
Abstract
The lung is a vulnerable organ. Along with the skin and the gut, the lung is at the interface of the sterile body sanctuary and the environment and is thus exposed to numerous agents capable of inflicting injury (Fraire 2008; Berman and Center 1998). These agents include viruses, bacteria and physicochemical factors, against which the lung may respond by eliciting an inflammatory reaction and/or mounting an immune response. This chapter briefly considers the remarkably effective and biologically complex defense mechanisms of the lung, particularly those that apply to viral-associated infections.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
The lung is a vulnerable organ. Along with the skin and the gut, the lung is at the interface of the sterile body sanctuary and the environment and is thus exposed to numerous agents capable of inflicting injury (Fraire 2008; Berman and Center 1998). These agents include viruses, bacteria and physicochemical factors, against which the lung may respond by eliciting an inflammatory reaction and/or mounting an immune response. This chapter briefly considers the remarkably effective and biologically complex defense mechanisms of the lung, particularly those that apply to viral-associated infections.
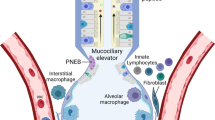
Classic defense mechanisms such as cough and mucociliary clearance offer only limited protection against viruses, primarily due to their exceedingly small size. Given these limitations, much of the needed defense to combat viral infections is derived from soluble mediators. Two of the most abundant soluble mediators found in airway secretions are lysozyme and lactoferrin. Most of the antimicrobial effects of these two mediators are focused on gram-negative bacteria in the case of lysozyme and both gram-negative and gram-positive bacteria in the case of lactoferrin (Kobzik 2008). In vitro defensins are potent microbicidal agents against many gram-positive and gram-negative bacteria, yeast, fungi, and some enveloped viruses (Kobzik 2008).
The immune response to infection encompasses innate immunity and adaptive immunity. Innate immunity is provided by physical, cellular, and chemical systems that are in place and respond to all classes of foreign invaders (Sherris 2010). These include mucosal barriers, phagocytic cells, and the action of circulating proteins such as complement molecules and mannose-binding protein (Sherris 2010). Further, many cells, including dendritic cells (DC) and macrophages, have recognition molecules that detect pathogen-associated molecular patterns (PAMPS), including nucleic acids, proteins, and glycolipids. Pathogen engagement of these recognition molecules, which include Toll-like receptors, cytoplasmic helicases, and other cellular sensors, can stimulate the synthesis of inflammatory cytokines such as interleukin 1 (IL-1), IL-6, and tumor necrosis factor (TNF) and antiviral cytokines including interferons. These and other cytokines will activate innate cells such as DC, macrophages, and natural killer (NK) cells and contribute to an early antiviral response that sets the stage for the adaptive immune response. Adaptive immunity, which is also called acquired immunity, refers to the ability to develop new responses that are highly specific to antigens derived from infectious agents. This is a consequence of the expansion of antigen-specific clones of T cells and antibody-producing B cells. In regard to viruses, it has been shown that previous immunity to one certain virus can influence a host response to an infection by a subsequent unrelated virus or viruses (Chen et al. 2003). Immunity to viral infections can also be influenced by commensal bacteria. Commensal bacteria are crucial in maintaining immune homeostasis in the gut, but the role of such bacteria in immune response of the lung and other mucosal surfaces is not well known (Ichinohe et al. 2011). Ichinohe examined the role of commensal bacteria in the initiation of adaptive immunity after respiratory infection with influenza virus and demonstrated that commensal bacteria contributed to immunocompetence of the lung. This was mediated in part by providing signals for robust priming of pro-IL-1B and pro-IL-18 expression, at steady state. These authors concluded that a key role exists for commensal bacteria in controlling adaptive immunity against respiratory virus infections, in this case influenza.
As noted by Land, viruses are macromolecular complexes made up of proteins plus DNA or RNA. Upon gaining access to the host cell source of energy and synthetic organelles, viruses effectively redirect host cell metabolism and synthetic capabilities in order to replicate and transmit their progeny (Land 2008). At this point, the immune system is activated and recognizes the virus as a foreign antigen. This protective mechanism is accomplished via the innate mechanisms cited above or by adaptive immunity. Soon after gaining entry into a host, viruses disseminate further by spreading via lymphocytes or the blood stream, leading to damage manifested as inflammation. Damage is the result of cell lysis and influx of cytokines (Land 2008).
1 Cellular Elements
In the complex process of cellular defense in the lung, lymphocytes and pulmonary macrophages stand out, as they play key roles in the inflammatory response, with some direction by dendritic cells . T and B lymphocytes display antigen specificity and play a significant role in defense against viruses. NK cells can also display some antiviral activity by virtue of their ability to sometimes distinguish between self and virus-infected self (Berman and Center 1998). Lymphocytes derive from progenitor stem cells in the marrow and thymus, leaving their initial milieu in a static state, designated to become B cells or T cells. The B cells mature in the bone marrow and then circulate in the blood to lymphoid organs (Sherris 2010). T cells mature in the thymus and then, like the B cells, circulate into the lymphoid organs, where they can interact with the B cells. NK cells are also derived from the bone marrow and circulate throughout the body. In the lymph nodes, B cells transform into plasmacytoid lymphocytes and eventually into plasma cells, which produce antibodies. T cells, on the other hand, mature in the thymus and then circulate, awaiting activation by antigen presented most efficiently by DC. This activation results in production of cytokines, which are effective molecules for multiple immunocytes and somatic cells (Sherris 2010).
2 Cytokines
Cytokines, broadly defined as active molecules released from a certain cell type and designed to have an effect on another or different cell population, play a critical role in antiviral defense mechanisms (Sherris 2010). Cytokines include chemokines, which are chemotactic factors for inflammatory cell migration, and interleukins, which are regulators of growth and differentiation of lymphocytes, monocytes, and DC. Cytokines also include TNF, an inducer of apoptosis, and interferons, which block viral replication and are critical to activation of NK cells, T cells, DC, and macrophages. Among these cytokines, interferons are of particular interest due to their antiviral properties. Interferons are regulatory cytokines produced by many nucleated cells, especially DC, in response to viral infection and are crucial in the early stage of viral infection. Interferon-stimulated cells shut down viral protein synthesis and effectively destroy viral mRNA. Interferons can also stimulate the activity of T cells and NK cells, thus accelerating the immune response to viral infection. The complex antiviral effects of interferon are graphically illustrated in Fig. 2.1.

Antiviral action of interferon: Interferon (IFN) synthesis and release are often induced by a virus infection when cellular sensors recognize their pathogen-associated molecular patterns such as double-stranded RNA or cytoplasmic DNA. This releases transcription factors that stimulate the synthesis of mRNA, and the translated IFN protein is secreted onto a neighboring cell. IFN binds to receptor on the plasma membrane of a second cell, and that binding stimulates the activation of genes encoding antiviral proteins. Two important antiviral proteins are the enzymes 2–5 oligo (A) synthetase and protein kinase R (PKR). When an IFN-stimulated cell is infected, viral protein synthesis is inhibited by a 2–5 A-activated endoribonuclease that degrades viral mRNA and by the PKR, which phosphorylates and inactivates the initiation factor eIF-2 required for viral protein synthesis (Reproduced with permission from Wiley J, Sherwood L, Woolverton C, Eds. Prescott’s Principles of Microbiology, New York: McGraw-Hill; 2008)
References
Immune Response to Infection Ch 2. (2010) In: Sherris Medical Microbiology 5th Ed. KJ Ryan and C George Ray, Eds. The McGraw Hill Co, New York, Chicago, San Francisco. pp 19–43
Berman JS, Center DM (1998) Lymphocyte and macrophage mediated inflammation in the lung. In: Fishman AP (ed) Pulmonary diseases and disorders, vol 1, 3rd edn. McGraw-Hill, New York, pp 275–287
Chen HD, Fraire AE, Joris I et al (2003) Specific history of heterologous virus infections determines anti-viral immunity and immunopathology in the lung. Amer J Pathol 163:1341–1355
Fraire AE (2008) Inflammation, Chap 43. In: Zander DS, Popper HH, Jagirdar J, Haque AK, Cagle PT, Barrios R (eds) Molecular pathology of lung diseases. Springer, New York, pp 459–469
Ichinohe T, Pang IK, Kumamoto Y et al (2011) Microbiota regulates immune defense against respiratory tract influenza virus infection. PNAS 108:5354–5359
Kobzik L (2008) Lung defenses. In: Tomashefski JF, Cagle PT, Farver CF, Fraire AE (eds) Dail and Hammar’s pulmonary pathology, vol 1, Non neoplastic lung diseases. Springer, New York/Heidelberg, pp 49–63
Land GA (2008) Molecular pathology of viral respiratory diseases, Chap 37. In: Zander DS, Popper HH, Jagirdar J, Haque AK, Cagle PT, Barrios R (eds) Molecular pathology of lung diseases. Springer, New York, pp 382–396
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Fraire, A.E., Welsh, R.M. (2014). Lung Defenses. In: Fraire, A., Woda, B., Welsh, R., Kradin, R. (eds) Viruses and the Lung. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-40605-8_2
Download citation
DOI: https://doi.org/10.1007/978-3-642-40605-8_2
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-40604-1
Online ISBN: 978-3-642-40605-8
eBook Packages: MedicineMedicine (R0)