, Jiang R, Petersen Y, Elnif J, Buddington RK, Schmidt M, Holst JJ, Hartmann B, Sangild PT (2000) GLP-2 stimulates intestinal growth in premature TPN-fed pigs by suppressing proteolysis and apoptosis. Am J Physiol Gastrointest Liver Physiol 279:G1249–G1256
Caicedo RA, Schanler RJ, Li N, Neu J (2005) The developing intestinal ecosystem: implications for the neonate. Pediatr Res 58:625–628
Chapman KP, Courtney-Martin G, Moore AM, Ball RO, Pencharz PB (2009) Threonine requirement of parenterally fed postsurgical human neonates. Am J Clin Nutr 89:134–141
Chauhan M, Henderson G, McGuire W (2008) Enteral feeding for very low birth weight infants: reducing the risk of necrotising enterocolitis. Arch Dis Child Fetal Neonatal Ed 93:F162–166
Christmas DM, Potokar J, Davies SJ (2011) A biological pathway linking inflammation and depression: activation of indoleamine 2,3-dioxygenase. Neuropsychiatr Dis Treat 7:431–439
Cilieborg M, Boye M, Thymann T, Mølbak L, Sangild PT (2010) Preterm birth and necrotizing enterocolitis alter gut colonization in pigs. Pediatr Res 69:10–16
Clark JA, Lane RH, Maclennan NK, Holubec H, Dvorakova K, Halpern MD, Williams CS, Payne CM, Dvorak B (2005) Epidermal growth factor reduces intestinal apoptosis in an experimental model of necrotizing enterocolitis. Am J Physiol Gastrointest Liver Physiol 288:G755–G762
Claud EC, Walker WA (2001) Hypothesis: inappropriate colonization of the premature intestine can cause neonatal necrotizing enterocolitis. FASEB J 15:1398–1403
Claud EC, Savidge T, Walker WA (2003) Modulation of human intestinal epithelial cell IL-8 secretion by human milk factors. Pediatr Res 53:419–425
Clouard C, Meunier-Salaün MC, Val-Laillet D (2012) Food preferences and aversions in human health and nutrition: how can pigs help the biomedical research? Animal 6:118–36
Courtney-Martin G, Chapman KP, Moore AM, Kim JH, Ball RO, Pencharz PB (2008) Total sulfur amino acid requirement and metabolism in parenterally fed postsurgical human neonates. Am J Clin Nutr 88:115–124
Courtney-Martin G, Moore AM, Ball RO, Penchar PB (2010) The addition of cysteine to the total sulphur amino acid requirement as methionine does not increase erythrocytes glutathione synthesis in the parenterally fed human neonate. Pediatr Res 67:320–324
Crissinger KD, Burney DL, Velasquez OR, Gonzalez E (1994) An animal model of necrotizing enterocolitis induced by infant formula and ischemia in developing piglets. Gastroenterology 106:1215–1222
Cvitkovic S, Bertolo RF, Brunton JA, Pencharz PB, Ball RO (2004) Enteral tryptophan requirement determined by oxidation of gastrically or intravenously infused phenylalanine is not different from the parenteral requirement in neonatal piglets. Pediatr Res 55:630–636
Dekaney CM, Wu G, Jaeger LA (2003) Gene expression and activity of enzymes in the arginine biosynthetic pathway in porcine fetal small intestine. Pediatr Res 53:274–80
Dyess DL, Peeples GL, Ardell JL, Tacchi EJ, Roberts WS, Ferrara JJ, Powell RW (1993) Indomethacin-induced blood flow distribution in premature and full-term piglets. J Pediatr Surg 28:1396–1400
Elango R, Pencharz PB, Ball RO (2002) The branched-chain amino acid requirement of parenterally fed neonatal piglets is less than the enteral requirement. J Nutr 132:3123–3129
Estall JL, Drucker DJ (2005) Tales beyond the crypt: glucagon-like peptide-2 and cytoprotection in the intestinal mucosa. Endocrinology 146:19–21
Ewaschuk JB, Murdoch GK, Johnson IR, Madsen KL, Field CJ (2011) Glutamine supplementation improves intestinal barrier function in a weaned piglet model of Escherichia coli infection. Br J Nutr 106:870–877
Fanaro S, Chierici R, Guerrini P, Vigi V (2003) Intestinal microflora in early infancy: composition and development. Acta Paediatr Suppl 91:48–55
Flynn NE, Wu G (1996) An important role for endogenous synthesis of arginine in maintaining arginine homeostasis in neonatal pigs. Am J Physiol Regul Integr Comp Physiol 271:R1149–R1155
Flynn NE, Meininger CJ, Haynes TE, Wu G (2002) The metabolic basis of arginine nutrition and pharmacotherapy. Biomed Pharmacother 56:427–438
Grishin AV, Wang J, Potoka DA, Hackam DJ, Upperman JS, Boyle P, Zamora R, Ford HR (2006) Lipopolysaccharide induces cyclooxygenase-2 in intestinal epithelium via a noncanonical p38 MAPK pathway. J Immunol 176:580–588
Hallstrom M, Eerola E, Vuento R, Janas M, Tammela O (2004) Effects of mode of delivery and necrotising enterocolitis on the intestinal microflora in preterm infants. Eur J Clin Microbiol Infect Dis 23:463–470
He QH, Kong XF, Wu GY, Ren PP, Tang HR, Hao FH, Huang RL, Li TJ, Tan BE, Li P, Tang ZR, Yin YL, Wu YN (2009) Metabolomic analysis of the response of growing pigs to dietary L-arginine supplementation. Amino Acids 37:199–208
Heird WC, Gomez MR (1996) Parenteral nutrition in low-birth-weight infants. Annu Rev Nutr 16:471–499
Hunter CJ, Upperman JS, Ford HR, Camerini V (2008) Understanding the susceptibility of the premature infant to necrotizing enterocolitis (NEC). Pediatr Res 63:117–123
Kadrofske MM, Parimi PS, Gruca LL, Kalhan SC (2006) Effect of intravenous amino acids on glutamine and protein kinetics in low-birth-weight preterm infants during the immediate neonatal period. Am J Physiol Endocrinol Metab 290:E622–E630
Kim SW, Mateo RD, Yin YL, Wu GY (2007) Functional amino acids and fatty acids for enhancing production performance of sows and piglets. Asian Australas J Anim Sci 20:295–306
Kuitunen M, Savilahti E (1995) Mucosal IgA, mucosal cow’s milk antibodies, serum cow’s milk antibodies and gastrointestinal permeability in infants. Pediatr Allergy Immunol 6:30–35
Le Floc’h N, Melchior D, Sève B (2008) Dietary tryptophan helps to preserve tryptophan homeostasis in pigs suffering from lung inflammation. J Anim Sci 86:3473–3479
Li N, DeMarco VG, West CM, Neu J (2003) Glutamine supports recovery from loss of transepithelial resistance and increase of permeability induced by media change in Caco-2 cells. J Nutr Biochem 14:401–408
Li P, Yin YL, Li DF, Kim SW, Wu GY (2007) Amino acids and immune function. Br J Nutr 98:237–252
Li FN, Yin YL, Tan B, Kong XF, Wu GY (2011) Leucine nutrition in animals and humans: mTOR signaling 3 and beyond. Amino Acids 41:1185–1193
Lin J (2004) Too much short chain fatty acids cause neonatal necrotizing enterocolitis. Med Hypotheses 62:291–293
Liu YL, Huang JJ, Hou YQ, Zhu HL, Zhao SJ, Ding BY, Yin YL, Yi GF, Shi JX, Fan W (2008) Dietary arginine supplementation alleviates intestinal mucosal disruption induced by Escherichia coli lipopolysaccharide in weaned pigs. Br J Nutr 100:552–560
Maaike AR, van Beek RH, Gardi V, de Bie MH, Dassel AC, van Goudoever JB (2007a) Cysteine: a conditionally essential amino acid in low-birth-weight preterm infants? Am J Clin Nutr 86:1120–1125
Maaike AR, Barbara S, Shaji C, Henk S, Agneta LS, Johannes BG, Douglas GB (2007b) Methionine transmethylation and transsulfuration in the piglet gastrointestinal tract. Proc Natl Acad Sci U S A 104:3408–3413
Maingay-de Groof F, Huang L, Voortman GJ, Chao C, Huang Y, Van Goudoeve JB (2010) 148 branched chain amino acid requirements for term neonates. Pediatr Res 68:78
McIntire DD, Bloom SL, Casey BM, Leveno KJ (1999) Birth weight in relation to morbidity and mortality among newborn infants. N Engl J Med 340:1234–1238
Miller RG, Jahoor F, Jaksic T (1995) Decreased cysteine and praline synthesis in parenterally fed, premature infants. J Pediatr Surg 30:953–958
Møller HK, Thymann T, Fink LN, Frokiaer H, Kvistgaard AS, Sangild PT (2011) Bovine colostrum is superior to enriched formulas in stimulating intestinal function and necrotising enterocolitis resistance in preterm pigs. Br J Nutr 105:44–53
Neu J (1996) Necrotizing enterocolitis: the search for a unifying pathogenic theory leading to prevention. Pediatr Clin North Am 43:409–32
Neu J (2007) Gastrointestinal development and meeting the nutritional needs of premature infants. Am J Clin Nutr 85:629S–634S
Nusrat A, Turner JR, Madara JL (2000) Molecular physiology and pathophysiology of tight junctions. IV. Regulation of tight junctions by extracellular stimuli: nutrients, cytokines, and immune cells. Am J Physiol Gastrointest Liver Physiol 279:G851–G857
Omari TI, Davidson GP (2003) Multipoint measurement of intragastric pH in healthy preterm infants. Arch Dis Child Fetal Neonatal Ed 88:F517–F520
Oste M, Van Ginneken C, Van Haver E, Bjornvad CR, Thymann T, Sangild PT (2005) The intestinal trophic response to enteral food is reduced in parenterally fed preterm pigs and is associated with more nitrergic neurons. J Nutr 135:2657–2663
Parimi PS, Kalhan SC (2007) Glutamine supplementation in the newborn infant. Semin Fetal Neonatal Med 12:19–25
Patrycja P, Barbara S (2008) Animal models to study neonatal nutrition in humans. Curr Opin Clin Nutr Metab Care 11:601–606
Patterson JK, Lei XG, Miller DD (2008) The pig as an experimental model for elucidating the mechanisms governing dietary influence on mineral absorption. Exp Biol Med (Maywood) 233:651–64
Rhoads JM, Chen W, Gookin J, Wu GY, Fu Q, Blikslager AT, Rippe RA, Argenzio RA, Cance WG, Weaver EM, Romer LH (2004) Arginine stimulates intestinal cell migration through a focal adhesion kinase dependent mechanism. Gut 53:514–522
Riezzo G, Indrio F, Raimondi F, Montagna O, Salvia G, Massimo B, Polimeno L, Cavallo L, Francavilla R (2009) Maturation of gastric electrical activity, gastric emptying and intestinal permeability in preterm newborns during the first month of life. Ital J Pediatr 35:6
Roberts SA, Ball RO, Moore AM, Filler RM, Pencharz PB (2001) The effect of graded intake of glycyl-L-tyrosine on phenylalanine and tyrosine metabolism in parenterally fed neonates with an estimation of tyrosine requirement. Pediatr Res 49:111–119
Rouwet EV, Heineman E, Buurman WA, ter Riet G, Ramsay G, Blanco CE (2002) Intestinal permeability and carrier-mediated monosaccharide absorption in preterm neonates during the early postnatal period. Pediatr Res 51:64–70
Sangild PT (2006) Gut responses to enteral nutrition in preterm infants and animals. Exp Biol Med 231:1695–1711
Sangild PT, Schmidt M, Jacobsen H, Fowden AL, Forhead A, Avery B, Greve T (2000) Blood chemistry, nutrient uptake and organ weights of fetal and newborn calves derived from in vitro produced embryos. Biol Reprod 62:1495–1504
Sangild PT, Petersen YM, Schmidt M, Elnif J, Petersen TK, Buddington RK, Greisen G, Michaelsen KF, Burrin DG (2002a) Preterm birth affects the intestinal response to parenteral and enteral nutrition in newborn pigs. J Nutr 132:3786–3794
Sangild PT, Petersen YM, Schmidt M, Elnif J, Petersen TK, Buddington RK, Michaelsen KF, Greisen G, Burrin DG (2002b) Preterm birth affects the gastrointestinal responses to parenteral and enteral nutrition in the newborn pig. J Nutr 132:2673–2681
Sangild PT, Schmidt M, Elnif J, Bjornvad CR, Buddington RK (2002c) Prenatal development of the gastrointestinal tract in pigs and the effect of fetal gut obstruction. Pediatr Res 52:416–424
Sangild PT, Siggers RH, Schmidt M, Elnif J, Bjornvad CR, Thymann T, Grondahl ML, Hansen AK, Jensen SK, Boye M, Moelbak L, Buddington RK, Westrom BR, Holst JJ, Burrin DG (2006) Diet- and colonization-dependent intestinal dysfunction predisposes to necrotizing enterocolitis in preterm pigs. Gastroenterology 130:1776–1792
Schaart MW, de Bruijn AC, Schierbeek H, Tibboel D, Renes IB, van Goudoever JB (2009) Small intestinal MUC2 synthesis in human preterm infants. Am J Physiol Gastrointest Liver Physiol 296:G1085–G1090
Schmidt M, Buddington RK, Sangild PT, Siggers RH, Thymann T, Jensen BB, Mølbak L, Heegaard PM (2008) Elective cesarean delivery affects gut maturation and delays microbial colonization but does not increase necrotizing enterocolitis in preterm pigs. Am J Physiol Regul Integr Comp Physiol 294:R929–R938
Schwiertz A, Gruhl B, Löbnitz M, Michel P, Radke M, Blaut M (2003) Development of the intestinal bacterial composition in hospitalized preterm infants in comparison with breast-fed, full-term infants. Pediatr Res 54:393–399
Shoveller AK, House JD, Brunton JA, Pencharz PB, Ball RO (2004) The balance of dietary sulfur amino acids and the route of feeding affect plasma homocysteine concentrations in neonatal piglets. J Nutr 134:609–612
Shulman RJ (2002) Effect of enteral administration of insulin on intestinal development and feeding tolerance in preterm infants: a pilot study. Arch Dis Child Fetal Neonatal 86:F131–F133
Siggers RH, Siggers J, Thymann T, Boye M, Sangild PT (2011) Nutritional modulation of the gut microbiota and immune system in preterm neonates susceptible to necrotizing enterocolitis. J Nutr Biochem 22:511–521
Sodhi C, Richardson W, Gribar S, Hackam DJ (2008) The development of animal models for the study of necrotizing enterocolitis. Dis Model Mech 1:94–98
Stechmiller JK, Childress B, Porter T (2004) Arginine immunonutrition in critically ill patients: a clinical dilemma. Am J Crit Care 13:17–23
Suryawan A, Nguyen HV, Almonaci RD, Davis TA (2012) Differential regulation of protein synthesis in skeletal muscle and liver of neonatal pigs by leucine through an mTORC1-dependent pathway. J Anim Sci Biotechnol 3:3
Tan B, Yin YL, Liu ZQ, Li XG, Xu HJ, Kong XF, Huang RL, Tang WJ, Shinzato I, Smith SB, Wu GY (2009) Dietary L-arginine supplementation increases muscle gain and reduces body fat mass in growing-finishing pigs. Amino Acids 37:169–175
Tan BE, Yin YL, Liu ZQ, Tang WJ, Xu HJ, Konga XF, Li XG, Yao K, Gu WT, Smith SB, Wu GY (2011) Dietary L-arginine supplementation differentially regulates expression of fat-metabolic genes in porcine adipose tissue and skeletal muscle. J Nutr Biochem 22:441–445
Thomas B, Gruca LL, Bennett C, Parimi PS, Hanson RW, Kalhan SC (2008) Metabolism of methionine in the newborn infant: response to the parenteral and enteral administration of nutrients. Pediatr Res 64:381–386
Thompson SW, McClure BG, Tubman TRJ (2003) A randomized, controlled trial of parenteral glutamine in ill, very low birth-weight neonates. J Pediatr Gastroenterol Nutr 37:550–553
Tubman TR, Thompson SW (2001) Glutamine supplementation for prevention of morbidity in preterm infants. Cochrane Database Syst Rev 4:CD001457
Upperman JS, Potoka D, Grishin A, Hackam D, Zamora R, Ford HR (2005) Mechanisms of nitric oxide-mediated intestinal barrier failure in necrotizing enterocolitis. Semin Pediatr Surg 14:159–66
Urschel KL, Evans AR, Wilkinson CW, Pencharz PB, Ball RO (2007) Parenterally fed neonatal piglets have a low rate of endogenous arginine synthesis from circulating proline. J Nutr 137:601–606
van den Akker CH, Schierbeek H, Minderman G, Vermes A, Schoonderwaldt EM, Duvekot JJ, Steegers EA, van Goudoever JB (2011) Amino acid metabolism in the human fetus at term: leucine, valine, and methionine kinetics. Pediatr Res 70:566–71
van Elburg RM, Fetter WP, Bunkers CM, Heymans HS (2003) Intestinal permeability in relation to birth weight and gestational and postnatal age. Arch Dis Child Fetal Neonatal Ed 88:F52–F55
Verner A, Craig S, McGuire W (2007) Effect of taurine supplementation on growth and development in preterm or low birth weight infants. Cochrane Database Syst Rev 17:CD006072
Vlaardingerbroek H, van den Akker CHP, de Groof F, Hogewind-Schoonenboom JE, Huang L, Riedijk MA, van der Schoor SRD, Huang Y, van Goudoever JB (2011) Amino acids for the neonate: search for the ideal dietary composition. NeoRev 12:e506–e516
Wang X, Qiao SY, Yin YL, Yu LG, Wang ZY, Wu GY (2007) A deficiency or excess of dietary threonine reduces protein synthesis in jejunum and skeletal muscle of young pigs. J Nutr 137:1442–1446
Wang JJ, Chen LX, Li P, Li XL, Zhou HJ, Wang FL, Li DF, Yin YL, Wu GY (2008) Gene expression is altered in piglet small intestine by weaning and dietary glutamine supplementation. J Nutr 138:1025–1032
Wang J, Ford HR, Grishin AV (2010) NF-κB-mediated expression of MAPK phosphatase-1 is an early step in desensitization to TLR ligands in enterocytes. Mucosal Immunol 3:523–534
Wang SP, Blachier F, Zhao F, Yin YL (2011) Intestinal microbiota: development, metabolism and functions. J Food Agric Environ 9:121–129
Weaver LT, Austin S, Cole TJ (1991) Small intestinal length: a factor essential for gut adaptation. Gut 32:1321–1323
WHO (2007) Protein and amino acid requirements in human nutrition. WHO Tech Rep Ser 935:1–265
Wu G, Ott TL, Knabe DA, Bazer FW (1999) Amino acid composition of the fetal pig. J Nutr 129:1031–8
Wu G, Meininger CJ, Kelly K, Watford M, Morris SM Jr (2000a) A cortisol surge mediates the enhanced expression of pig intestinal pyrroline-5-carboxylate synthase during weaning. J Nutr 130:1914–1919
Wu G, Meininger CJ, Knabe DA, Bazer FW, Rhoads JM (2000b) Arginine nutrition in development, health and disease. Curr Opin Clin Nutr Metab Care 3:59–66
Wu G, Becker RM, Bose CL, Rhoads JM (2001) Serum amino acid concentrations in preterm infants. J Pediatr 139:334–337
Wu G, Jaeger LA, Bazer FW, Rhoads JM (2004) Arginine deficiency in preterm infants: biochemical mechanisms and nutritional implications. J Nutr Biochem 15:442–451
Wu GY, Bazer FW, Davis TA, Johnson GA, Kim SW, Knabe DA, Spencer TE, Yin YL (2007) Important roles for arginine-family amino acids in swine nutrition and production. Livest Sci 122:8–22
Wu GY, Bazer FW, Davis TA, Kim SW, Li P, Rhoads JM, Satterfield MC, Smith SB, Spencer TE, Yin YL (2009) Arginine metabolism and nutrition in growth, health and disease. Amino Acids 37:169–175
Yao K, Yin YL, Chu WY, Liu ZQ, Dun D, Li TJ, Huang RL, Zhang JS, Bie T, Wang WC, Wu GY (2008) Dietary arginine supplementation increases mTOR signaling activity in skeletal muscle of neonatal pigs. J Nutr 138:867–872
Yao K, Guan S, Li TJ, Huang RL, Wu GY, Ruan Z, Yin YL (2011) Dietary L-arginine supplementation enhances intestinal development and expression of vascular endothelial growth factor in weanling piglets. Br J Nutr 105:703–709
Yin YL, Tan BE (2010) Manipulation of dietary nitrogen, amino acids and phosphorus to reduce environmental impact of swine production and enhance animal health. J Food Agric Environ 8:447–462
Yin YL, Yao K, Liu ZJ, Gong M, Ruan Z, Deng D, Tan BE, Liu ZQ, Wu GY (2010) Supplementing L-leucine to a low-protein diet increases tissue protein synthesis in weanling pigs. Amino Acids 39:1477–1486
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer-Verlag Wien
About this chapter
Cite this chapter
Hu, C., Yin, Y., Shan, A., He, X., Li, W., Fang, J. (2013). The Pig Model for Studying Amino Acid-Related Human Diseases: Amino Acids and Intestinal Diseases in Preterm Infants. In: Blachier, F., Wu, G., Yin, Y. (eds) Nutritional and Physiological Functions of Amino Acids in Pigs. Springer, Vienna. https://doi.org/10.1007/978-3-7091-1328-8_13
Download citation
DOI: https://doi.org/10.1007/978-3-7091-1328-8_13
Published:
Publisher Name: Springer, Vienna
Print ISBN: 978-3-7091-1327-1
Online ISBN: 978-3-7091-1328-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)