
Abstract
In nature, microorganisms prefer to live in structured microbial communities rather than as free-floating planktonic cells. These dynamic microbial communities are termed biofilms, in which transitions between planktonic and sessile modes of growth occur interchangeably in response to different environmental cues. Such phenomenas are advantageous for microbial pathogens but disadvantageous for human health. Due to the increased resistance/tolerance of biofilm cells to antimicrobial treatment, it becomes difficult to eradicate pathogens, which results in relapses of infections even after appropriate therapy. In clinically relevant biofilms, Pseudomonas spp., Staphylococcus spp., and Candida spp. are the most frequently isolated microorganisms. These microorganisms are able to adhere to and colonize surfaces of medical devices such as central venous catheters, intrauterine devices, voice prostheses, and prosthetic joints, resulting in the development of a biofilm. Many antimicrobial agents are now being used against microbial biofilms. However, inappropriate use of conventional antibiotic therapy may also contribute to inefficient biofilm control and to the dissemination of resistance. Consequently, new control strategies are constantly emerging to control biofilm-associated infections, such as the antifungal lock therapy, improved drug delivery, penetration of matrix-attacking extracellular polymetric substances, and regulation of biofilm inhibition/disruption by manipulating small molecules. The present chapter is focused on describing the clinical aspects of biofilm formation and deleterious effects associated with their presence. This chapter will highlight current and emergent control strategies for biofilms.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
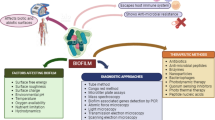
Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
While microbes are often thought to be multiplying and growing as free floating cells, most microbes live in aggregations and form complex structures termed biofilms. These organized structures are communities of microorganisms that form on solid or liquid interfaces and provide protection to individual cells by producing extracellular polymeric substances (EPS). The cells in the biofilms exhibit an altered phenotype compared with corresponding planktonic cells, especially in regard to gene transcription, and in interacting with each other (Donlan 2002; Hall-Stoodley et al. 2004). Biofilms result from a natural tendency of microbes to attach to biotic or abiotic surfaces. The formation of biofilms starts by irreversible attachment of microorganisms to a surface, which can vary from mineral surfaces and mammalian tissues to synthetic polymers and indwelling medical devices, followed by the production of extracellular substances by one or more of the attached microorganisms (Nikolaev and Plankunov 2007; Dongari-Bagtzoglou 2008).
Typically, most of the research on infectious microorganisms is conducted on single-celled (planktonic forms) of bacteria and fungi because of ease of study and manipulation. Consequently, most of the drugs developed have efficacy against planktonic forms of microbes, and unfortunately these drugs do not work or work poorly against the same organisms in their biofilm form. Moreover, the failure of antibacterial and antifungal drugs to combat such infections is due to the increased resistance and/or tolerance of the organisms in their biofilm state. The National Institute of Health estimates that biofilms cause more than 80 % of infections, which have imposed an enormous cost on human health (Sachachter 2003). Most infections on biomedical devices and mucosal surfaces, including oral and uro-genital tracts, are reportedly caused by the biofilm growth of Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus, Streptococcus pyrogens, and Candida albicans (Donlan 2001; Wilson 2001; Douglas 2002).
Development of effective strategies to control or prevent biofilm-associated infections requires a thorough understanding of the biofilm development process (Jain et al. 2007). The adhesion of bacteria to a surface depends on a number of microbiological, physical, chemical, and material-related parameters. Biofilms may consist of mono or mixed species, are highly interactive, and employ a range of cell-to-cell communication or “quorum sensing” (QS) systems (Hogan 2006; Jayaraman and Wood 2008). This phenomenon for promoting collective behavior within a population is important for ensuring survival and propagation by enhancing access to nutrients and niches, as well as for providing protection (Nikolaev and Plankunov 2007). The dense population structure in biofilms also increases the opportunity of gene transfer between the species which can convert a previously avirulent commensal organism into a highly virulent pathogen (Molin and Tolker-Nielson 2003). The enhanced efficiency of gene transfer in biofilms induces enhanced stabilization of the biofilm structure but, more importantly, also facilitates the spread of antibiotic resistance (Molin and Tolker-Nielson 2003; Wuertz and Hausner 2004). The increasing emergence of drug resistance to commonly used antibiotics and antifungals has increased the need for the identification of novel therapeutics and approaches. Therefore, understanding how antibiotic resistance develops is a prerequisite to the design of intervention strategies intended to minimize the threat of biofilm-associated infections. This chapter outlines our understanding and current state of knowledge of the nature of microbial biofilms in clinical context with emphasis on novel prophylactic and therapeutic strategies targeting prevention and management of biofilms.
2 Clinical Significance of Biofilms
It is estimated that the majority of clinical infections exist as biofilms rather than as planktonic cells. In medical settings, biofilms can occur in several places, such as the intestinal brush border (e.g., Vibrio cholerae), urethral lining (e.g., Neisseria gonorrhoeae), lymphoid patches in the intestine (e.g., Salmonella typhimurium) (Costerton et al. 1999), antibiotic-recalcitrant acne (Coates et al. 2003), chronically infected tonsils (Chole and Faddis 2003), cystic fibrosis (lungs) (Prince 2002), urinary and central venous catheters, and mechanical heart valves (Donlan 2002). A list of microorganisms causing infection due to biofilm growth on tissues or medical devices is given in Table 1. Intravascular administration of antibiotics is used to prevent surgical site and other infections, but the formation of a biofilm makes antibiotic therapy ineffective at eradicating the bacteria or fungi. The formation of biofilms with low sensitivity to antibiotics in the course of chronic infections, such as cystic fibrosis, is a matter of great concern (Il’ina et al. 2004).
A range of mucosal to systemic fungal infections have been reported to be caused by opportunistic pathogen Candida spp. such as oral candidiasis, vaginitis, and candidemia. Vulvovaginal infections are among the most common infections caused by C. albicans. Most women experience a vaginal Candida infection at some point in their lifetimes (Mardh et al. 2002). Oropharangeal candidiasis occurs most commonly in immunocompromised individuals, especially people infected with HIV and cancer patients (De Repentigny et al. 2004; Davies et al. 2006). Recent evidence suggests that the majority of such diseases produced by this pathogen are associated with biofilm growth (Ramage et al. 2005; Hasan et al. 2009; Dongari-Bagtzoglou et al. 2009).
The polymicrobial nature of oral biofilms associated with dental plaque and periodontitis has made them a pioneering model of interspecies interactions and highlights the level of complexity in biofilm research (Kuramitsu et al. 2007; Shirtliff et al. 2009). Such a complex interaction can be seen in the biofilms formed between C. albicans and S. epidermidis (Fig. 1). A study conducted by Harriott and Noverr (2009, 2010) on polymicrobial versus monomicrobial biofilms suggested that S. aureus may become coated in the matrix secreted by C. albicans. The enhancement in S. aureus resistance to vancomycin within the polymicrobial biofilm required viable C. albicans, and this was in part facilitated by C. albicans matrix. However, the growth or sensitivity to amphotericin B (AMB) of C. albicans was not altered in the polymicrobial biofilm. Peters et al. (2010) reported that the pathogenicity of S. aureus was increased due to its interaction with C. albicans in a mixed biofilm.

Scanning electron micrograph of a mixed-species biofilm of Candida albicans and Staphylococcus epidermidis. Smaller bacterial cells can be seen adherent to both yeasts and hyphae (Shirtliff et al. 2009)
Overall, biofilms are increasingly being recognized by the public health community as an important source of bacterial and fungal pathogens for all classes of patients, especially immunocompromised individuals and those with indwelling medical devices. Biofilm device infections can lead to significant morbidity and mortality, and may impair device function. Removal and/or replacement of devices are often the only treatment options, which can be very costly and also risky to the patients.
3 Biofilms: A Challenge for Antibiotic Therapy
The pathogenicity of biofilms is amplified by two of their major characteristics: (1) their increased tolerance to antimicrobials; (2) their protection of cells against the host’s defense mechanisms. Overall, the combined action of different mechanisms is believed to contribute to increased resistance and tolerance in biofilms: slow growth; phenotypic variation and differential regulation of the cell metabolic activity caused by nutrient limitation, stress, and cell density; over-expression of resistance genes and amplified expression of efflux pumps; a changing sterol composition in the membrane; limited diffusion of antibiotics and immunological molecules through the extracellular matrix; and presence of persisters in the biofilm, which are able to tolerate high concentrations of antibiotics.
Microbes within biofilms are significantly more resistant to standard antibiotic therapy and may require up to 1,000 times the antibiotic dose to achieve efficacy (Davies 2003; Lewis 2005). Therefore, the doses of antibiotics used effectively against planktonic cells are usually not enough to tackle biofilms, leading to resistant subpopulations remaining in the biofilm and causing recurring infections. This has led to a more judicious approach for antibiotic use in order to limit further development of resistant strains. The emergence of biofilm-associated infections and rise in resistant strains has threatened the efficacy of current antimicrobial agents, and therefore newer antimicrobial tools and strategies are needed to combat such infections. Here, we have reviewed some of the novel and successful strategies and approaches being used to prevent and control the biofilm infections.
4 Novel Strategies to Combat Biofilm-Associated Infections
Efforts to develop successful treatments for biofilm-associated infections are urgently needed in clinical practice. These new strategies must take into account the differences in physiology and antibiotic/host defense susceptibility of biofilm embedded microorganisms. The genetic and phenotypic versatility of the cells within biofilms represent a challenge for discovering new methods of treatment and prevention of biofilm-associated infections. Biofilm penetration by biocides or antibiotics is typically strongly hindered. To increase the efficiency of new treatment strategies against bacterial and fungal infections, factors that lead to inhibition of biofilm growth, disruption, or eradication of biofilms are being sought (Francolini and Donelli 2010). These factors include microbial products, enzymes, sodium salts, metal nanoparticles, antibiotics, acids, chitosan and its derivatives, or plant products. All of these factors influence biofilm structure via various mechanisms and with different efficiencies.
4.1 Use of Combination Therapy
Conventional therapies target individual microbial species without consideration that most biofilms are polymicrobial. However, a careful attempt should be made to identify the causative microorganisms in a biofilm community. Appropriate management of mixed infections requires the administration of antimicrobials that are effective against all the components of the biofilms. Many nosocomial infections involve microbial biofilms and persistence of chronic infections is attributed to the persistence of polymicrobial biofilms (Brogden and Guthmiller 2002; Hall-Stoodley and Stoodley 2009). The standard treatment for such infections involves two or more antibiotics, referred to as combination therapy (Brook 2002). The use of novel antibiotic combinations may increase the effectiveness of antibiotic therapies.
Another potential strategy could be to sensitize the bacteria or fungal biofilms by synthetic or natural compounds (other than antibiotics). For example, Jabra-Rizk et al. (2006) reported the sensitization of S. aureus bioflms by farnesol, a fungal QS molecule. The combined effect of gentamicin at 2.5 times the MIC and farnesol at 100 μM (22 μg/mL) was able to reduce bacterial populations by more than 2 log units and demonstrated a synergy between the two agents. This observed sensitization of resistant strains to antimicrobials and the observed synergistic effect with gentamicin indicates a potential application for farnesol as an adjuvant therapeutic agent for the prevention of biofilm-related infections. Using a combination approach we have demonstrated that the phenolic compounds eugenol and phenyl aldehyde cinnamdehyde potentiate the activity of flucoanzole against biofilm forming drug-resistant strains of C. albicans (Khan and Ahmad 2012a).
4.2 Prevention Against Catheter-Related Blood Stream Infections
Biofilms play a pivotal role in healthcare-associated infections, especially those related to the implantation of medical devices, such as intravascular catheters, urinary catheters, and orthopaedic implants. Implants act as passive surfaces prone to bacterial adhesion and biofilm formation. This tendency can result in implant-associated infection of the surgical site. In spinal surgery, implant-associated deep body infections are still a major problem (Trampuz and Widmer 2006). Some bacteria produce slime, which is responsible for bacterial adhesion and formation of biofilms on artificial surfaces. This slime is composed of proteins, hexosamines, neutral sugars, and phosphorus-containing compounds. If slime-forming bacteria colonize an artificial surface and develop a biofilm, this layer protects the bacteria from antibiotic agents. Thus, treatment against implant-associated infection must target the development of a biofilm (Secinti et al. 2011).
The most successful approaches for the control and prevention of infections due to adhesion, colonization, and biofilm formation on medical devices have been described in a review article by Francolini and Donelli (2010). Readers are suggested to go through this article for more detailed strategies currently in use for preventing biofilm formation on medical implants. In this chapter we will be reviewing developments in novel strategies to prevent biofilm infection of implants and tissues.
4.2.1 Lock Therapy Approach
Nosocomial infections associated with medical devices represent a large proportion of all cases of hospital-acquired infections (Bell 2001). In particular, insertion of any vascular catheter can result in a catheter-related infection, as microorganisms can colonize external and internal catheter surfaces. Adherence to the catheter surface is facilitated by host proteins such as fibronectin and fibrinogen, which can then lead to biofilm formation (Christner et al. 2010). Such problems can be overcome by one of the approaches termed lock therapy. This approach is currently recommended and employed in treating catheter-related bloodstream infections (CRBSI), in particular for long-term catheters, according to the Infectious Diseases Society of America’s guidelines (Mermel et al. 2009). The choice of antibiotics used in the lock technique is dependent on the pathogen suspected of infecting the catheter lumen, characteristics of the organism (i.e., ability to produce slime, adherence to host proteins), and the pharmacodynamic properties of the antimicrobial agent. Lock therapy involves the coating of high doses of an antimicrobial agent [from 100- to 1,000-fold the minimal inhibitory concentration, (MIC)] directly into the catheter in order to “lock” it for a certain period of time (from hours to days) (Carratala 2002). If host proteins such as fibronectin, fibrinogen, and fibrin are present in the catheter lumen, heparin may increase the efficacy of the antibiotics. Liposomal AMB and echinocandisn have been used successfully in a rabbit model of C. albicans biofilm infection (Schinabeck et al. 2004; Donlan 2008). While these results are promising for potential use of the lock technique to treat infected catheters, 100 % biofilm inhibition could not be achieved (Tournu and Dijck 2012). Synergistic antibiofilm combinations, between classical antimicrobial agents and other compounds such as the mucolytic agent N-acetylcysteine, ethanol, or the chelating agent EDTA, are being used as lock solutions and appeared to be very effective against S. epidermidis and C. albicans individual and mixed biofilms (Venkatesh et al. 2009). In a similar approach, recent findings suggest that the combination of antibacterial agents with Gram-positive activity, including doxycycline and tigecycline, with known antifungals, such as AMB, caspofungin, and fluconazole, can be useful for the treatment of C. albicans biofilms (Miceli et al. 2009; Ku et al. 2010).
The prevention of CRBSI has also been the focus of research and randomized controlled trials. The clinical effectiveness of central venous catheters (CVCs) treated with anti-infective agents (AI-CVC) in preventing CRBSI has been shown by Hockenhull et al. (2009). Antifungal impregnated CVCs have also been tested in animal models. Caspofungin was employed to prevent C. albicans biofilm formation in a murine biofilm model. C. albicans biofilm formation was reported to be greatly reduced in CVCs that had been pretreated for 24 h with high doses of caspofungin (Lazzell et al. 2009). The antibiofilm potential of liposomal AMB as a lock solution to inhibit C. albicans, Candida glabrata, and Candida parapsilosis biofilms in vitro has been reported by Toulet et al. (2012). Thus, the use of the lock technique or preventive impregnation of antifungals in combating catheter-associated infection seems promising, but not yet convincing from a cost-effective point of view, as huge doses are still needed to eradicate microbial growth.
4.2.2 Material Coatings and Novel Antibiofilm Surfaces
Among the promising approaches to combat biofilm infections is the generation of surface modification of devices to reduce microbial attachment and biofilm development. Typically, this strategy uses the incorporation of antimicrobial agents to prevent colonization (Smith 2005). Implanted materials are prone to biofilm formation affecting health in general and duration of the implant in particular. Surface characteristics, such as roughness, free energy, and chemistry, can influence the type and the feature of the biofilms (Teughels et al. 2006). For example, C. albicans adhesion is enhanced if the roughness of denture materials is increased (Radford et al. 1998). Currently, coatings may be engineered to promote selective adhesion to cells or tissue in bone implants but not to microbes. They may also address the second phase of biofilm development involving QS, by inhibiting cell–cell communication signals (Bruellhoff et al. 2010; Xiong and Liu 2010). Biomaterial modifications are a way to prevent biofilm development and have been the focus of intense research. While most research has focused on bacterial biofilms, the efficacy of biomaterial modifications also appears to inhibit Candida biofilms (Tournu and Dijck 2012).
4.2.2.1 Surface Modifications
The surface properties of medical devices constitute a major factor contributing not only to their stability in the body but also to their performance and lifetime in vivo and their colonization by microorganisms. Accordingly, albumin adhesion to surfaces is potentially beneficial since it has been shown to prevent binding of microorganisms, while fibrinogen has the opposite effect (Anderson et al. 2008). Chemical grafting of polyethylene and polypropylene surfaces with functionalized cyclodextrins changes the protein adsorption profile of these polymers by promoting adsorption of albumin and reducing the adhesion of fibrinogen to the material surface (Nava-Ortiz et al. 2010). These modified substrates were able to incorporate the antifungal agent miconazole very well and retarded biofilm formation by C. albicans. Modified polyethylene and silicone rubbers proved to be very efficient in inhibiting C. albicans biofilm formation (Contreras-Garcia et al. 2011). These materials are cytocompatible and also capable of releasing considerable amounts of nalidixic acid for several hours. This may further potentiate efficacy of treated surfaces to prevent formation of biofilms.
Biofilms on voice prostheses consist of mixed populations. Modification of the silicone surface of the prostheses has been employed to limit C. albicans colonization, as opposed to incorporation of antimicrobial agents in order to avoid the occurrence of resistance (De Prijck et al. 2010a). Silicone disks grafted with C1 and C8 alkyl side chains demonstrated reduced microbial adherence and inhibited biofilm formation by C. albicans by up to 92 %. Similarly, grafting of silicone rubber with cationic peptides, such as the salivary peptide Hst5 and synthetic variants, inhibited biofilm formation by up to 93 %, in a peptide-dependent manner (De Prijck et al. 2010b).
Preconditioning surfaces with surfactants also has potential to prevent bacterial adhesion and inhibit formation of biofilms. Splendiani et al. (2006) screened 22 surfactants for their potential to increase the cell wall charge of a Burkholderia sp. strain and reduce the ability to attach and form biofilms. The authors demonstrated that some surfactants affected the development of flagella, demonstrating significant changes in the ability of bacteria to attach in the presence of the surfactant. In addition to surfactants, biosurfactants synthesized by microbes have also been used as coating agents for medical implants leading to a reduction in hospital infections caused by biofilm growth (Rodrigues et al. 2006).
4.2.2.2 Surface Coatings
Microbicidal or static materials have been employed to fabricate or coat the surfaces of medical devices and have a great potential in reducing or eliminating the incidence of biofilm-related infections. Studies have reported the use of several compounds and synthetic analogues to prevent biofilm formation such as farnesol, quaternary ammonium salts, and silver ions, which were shown to effectively inhibit both bacterial and fungal biofilm formation (Gottenbos et al. 2001; Hashimoto 2001; Jabra-Rizk et al. 2006; Shirtliff et al. 2009). One particular study highlighted the role of two quaternary ammonium silanes (QAS) to coat silicone rubber tracheoesophageal shunt prostheses, yielding a positively charged surface. One QAS coating [(trimethoxysilyl)-propyldimethyloctadecylammonium chloride] was applied through chemical bonding, while the other coating, Biocidal ZF, was sprayed onto the silicone rubber surface. This was the first report on the inhibitory effects of positively charged coatings of tracheoesophageal shunt prostheses on the viability of yeasts and bacteria in mixed biofilms (Oosterhof et al. 2006). Although the study initially aimed at reducing voice prosthetic biofilms, its relevance extends to all biomedical surfaces where mixed biofilms develop and become problematic. Similarly, dental resin material coated with thin-film polymer formulations containing the polyene antifungal nystatin, AMB, or the antiseptic agent chlorhexidine were used in C. albicans and mixed biofilm prevention (Redding et al. 2009).
The polysaccharide dextran is widely used in medicine and is also one of the main components of dental plaque. Cross-linked dextran disks soaked with AMB solutions, described as amphogel, killed fungi within 2 h of contact and could be reused for almost 2 months without losing their efficacy against C. albicans (Zumbuehl et al. 2007). This antifungal material is biocompatible and could be used to coat medical devices to prevent microbial attachment.
Another option is to coat biomaterial surfaces with organic molecules to prevent protein adsorption which may also inhibit biofilm formation (Njoroge and Sperandio 2009). Coating of medical material surfaces has been employed and tested with several types of coating molecules, including the naturally occurring polymer chitosan and antimicrobial peptides such as Histatin 5 (Hst5). Histatins, a family of histidine-rich cationic peptides, are secreted by the major salivary glands in humans, especially histatin 5, which possess significant antifungal properties. A recent study demonstrated that histatin 5 exhibited antifungal activity against C. albicans biofilms and to a lesser extent against C. glabrata biofilms developed on denture acrylic (Konopka et al. 2010).
Naturally occurring antimicrobial peptides are promising therapeutic agents against pathogens such as C. albicans. But they are difficult and expensive to produce in large quantities and are also often sensitive to protease digestion. Therefore, their development as coating agents has been hampered. The search for new and improved antimicrobial peptides has led to the study of peptide mimetics. Synthetic analogs that mimic the properties of these peptides have many advantages and exhibit potent and selective antimicrobial activity (Tew et al. 2002). New classes of antimicrobial peptides were designed to mimic transmembrane segments of integral membrane proteins and were tagged with lysine residues to facilitate solubilization in aqueous media. These peptides, designated kaxins, have a non-amphipathic hydrophobic core segment, which distinguishes them from many natural linear cationic antimicrobial peptides. With this peptide Stark et al. (2002) showed that placing all of the K residues on the N-terminus and generating all-D enantiomeric versions, in combination with decreasing the length of the hydrophobic segment, resulted in shorter peptides that generally displayed increased antimicrobial activity. Generation of these shorter peptides is cost-effective and has shown potential applications in surface coatings. Karlsson and coworkers showed in 2009 that β-peptides (β-amino acid oligomers), at a concentration near the MIC, completely inhibited C. albicans planktonic cells from forming a biofilm by a toxicity mechanism involving membrane disruption. The same group reported in 2010 that fabrication of multilayered polyelectrolyte thin films promoted the surface-mediated release of an antifungal β-peptide. These films inhibited the growth of C. albicans on film-coated surfaces. In addition, β-peptide-containing films inhibited hyphal elongation by 55 %. This approach could ultimately be used to coat the surfaces of catheters, surgical instruments, and other devices to inhibit drug-resistant C. albicans biofilm formation in clinical settings (Karlsson et al. 2010). The utility and potential of selected peptides as therapeutic molecules, including the β-glucan synthesis inhibitors, the histidine-rich peptides, and the LL-37 cathelicidin family, are being determined and could be used as coating compounds against adherence and biofilm formation (Matejuk et al. 2010; Tsai et al. 2011).
Chitosan, a polymer isolated from crustacean exoskeletons, recently proved to be active against Candida biofilms in vitro. Surfaces coated with chitosan reduced the viable cell number in biofilms by more than 95 % in the case of C. albicans and also for many bacteria such as S. aureus (Carlson et al. 2008). Chitosan is a hydrophilic biopolymer that is industrially obtained by means of N-deacetylation of crustacean chitin. It is active against a wide range of pathogenic microbes including fungi, bacteria, and viruses (Rabea et al. 2003) by disrupting cell membranes as cells settle on to its surface. The use of such polymers offers a biocompatible tool for coating medical devices. Chitosan has been used to pretreat catheters and prevent C. albicans biofilm formation as validated in an in vivo CVC biofilm model by Martinez et al. (2010). The investigators demonstrated that mature C. albicans and C. parapsilosis biofilms were susceptible to chitosan in vitro. Chitosan decreased the metabolic activity and survival of Candida species biofilms, with more than 95 % killing of the sessile cells after 0.5 h treatment with 2.5 mg/mL chitosan.
4.2.2.3 Use of Nanoparticles
Nanotechnology is providing new ways to manipulate the structure and chemistry of surfaces to inhibit bacterial colonization. It is a new discipline with many applications in biological sciences and medicine, and is discussed in detail in other chapters included in this book. Nanomaterials are applied as coating materials, as well as in treatment and diagnosis (Colvin 2003). The advantages of nanoparticles are their high surface-to-volume ratios, quantum confinement, and nanoscale sizes. These properties allow more active sites of nanoparticles to interact with biological systems, including bacteria and fungi. This is the most important difference between nanoparticles and typical antimicrobial agents and could minimize the risk of developing antimicrobial resistance (Hernandez-Delgadillo et al. 2012). The mechanism of antimicrobial activity for nanoparticles is not completely understood. However, the positive charge of metal ions is known to be critical for antimicrobial activity, because it allows for their electrostatic attraction with the negative charge of the bacterial cell membrane. It has been reported that silver nanoparticles can damage DNA, alter gene expression, and affect membrane-bound respiratory enzymes (Kim et al. 2007). Nanoparticles of titanium, silver, copper oxide, selenium diamond, iron oxide, carbon nanotubes, and biodegradable polymers have also been studied for their use in diagnosis and treatment and their reported antimicrobial activities are summarized below.
As shown in Fig. 2, incorporation of cross-linked quaternary ammonium polyethylenimine (QPEI) nanoparticles in dental resin composite at a low concentration exerted a significant in vivo antibiofilm activity and potent broad spectrum antibacterial activity against salivary bacteria (Beyth et al. 2010). The antibacterial and antifungal effects of silver ions have long been known, and silver seems to inhibit biofilm formation by S. epidermidis, P. aeruginosa, and Candida spp. as evident from various studies (Kalishwaralal et al. 2010; Secinti et al. 2011; Monteiro et al. 2011a, b). These findings highlighted that nanoparticle silver ion-coated titanium implants are safe and provide a means to treat Candida-associated denture stomatitis. Recently, inhibition of biofilm formation by a S. aureus clinical isolate, with silver nanoparticle-coated catheters, was reported by Namasivayam et al. (2012). They also found that these naonoparticles exhibited synergistic effects to eradicate biofilms with the antibiotics ofloxacin, cephalexin, and neoflaxin.

Biofilms formed on resin composite incorporating QPEI nanoparticles and on nonmodified resin composite. Scanning electron micrographs (×10,000) of biofilms formed on resin composite (a) and resin composite with incorporated QPEI nanoparticles (b) (Beyth et al. 2010)
In another study, glass slides coated with zinc oxide (ZnO) nanoparticles restricted the biofilm formation of common bacterial pathogens. The generation of hydroxyl radicals, originating from the coated surface, was found to play a key role in antibiofilm activity (Applerot et al. 2012). Functionalized magnetite (Fe3O4/C18) nanoparticles have the potential to improve the antibiofilm properties of textile dressings against C. albicans biofilms (Anghel et al. 2012). In addition, these functionalized surface-based approaches are very useful in the prevention of wound microbial contamination and subsequent biofilm development on viable tissues or implanted devices. Recently, zerovalent stable colloidal bismuth nanoparticles were shown to possess antimicrobial activity against S. mutans and C. albicans growth and completely inhibited their biofilm formation. The results are similar to those obtained with chlorhexidine, the most commonly used oral antiseptic agent, and suggest that zerovalent bismuth nanoparticles could be an interesting antimicrobial agent to incorporate into an oral antiseptic preparation (Hernandez-Delgadillo et al. 2012, 2013a, b).
4.3 Disruption of Biofilms
Since biofilms must release and disperse cells into the environment in order to colonize new sites (Kaplan 2010), biofilm dispersal is another promising area of research that may lead to the development of novel agents to promote biofilm cell detachment. Furthermore, since the biofilm matrix also contains polysaccharides and DNA, a promising strategy could be the use of enzymes (e.g., DNase and alginate lyase) that can disrupt and dissolve biofilms by attacking surface polysaccharides and the extracellular DNA which is critical for the early development of biofilms (Arciola 2009; Taraszkiewicz et al. 2013).
Xavier et al. (2005) proposed a kinetic model to assess the feasibility of strategies for the removal of biofilms by using substances that induce detachment by affecting the cohesiveness of the EPS. Detachment-promoting agents are enzymes, chelating agents, or any other agents that reduce EPS cohesiveness through a variety of mechanisms. Promoting detachment is the least investigated of the possible strategies to remove unwanted biofilms. However, the use of substances to induce biofilm removal directly by destroying the physical integrity of the biofilm matrix would be an attractive alternative for both medical and industrial applications where complete biofilm removal is essential. This approach could also overcome the problem of recalcitrant infections of biofilms due to persister cells.
4.3.1 Biofilm-Disrupting Enzymes
The two most well-studied biofilm-dispersing enzymes are deoxyribonuclease I (DNase I) and dispersin B (DspB) (Kaplan 2009), but other extracellular enzymes have also been explored as antibiofilm agents.
Deoxyribonuclease I
Deoxyribonuclease I (DNase I) degrades extracellular DNA (eDNA), a newly highlighted structural component of biofilms that confers firmness and stability. Tetz et al. (2009) reported a strong negative impact of DNase I on the structures of biofilms formed by Acinetobacter baumannii, Haemophilus influenzae, K. pneumoniae, E. coli, P. aeruginosa, S. aureus, and S. pyogenes. Using DNase I at a concentration of 10 μg/mL, they observed degradation of mature 24 h-old biofilms by 53.85, 52.83, 50.24, 53.61, 51.64, 47.65, and 49.52 %, respectively. Moreover, bacterial susceptibility to selected antibiotics (azithromycin, rifampin, levofloxacin, ampicillin, and cefotaxime) increased in the presence of 5 μg/mL DNase I.
The antibiofilm activity of DNase I (130 μg/mL) in combination with selected antibiotics toward C. albicans biofilms has been estimated by Martins et al. (2012). In their study, reduction of viable counts by 0.5 log10 units was observed for C. albicans biofilms incubated with DNase I. Treatment of C. albicans with AMB alone (1 μg/mL) resulted in a 1 log10 unit reduction in cell viability, which increased to 3.5 log10 units in combination with DNase I. At higher concentrations of AMB (>2 μg/mL) and DNase I, cell viability was reduced by 5 log10 units.
DispersinB
DispersinB is a naturally occurring N-acetylglucosaminidase enzyme produced by a periodontal disease-associated oral bacterium, Aggregatibacter actinomycetemcomitans. This 41 kDa enzyme consists of a single chain containing 361 amino acid residues and is a highly active and stable glycoside hydrolase that functions in a narrow pH range. DispersinB specifically hydrolyses the glycosidic linkages of poly-β-1, 6-N-acetylglucosamine in the polysaccharide adhesins of bacteria, which are needed for biofilm formation, and are present in the polysaccharide matrix of mature biofilms, without affecting bacterial growth (Itoh et al. 2005). Thus, it inhibits as well as disperses bacterial biofilms and has been reported to be active against biofilms produced by various organisms such as E. coli, S. aureus, S. epidermidis, and P. fluorescence (Itoh et al. 2005; Rohde et al. 2007).
Lysostaphin
Lysostaphin is a natural staphylococcal endopeptidase that can penetrate bacterial biofilms (Belyansky et al. 2011). Promising antibiofilm results have been obtained for lysostaphin. The antimicrobial properties of lysostaphin were analyzed by Walencka et al. (2005), who reported the biofilm inhibitory concentration (BIC) of the enzyme for various S. aureus and S. epidermidis clinical strains. In addition, the combined use of lysostaphin with oxacillin resulted in increased susceptibility of the biofilm-growing bacteria to the antibiotic. Likewise, Aguinaga et al. (2011) reported a synergistic effect of lysostaphin in combination with doxycycline leading to significantly increased antibiotic susceptibility against methicillin-resistant S. aureus (MRSA) and methicillin-sensitive S. aureus (MSSA) strains.
Other Enzymes
Alkawash et al. (2006) showed application for lyase in the destruction of biofilms made by two mucoid P. aeruginosa strains. Treatment of the biofilms with gentamycin (64 μg/mL) in combination with alginate lyase (20 U/mL) resulted in biofilm matrix liquefacation. Incubation of the biofilm with lyase and gentamycin for 96 h resulted in the complete eradication of the biofilm structure and living bacteria. The antibiofilm activity of α-amylases against strains of S. aureus was analyzed by Craigen et al. (2011). This enzyme effectively reduced biofilm formation in the case of S. aureus. Time-course experiments for S. aureus showed that biofilms were degraded by 79 % within 5 min and by 89 % within 30 min of incubation with α-amylases. Amylase at doses of 10, 20, and 100 mg/mL reduced biofilms by 72, 89, and 90 %, respectively, and inhibited matrix formation by 82 %. In addition, they also investigated antibiofilm activities of amylases from different biological sources. The most effective biofilm reduction was reported for the α-amylase isolated from Bacillus subtilis. Although enzymes derived from human saliva and sweet potato had no effect against preformed biofilms, all of the tested enzymes, regardless of origin, were highly effective in inhibiting biofilm formation. Lactonase was also identified as a potential antibiofilm agent. Kiran et al. (2011) showed that biofilms formed by P. aeruginosa strains exhibited growth inhibition of 68.8–76.8 % in the presence of the enzyme (1 U/mL). They also found that 0.3 U/mL of the enzyme disrupted the biofilm structure and led to increased ciprofloxacin and gentamycin penetration and antimicrobial activity.
4.3.2 Photodynamic Therapy
Another innovative approach to disrupt biofilms is to expose them to photodynamic substances (Njoroge and Sperandio 2009). Antimicrobial Photodynamic Therapy (APDT) consists of three major components: light, a chemical molecule known as a photosensitizer, and oxygen. Photodynamic therapy (PDT) is based on the concept that a certain nontoxic photoactivatable compound or photosensitizer (PS) can be preferentially localized in certain tissues. These photosensitizers can be excited by absorbing a certain amount of energy from light of the appropriate wavelength. After excitation, photosensitizers usually form a long-lived triplet-excited state that will then generate reactive oxygen species (ROS), such as singlet oxygen and superoxide from which energy can be transferred to biomolecules or directly to molecular oxygen, depending on the reaction type. This results in oxidation of biomolecules, especially proteins involved in transport and membrane structure, in microorganisms leading to cell damage and death (Hamblin and Hasan 2004). Recent studies have shown that antimicrobial effects can be obtained with the use of photosensitizers belonging to different chemical groups such as phenothiazine dyes [methylene blue (MB) and toluidine blue O (TBO)]; porphyrin and its derivatives, TMPyP (5-,10-,15-,20-tetrakis (1-methylpyridinium-4-yl)-porphyrin), tetra p-toluenesulfonate; fullerenes; and cyanines and its derivatives (Taraszkiewicz et al. 2013).
The phenothiazinium salts are most commonly used in the clinic, and combinations of MB or TBO together with red light are used to disinfect blood products, sterilize dental cavities and root canals, and treat periodontitis (Wainwright 2003) and also have been actively investigated for the eradication of bacterial biofilm growing on dental plaques and oral implants (Saino et al. 2010). Tri-meso (N-methyl-pyridyl), meso (N-tetradecyl-pyridyl) porphine (C14) was exploited for inactivation of two structurally distinct S. epidermidis biofilms grown on Ti6Al4V alloy by Saino et al. (2010). They also compared its photosensitizing efficiency with that of the parent molecule, tetra-substituted N-methyl-pyridyl-porphine (C1). Their data suggested that C14 is a potential photosensitizer for the inactivation of staphylococcal biofilms for many device-related infections which are accessible to visible light.
Kishen et al. (2010) evaluated the ability of a cationic, phenothiazinium photosensitizer, methylene blue (MB), and an anionic, xanthene photosensitizer, rose bengal (RB), to inactivate and disrupt biofilms produced by E. faecalis (OGIRF and FA 2-2). The role of a specific microbial efflux pump inhibitor (EPI), verapamil hydrochloride in the MB-mediated antimicrobial photodynamic inactivation (aPDI) of E. faecalis biofilms, was also investigated. Their results showed that APDT with cationic MB produced superior inactivation of E. faecalis strains in a biofilm along with significant destruction of the biofilm structure when compared to anionic RB (P < 0.05). The ability to inactivate biofilm bacteria was further enhanced when the EPI was used with MB (P < 0.001). These experiments demonstrated the advantage of a cationic phenothiazinium photosensitizer combined with an EPI to inactivate biofilm bacteria and disrupt biofilm structure as shown in Fig. 3.

The three-dimensional CLSM reconstruction of E. faecalis biofilms subjected to aPDI (inset shows the sagittal section). (a) The untreated biofilm. (b) The biofilm incubated with 100 μM RB followed by irradiation at 40 J/cm2. (c) The biofilm incubated with 100 μM MB followed by irradiation at 40 J/cm2 (The colors represent: green viable, red dead, yellow intermediate) (Kishen et al. 2010)
Collins et al. (2010) studied the effect of TMP on P.aeruginosa biofilms. In their study, a significant decrease in biofilm density was observed, and the majority of the cells within the biofilm were nonviable when 100 μM TMP and 10 min of irradiation (mercury vapor lamp, 220–240 J/cm2) were used. Moreover, the use of 225 μM TMP and the same light dose resulted in almost complete disruption and clearance of the biofilm. Biel et al. (2011a, b) demonstrated that MB-mediated APDT was highly effective in the photo-eradication of multispecies bacterial biofilms (multidrug-resistant P. aeruginosa and MRSA). They observed a significant decrease in CFU/mL (>6 log10 units) when 300 μg/mL MB and a light dose of 60 J/cm2 (diode laser, 664 nm) were used. The reduction was >7 log10 units when 500 μg/mL MB and two light doses of 55 J/cm2 separated by a 5-min break were used. Recently, Meire et al. (2012) observed a statistically significant 1.9 log10 reduction in the viable counts of E. faecalis biofilms treated with 10 mg/mL MB and exposed to a soft laser at an output power of 75 mW (660 nm) for 2 min.
Since APDT represents an alternative method of killing resistant pathogens, efforts have been made to develop delivery systems for hydrophobic drugs to improve the photokilling. In this regard a study was conducted by Ribeiro et al. (2013) to evaluate the photodynamic effect of chloro-aluminum phthalocyanine (ClAlPc) encapsulated in nanoemulsions (NE) on MRSA and MSSA suspensions and biofilms. Suspensions and biofilms were treated with different delivery systems containing ClAlPc. For biofilms, cationic NE-ClAlPc reduced cell metabolism by 80 and 73 % of susceptible and resistant strains, respectively. Although anionic NE-ClAlPc caused a significant CFU/mL reduction for MSSA and MRSA, it was not capable of reducing MRSA biofilm metabolism. Moreover, a very recent study has shown improved efficacy of twofold positively charged porphyrin (XF-73) in comparison to fourfold positively charged porphyrin [5,10,15,20-tetrakis(1-methyl-4-pyridyl)-21H,23H-porphine, tetra-p-tosylate salt] against C. albicans planktonic cells and biofilms (Gonzales et al. 2013). Overall, this therapy is a very effective means of biofilm disruption and may represent an alternative treatment for eradicating resistant strains.
4.4 Biofilm Control Through Microbial Interactions or Interference
The existence of multiple interspecies interactions or the simple production of a metabolite can interfere with biofilm formation and development (Rossland et al. 2005; Valle et al. 2006). Competition for substrates is considered to be one of the major evolutionary driving forces in the bacterial world, and numerous experimental data obtained in the laboratory, under controlled conditions, have shown how different microorganisms may effectively outcompete others because they are better able to utilize a given energy source (Simoes et al. 2007). P. aeruginosa and Candida in a dual species environment mutually suppress biofilm development, both quantitatively and qualitatively (Bandara et al. 2010). In their study, Bandara et al. found that P. aeruginosa attached to C. albicans hyphae in a mixed-species biofilm and killed the fungi, whereas the yeast forms could not be killed. Isolation and purification of microbial compounds that mediate these types of interactions could lead to the development of new antibiofilm agents.
Commensal bacteria are known to inhibit pathogen colonization; however, complex host–microbe and microbe–microbe interactions have made it difficult to gain a detailed understanding of the mechanisms involved in the inhibition of colonization (Wertheim et al. 2005). In an attempt to understand these relationships, Iwase et al. (2010) found that the serine protease Esp, secreted by a subset of S. epidermidis, which are commensals, inhibited biofilm formation and nasal colonization by S. aureus. Furthermore, Esp enhanced the susceptibility of S. aureus biofilms to immune system components. In vivo studies have also shown that Esp-secreting S. epidermidis eliminates S. aureus nasal colonization. These findings indicate that Esp hinders S. aureus colonization in vivo through a novel mechanism of bacterial interference, which could lead to the development of novel therapeutics to prevent S. aureus colonization and infection. Two of the most commonly used strategies, designed to exploit microbe–microbe interactions, are discussed below.
4.4.1 Use of Probiotics
A novel mechanism for prophylactic or therapeutic management of biofilm-associated diseases is by microbial interference, through the use of probiotics. Probiotics are live microbial supplements which beneficially affect the host by improving its microbial balance, producing metabolites which inhibit the colonization or growth of other microorganisms, or by competing with them for resources such as nutrients or space. The use of antibiotics and immunosuppressive drugs often causes alterations in the composition of host microflora particularly in the oral cavity and intestinal and urogenital tracts. Therefore, the introduction of beneficial microbial species is a very attractive option to reestablish the microbial equilibrium and prevent disease (Gupta 2009).
The most commonly used genera in probiotic preparations are Lactobacillus, Bifidobacterium, Escherichia, Enterococcus, Bacillus, Streptococcus, and Saccharomyces. The use of probiotics creates a biofilm niche less conducive to proliferation of pathogens and their virulence factors via immune modulation and pathogen displacement activity. This phenomenon has been shown to be effective in varied clinical conditions such as antibiotic-associated diarrhea and Helicobacter pylori infections (Gupta 2009; Dobrogosz et al. 2010). One example is use of lactobacilli to improve urogenital health in women. Four probiotic strains Lactobacillus rhamnosus GG, L. plantarum 299v, and L. reuteri strains PTA 5289 and SD2112 were shown to interfere with the biofilms of salivary Streptococcus mutans. This antimicrobial activity against S. mutans was found to be pH dependent (Soderling et al. 2011).
There have also been reports of the inhibitory effect of probiotic Enterococcus faecium WB2000 on biofilm formation by cariogenic streptococci. Dental caries is a very common chronic disease arising from the interplay among the oral flora, teeth, and dietary factors. The major etiological players are the two α-hemolytic “mutans group” streptococci: S. mutans and Streptococcus sobrinus. The effect of Lactobacillus acidophilus DSM 20079 as a probiotic strain on the adhesion of some of the selected streptococcal strains was reported by Tahmourespour and Kermanshahi (2011). This strain was used for its ability to inhibit biofilm formation among mutans and non-mutans oral streptococci. In the presence of the probiotic strain, streptococcal adhesion was reduced and this reduction was not significantly higher if the probiotic strain was inoculated before the oral bacteria. The Lactobacillus acidophilus had a significantly higher effect on adherence of mutans streptococci than non-mutans streptococci (p < 0.05). It is expected that adhesion reduction is likely due to bacterial interactions and colonization of adhesion sites by the probiotic strain before the streptococci. Adhesion reduction can be an effective way to decrease the cariogenic potential of oral streptococci. Moreover, the ability of E. faecium WB2000 and JCM5804 and Enterococcus faecalis JCM5803 to inhibit biofilm formation by seven laboratory oral streptococcal strains and 13 clinical mutans streptococcal strains was assayed by Suzuki et al. (2011). E. faecium WB2000 inhibited biofilm formation by 90.0 % (9/10) of the clinical S. mutans strains and 100 % (3/3) of the clinical S. sobrinus strains.
4.4.2 Use of Phages as Antibiofilm Agents
Phages are ubiquitous in nature. Bacteriophages are viruses that infect bacteria and may provide a natural, highly specific, nontoxic, feasible approach for controlling several microorganisms involved in biofilm formation (Kudva et al. 1999). The ability of these phages to inhibit and/or eradicate biofilms has been demonstrated for biofilms of several pathogens including P. aeruginosa, K. pneumonia, E. coli, Proteus mirabilis, and S. epidermidis, and these studies are summarized here briefly.
Biofilms of E. coli strains 3000 XIII developed on the surfaces of polyvinylchloride coupons in a modified Robbins device were infected and lysed using bacteriophage T4D. Similar studies with phage E79 infecting P. aeruginosa indicated that phages were infecting the surface organisms but access to the cells deep in the biofilm was restricted (Doolittle et al. 1995). Investigators have demonstrated the use of bacteriophages in killing S. aureus and P. fluorescens biofilms; however, the infection of biofilm cells by phages is extremely conditional on their chemical composition and environmental factors such as temperature, growth stage, media, and phage concentration (Sillankorva et al. 2004; Chaignon et al. 2007).
The crucial role of titers of specifically selected phages with a proper virion-associated exopolysaccharide (EPS) depolymerase in the development of phage therapy were shown by Cornelissen et al. (2011). They carried out an experiment to investigate the in vitro degradation of single-species Pseudomonas putida biofilms, PpG1 and RD5PR2, by the novel phage Q15, a “T7-like virus” EPS depolymerase. Phage Q15 formed plaques surrounded by growing opaque halo zones, on seven out of 53 P. putida strains. This has happened because of EPS degradation. Since halos were absent on infection-resistant strains, they suggested that the EPS probably acts as a primary bacterial receptor for phage infection. EPS degrading activity of recombinantly expressed viral tail spike was also confirmed by capsule staining.
Application of bacteriophages in controlling mixed biofilms of Pseudomonas fluorescens and Staphylococcus lentus has also been reported. Sillankorva et al. (2010) challenged the biofilms with phage phiIBB-PF7A, specific for P. fluorescens, and the results obtained showed that phiIBB-PF7A readily reached the target host and caused a significant population decrease. This phage was also capable of causing partial damage to the biofilms leading to the release of the non-susceptible host (S. lentus) from the dual species biofilms.
Phage therapy has been successfully employed in the treatment of lung infections of cystic fibrosis caused by colonization of S. aureus and further predominant growth of P. aeruginosa biofilms. The treatment is very difficult with antibiotics due to several fold increased drug resistance (Brussow 2012). Applications of bacteriophages φMR299-2 and φNH-4 eliminated P. aeruginosa in the murine lung and cystic fibrosis lung airway cells (Alemayehu et al. 2012).
Recently, potential of the bacteriophage-derived peptidase, CHAPK, for the rapid disruption of biofilm was reported against staphylococci, associated with the bovine mastitis (Fenton et al. 2013). Purified CHAPK was able to prevent biofilm formation and also completely eliminated biofilms of S. aureus DPC5246 within 4 h. The CHAPK lysin also reduced S. aureus in a skin decolonization model. Furthermore, Shen et al. (2013) found rapid degradation of S. pyogenes biofilms by PlyC, a bacteriophage-encoded endolysin. Laser scanning confocal microscopy revealed that lytic action of PlyC destroys the biofilm as it diffuses through the matrix in a time-dependent fashion, and biofilm rapidly become refractory to traditional antibiotics.
Phage therapy is very effective in killing drug-resistant strains because of its specificity toward particular bacterial populations. Formation of a protected biofilm environment is one of the major causes of the increasing antibiotic resistance development. These facts emphasize the need to develop alternative antibacterial strategies, like phage therapy (Cornelissen et al. 2011).
4.5 Nature’s Own Biofilm Inhibitors
Interest in studying natural products derived from plant sources for the discovery of new biologically active compounds is not uncommon as many traditional medicines have been rooted. Some of the most active antibiofilm compounds discovered to date have been based upon the molecular scaffolds of natural products isolated from marine natural products (Worthington et al. 2012).
4.5.1 Plant Products
The prevention or control of biofilms by interfering with QS systems is one possible strategy; however, other studies have indicated that phytochemicals can inhibit interspecies coaggregation (Weiss et al. 1998), prevent bacterial adhesion (Kuzma et al. 2007), and inactivate mature single and multispecies biofilms (Niu and Gilbert 2004; Knowles et al. 2005). There is a novel trend in the antibiofilm research area toward the identification of natural products, such as plants and their extracts with antibiofilm activity. Plants offer a virtually inexhaustible and sustainable resource of very interesting classes of biologically active, low-molecular weight compounds. Several microbes in complex ecological niches or in association with biofilms produce compounds that act as antibiofilm agents to gain advantage over others. Certain marine plants are known to produce compounds that inhibit biofilm formation in order to prevent microbes from attaching and blocking the sunlight. The best characterized example is the red algae Delisea pulchra that produces halogenated furanones to ward off bacterial biofilms (Ren et al. 2004). Several marine plants and microbes have been shown to inhibit biofilm formation.
Plant extracts and essential oils from several medicinal plants have been exploited as antibiofilm agents for pathogenic biofilm forming bacteria and fungi. In this respect, xanthorrhizol isolated from Curcuma xanthorrhiza (Rukayadi et al. 2011) and the oil of Boesenbergia pandurata rhizomes (Taweechaisupapong et al. 2010) and Ocimum americanum (Thaweboon and Thaweboon 2009) showed potent in vitro activity against Candida biofilms. Nostro et al. (2007) studied the effect of oregano essential oil, carvacrol, and thymol on biofilm made by S. aureus and Staphylococcus epidermidis strains. They found that sub-inhibitory concentrations of the oils attenuated biofilm formation by S. aureus and S. epidermidis strains on polystyrene microtiter plates. Agarwal et al. (2008) studied 30 plant oils for their activity against C. albicans biofilms. Peppermint, eucalyptus, ginger grass, and clove oils resulted in a reduction in C. albicans biofilm formation. Dalleau et al. (2008) performed a study on 10 terpenic derivatives, corresponding to major components of essential oils, for their activity against C. albicans biofilms. Almost all the studied terpenic derivatives showed antibiofilm activity; however, carvacrol, geraniol, and thymol exhibited the strongest activity. Moreover, these compounds also proved to be efficient against biofilms made by C. glabrata and C. parapsilosis. In addition, Hendry et al. (2009) have shown potent antibiofilm activity from the main component of eucalyptus oil, 1,8-cineole, against C. albicans biofilms.
Harjai et al. (2010) reported anti-QS activity by fresh Allium sativum extract [fresh garlic extract (FGE)] and subsequently inhibited P. aeruginosa biofilm formation by 6 log10 units. Moreover, in vivo prophylactic treatment in a mouse model of kidney infection with FGE (35 mg/mL) for 14 days resulted in a 3 log10 unit decrease in the bacterial load on the fifth day after infection compared to untreated animals. They found that FGE also protected renal tissue from bacterial adherence and resulted in a milder inflammatory response and histopathological changes in infected tissues. FGE inhibited expression of P. aeruginosa virulence factors such as pyoverdin, hemolysin, and phospholipase C. Moreover, killing efficacy and phagocytic uptake of bacteria by peritoneal macrophages was enhanced by administration of garlic extract.
Issac Abraham et al. (2011) reported efficacy of Capparis spinosa (caper bush) extract to inhibit biofilm formation by 73 %, at a concentration of 2 mg/mL, in E. coli. Also, for the pathogens Serratia marcescens, P. aeruginosa, and P. mirabilis, biofilm biomass was reduced by 79, 75, and 70 %, respectively. Moreover, the mature biofilm structure was disrupted for all of the studied pathogens. Furthermore, the addition of C. spinosa extract (100 μg/mL) to a bacterial culture resulted in swimming and swarming inhibition. Similarly, Melia dubia (bead tree) bark extracts were examined by Ravichandiran et al. (2012) at a concentration of 30 mg/mL. In their study, these extracts reduced E. coli biofilm formation by 84 % and inhibited expression of virulence factors, such as hemolysins, by 20 %. Bacterial swarming regulated by QS was inhibited by 75 %, resulting in decreased biofilm expansion. Recently, our group (Khan and Ahmad 2012a, b) has shown antibiofilm activity by Cymbopogon citratus and Syzygium aromaticum essential oils and active compounds, namely cinnamaldehyde and eugenol, in drug-resistant strains of C. albicans (Fig. 4).

Scanning electron micrograph of the 48 h-old C. albicans 04 biofilm formed on catheter discs in the absence and presence of eugenol or cinnamaldehyde. In (a) biofilm formed in the absence of active compounds, dense network of cells and hyphae along with exopolysaccharide material are observed, and in (b) biofilm formed in the presence of eugenol at 100 mg/L, no exopolysaccharide material and aggregation of cells are observed (Khan and Ahmad 2012b)
4.5.1.1 Interference with Quorum Sensing
A new drug target is to interfere with the process of QS, a phenomenon of communication cross talk. This phenomenon is used by many pathogenic microorganisms to establish a biofilm and control much of their virulence arsenal. The process is regulated by means of extracellular signal molecules (Rasmussen et al. 2005). Although the exact role of QS in various stages of biofilm formation, maturation, and dispersal and in biofilm resistance is not entirely clear, the use of QS inhibitors (QSI) has been proposed as a potential antibiofilm strategy. It is conceivable that QS inhibition may represent a natural, widespread, antimicrobial strategy with significant impact on biofilm formation (Dong et al. 2002). Acting on biofilms by interfering with their command language, QS can provide an alternative to the ineffective conventional biofilm control strategies (Rasmussen and Givskov 2006).
QS has been shown to be responsible for the development of resistance to various antimicrobial agents and immune modulation in biofilm entities. Several organisms seem to have evolved the ability to interrupt this process. Examples include plants (e.g., tomato, rice, and pea) and soil bacteria that secrete compounds that alter homoserine lactone activity and Delisea pulchra, which secretes a halogenated furanone that inhibits QS signaling (Bauer and Robinson 2002). This suggests that synthetic analogs of such substances, or novel compounds from drug discovery efforts, could interrupt QS in one or more (Stewart 2003) ways. QS signaling can be interrupted in several manners like targeting to ligand-receptor pathways, i.e., by inhibiting ligand synthesis, transport, or release; inhibiting receptor synthesis and processing; and perhaps most analogous to current pharmacotherapy, inhibiting enzyme activity or ligand-receptor binding (Raffa et al. 2005). Furthermore, the use of QS inhibitors may control biofilm formation by making biofilms more susceptible to antibiotics as well as to host defenses (Bjarnsholt and Givskov 2007; Jayaraman and Wood 2008; Hoiby et al. 2010). Attenuation of bacterial virulence or biofilms by QSI rather than by antibiotics is a very interesting concept, which could prove to be a new target with less risk of inducing resistance (Lazar 2011). This strategy could lead to the development of new and efficient natural products for biofilm control.
Adonizio et al. (2008) have shown that certain plant extracts from Southern Florida caused the inhibition of QS genes and QS-controlled factors, with marginal effects on the growth of P. aeruginosa. Lonn-Stensrud et al. (2009) have shown that furanones may inhibit biofilm formation through interference with QS and thus represent promising agents for protecting surfaces from being colonized by S. epidermidis. Our group has also shown that pea seedling inhibits QS in P. aeruginosa PA01 and Cromobacterium violaceum CV12472 (Fatima et al. 2010).
Ding et al. (2011) screened 46 active components found in traditional Chinese medicines (TCMs) that inhibit bacterial biofilm formation. Six of 46 active components found in TCMs were identified as putative QSIs based on molecular docking studies. Of these, three compounds inhibited biofilm formation by P. aeruginosa and Stenotrophomonas maltophilia at a concentration of 200 μM. A fourth compound (emodin) significantly inhibited biofilm formation at 20 μM and induced proteolysis of the QS signal receptor TraR in Escherichia coli at a concentration of 3–30 mM. Emodin also increased the activity of ampicillin against P. aeruginosa. Therefore, they suggested that emodin might be suitable for development into an antivirulence and antibacterial agent based on disruption of biofilms. Brackman et al. (2011) have shown that QSI (baicalin hydrate, cinnamaldehyde, and hamamelitannin) increased the success of the antibiotics vancomycin, tobramycin, and clindamicin by increasing the susceptibility of bacterial biofilms and/or by increasing host survival following infection. Damte et al. (2013) screened for anti-QS activity in 97 indigenous plant extracts from Korea, through biomonitor bacterial strains, Chromobacterium violaceum (CV12472) and P. aeruginosa (PAO1), and found 18 plant extracts to exhibit anti-QS activity against both reporter systems. Sarabhai et al. (2013) have published a first report on the anti-QS activity of ellagic acid derivative compounds from T. chebula fruit. They found that these compounds downregulated the expression of the lasI/R and rhlI/R genes with concomitant decreases in N-acyl homoserine lactones (AHLs) in P. aeruginosa PAO1 causing attenuation of its virulence and enhanced sensitivity of its biofilm to tobramycin.
These data confirm that plant and microbial products have anti-QS, antiseptic, and antivirulence factor properties and can easily inhibit biofilm formation as well as disrupt the mature biofilm structure. These natural products represent a virtually inexhaustible and sustainable source of biocide-free antibiofilm agents with novel targets, unique modes of action, and proprieties with potential for utilization in clinical perspectives. Testing sublethal concentrations of plant-derived compounds for disrupting microbial biofilms could be of great importance to reveal mechanisms other than killing activity to overcome the emergence of drug-resistant strains. This strategy could offer an elegant way to develop novel biocide-free antibiofilm strategies. The studies conducted in this regard and significance and future prospects are well reviewed by Villa and Cappitelli (2013), and readers are directed to this chapter to get more insight on plant-derived products as effective antibiofilm agents.
4.5.2 Microbial Metabolites
The secondary metabolites of several microorganisms, ranging from furanone to exo-polysaccharides, have been suggested to have antibiofilm activity in various recent studies. Among these, E. coli group II capsular polysaccharides were shown to inhibit biofilm formation of a wide range of organisms, and marine Vibrio sp. were found to secrete complex exopolysaccharides having the potential for broad-spectrum biofilm inhibition and disruption (Abu Sayem et al. 2011). Extracts from coral associated Bacillus horikoshii (Thenmozhi et al. 2009) and actinomycetes (Nithyanand et al. 2010) inhibit biofilm formation of S. pyogenes. The exoproducts of marine Pseudoalteromonas impair biofilm formation by a wide range of pathogenic strains (Dheilly et al. 2010). Most recently, exopolysaccharides from the marine bacterium Vibrio sp. QY101 were shown to control biofilm-associated infections (Jiang et al. 2011). Abu Sayem et al. (2011) reported antibiofilm activity from a newly identified ca. 1,800 kDa polysaccharide, which has simple monomeric units of α-d-galactopyranosyl-(1→2)-glycerol-phosphate, against a number of both pathogenic and nonpathogenic strains without bactericidal effects. This polysaccharide was extracted from a Bacillus licheniformis strain associated with the marine organism Spongia officinalis. Musthafa et al. (2011) have shown the antibiofilm potential of ethyl acetate extract of marine Bacillus sp. SS4 using a static biofilm ring assay. Their study showed a concentration-dependent reduction in the biofilm-forming ability of PAO1 by these compounds. Members of the actinomycetes family are a rich source of bioactive compounds including diverse antibiotics.
4.5.2.1 Role of QS Molecules
Gram-negative bacteria predominantly use AHLs as autoinducers, which show variation in the length and oxidation state of the acyl side chain. In V. fischeri, AHL synthesis occurs when the luxI gene is activated to produce the AHL synthase enzyme LuxI. When these AHLs reach a threshold intracellular concentration, they bind to the transcriptional activator LuxR and lead to activation of the luxR gene set. AHLs are able to freely diffuse in and out of bacterial cells, allowing the total AHL concentration to correlate to the total bacterial concentration, thus enabling population density-based control of gene expression. This cascade of events ultimately leads to the control of gene expression resulting in the control of virulence factor production and biofilm formation and maintenance (Finch et al. 1998). A huge amount of work has been reported involving the biological consequences of chemically modified AHL derivatives in a variety of QS systems. Work from the Blackwell group has documented the synthesis and identification of a number of natural and synthetic AHLs with the ability to modulate QS in P. aeruginosa and Agrobacterium tumefaciens (Geske et al. 2005). They also found two of their most active synthetic AHLs retarded biofilm formation in P. aeruginosa PA01.
QS in Gram-positive bacteria is predominantly mediated by autoinducing peptides (AIPs) but may not exclusively utilize peptide signaling molecules for communication. Small molecules known as γ-butyrolactones have been identified as signaling molecules in some species of Streptomyces (Takano et al. 2001). The agr and TRAP (target of RNA-III activating peptide) QS systems in S. aureus regulate a number of virulence phenotypes, including biofilm formation. The RNA-III activating protein (RAP) activates TRAP via phosphorylation, leading to increased cell adhesion and biofilm formation, in addition to inducing expression of the agr operon (Fux et al. 2003). It has been demonstrated that the RNA-III inhibiting peptide (RIP) inhibits phosphorylation of TRAP, leading to reduced biofilm formation (Giacometti et al. 2003).
A small molecule termed autoinducer-2 (AI-2) is one of the putative universal QS mechanisms shared by both Gram-negative and Gram-positive bacteria. AI-2 molecules are derived from the precursor molecule (S)-4,5-dihydroxy-2,3-pentanedione (DPD), and the synthase enzyme that drives DPD production has been found to be conserved in over 55 bacterial species (Waters and Bassler 2005). An adenosine analogue of DPD was found to block AI-2-based QS without interfering with bacterial growth. This compound was subsequently shown to affect biofilm formation in Vibrio anguillarum, Vibrio vulnificus, and V. cholerae (Brackman et al. 2009).
Indole is a putative universal intercellular signal molecule amongst diverse bacteria that plays a direct role in the control of biofilm formation (Lee et al. 2007). Another attractive target for control of biofilm formation is interfering with c-di-GMP signaling using either c-di-GMP analogues or with small molecules that interfere with the synthesis or degradation of c-di-GMP (Sintim et al. 2010). Bis-(3′5′)-cyclic di-guanylic acid (c-di-GMP) is a second messenger signaling molecule that is thought to be ubiquitous in bacteria. Diguanylate cyclases (DGCs) and phosphodiesterases (PDEs) are responsible for the synthesis and breakdown of c-di-GMP, respectively (Yan and Chen 2010). There is increasing evidence that the transition between the planktonic and biofilm lifestyle of P. aeruginosa is regulated via proteins with DGC or PDE activities through control of c-di-GMP levels (Tamayo et al. 2007). It has also been observed that exopolysaccharide synthesis (and thus the exopolysaccharide-dependent formation of biofilms) is regulated by c-di-GMP in various proteobacterial species such as V. cholera, P. aeruginosa, P. fluorescens, A. tumefaciens, E. coli, and Salmonella enterica (Ryjenkov et al. 2005).
4.5.2.2 Role of Biosurfactants
Many bacteria are capable of synthesizing and excreting biosurfactants with anti-adhesive properties (Rodrigues et al. 2004; van Hamme et al. 2006). Biosurfactants are amphihilic biological compounds that are produced extracellularly or intracellularly by a wide variety of microorganisms, which include bacteria, yeasts, and filamentous fungi (Cameotra and Makkar 2004). These biosurfactants have promising applications in biomedical sciences (Singh and Cameotra 2004). Biosurfactants produced by Lactococcus lactis impaired biofilm formation on silicone rubber (Rodrigues et al. 2004). Surfactin from Bacillus subtilis dispersed biofilms without affecting cell growth and prevented biofilm formation by microorganisms such as Salmonella enterica, E. coli, and P. mirabilis (Mireles et al. 2001). Valle et al. (2006) demonstrated that E. coli expressing group II capsules released a soluble polysaccharide into their environment that induced physicochemical surface alterations, which prevented biofilm formation by a wide range of Gram-positive and Gram-negative bacteria. Many other researchers have demonstrated the potential for biofilm control by various other biosurfactants made by bacteria and fungi (Davey et al. 2003; Walencka et al. 2008). Two lipopeptide biosurfactants produced by B. subtilis and B. licheniformis have been shown by Rivardo et al. 2009 to exhibit anti-adhesive activity by selectively inhibiting biofilm formation of two human pathogenic strains, E. coli CFT073 and S. aureus ATCC29213. Davies and Marques (2009) found that P. aeruginosa produces cis-2-decenoic acid, which is capable of inducing the dispersion of established biofilms and of inhibiting biofilm development by B. subtilis, E. coli, S. aureus, Klebsiella pneumoniae, P. aeruginosa, P. mirabilis, S. pyogenes, and the yeast C. albicans, when applied exogenously. The authors also suggested that this molecule is functionally and structurally related to a class of short-chain fatty acid signaling molecules.
Fracchia et al. (2010) reported biofilm inhibitory activity by a Lactobacillus-derived biosurfactant against human pathogenic C. albicans. Rufino et al. (2011) have isolated a biosurfactant rufisan from the yeast Candida lipolytica UCP0988 that exhibited antimicrobial and anti-adhesive activities against many Streptococcus spp. Monteiro et al. (2011a, b) have evaluated the effects of a glycolipid-type biosurfactant produced by Trichosporon montevideense CLOA72 in the formation of biofilms in polystyrene plate surfaces by C. albicans CC isolated from the apical tooth canal. Biofilm formation was reduced up to 87.4 % with use of this biosurfactant at a 16 mg/mL concentration. This biomolecule did not present any cytotoxic effects in a HEK 293A cell line at concentrations of 0.25–1 mg/mL. Their studies indicated a possible application of the referred biosurfactant in inhibiting the formation of biofilms on plastic surfaces by C. albicans.
Recently, Padmapriya and Suganthi (2013) found antimicrobial and anti-adhesive activity of a biosurfactant produced by Candida tropicalis and C. albicans against a variety of urinary and clinical pathogens such as Bacillus, C. albicans, Citrobacter, E. coli, K. pneumoniae, P. mirabilis, P. aeruginosa, Salmonella, and S. aureus. A study from our group, Singh et al. (2013) has demonstrated Candida biofilm disrupting ability by di-rhamnolipid (RL-2) produced by P. aeruginosa DSVP20.
The antibiofilm activity of a glycolipid biosurfactant isolated from the marine actinobacterium Brevibacterium casei MSA19 was evaluated by Kiran et al. (2010) against pathogenic biofilms in vitro. The disruption of the biofilm by the MSA19 glycolipid was consistent against mixed pathogenic biofilm bacteria. Therefore, it could be suggested that the glycolipid biosurfactant can be used as a lead compound for the development of novel antibiofilm agents.
Janek et al. (2012) have recently identified a biosurfactant, Pseudofactin II, secreted by Pseudomonas fluorescens BD5, the strain obtained from freshwater from the Arctic Archipelago of Svalbard. Pseudofactin II showed anti-adhesive activity against several pathogenic microorganisms (E. coli, E. faecalis, Enterococcus hirae, S. epidermidis, P. mirabilis, and two C. albicans strains), which are potential biofilm formers on catheters, implants, and internal prostheses. Up to 99 % prevention was achieved by 0.5 mg/mL pseudofactin II. In addition, pseudofactin II dispersed preformed biofilms. Pseudofactin II can be used as a disinfectant or surface coating agent against microbial colonization of different surfaces, e.g., implants or urethral catheters.
An overview of all the above-discussed strategies to control biofilms is given in Fig. 5. The targets of each approaches at different stages of biofilms and their interrelation are depicted.

Depiction of targets of various approaches acting at different stages of biofilm growth and their interrelationship to control biofilm infections
4.6 Small Molecule Control of Biofilms
Given the prominence of biofilms in infectious diseases, there has been an increased effort toward the development of small molecules that inhibit and/or disperse bacterial biofilms through non-microbicidal mechanisms. It will be meaningful to distinguish molecules that have the ability to affect biofilm development via non-microbicidal mechanisms, as the pressure on bacteria or fungi to evolve resistance to these agents will be significantly reduced or even eliminated (Worthington et al. 2012). Due to the scarcity of known molecular scaffolds that inhibit/disperse bacterial biofilms, high throughput screening (HTS) has been employed in attempts to discover leads for new anti-biofilm modulators. Here, we have briefly summarized the application of chemical databases for the discovery of lead small molecules, using HTS approaches, which mediate biofilm development.
These approaches are grouped into three steps:
-
1.
The identification and development of small molecules that target one of the bacterial signaling pathways involved in biofilm regulation
-
2.
Chemical library screening for compounds with antibiofilm activity
-
3.
The identification of natural products that possess antibiofilm activity, and the chemical manipulation of these natural products to obtain analogues (using structure activity relationship (SAR) method) with increased activity.
Natural products provide a diverse array of chemical structures and possess a plethora of biological activities. A number of natural products that possess the ability to inhibit or disperse bacterial biofilms have been used as the starting points for medicinal chemistry programs in which synthetic manipulation of the natural product scaffold has allowed for the design of more efficacious compounds. Much of the natural product inspiration for these programs has come from compounds isolated from plants and marine organisms. It is known that QS pathways heavily influence the formation of biofilms, in addition to the production of other virulence factors. A diverse range of biomolecules serve as the facilitators for QS systems in bacteria. Therefore, extensive research in this area has produced a number of analogues with the ability to modulate QS-dependent enzymes. These molecules compose the vast majority of compounds thus far investigated for biofilm control. AHLs have served as one of the primary scaffolds studied over the past 30 years for the design of potential biofilm inhibitors (Geske et al. 2008). A considerable amount of work has been published involving the biological consequences of chemically modified AHL derivatives in a variety of QS systems and reviewed by Worthington et al. (2012). Here, we focus on using small molecules to derive novel compounds capable of controlling biofilm-associated infections (Fig. 6).

Principle of small molecule high throughput screening from natural products for development of anti-biofilm agents
4.6.1 Application of Chemical Library Screening
One of the first reports detailing the screening of a large library of compounds with the objective of identifying novel small molecules that possessed antibiofilm activity was reported from Biosignals (Sydney, Australia). It has developed over 200 furanone-like compounds and evaluated them as biofilm inhibitors in preventive therapy. Other natural product compounds from plants make up the material for Sequoia Sciences (San Diego, CA) to design biofilm inhibitors. They have developed a high-throughput strategy for extracting, purifying, and structurally characterizing libraries of natural product compounds from plants. They generated a library of over 150,000 natural compounds for evaluation as antibiofilm compounds (Sachachter 2003). In 2005, workers from the Wood group (Ren et al. 2005) screened 13,000 compounds. The study revealed a hit (0.08 %), identified as ursolic acid 57, as a compound which effectively inhibited E. coli biofilm formation at concentrations as low as 22 μM without affecting growth. In the same year, the Hergenrother group reported the identification of iron salts as effective nonantibiotic inhibitors and disruptors of P. aeruginosa biofilms from a screen of over 4,500 compounds which belonged to the University of Illinois Marvel Library Compound Collection (MLCC) (Musk et al. 2005).
Work from the Blackwell group has documented the synthesis and identification of a number of natural and unnatural AHLs with the ability to modulate QS in P. aeruginosa and Agrobacterium tumefaciens (Geske et al. 2005). They also demonstrated that two of their most active synthetic AHLs could retard biofilm formation in P. aeruginosa PA01. Other research that includes the modification of AHLs to discern their effects on QS and biofilm formation in P. aeruginosa comes from the Suga group. The work exploited the synthesis of a 96-member library constructed through solid phase protocols to mimic AHLs by replacing the homoserine lactone moiety with a variety of functionalities. A noteworthy compound identified within this study was AHL derivative 8, which had no effect on biofilm growth, yet elicited a noticeable change in the biofilm morphology of P. aeruginosa PA01 (Smith et al. 2003a, b). Analogues of P. aeruginosa AHLs in which the lactone functionality was replaced by a ketone had additional difluorination between the β-keto amide positions (Glansdorp et al. 2004).
Junker and Clardy (2007) have developed a HTS method for small molecule inhibitor of P. aeruginosa biofilms at the Institute of Chemistry and Cell Biology-Longwood (ICCB-L) at Harvard Medical School, Boston, MA (http://iccb.med.harvard.edu/). They have obtained 66,095 compounds, from natural products of microbial or plant origin and also some commercial chemical compounds, to identify those that prevent biofilm formation without affecting planktonic bacterial growth. The screen is a luminescence-based attachment assay that has been validated with several strains of P. aeruginosa and compared to a well-established but low-throughput crystal violet staining biofilm assay. They have determined the potencies of 61 compounds against biofilm attachment and have identified 30 compounds that fall into different structural classes as biofilm attachment inhibitors with 50 % effective concentrations of less than 20 μM. The most active compound discovered was shown to possess an IC50 value of 530 nM for biofilm inhibition. This makes this compound as one of the most active biofilm modulators ever disclosed against either Gram-positive or Gram-negative bacteria. Their study has highlighted these small-molecule inhibitors for identification of their relevant biofilm targets or potential therapeutics for P. aeruginosa infections.
A structure-based virtual screen (SB-VS) for the identification of putative QS inhibitors was carried out using a focused database comprising compounds that possess structural similarities to the known QS inhibitors furanone C30, patulin, the P. aeruginosa LasR natural ligand (3-oxo-C12-AHL 5), and a known QS receptor agonist TP-1, (Yang et al. 2009). This screen led to the discovery of three compounds, which were all recognized drugs, salicylic acid, nifuroxazide, and chlorzoxazone, and were subsequently shown to significantly inhibit QS-regulated gene expression at concentrations at which they did not affect bacterial growth. In addition to affecting QS regulated virulence factor production, these compounds were shown to affect biofilm formation by PA01. Screening of approximately 66,000 compounds and natural product extracts from the Center for Chemical Genomics at the University of Michigan to identify compounds that affected induction of a V. cholerae c-di-GMP-inducible transcriptional fusion led to the discovery of a novel benzimidazole (Sambanthamoorthy et al. 2011). This compound was examined for its ability to inhibit biofilm formation by a number of pathogenic bacterial strains. Compound 61 was shown to be a broad spectrum inhibitor of biofilm formation, significantly inhibiting biofilm formation by P. aeruginosa (CF-145), K. pneumoniae, Erwinia amylovora, and Shigella boydii at 100 μM, by MRSA USA300, and by S. aureus Newman at 25 μM, using the minimum biofilm eradication concentration (MBEC) static assay, without affecting bacterial growth. These signaling molecule derivatives are particularly important because the biological activity of nearly every compound in this class is not driven by microbicidal properties.
5 Conclusions
Biofilms have great importance for public health because of their role in certain infectious diseases and their importance in a variety of device-related infections. Most of our understanding of infections is based on research that has examined free-living organisms. The results do not necessarily apply to biofilm organisms, since metabolic and synthetic characteristics of free-living organisms can change when they assume the biofilm mode of growth. Microbial adhesion and biofilm formation are major concerns in control strategies. Drug resistance, virulence, and pathogenicity of microorganisms are often enhanced when growing in a biofilm, and new strategies are therefore required to control biofilm formation and development. A greater understanding of biofilm processes should lead to novel, effective control strategies for biofilm control and a resulting improvement in disease management. The similarity of QS processes to ligand-receptor binding could be exploited as a guide to direct novel antibiotic drug design efforts based on standard pharmacologic principles and drug discovery processes. The unique nature of their mechanism should provide these new antibiotics with greater activity against currently resistant bacteria. In addition, plant and microbial products in combination with other antimicrobial strategies such as antibiotics or photodynamic inactivation could provide an effective bactericidal tool for the treatment of various bacterial and yeast infections. Furthermore, development of high throughput methods to identify natural compounds or their analogues will be a promising strategy to overcome the problem of biofilm management.
References
Abu Sayem AM, Manzo E, Ciavatta L, Tramice A, Cordone A, Zanfardino A, De Felice M, Varcamonti M (2011) Anti-biofilm activity of an exopolysaccharide from a sponge-associated strain of Bacillus licheniformis. Microb Cell Fact 10:74
Adonizio A, Kong KF, Mathee K (2008) Inhibition of quorum sensing-controlled virulence factor production in Pseudomonas aeruginosa by South Florida plant extracts. Antimicrob Agents Chemother 52:198–203
Agarwal V, Lal P, Pruthi V (2008) Prevention of Candida albicans biofilm by plant oils. Mycopathologia 165:13–19
Aguinaga A, Francés ML, Del Pozo JL, Alonso M, Serrera A, Lasa I, Leiva J (2011) Lysostaphin and clarithromycin: a promising combination for the eradication of Staphylococcus aureus biofilms. Int J Antimicrob Agents 37:585–587
Alemayehu D, Casey PG, McAuliffe O, Guinane CM, Martin JG, Shanahan F, Coffey A, Ross RP, Hill C (2012) Bacteriophages φMR299-2 and φNH-4 can eliminate Pseudomonas aeruginosa in the murine lung and on cystic fibrosis lung airway cells. mBio 3:e00029-12
Alkawash MA, Soothill JS, Schiller NL (2006) Alginate lyase enhances antibiotic killing of mucoid Pseudomonas aeruginosa in biofilms. Acta Pathol Microbiol Immunol Scand 114:131–138
Anderson JM, Rodriguez A, Chang DT (2008) Foreign body reaction to biomaterials. Semin Immunol 20:86–100
Anghel I, Grumezescu AM, Andronescu E, Anghel AG, Ficai A, Saviuc C, Grumezescu V, Vasile BS, Chifiriuc MC (2012) Magnetite nanoparticles for functionalized textile dressing to prevent fungal biofilms development. Nanoscale Res Lett 7:501
Applerot G, Lellouche J, Perkas N, Nitzan Y, Gedanken A, Banin E (2012) ZnO nanoparticle-coated surfaces inhibit bacterial biofilm formation and increase antibiotic susceptibility. RSC Adv 2:2314–2321
Arciola C (2009) New concepts and new weapons in implant infections. Int J Artif Organs 32:533–536
Bandara HMHN, Yau JYY, Watt RM, Jin LJ, Samaranayake LP (2010) Pseudomonas aeruginosa inhibits in-vitro Candida biofilm development. BMC Microbiol 10:125
Bauer WD, Robinson JB (2002) Disruption of bacterial quorum sensing by other organisms. Curr Opin Biotechnol 13:234–237
Bell M (2001) Biofilms: a clinical perspective. Curr Infect Dis Rep 3:483–486
Belyansky I, Tsirline VB, Martin TR, Klima DA, Heath J, Lincourt AE, Satishkumar R, Vertegel A, Heniford BT (2011) The addition of lysostaphin dramatically improves survival, protects porcine biomesh from infection, and improves graft tensile shear strength. J Surg Res 171:409–415
Beyth N, Yudovin-Farber I, Perez-Davidi M, Domb AJ, Weiss EI (2010) Polyethyleneimine nanoparticles incorporated into resin composite cause cell death and trigger biofilm stress in vivo. Proc Natl Acad Sci USA 107:22038–22043
Biel MA, Sievert C, Usacheva M, Teichert M, Balcom J (2011a) Antimicrobial photodynamic therapy treatment of chronic recurrent sinusitis biofilms. Int Forum Allergy Rhinol 1:329–334
Biel MA, Sievert C, Usacheva M, Teichert M, Wedell E, Loebel N, Rose A, Zimmermann R (2011b) Reduction of endotracheal tube biofilms using antimicrobial photodynamic therapy. Laser Surg Med 43:586–590
Bjarnsholt T, Givskov M (2007) Quorum-sensing blockade as a strategy for enhancing host defences against bacterial pathogens. Philos Trans R Soc B 362:1213–1222
Brackman G, Celen S, Baruah K, Bossier P, Van Calenbergh S, Nelis HJ, Coenye T (2009) AI-2 quorum-sensing inhibitors affect the starvation response and reduce virulence in several Vibrio species, most likely by interfering with LuxPQ. Microbiology 155:4114–4122
Brackman G, Cos P, Maes L, Nelis HJ, Coenye T (2011) Quorum sensing inhibitors increase the susceptibility of bacterial biofilms to antibiotics in vitro and in vivo. Antimicrob Agents Chemother 55:2655–2661
Brogden KA, Guthmiller JM (2002) Polymicrobial diseases. ASM Press, Washington, DC
Brook I (2002) Microbiology of polymicrobial abscesses and implications for therapy. J Antimicrob Chemother 50:805–810
Bruellhoff K, Fiedler J, Oller MM, Groll J, Brenner RE (2010) Surface coating strategies to prevent biofilm formation on implant surfaces. Int J Artif Organs 33:646–653
Brussow H (2012) Pseudomonas biofilms, cystic fibrosis, and phage: a silver lining? mBio 3:e00061-12. doi:10.1128/mBio.00061-12
Cameotra SS, Makkar RS (2004) Recent applications of biosurfactants as biological and immunological molecules. Curr Opin Microbiol 7:262–266
Carlson RP, Taffs R, Davison WM, Stewart PS (2008) Anti-biofilm properties of chitosan-coated surfaces. J Biomat Sci 19:1035–1046
Carratala J (2002) The antibiotic-lock technique for therapy of “highly needed” infected catheters. Clin Microbiol Infect 8:282–289
Chaignon P, Sadovskaya I, Ragunah C, Ramasubbu N, Kaplan JB, Jabbouri S (2007) Susceptibility of staphylococcal biofilms to enzymatic treatments depends on their chemical composition. Appl Microbiol Biotechnol 75:125–132
Chole R, Faddis B (2003) Anatomical evidence of microbial biofilms in tonsillar tissues, a possible mechanism to explain chronicity. Arch Otolaryngol Head Neck Surg 129:634–636
Christner M, Franke GC, Schommer NN et al (2010) The giant extracellular matrix-binding protein of Staphylococcus epidermidis mediates biofilm accumulation and attachment to fibronectin. Mol Microbiol 75:187–207
Chuang YP, Fang CT, Lai SY, Chang SC, Wang JT (2006) Genetic determinants of capsular serotype K1 of Klebsiella pneumoniae causing primary pyogenic liver abscess. J Infect Dis 193:645–654
Coates T, Eady A, Cove J (2003) Propionibacterial biofilms cannot explain antibiotic resistance but might contribute to some cases of antibiotic recalcitrant acne. Br J Dermatol 148:366–367
Collins TL, Markus EA, Hassett DJ, Robinson JB (2010) The effect of a cationic porphyrin on Pseudomonas aeruginosa biofilms. Curr Microbiol 61:411–416
Colvin VL (2003) The potential environmental impact of engineered nanomaterials. Nat Biotechnol 21:1166–1170
Contreras-Garcia A, Bucio E, Brackmanc G, Coenye T, Concheiro A, Alvarez-Lorenzo C (2011) Biofilm inhibition and drug-eluting properties of novel DMAEMA-modified polyethylene and silicone rubber surfaces. Biofouling 27:123–135
Cornelissen A, Ceyssens PJ, T’Syen J, Van Praet H, Noben JP, Shaburova OV, Krylov VN, Volckaert G, Lavigne R (2011) The T7-related Pseudomonas putida phage Q15 displays virion-associated biofilm degradation properties. PLoS ONE 6:e18597. doi:10.1371/journal.pone.001 8597
Costerton JW, Stewart PS, Greenberg EP (1999) Bacterial biofilms: a common cause of persistent infections. Science 284:1318–1322
Craigen B, Dashiff A, Kadouri DE (2011) The use of commercially available alpha-amylase compounds to inhibit and remove Staphylococcus aureus biofilms. Open Microbiol J 5:21–31
Dalleau S, Cateau E, Berges T, Berjeaud JM, Imbert C (2008) In vitro activity of terpenes against Candida biofilms. Int J Antimicrob Agents 31:572–576
Damte D, Gebru E, Lee SJ, Suh JW, Park SC (2013) Evaluation of anti-quorum sensing activity of 97 indigenous plant extracts from Korea through bioreporter bacterial strains Chromobacterium violaceum and Pseudomonas aeruginosa. J Microb Biochem Technol 5:42–46
Davey ME, Caiazza NC, O’Toole GA (2003) Rhamnolipid surfactant production affects biofilm architecture in Pseudomonas aeruginosa PAO1. J Bacteriol 185:1027–1036
Davies D (2003) Understanding biofilm resistance to antibacterial agents. Nat Rev Drug Discov 2:114–122
Davies DG, Marques CN (2009) A fatty acid is responsible for inducing dispersion in microbial biofilms. J Bacteriol 191:1393–1403
Davies AN, Brailsford SR, Beighton D (2006) Oral candidosis in patients with advanced cancer. Oral Oncol 42:698–702
De Prijck K, Smet D, Coenye T, Schacht E, Nelis HJ (2010a) Prevention of Candida albicans biofilm formation by covalently bound dimethylaminoethylmethacrylate and polyethylenimine. Mycopathologia 170:213–221
De Prijck K, Smet D, Rymarczyk-Machal M, Van Driessche G, Devreese B, Coenye T, Schacht E, Nelis HJ (2010b) Candida albicans biofilm formation on peptide functionalized polydimethylsiloxane. Biofouling 26:269–275
Dheilly A, Soum-Soutera E, Klein GL, Bazire A, Compère C, Haras D, Dufour A (2010) Antibiofilm activity of the marine bacterium Pseudoalteromonas sp. strain 3J6. Appl Environ Microbiol 76:3452–3461
Ding X, Yin B, Qian L, Zeng Z, Yang Z, Li H, Lu Y, Zhou S (2011) Screening for novel quorum-sensing inhibitors to interfere with the formation of Pseudomonas aeruginosa biofilm. J Med Microbiol 60:1827–1834
Dobrogosz WJ, Peacock TJ, Hassan HM, Allen I. Laskin SS, Geoffrey MG (2010) Evolution of the probiotic concept: from conception to validation and acceptance in medical science. Adv Appl Microbiol 72:1–41
Dong YH, Gusti AR, Zhang Q, Xu JL, Zhang LH (2002) Identification of quorum-sensing N-acyl homoserine lactonases from Bacillus species. Appl Environ Microbiol 68:1754–1759
Dongari-Bagtzoglou A (2008) Pathogenesis of mucosal biofilm infections: challenges and progress. Expert Rev Anti Infect Ther 6:201–208
Dongari-Bagtzoglou A, Kashleva H, Dwivedi P, Diaz P, Vasilakos J (2009) Characterization of mucosal Candida albicans biofilms. PLoS ONE. doi:10.1371/ journal.pone.0007967
Donlan RM (2001) Biofilm formation: a clinically relevant microbiological process. Clin Infect Dis 33:1387–1392
Donlan RM (2002) Biofilms: microbial life on surfaces. Emerg Infect Dis 8:881e890
Donlan RM (2008) Biofilms on central venous catheters: is eradication possible? Curr Top Microbiol Immunol 322:133–161
Donlan RM, Costerton JW (2002) Biofilms: survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev 15:167–193
Doolittle MM, Cooney JJ, Caldwell DE (1995) Lytic infection of Escherichia coli biofilms by bacteriophage-T4. Can J Microbiol 41:12–18
Douglas LJ (2002) Medical importance of biofilms in Candida infections. Rev Iberoam Micol 19:139–143
Fatima Q, Zahin M, Khan MSA, Ahmad I (2010) Modulation of quorum sensing controlled behaviour of bacteria by growing seedling, seed and seedling extracts of leguminous plants. Indian J Microbiol 50:238–242
Fenton M, Keary R, McAuliffe O, Ross RP, O’Mahony J, Coffey A (2013) Bacteriophage-derived peptidase CHAPK eliminates and prevents staphylococcal biofilms. Int J Microbiol. Article ID 625341. doi:10.1155/2013/625341
Finch RG, Pritchard DI, Bycroft BW, Williams P, Stewart GS (1998) Quorum sensing: a novel target for anti-infective therapy. J Antimicrob Chemother 42:569–571
Fracchia L, Cavallo M, Allegrone G, Martinotti MG (2010) A Lactobacillus-derived biosurfactant inhibits biofilm formation of human pathogenic Candida albicans biofilm producers. In: Mendez-Vilas A (ed) Current research, technology and education topics in applied microbiology and microbial biochemistry. Formatex, Spain, pp 827–837
Francolini I, Donelli G (2010) Prevention and control of biofilm-based medical-device-related infections. FEMS Immunol Med Microbiol 59:227–238
Fux CA, Stoodley P, Hall-Stoodley L, Costerton JW (2003) Bacterial biofilms: a diagnostic and therapeutic challenge. Expert Rev Anti Infect Ther 1:667–683
Geske GD, Wezeman RJ, Siegel AP, Blackwell HE (2005) Small molecule inhibitors of bacterial quorum sensing and biofilm formation. J Am Chem Soc 127:12762–12763
Geske GD, O’Neill JC, Blackwell HE (2008) Expanding dialogues: from natural autoinducers to non-natural analogues that modulate quorum sensing in Gram-negative bacteria. Chem Soc Rev 37:1432–1447
Giacometti A, Cirioni O, Gov Y, Ghiselli R, Del Prete MS, Mocchegiani F, Saba V, Orlando F, Scalise G, Balaban N, Dell’Acqua G (2003) RNA III inhibiting peptide inhibits in vivo biofilm formation by drug-resistant Staphylococcus aureus. Antimicrob Agents Chemother 47:1979–1983
Glansdorp FG, Thomas GL, Lee JJ, Dutton JM, Salmond GP, Welch M, Spring DR (2004) Synthesis and stability of small molecule probes for Pseudomonas aeruginosa quorum sensing modulation. Org Biomol Chem 2:3329–3336
Gonzales FP, Felgentrager A, BaumLer W, Maisch T (2013) Fungicidal photodynamic effect of two fold positively charged porphyrin against Candida albicans planktonic cells and biofilms. Future Microbiol 8:785–797
Gottenbos B, van der Mei HC, Klatter F, Nieuwenhuis P, Busscher HJ (2001) In vitro and in vivo antimicrobial activity of covalently coupled quaternary ammonium silane coatings on silicone rubber. Biomaterials 23:1417–1423
Gupta VGR (2009) Probiotics. Indian J Med Microbiol 27:202–209
Hall-Stoodley L, Stoodley P (2009) Evolving concepts in biofilm infections. Cell Microbiol 11:1034–1043
Hall-Stoodley L, Costerton JW, Stoodley P (2004) Bacterial biofilms: from the natural environment to infectious diseases. Nat Rev Microbiol 2:95e108
Hamblin MR, Hasan T (2004) Photodynamic therapy: a new antimicrobial approach to infectious disease? Photochem Photobiol Sci 3:436–450
Harjai K, Kumar R, Singh S (2010) Garlic blocks quorum sensing and attenuates the virulence of Pseudomonas aeruginosa. FEMS Immunol Med Microbiol 58:161–168
Harriott MM, Noverr MC (2009) Candida albicans and Staphylococcus aureus form polymicrobial biofilms: effects on antimicrobial resistance. Antimicrob Agents Chemother 53:3914–3922
Harriott MM, Noverr MC (2010) Ability of Candida albicans mutants to induce Staphylococcus aureus vancomycin resistance during polymicrobial biofilm formation. Antimicrob Agents Chemother 54:3746–3755
Hasan F, Xess I, Wang X, Jain N, Fries BC (2009) Biofilm formation in clinical Candida isolates and its association with virulence. Microbes Infect 11:753–761
Hashimoto H (2001) Evaluation of the anti-biofilm effect of a new antibacterial silver citrate/lecithin coating in an in-vitro experimental system using a modified Robbins device. J Jpn Assoc Infect Dis 75:678–685
Hendry ER, Worthington T, Conway BR, Lambert PA (2009) Antimicrobial efficacy of eucalyptus oil and 1,8-cineole alone and in combination with chlorhexidine digluconate against microorganisms grown in planktonic and biofilm cultures. J Antimicrob Chemother 64:1219–1225
Hernandez-Delgadillo R, Velasco-Arias D, Diaz D, Arevalo-Niño K, Garza-Enriquez M, De la Garza-Ramos MA, Cabral-Romero C (2012) Zerovalent bismuth nanoparticles inhibit Streptococcus mutans growth and formation of biofilm. Int J Nanomed 7:2109–2113
Hernandez-Delgadillo R, Velasco-Arias D, Diaz D, Arevalo-Nino K, Garza-Enriquez M, Garza-Ramos MAD, Cabral-Romero C (2013a) Zerovalent bismuth nanoparticles inhibit Streptococcus mutans growth and formation of biofilm. Int J Nanomed 7:2109–2113
Hernandez-Delgadillo R, Velasco-Arias D, Diaz D, Arevalo-Nino K, Garza-Enriquez M, Garza-Ramos MAD, Cabral-Romero C (2013b) Bismuth oxide aqueous colloidal nanoparticles inhibit Candida albicans growth and biofilm formation. Int J Nanomed 8:1645–1652
Hockenhull JC, Dwan KM, Smith GW, Gamble CL, Boland A, Walley TJ, Dickson RC (2009) The clinical effectiveness of central venous catheters treated with anti infective agents in preventing catheter-related bloodstream infections: a systematic review. Crit Care Med 37:702–712
Hogan DA (2006) Talking to themselves: autoregulation and quorum sensing in fungi. Eukaryot Cell 5:613–619
Hoiby N, Bjarnsholt T, Givskov M, Molin S, Ciofu O (2010) Antibiotic resistance of bacterial biofilms. Int J Antimicrob Agents 35:322–332
Il’ina TS, Romanova YM, Gintsburg AL (2004) Biofilms as a mode of existence of bacteria in external environment and host body: the phenomenon, genetic control, and regulation systems of development. Genetika 40:1445–1456
Issac Abraham SV, Palani A, Ramaswamy BR, Shunmugiah KP, Arumugam VR (2011) Antiquorum sensing and antibiofilm potential of Capparis spinosa. Arch Med Res 42:658–668
Itoh Y, Wang X, Hinnebusch BJ, Preston JF III, Romeo T (2005) Depolymerization of β-1, 6-N-acetyl-D-glucosamine disrupts the integrity of diverse bacterial biofilms. J Bacteriol 187:382–387
Iwase T, Uehara Y, Shinji H, Tajima A, Seo H, Takada K, Agata T, Mizunoe Y (2010) Staphylococcus epidermidis Esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature 465:346–351
Jabra-Rizk MA, Meiller TF, James C, Shirtliff ME (2006) Effect of farnesol on Staphylococcus aureus biofilm formation and antimicrobial susceptibility. Antimicrob Agents Chemother 50:1463–1469
Jain AGY, Agrawal R, Khare P, Jain SK (2007) Biofilms a microbial life perspective: a critical review. Crit Rev Ther Drug Carrier Syst 24:393–443
Janek T, Łukaszewicz M, Krasowska A (2012) Antiadhesive activity of the biosurfactant pseudofactin II secreted by the Arctic bacterium Pseudomonas fluorescens BD5. BMC Microbiol 12:24
Jayaraman A, Wood TK (2008) Bacterial quorum sensing: signals, circuits, and implications for biofilms and disease. Annu Rev Biomed Eng 10:145–167
Jiang P, Li J, Han F, Duan G, Lu X, Gu Y, Yu W (2011) Antibiofilm activity of an exopolysaccharide from marine bacterium Vibrio sp. QY101. PLoS ONE 6:e18514
Junker LM, Clardy J (2007) High-throughput screens for small-molecule inhibitors of Pseudomonas aeruginosa biofilm development. Antimicrob Agents Chemother 51:3582–3590
Kalishwaralal K, BarathManiKanth S, Pandian SR, Deepak V, Gurunathan S (2010) Silver nanoparticles impede the biofilm formation by Pseudomonas aeruginosa and Staphylococcus epidermidis. Colloids Surf B Biointerfaces 79:340–344
Kaplan JB (2009) Therapeutic potential of biofilm-dispersing enzymes. Int J Artif Organs 32:545–554
Kaplan JB (2010) Biofilm dispersal: mechanisms, clinical implications and potential therapeutic uses. J Dent Res 89:205–218
Karlsson AJ, Pomerantz WC, Neilsen KJ, Gellman SH, Palecek SP (2009) Effect of sequence and structural properties on 14-helical betapeptide activity against Candida albicans planktonic cells and biofilms. ACS Chem Biol 4:567–579
Karlsson AJ, Flessner RM, Gellman SH, Lynn DM, Palecek SP (2010) Polyelectrolyte multilayers fabricated from antifungal β-peptides: design of surfaces that exhibit antifungal activity against Candida albicans. Biomacromolecules 11:2321–2328
Khan MSA, Ahmad I (2012a) Biofilm inhibition by Cymbopogon citratus and Syzygium aromaticum essential oils in the strains of Candida albicans. J Ethnopharmacol 140:416–423
Khan MSA, Ahmad I (2012b) Antibiofilm activity of certain phytocompounds and their synergy with fluconazole against Candida albicans biofilms. J Antimicrob Chemother 67:618–621
Kim JS, Kuk E, Yu KN, Kim JH, Park SJ, Lee HJ, Kim SH, Park YK, Park YH, Hwang CY, Kim YK, Lee YS, Jeong DH, Cho MH (2007) Antimicrobial effects of silver nanoparticles. Nanomedicine 3:95–101
Kishen A, Upadya M, Tegos GP, Hamblin MR (2010) Efflux pump inhibitor potentiates antimicrobial photodynamic inactivation of Enterococcus faecalis biofilm. Photochem Photobiol 86:1343–1349
Kiran GS, Sabarathnam B, Selvin J (2010) Biofilm disruption potential of a glycolipid biosurfactant from marine Brevibacterium casei. FEMS Immunol Med Microbiol 59:432–438
Kiran S, Sharma P, Harjai K, Capalash N (2011) Enzymatic quorum quenching increases antibiotic susceptibility of multidrug resistant Pseudomonas aeruginosa. Iran J Microbiol 3:1–12
Knowles JR, Roller S, Murray DB, Naidu AS (2005) Antimicrobial action of carvacrol at different stages of dual-species biofilm development by Staphylococcus aureus and Salmonella enterica Serovar Typhimurium. Appl Environ Microbiol 71:797–803
Kokare CR, Chakraborty S, Khopade AN, Mahadik KR (2009) Biolfilm: importance and applications. Indian J Biotechnol 8:159–168
Konopka K, Dorocka-Bobkowska B, Gebremedhin S, Duzguneş N (2010) Susceptibility of Candida biofilms to histatin 5 and fluconazole. Antonie Van Leeuwenhoek 97:413–417
Ku TSN, Palanisamy SKA, Lee SA (2010) Susceptibility of Candida albicans biofilms to azithromycin, tigecycline and vancomycin and the interaction between tigecycline and antifungals. Int J Antimicrob Agents 36:441–446
Kudva IT, Jelacic S, Tarr PI, Youderian P, Hovde CJ (1999) Biocontrol of Escherichia coli O157 with O157-specific bacteriophages. Appl Environ Microbiol 65:3767–3773
Kuramitsu HK, He X, Lux R, Anderson MH, Shi W (2007) Interspecies interactions within oral microbial communities. Microbiol Mol Biol Rev 71:653–670
Kuzma L, Ozalski MR, Walencka E, Ozalska BR, Wysokinska H (2007) Antimicrobial activity of diterpenoids from hairy roots of Salvia sclarea L.: salvipisone as a potential anti-biofilm agent active against antibiotic resistant Staphylococci. Phytomedicine 14:31–35
Lazar V (2011) Quorum sensing in biofilms: how to destroy the bacterial citadels or their cohesion/power? Anaerobe 17:280e285
Lazzell AL, Chaturvedi AK, Pierce CG, Prasad D, Uppuluri P, Lopez-Ribot JL (2009) Treatment and prevention of Candida albicans biofilms with caspofungin in a novel central venous cathetermurine model of candidiasis. J Antimicrob Chemother 64:567–570
Lee J, Jayaraman A, Wood TK (2007) Indole is an inter-species biofilm signal mediated by SdiA. BMC Microbiol 7:42
Lewis K (2005) Persister cells and the riddle of biofilm survival. Biokhimiya 70:327–336
Lonn-Stensrud J, Landin MA, Benneche T, Petersen FC, Scheie AA (2009) Furanones, potential agents for preventing Staphylococcus epidermidis biofilm infections? J Antimicrob Chemother 63:309–331
Mardh PA, Rodrigues AG, Genc M, Novikova N, Martinez-de-Oliveira J, Guaschino S (2002) Facts and myths on recurrent vulvovaginal candidosis—a review on epidemiology, clinical manifestations, diagnosis, pathogenesis and therapy. Int J STD AIDS 13:522–539
Martinez LR, Mihu MR, Tar M, Cordero RJ, Han G, Friedman AJ, Friedman JM, Nosanchuk JD (2010) Demonstration of antibiofilm and antifungal efficacy of chitosan against candidal biofilms, using an in vivo central venous catheter model. J Infect Dis 201:1436–1440
Martins M, Henriques M, Lopez-Ribot JL, Oliveira R (2012) Addition of DNase improves the in vitro activity of antifungal drugs against Candida albicans biofilms. Mycoses 55:80–85
Matejuk A, Leng Q, Begum MD, Woodle MC, Scaria P, Chou ST, Mixson AJ (2010) Peptide-based antifungal therapies against emerging infections. Drugs Future 35:197–217
Meire MA, Coenye T, Nelis HJ, De Moor RC (2012) Evaluation of Nd:YAG and Er:YAG irradiation, antibacterial photodynamic therapy and sodium hypochlorite treatment on Enterococcus faecalis biofilms. Int Endod J 45:482–491
Mermel LA, Allon M, DE Bouza C, Flynn P, O’Grady NP, Raad II, Rijnders BJ, Sherertz RJ, Warren DK (2009) Clinical practice guidelines for the diagnosis andmanagement of intravascular catheter-related infection: 2009 update by the infectious diseases society of America. Clin Infect Dis 49:1–45
Miceli MH, Bernardo SM, Lee SA (2009) In vitro analyses of the combination of high-dose doxycycline and antifungal agents against Candida albicans biofilms. Int J Antimicrob Agents 34:326–332
Mireles JR, Toguchi A, Harshey RM (2001) Salmonella enterica serovar Typhimurium swarming mutants with altered biofilm-forming abilities: surfactin inhibits biofilm formation. J Bacteriol 183:5848–5854
Molin S, Tolker-Nielson T (2003) Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced stabilisation of the biofilm structure. Curr Opin Biotechnol 14:255–261
Monteiro AS, Miranda TT, Lula I, Denadai AML, Sinisterra RD, Santoro MM, Santos VL (2011a) Inhibition of Candida albicans CC biofilms formation in polystyrene plate surfaces by biosurfactant produced by Trichosporon montevideense CLOA72. Colloids Surf B Biointerfaces 84:467–476
Monteiro DR, Gorup LF, Silva S, Negri M, de Camargo ER, Oliveira R, Barbosa DB, Henriques M (2011b) Silver colloidal nanoparticles: antifungal effect against adhered cells and biofilms of Candida albicans and Candida glabrata. Biofouling 27:711–719
Muller FM, Seidler M, Beauvais A (2011) Aspergillus fumigatus biofilms in the clinical setting. Med Mycol 49(Suppl 1):S96–S100
Musk DJ, Banko DA, Hergenrother PJ (2005) Iron salts perturb biofilm formation and disrupt existing biofilms of Pseudomonas aeruginosa. Chem Biol 12:789–796
Musthafa KS, Saroja V, Pandian SK, Ravi AV (2011) Antipathogenic potential of marine Bacillus sp. SS4 on N-acyl-homoserine-lactonemediated virulence factors production in Pseudomonas aeruginosa (PAO1). J Biosci 36:55–67
Namasivayam SKR, Preethi M, Bharani ARS, Robin G, Latha B (2012) Biofilm inhibitory effect of silver nanoparticles coated catheter against Staphylococcus aureus and evaluation of its synergistic effects with antibiotics. Int J Biol Pharm Res 3:259–265
Nava-Ortiz CAB, Burillo G, Concheiro A et al (2010) Cyclodextrin-functionalized biomaterials loaded with miconazole prevent Candida albicans biofilm formation in vitro. Acta Biomater 6:1398–1404
Nikolaev YA, Plankunov VK (2007) Biofilm- “city of microbes” or an analogue of multicellular organisms? Microbiology 76:125–138
Nithyanand P, Thenmozhi R, Rathna J, Pandian SK (2010) Inhibition of biofilm formation in Streptococcus pyogenes by coral associated Actinomycetes. Curr Microbiol 60:454–460
Niu C, Gilbert ES (2004) Colorimetric method for identifying plant essential oil components that affect biofilm formation and structure. Appl Environ Microbiol 70:6951–6956
Njoroge J, Sperandio V (2009) Jamming bacterial communication: new approaches for the treatment of infectious diseases. EMBO Mol Med 1:201–210
Nostro A, Roccaro AS, Bisignano G, Marino A, Cannatelli MA, Pizzimenti FC, Cioni PL, Procopio F, Blanco AR (2007) Effects of oregano, carvacrol and thymol on Staphylococcus aureus and Staphylococcus epidermidis biofilms. J Med Microbiol 56:519–523
Oosterhof JJH, Buijssen KJDA, Busscher HJ, van der Laan BFAM, van der Mei HC (2006) Effects of quaternary ammonium silane coatings on mixed fungal and bacterial biofilms on tracheoesophageal shunt prostheses. Appl Environ Microbiol 72:3673–3677
Padmapriya B, Suganthi S (2013) Antimicrobial and anti adhesive activity of purified biosurfactants produced by Candida species. Middle East J Sci Res 14:1359–1369
Peters BM, Jabra-Rizk MA, Scheper MA, Leid JG, Costerton JW, Shirtliff ME (2010) Microbial interactions and differential protein expression in Staphylococcus aureus–Candida albicans dual-species biofilms. FEMS Immunol Med Microbiol 59:493–503
Prince AS (2002) Biofilms, antimicrobial resistance, and airway infection. N Engl J Med 347:1110–1111
Rabea EI, Badawy ME, Stevens CV, Smagghe G, Steurbaut W (2003) Chitosan as antimicrobial agent: applications and mode of action. Biomacromolecules 4:1457–1465
Radford DR, Sweet SP, Challacombe SJ, Walter JD (1998) Adherence of Candida albicans to denture-base materials with different surface finishes. J Dent 26:577–583
Raffa RB, Iannuzzo JR, Levine DR, Saeid KK, Schwartz RC, Sucic NT, Terleckyj OD, Jeffrey M (2005) Young bacterial communication (“Quorum Sensing”) via ligands and receptors: a novel pharmacologic target for the design of antibiotic drugs. J Pharmacol Exp Ther 312:417–423
Ramage G, Saville DO, Lopez-Ribot JL (2005) Candida biofilm: an update. Eukaryot Cell 4:633–638
Rasmussen TB, Givskov M (2006) Quorum sensing inhibitors: a bargain of effects. Microbiology 152:895–904
Rasmussen TB, Skindersoe ME, Bjarnsholt T, Phipps RK, Christensen KB, Jensen PO, Andersen JB, Koch B, Larsen TO, Hentzer M, Eberl L, Hoiby N, Givskov M (2005) Identity and effects of quorum-sensing inhibitors produced by Penicillium species. Microbiology 151:1325–1340
Ravichandiran V, Shanmugam K, Anupama K, Fomas S, Princy A (2012) Structure-based virtual screening for plant-derived SdiA-selective ligands as potential antivirulent agents against uropathogenic Escherichia coli. Eur J Med Chem 48:200–205
Redding S, Bhatt HR, Rawls G, Siegel K, Scott Lopez-Ribot J (2009) Inhibition of Candida albicans biofilm formation on denture material. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 107:669–672
Ren D, Bedzyk LA, Ye RW, Thomas SM, Wood TK (2004) Stationary-phase quorum-sensing signals affect autoinducer-2 and gene expression in Escherichia coli. Appl Environ Microbiol 70:2038–2043
Ren DC, Zuo RJ, Barrios AF, Bedzyk LA, Eldridge GR, Pasmore ME, Wood TK (2005) Differential gene expression for investigation of Escherichia coli biofilm inhibition by plant extract ursolic acid. Appl Environ Microbiol 71:4022–4034
Repentigny D, Lewandowski D, Jolicoeur P (2004) Immunopathogenesis of oropharyngeal candidiasis in human immunodeficiency virus infection. Clin Microbiol Rev 17:729–759
Ribeiro APD, Andrade MC, Bagnato VS, Vergani CE, Primo FL, Tedesco AC, Pavarina AC (2013) Antimicrobial photodynamic therapy against pathogenic bacterial suspensions and biofilms using chloro-aluminum phthalocyanine encapsulated in nanoemulsions. Lasers Med Sci. doi:10.1007/s10103-013-1354-x
Rivardo F, Turner RJ, Allegrone G, Ceri H, Martinotti MG (2009) Anti-adhesion activity of two biosurfactants produced by Bacillus spp. prevents biofilm formation of human bacterial pathogens. Appl Microbiol Biotechnol 83:541–553
Rodrigues LR, van der Mei HC, Teixeira JA, Oliveira R (2004) Biosurfactant from Lactococcus lactis 53 inhibits microbial adhesion on silicone rubber. Appl Microbiol Biotechnol 66:306–311
Rodrigues L, Banat IM, Teixeira J, Oliveira R (2006) Biosurfactants: potential applications in medicine. J Antimicrob Chemother 57:609–618
Rohde H, Burandt EC, Siemssen N, Frommelt L, Burdlski C, Wurster S, Scherpe S, Davies AP, Harris LG, Horstkotte MA, Knobloch JKM, Ragunath C, Kaplan JB, Mack D (2007) Polysaccharide intercellular adhesin or protein factors in biofilm accumulation of Staphylococcus epidermidis and Staphylococcus aureus isolated from prosthetic hip and knee joint infections. Biomaterials 28:1711–1720
Rossland E, Langsrud T, Granum PE, Sorhaug T (2005) Production of antimicrobial metabolites by strains of Lactobacillus or Lactococcus co-cultured with Bacillus cereus in milk. Int J Food Microbiol 98:193–200
Rufino RD, Luna JM, Sarubbo LA, Rodrigues LRM, Teixeira JAC, Campos-Takaki GM (2011) Antimicrobial and anti-adhesive potential of a biosurfactant rufisan produced by Candida lipolytica UCP0988. Colloids Surf B Biointerfaces 84:1–5
Rukayadi Y, Han S, Yong D, Hwang JK (2011) In vitro activity of xanthorrhizol against Candida glabrata, C. guilliermondii, and C. parapsilosis biofilms. Med Mycol 49:1–9
Ryjenkov DA, Tarutina M, Moskvin OV, Gomelsky M (2005) Cyclic diguanylate is a ubiquitous signaling molecule in bacteria: insights into biochemistry of the GGDEF protein domain. Bacteriology 187:1792–1798
Sachachter B (2003) Slimy business-the biotechnology of biofilms. Nat Biotechnol 21:361–365
Saino E, Sbarra MS, Arciola CR, Scavone M, Bloise N, Nikolov P, Ricchelli F, Visai L (2010) Photodynamic action of Tri-meso (N-methyl-pyridyl), meso (N-tetradecyl-pyridyl) porphine on Staphylococcus epidermidis biofilms grown on Ti6Al4V alloy. Int J Artif Organs 33:636–645
Sambanthamoorthy K, Gokhale AA, Lao W, Parashar V, Neiditch MB, Semmelhack MF, Lee I, Waters CM (2011) Identification of a novel benzimidazole that inhibits bacterial biofilm formation in a broad-spectrum manner. Antimicrob Agents Chemother 55:4369–4378
Sarabhai S, Sharma P, Capalash N (2013) Ellagic acid derivatives from Terminalia chebula Retz. downregulate the expression of quorum sensing genes to attenuate Pseudomonas aeruginosa PAO1 virulence. PLoS ONE 8:e53441
Schinabeck MK, Long LA, Hossain MA, Chandra J, Mukherjee PK, Mohamed S, Ghannoum MA (2004) Rabbit model of Candida albicans biofilm infection: liposomal amphotericin B antifungal lock therapy. Antimicrob Agents Chemother 48:1727–1732
Secinti KD, Ozalp H, Attar A, Sargon MF (2011) Nanoparticle silver ion coatings inhibit biofilm formation on titanium implants. J Clin Neurosci 18:391–395
Shen Y, Koller T, Kreikemeyer B, Nelson DC (2013) Rapid degradation of Streptococcus pyogenes biofilms by PlyC, a bacteriophage-encoded endolysin. J Antimicrob Chemother. doi:10.1093/jac/dkt104
Shirtliff ME, Krom BP, Meijering RA, Peters BM, Zhu J, Scheper MA, Harris ML, Jabra-Rizk MA (2009) Farnesol-induced apoptosis in Candida albicans. Antimicrob Agents Chemother 53:392–2401
Sillankorva S, Oliveira DR, Vieira MJ, Sutherland IW, Azeredo J (2004) Bacteriophage V S1 infection of Pseudomonas fluorescens planktonic cells versus biofilms. Biofouling 20:133–138
Sillankorva S, Neubauer P, Azeredo J (2010) Phage control of dual species biofilms of Pseudomonas fluorescens and Staphylococcus lentus. Biofouling 26:567–575
Simoes M, Sillankorva S, Pereira MO, Azeredo J, Vieira MJ (2007) The effect of hydrodynamic conditions on the phenotype of Pseudomonas fluorescens biofilms. Biofouling 24:249–258
Singh P, Cameotra S (2004) Potential applications of microbial surfactants in biomedical sciences. Trends Biotechnol 22:142–146
Singh N, Pemmaraju SC, Pruthi PA, Cameotra SS, Pruthi V (2013) Candida biofilm disrupting ability of di-rhamnolipid (RL-2) produced from Pseudomonas aeruginosa DSVP20. Appl Biochem Biotechnol 169:2374–2391
Sintim HO, Smith JA, Wang J, Nakayama S, Yan L (2010) Paradigm shift in discovering next-generation anti-infective agents: targeting quorum sensing, c-di-GMP signaling and biofilm formation in bacteria with small molecules. Future Med Chem 2:1005–1035
Smith AW (2005) Biofilms and antibiotic therapy: is there a role for combating bacterial resistance by the use of novel drug delivery systems. Adv Drug Deliv Rev 57:1539–1550
Smith KM, Bu YG, Suga H (2003a) Library screening for synthetic agonists and antagonists of a Pseudomonas aeruginosa autoinducer. Chem Biol 10:563–571
Smith KM, Bu YG, Suga H (2003b) Induction and inhibition of Pseudomonas aeruginosa quorum sensing by synthetic autoinducer analogs. Chem Biol 10:81–89
Soderling EM, Marttinen AM, Haukioja AL (2011) Probiotic lactobacilli interfere with Streptococcus mutans biofilm formation in vitro. Curr Microbiol 62:618–622
Splendiani A, Livingston AG, Nicolella C (2006) Control membrane-attached biofilms using surfactants. Biotechnol Bioeng 94:15–23
Stark M, Liu LP, Deber CM (2002) Cationic hydrophobic peptides with antimicrobial activity. Antimicrob Agents Chemother 46:3585–3590
Stewart PS (2003) New ways to stop biofilm infections. Lancet 361:97
Suzuki N, Yoneda M, Hatano Y, Iwamoto T, Masuo Y, Hirofuji T (2011) Enterococcus faecium WB2000 inhibits biofilm formation by oral cariogenic Streptococci. Int J Dent. doi:10.1155/2011/834151
Tahmourespour A, Kermanshahi RK (2011) The effect of a probiotic strain (Lactobacillus acidophilus) on the plaque formation of oral Streptococci. Bosn J Basic Med Sci 11:37–40
Takano E, Chakraburtty R, Nihira T, Yamada Y, Bibb MJ (2001) A complex role for the γ-butyrolactone SCB1 in regulating antibiotic production in Streptomyces coelicolor A3(2). Mol Microbiol 41:1015–1028
Tamayo R, Pratt JT, Camilli A (2007) Roles of cyclic diguanylate in the regulation of bacterial pathogenesis. Annu Rev Microbiol 61:131–148
Taraszkiewicz A, Fila G, Grinholc M, Nakonieczna J (2013) Innovative strategies to overcome biofilm resistance. Bio Med Res Int. doi:10.1155/2013/150653
Taweechaisupapong S, Singhara S, Lertsatitthanakorn P, Khunkitti W (2010) Antimicrobial effects of Boesenbergia pandurata and Piper sarmentosum leaf extracts on planktonic cells and biofilm of oral pathogens. Pak J Pharm Sci 23:224–231
Tetz GV, Artemenko NK, Tetz VV (2009) Effect of DNase and antibiotics on biofilm characteristics. Antimicrob Agents Chemother 53:1204–1209
Teughels W, Van Assche N, Sliepen I, Quirynen M (2006) Effect of material characteristics and/or surface topography on biofilm development. Clin Oral Implants Res 17:68–81
Tew GN, Liu D, Chen B, Doerksen RJ, Kaplan J, Carroll PJ, Klein ML, DeGrado WF (2002) De novo design of biomimetic antimicrobial polymers. Proc Natl Acad Sci USA 99:5110–5114
Thaweboon S, Thaweboon B (2009) In vitro antimicrobial activity of Ocimum americanum L. essential oil against oral microorganisms. Southeast Asian J Trop Med Public Health 40:1025–1033
Thenmozhi R, Nithyanand P, Rathna J, Pandian SK (2009) Antibiofilm activity of coral associated bacteria against different clinical M serotypes of Streptococcus pyogenes. FEMS Immunol Med Microbiol 57:284–294
Toulet D, Debarre C, Imbert C (2012) Could liposomal amphotericin B (L-AMB) lock solutions be useful to inhibit Candida spp. biofilms on silicone biomaterials? J Antimicrob Chemother 67:430–432
Tournu H, Dijck PV (2012) Candida biofilms and the host: models and new concepts for eradication. Int J Microbiol. doi:10.1155/2012/845352
Trampuz A, Widmer AF (2006) Infections associated with orthopedic implants. Curr Opin Infect Dis 19:349–356
Tsai PW, Yang CY, Chang HT, Lan CY (2011) Human antimicrobial peptide LL-37 inhibits adhesion of Candida albicans by interacting with yeast cell-wall carbohydrates. PLoS ONE 6:e17755. doi:10.1371/journal.pone.0017755
Valle J, Re DS, Henry N, Fontaine T, Balestrino D, Latour-Lambert P, Ghigo JM (2006) Broad-spectrum biofilm inhibition by a secreted bacterial polysaccharide. Proc Natl Acad Sci USA 103:12558–12563
van Hamme JD, Singh A, Ward OP (2006) Physiological aspects Part 1 in a series of papers devoted to surfactants in microbiology and biotechnology. Biotechnol Adv 24:604–620
Venkatesh M, Rong L, Raad I, Versalovic J (2009) Novel synergistic antibiofilm combinations for salvage of infected catheters. J Med Microbiol 58:936–944
Villa F, Cappitelli F (2013) Plant-derived bioactive compounds at sub-lethal concentrations: towards smart biocide-free antibiofilm strategies. Phytochem Rev 12:245–254
Wainwright M (2003) The use of dyes in modern biomedicine. Biotech Histochem 78:147–155
Walencka E, Sadowska B, Rozalska S, Hryniewicz W, Rozalska B (2005) Lysostaphin as a potential therapeutic agent for staphylococcal biofilm eradication. Pol J Microbiol 54:191–200
Walencka E, Rozalska S, Sadowska B, Rozalska B (2008) The influence of Lactobacillus acidophilus-derived surfactants on staphylococcal adhesion and biofilm formation. Folia Microbiol 53:61–66
Waters CM, Bassler BL (2005) Quorum sensing: cell-to-cell communication in bacteria. Annu Rev Cell Dev Biol 21:319–346
Weiss EI, Lev-Dor R, Kashamn Y, Goldhar J, Sharon N, Ofek I (1998) Inhibiting interspecies coaggregation of plaque bacteria with a craneberry juice constituent. J Am Dent Assoc 129:1719–1723
Wertheim HF, Melles DC, Vos MC, van Leeuwen W, van Belkum A, Verbrugh HA, Nouwen JL (2005) The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect Dis 5:751–762
Wilson M (2001) Bacterial biofilms and human disease. Sci Prog 84:235–254
Worthington RJ, Justin J, Richards MC (2012) Small molecule control of bacterial biofilms. Org Biomol Chem 10:7457–7474
Wuertz SOS, Hausner M (2004) Microbial communities and their interactions in biofilm systems: an overview. Water Sci Technol 49:327–336
Xavier JB, Picioreanu C, Rani SA, van Loosdrecht MCM, Stewart PS (2005) Biofilm-control strategies based on enzymic disruption of the extracellular polymeric substance matrix: a modelling study. Microbiology 151:3817–3832
Xiong Y, Liu Y (2010) Biological control of microbial attachment: a promising alternative for mitigating membrane biofouling. Appl Microbiol Biotechnol 86:825–837
Yan H, Chen W (2010) 3′,5′-cyclic diguanylic acid: a small nucleotide that makes big impacts. Chem Soc Rev 39:2914–2924
Yang L, Rybtke MT, Jakobsen TH, Hentzer M, Bjarnsholt T, Givskov M, Tolker-Nielsen T (2009) Computer-aided identification of recognized drugs as Pseudomonas aeruginosa quorum-sensing inhibitors. Antimicrob Agents Chemother 53:2432–2443
Zumbuehl A, Ferreira L, Kuhn D, Astashkina A, Long L, Yeo Y, Iaconis T, Ghannoum M, Fink GR, Langer R, Kohane DS (2007) Antifungal hydrogels. Proc Natl Acad Sci USA 104:12994–12998
Acknowledgments
We would like to thank University Grants Commission, and Indian Council of Medical Research, New Delhi for their financial support.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Khan, M.S.A., Ahmad, I., Sajid, M., Cameotra, S.S. (2014). Current and Emergent Control Strategies for Medical Biofilms. In: Rumbaugh, K., Ahmad, I. (eds) Antibiofilm Agents. Springer Series on Biofilms, vol 8. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-53833-9_7
Download citation
DOI: https://doi.org/10.1007/978-3-642-53833-9_7
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-53832-2
Online ISBN: 978-3-642-53833-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)